Для большинства палеонтологических объектов биологическая природа видна уже при первоначальном рассмотрении их строения. При изучении строматолитов – минерализованных сообществ нитчатых цианобактерий – их генезис, соподчинение морфологических признаков, механизм образования и другие важные параметры не столь очевидны. Поэтому дополнительным инструментом в выяснении природы строматолитов может выступить характер их изменения в геологическом времени как отражение их эволюции.
Поскольку строматолиты обладают крайне невнятной и неустойчивой морфологией по сравнению с остатками заведомых организмов, долгое время их исследования ограничивались упоминаниями в качестве объектов неясной природы, занимающих промежуточное положение между текстурами пород и палеонтологическими объектами. Практическая необходимость расчленения и корреляции докембрийских карбонатных толщ, в которых по эволюционным причинам нет скелетных остатков, вызвала особый интерес к строматолитам как единственным макроскопическим ископаемым, содержащимся в них. Основные работы в этой области проводились в течение 50–80-х гг. ХХ в. – соответственно, на этот период приходится основной массив публикаций. Когда задачи геологического характера в целом были выполнены, их интенсивность резко снизилась. С одной стороны, за этот период был решен ряд сугубо биологических задач – разработаны принципы морфологического описания, классификации, доказано наличие у строматолитов биологической эволюции. Но, с другой стороны, эти исследования вывели на первый план вопрос о носителе организменных свойств строматолитообразователя.
Строматолиты образуются в результате жизнедеятельности сообщества бентосных нитчатых цианобактерий. Они являются микроскопическими фотосинтетиками и по этой причине не могут существовать как индивидуальные организмы, поскольку будут погребены осадком даже при минимальном его взмучивании. Благодаря действию этого и других факторов нитчатые бентосные цианобактерии существуют в виде макроскопических сообществ – кожистых пленок. Физическая целостность пленок активно поддерживается составляющими их микроорганизмами путем направленного движения и выделения слизистого матрикса. Благодаря этому бентосное сообщество цианобактерий не может рассматриваться как простая сумма индивидуальных микроорганизмов.
Природа сообщества нитчатых цианобактерий как целостного индивидуума подтверждена экспериментальными данными (Греченко и др., 2013; Сумина, Сумин, 2013; Сумина и др., 2020). Обнаружение явления морфогенеза в сообществе показывает, что оно способно контролировать свою форму и направленно изменять ее в зависимости от условий среды. Таким образом, форма строматолита, точнее, форма каждого его слоя, является достаточно полным отражением функциональной формы сообщества. А изменение формы слоев, как в процессе нарастания отдельного строматолита, так и при смене их комплексов в геологическом времени, является отражением изменения сообществом своей формы в ответ на изменения факторов среды.
Успешное использование строматолитов в биостратиграфии предполагает наличие у них целостности организменного уровня, выражающейся в иерархичности строения, дифференцированности на отдельные морфологически выделенные участки, связи этих участков между собой, в наличии приспособительного смысла черт строения, направленной и необратимой их смене во времени.
Основным биологически значимым результатом стратиграфического изучения строматолитов является установление того факта, что эволюция строматолитов сходна с таковой представителей фауны и флоры – объектов несомненной организменной природы. В общих чертах к такому выводу приходят М.А. Семихатов (1962), В.А. Комар (1964), И.Н. Крылов (1963). О направленном изменении ряда признаков строматолитов писали И.К. Королюк (1965), М.Е. Раабен с соавторами (Раабен, 1960; Комар и др., 1965; Раабен, Забродин, 1972).
Из приведенных данных следует, что строматолит, по-видимому, и должен рассматриваться как опорное минеральное образование некоего макроскопического организма. Эти организмы обладают способностью к морфогенетическим реакциям, а также филогенетическим изменениям. Приспособительный характер таких изменений был теоретически обоснован П.Ю. Петровым (1996). По его мнению, форма строматолита определяется не непосредственным воздействием внешних условий, а зависит от “внутренних функциональных причин развития и роста микробного сообщества” (Петров, 1996, с. 261). Однако в качестве носителя внутренних функциональных причин морфогенеза этот автор упоминает микробное сообщество, которое он понимает как биоценоз микроорганизмов.
Поскольку строматолитообразователь обладает функциями целого, эволюционные изменения строматолитов могут быть рассмотрены с точки зрения их приспособительного значения.
К ВОПРОСУ О ПРИЧИНАХ ЦЕЛОСТНОСТИ МИКРОБНЫХ СООБЩЕСТВ И НЕКОТОРЫХ ДРУГИХ ГРУПП ОРГАНИЗМОВ
Предваряя обсуждение темы о возможном уровне целостности древнейших микробных сообществ, одной из сторон которой является адаптивный характер морфологических изменений, необходимо остановиться на некоторых этапах развития представлений об “организмичности” на разных уровнях живого, и в первую очередь прокариот. Названия статей последних десятилетий в этой области знания говорят сами за себя – “Quorum sensing” или социальная жизнь бактерий” (Гинцбург и др., 2003), “Этология бактерий” (Смирнов, 2006), “Образование биопленок – пример “социального” поведения бактерий” (Романова и др., 2006), “Биопленка – “город микробов” или аналог многоклеточного организма?” (Николаев, Плакунов, 2007), “Биосоциальность одноклеточных (на материале исследований прокариот)” (Олескин, 2009), “Микробная популяция как многоклеточный организм” (Эль-Регистан, 2005).
Со времен Коха микробиология существовала в рамках парадигмы “чистых культур”, и, несмотря на колоссальные достижения, исследования экологии микробов оставались в стороне, хотя факт существования бактерий в виде сложных сообществ как в популяционном, так и в биоценотическом смысле был известен и во времена Левенгука. Сдвиг наметился, когда в середине прошлого века обратили внимание на взаимодействие микроорганизмов с поверхностями обитания в местах раздела фаз. Было показано, что большинство бактерий существует в виде биопленок на разнообразных биотических и абиотических поверхностях. Позже Г.А. Заварзин определил эту смену исследовательских подходов к биологическим объектам как переход от редукционизма к холизму – философии целостности, хотя сама идея существования микроорганизмов в природе не в виде свободно плавающих индивидов, а в виде структурированных сообществ была высказана Костертоном в 1978 г. (Costerton et al., 1978; Николаев, Плакунов, 2007).
В 2001 г. для особого типа микробных популяций был предложен специальный термин “структурированная бактериальная популяция”. Им обозначили бактериальные колонии, для которых характерна не только типичная для популяций вообще динамичная в пространстве и времени структура, но и особый высоко стабильный ее тип, который существует и регулируется более или менее как единое целое и совмещает свойства собственно популяции и организмов двух уровней (Головлев, 2001).
В биопленках бактерии объединены сложными межклеточными связями, которые осуществляют экспрессию разных генов в различных участках биопленки, что позволило рассматривать их как функциональный аналог многоклеточного организма. Биопленки имеют характерную архитектуру и окружены экзополимерным матриксом, одна из важнейших функций которого – защита бактерий от различных стрессовых факторов, а также создание структурной целостности биопленки. В матриксе расположены воздухоносные или заполненные жидкостью каналы, что рассматривается в качестве аналогов циркуляторной и дыхательной систем. Сложная архитектура пленок дает возможность метаболической кооперации внутри пространственно хорошо организованной системы, создавая условия для разнообразных взаимодействий между бактериями (Романова и др., 2006). Регуляция сложного поведения микроорганизмов в биопленках осуществляется не только сигналами из окружающей среды, но и межклеточными сигналами при участии механизма, получившего название “чувство кворума” (“quorum sensing”; Гинцбург и др., 2003).
Применительно к биопленкам используют термины “коммуникация”, “аффилиация” и “кооперация”. Эти сущностные характеристики позволяют проводить непротиворечивые аналогии с другими системами, состоящими из клеток (Олескин, 2009). В сообществах прокариот, как правило, отсутствует система “доминирования–подчинения”, однако отсутствие единого управляющего центра не препятствует эффективной координации социального поведения в сообществе, причем у микроорганизмов отмечены сетевые структуры нескольких уровней. Среди основных факторов такого поведения в бактериальных сообществах можно отметить межклеточные контакты, химические сигнальные агенты и наличие межклеточного матрикса (Олескин, 2009). Важно, что в микробных сообществах происходит синтез веществ с нейромедиаторной функцией животных и человека. В целом все это позволяет говорить о сходстве с нервной системой – удлиненные клетки в бактериальных колониях сравниваются с аксонами нервных клеток, матрикс – с глиальными клетками, а общая внешняя оболочка колонии – с оболочкой нервных структур. Как указывала Г.И. Эль-Регистан (2005, с. 14), “бактерии образуют сложные сообщества, коллективно охотятся за пищей и совершают совместные направленные перемещения и действия, координируя их посредством секретируемых внеклеточных сигнальных веществ – ауторегуляторов”.
Многочисленные примеры микробного взаимодействия приведены в ряде работ (Gray, 1997; Watnick, Kolter, 2000; Rickard et al., 2003; Jefferson, 2004; Beer, Stoodley, 2006).
Возможно, препятствием к пониманию бактериальных сообществ как структур организменного уровня целостности являлась внешняя простота их организации в сравнении с унитарными организмами, к которым могут быть отнесены прокариотные клетки, эукариотные клетки и многоклеточные организмы. Однако мы наблюдаем эти организмы на завершающих стадиях филогенеза, когда их черты приобрели устойчивый и определенный вид. Начальные же стадии становления организмов, будь то многоклеточность как эукариот, так и прокариот или образование модульных организмов, по понятным причинам значительно более просты и неопределенны. Это может быть отнесено и к их элементам.
Проблема целостности в разных ее аспектах, оценка стратегий модульных и унитарных организмов активно разрабатывается Н.Н. Марфениным (2016). Он на примере гидроидных полипов рассматривает переход от одиночных унитарных организмов к модульным и изменение целостности в процессе этого перехода. Разнообразие модульных организмов очень широко, если в него включать метамерные организмы с различной степенью олигомеризации. Трехслойные организмы имеют очень широкие возможности для реализации преимуществ централизации строения и поэтому большинство из них существенно удалены от первоначальных метамерных состояний, сохраняя только их реликты. Гидроиды же, как наиболее просто устроенные двуслойные, напротив, практически не имеют морфологических ресурсов для выхода из них и поэтому являются наиболее удобным объектом для изучения первичных стадий перехода от одиночных особей к модульным.
В начале этого перехода организмы двух рассматриваемых уровней скомпонованы различным образом. Первичный унитарный организм, как и его последующие копии, подвижен, хотя и без возможности передвижения по субстрату, дифференцирован и централизован. При почковании телесно непрерывное количество особей увеличивается. Большинство из них по строению не различаются между собой, но, благодаря наличию общей пищеварительной системы, уже появляются возможности для дифференциации с образованием непитающихся особей. Важным отличием отдельно взятого полипа от колонии является также ее неподвижность. В целом этот переход характеризуется тем, что “и анатомически, и функционально модульный организм оказывается более простым по сравнению с унитарным” (Марфенин, 2016, с. 317), т.е. эволюция организмов нового уровня начинается с первых недифференцированных стадий и впоследствии идет в том же направлении, что и эволюция образующих их унитарных организмов. Процесс дифференциации модульных организмов также идет в том же направлении, что и становление многоклеточности – объединение множества одинаковых самостоятельно питающихся организмов, образование распределительных и коммуникационных структур и дальнейшее выделение на этой базе разнообразия непитающихся специализированных особей, которые интенсифицируют функции организма следующего эволюционного уровня. “Особые органы управления (нервная и гуморальная системы) лишь надстроены над более простым типом взаимного согласования процессов, который является единственным у децентрализованных организмов, где интеграция основана на множественном взаимодействии относительно независимых паритетных частей целого” (Марфенин, 2016, с. 315). Здесь, начиная с самых первых этапов, мы видим, как сложность элементов постепенно переносится ими на организацию системы в направлении придания ей целостности унитарного организма.
Вопросы социальности трехслойных многоклеточных по сравнению со всеми вышеперечисленными объектами имеют гораздо более длительную историю осмысления, что связано с их размерностью и систематической близостью к исследователю. Фундаментальная общность социальных объединений животных (первично- и вторичноротых, как их назовут позднее) является одной из заметных составляющих культуры с древнейших времен.
Если человек максимально социален среди вторичноротых животных, то среди первичноротых лидируют муравьи. Наиболее полно к настоящему моменту эти вопросы проработаны А.А. Захаровым (2021), Э. Уилсоном и Б. Хёлльдоблером (Уилсон, 2020; Уилсон, Хёлльдоблер, 2022). В целом с эволюционной точки зрения рассматривается возможный переход от эгоистического конкурирующего поведения особи к групповому альтруистическому. Здесь нужно отметить, что сами по себе животные, как индивидуальные, так и общественные, являются высокоорганизованными сообществами клеток, сложность которых качественно превышает сложность сообществ, образуемых самими организмами. Н.Н. Марфенин (2016) указывает на это обстоятельство для отношений модульных организмов (“колоний”) и входящих в их состав элементарных унитарных организмов, но оно имеет универсальный характер.
Само образование сообществ вполне может быть отнесено к органопроекции (Флоренский, 1993). Иными словами, образование сообществ является фрактальным развертыванием сложности организмов за их физическими пределами (Сумин, Сумина, 2017). Этот процесс включает и клеточные организмы двух последовательных эволюционных уровней – прокариотные и эукариотные. Сам термин “эволюция” (развертывание) предполагает именно такое содержание.
В самом общем смысле как живые нами опознаются объекты, максимально независимые от среды. Это выражается в их строении, пространственном положении, химическом составе и других проявлениях. “Существование животного происходит не во внешней среде… но в жидкой внутренней среде”, “постоянство внутренней среды есть условие свободной, независимой жизни”, и “постоянство среды предполагает такое совершенство организма, чтобы внешние перемены в каждое мгновение компенсировались и уравновешивались” (Бернар, 1878, с. 96–97).
Социальность является еще одной границей, позволяющей организмам поддерживать свою независимость от ниже организованной внешней среды и иметь более широкие возможности для репарации нарушений, неизбежно возникающих при передаче наследственной информации. Иными словами – противостоять случайной изменчивости и естественному отбору и вести свою эволюцию в выбранном направлении (Дарвин, 1941; Rassoulzadegan et al., 2006; Dias, Ressler, 2014; Cossetti et al., 2014). Закономерно, что при такой высокой сложности организменных систем и при таком высоком градиенте организованности между ними и средой большинство эволюционных попыток оказываются неудачными. Удачные же решения никогда не имеют постоянного характера.
Жизнь организмов вообще и их социальность в частности связаны с развитием интеллектуальных способностей как в структурном, так и в функциональном смысле. По этой причине у животных независимо от их систематического положения просматривается прямая связь между сложностью поведения и “недоразвитостью” потомства. У животных с наиболее сложным поведением оно физиологически и морфологически максимально ограждено от упрощающего влияния на развитие когнитивных способностей внешней среды, в том числе социальной, что естественным образом продолжает изолированное развитие эмбриона в яйце или во время беременности. Это необходимо, так как даже у одиночных организмов молодь более социальна, чем взрослые особи, поскольку ее развитие на ранних стадиях онтогенеза фрактально определяется внутренними причинами, в первую очередь отношениями клеток многоклеточного организма. Однако для содержания беспомощной молоди требуются именно социальные условия. Поэтому возникает положительная обратная связь между социальностью, способностью к сложному поведению особей и их изоляцией от внешней среды на ранних стадиях онтогенеза.
Таким образом, для многоклеточных организмов эволюционно первичным логично предполагать социальное, а не индивидуальное существование. При отсутствии других проявлений социальности размножение может рассматриваться в качестве ее реликта. Если с этой точки зрения рассмотреть структуру разнообразия, то она включает абсолютное большинство индивидуальных организмов и меньшинство социальных. Это объясняется тем, что при любых преобразованиях возникает большинство неустойчивых и тупиковых конструкций. Как современное, так и наблюдаемое ископаемое разнообразие в целом будет состоять из продуктов их распада с большим числом переходов между состояниями.
Целостность социальных систем поддерживается постоянными или временными механическими контактами между их элементами. Такие контакты имеют малую протяженность и охватывают малое количество элементов. Однако возможных вариантов этих контактов существует бесконечное множество. Целенаправленно поддерживаются именно те, которые динамически сохраняют целостность системы, т.е. в основе выбора типа и участников контакта лежит представление каждого элемента о системе как целом.
Терминов “эгоизм” и “альтруизм” для описания этих взаимодействий недостаточно. Эгоизм – это стремление особи к собственной пользе или выгоде, альтруизм же – бескорыстная забота о благополучии других индивидуумов. В случае эусоциальных взаимодействий каждый из участников проецирует на систему индивидуальную информацию о ней, и часто это несовместимо с продолжением жизни. Социальность, таким образом, является информационным суперэгоизмом, физически проявляемым с помощью локальных телесных отношений. Физический размер муравья или клетки качественно меньше муравейника или организма, но для динамического сохранения или направленного изменения структуры связей сообщества муравей и его клетка, как информационно, так и в смысле сложности, должны быть “больше” муравейника и организма.
Если социальные сообщества многоклеточных организмов еще находятся в стадии становления, то клеточная социальность, лежащая в основе строения самих организмов, обеспечивается вполне устойчивыми структурами. Эукариотным клеткам, чтобы построить многоклеточный организм, требуется универсальный геном, описывающий организм в целом. Для конкретизации его считывания необходимы общеорганизменные информационные системы, к которым относятся общая сеть цитоскелета, обмен экзосомами и др. Таким образом, реализация генетической информации является поведенческим актом, а не выполнением некой неконтролируемой клетками исторически возникшей программы, что наиболее явно проявляется в ходе эмбриональных регуляций. Также здесь важно учитывать существование химерных организмов, то есть то, что унификация генома является не единственным и не главным способом согласования деятельности клеток. Эти механизмы являются общими как для клеток многоклеточных, так и для самих многоклеточных в организмоподобных социальных сообществах.
Важным вопросом здесь является соотношение наследуемой инстинктивной информации и когнитивных процессов (Панов, 2012). Та же двойственность была отмечена В.Я. Бродским (2009) для микроорганизмов. Связь социальности и развития информационных структур менее однозначна, чем это представлялось ранее. Например, сложная нервная система и когнитивные способности головоногих с социальностью не связаны. Ядерный аппарат многих одноклеточных более сложен, чем у многоклеточных. В целом же можно наблюдать первичность когнитивных процессов и закрепление удачных решений в виде наследственности.
Необходимо также отметить, что у социальных первичноротых по объему развитие нервной системы несопоставимо ниже, чем у большинства социальных вторичноротых, но они качественно более социальны, чем общественные вторичноротые, за исключением человека. Но в обоих случаях социальность основывается на высоко развитой нервной системе, включающей сложно организованный головной мозг, несмотря на то, что они не гомологичны.
Рациональная архитектура гнезд муравьев (Захаров, 2021) неизбежно должна быть связана с рациональными способностями особей, а они, в свою очередь, – с развитием нервной системы и со сложностью поведения. Особую роль здесь играет ситуативная и закрепляющаяся дифференциация поведения, основанная на интеграционных процессах (Волкова, 2014), о чем также говорит сложное устройство нервной системы и ее наибольшее развитие у рабочих особей.
А.А. Захаров (2021) также указывает на наличие сети локальных контактов между муравьями, позволяющей им строить купол правильной геометрии. Сама сеть контактов дифференцирована и имеет на вершине большую плотность. Кроме того, область вершины купола является областью принятия структурных решений, определяющих конкретные изменения в функционировании колонии. Из этого следует, что физически локальные контакты являются проявлением общих в отношении муравейника, инстинктивно и рационально понимаемых связей. Иными словами, для того, чтобы муравейник приобретал определенную форму, каждый муравей должен строить именно муравейник в целом, а не осуществлять “локальные контакты”.
Как это ни удивительно, но просматривается большое сходство между способом образования купола муравейника и строматолитовой постройки. Несмотря на все различия, между этими биогенными образованиями существует прямая морфофункциональная связь – в обоих случаях сообщества, используя рыхлый материал, выполняют задачу подъема над субстратом, увеличения инсоляции, удаления осадков с поверхности обитания, увеличения ее площади и т.д. Эти общие принципы, конечно, реализуются по различным причинам, различными способами и в различных условиях. Однако согласованная морфогенетическая активность элементов не может осуществляться без ее позиционирования и, соответственно, координатной сети (Сумина и др., 2020). Также в обоих случаях эта сеть оказывается дифференцированной и имеющей центр на вершине. Деление купола начинается с деления вершины, что само по себе не имеет функционального смысла. Это говорит о том, что морфологическим преобразованиям предшествуют информационные. У строматолитов эти преобразования координатной сети иногда выражены морфологически (Власов, 1970).
Исследования столь различных групп организмов все в большей степени выявляют общность принципов организации их сообществ. По всей видимости, на этом основании можно говорить о сквозных принципах организации сообществ вообще – по крайней мере от бактерий до человека. Наибольшее внимание, в том числе и с практической точки зрения, привлекают взаимосвязанные вопросы происхождения сложности сообществ и их централизации как этапа повышения этой сложности.
Как было показано выше, источником сложности сообщества на любом эволюционном уровне организмов является сложность их элементов (Марфенин, 2016; Sumina, Sumin, 2022). Для образования сообщества элементарным организмам в своем поведении необходимо реализовывать в первую очередь всеобщие связи, определяющие структуру сообщества, исходя из его внешних отношений. Простейший пример – куполообразные постройки муравьев и цианобактерий, полезность формы которых определяется как минимум их взаимоотношением с осадками и освещением. Также в этом поведении должны учитываться конструкционные свойства применяемых материалов и их соотношение с формой будущей постройки. Здесь также наблюдается сходство между постройками общественных насекомых и строматолитами. В эволюции цианобактериальные сообщества постоянно увеличивают прочность построек как на уровне материала, так и на уровне его компоновки и переходят от куполообразных построек к ветвящимся (рис. 1, 2). Здесь уже просматривается сходство скорее с термитниками, при построении которых используется более прочный материал, чем в куполообразных муравейниках. У муравьев также отмечена эта тенденция – их земляные купола имеют более крутые склоны, чем строящиеся из хвои. Далее в поведении реализуются общие связи, определяющие внутреннюю структуру сообщества в соответствии с внешними условиями. И лишь в последнюю очередь они конкретизируются с помощью локальных взаимодействий.
Рис. 1.
Возможное направление морфологической эволюции архейских строматолитов, связанное с выделением столбчатых структур (по: Hofmann, 2000): а – возникновение выпуклых и вогнутых участков рельефа, б – увеличение крутизны выпуклых структур рельефа и расстояний между ними, в – формирование столбиков и промежутков между ними с ограниченным развитием цианобактериальной пленки, г – окончательное формирование закономерных столбчатых морфотипов, д – возникновение ветвления и дальнейшее становление его закономерности.
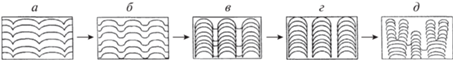
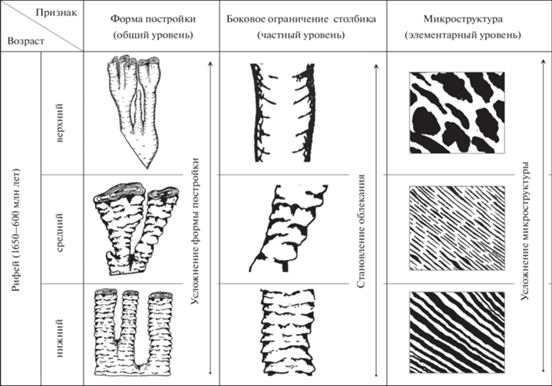
Рис. 2.
Закономерное изменение черт строения строматолитов на протяжении рифея (по данным В.А. Комара, 1964, 1966).
Согласованность деятельности большого количества самостоятельных индивидуумов не может быть достигнута без централизованного управления. Однако множество успешно живущих сообществ представляются нецентрализованными, и такая форма их организации обоснованно представляется эволюционно первичной. Исходя из высокого уровня организации взаимодействий, такие системы в действительности являются централизованными каждым индивидуумом – управляющим действиями всех остальных и управляемым ими. Такое базовое состояние сообществ, которое не утрачивается с нарастанием централизации, вероятнее всего, следует назвать взаимоорганизацией (Сумина, Сумин, 2013).
Возникновение централизации в сообществе является не сменой принципов организации, а надстройкой над взаимоорганизацией (Марфенин, 2016). В ее рамках возможно не только усиление управляющих функций некоторой части особей, но и сохранение восприятия получаемых от них сигналов как управляющих. Здесь важна возможность организации обратных связей в информационной системе сообщества. Если коммуникация клеток организма, как правило, основана на идентичности изначальной генетической информации, то коммуникация в сообществах перепончатокрылых, например, опирается на общность в 75% и качественно уступает по своей сложности организмам. Это показывает, что чем более равны элементы, тем более высокую и централизованную иерархию они могут построить – информационная протяженность устойчивых вертикальных связей всегда оказывается пропорциональной протяженности горизонтальных связей. Конфигурация же механических контактов может соотноситься с этой зависимостью любым образом.
В случае увеличения в сообществах необратимой дифференциации интенсивность его функционирования повышается. При этом начинает нарастать специализация во взаимоотношениях с внешней средой, что сужает ресурсную базу, повышает уязвимость и, главное, становится невозможным непрерывное эволюционное развитие. Для обеспечения возможности развития дифференцированных систем у многоклеточных эукариот возникли такие сложные механизмы, как половое размножение, старение и смерть. Это позволяет на время переводить клеточное сообщество в недифференцированное, нецентрализованное состояние и вводить в структуру организма новую информацию. Данный механизм наблюдается не только у организмов, но и у их сообществ – нецентрализованный муравейник переживает смену сезонов, меняет местоположение без сброса основной части биомассы, а централизованные семьи, например, осы обыкновенной, для этого должны ежегодно терять всех особей колоний, кроме покинувших ее молодых самок.
Сообщества нитчатых цианобактерий являются альтернативным многоклеточным эукариотам способом реализации двух уровней прокариотной многоклеточности (Гусев, Гохлернер, 1980). При качественно сопоставимых возможностях дифференциации они сохраняют универсальность элементов, и поэтому функциональная и морфологическая дифференциация сообществ является обратимой, а изменение их формы и строения, преодолевающее таксономические границы, происходит с одной и той же непрерывно существующей биомассой.
Отдельно может рассматриваться вопрос о соотношении собственной сложности цианобактерий и сложности образуемых ими сообществ. В отношении как эукариотных многоклеточных организмов (т.е. сообществ клеток), так и их социальных объединений наблюдается соответствие сложности информационных структур организмов, их индивидуального поведения и сложности социальных систем.
В отношении нитей цианобактерий и их клеток соответствующей сложности структурной организации не наблюдается. Здесь мы имеем дело с ситуацией, когда наблюдение в большей степени зависит от развития увеличительных приборов, чем от объектов, в частности это касается системы цитоскелета. И объективно окажется, что только поведение, являясь интегральной характеристикой, способно отражать реальную сложность организма.
Рассмотрим с точки зрения обсуждавшихся выше общебиологических закономерностей целостные адаптивные реакции строматолитообразующего сообщества.
Можно предположить, что основными тенденциями развития строматолитообразователя как бентосного макроскопического фотосинтетика в условиях интенсивного осадконакопления могут быть: уход из зоны взмучивания осадка, удаление осадка с поверхности, сохранение относительной поверхности фотосинтеза, поддержание оптимальной формы (табл. 1). При карбонатном осадконакоплении сообщество имеет возможность использовать биогенный карбонат, выпадающий в результате изъятия из среды растворенной в ней углекислоты. Этот карбонатный компонент может быть вовлечен в морфогенетические преобразования, которые способны компенсировать отрицательное влияние выпадения минерального осадка.
Таблица 1.
Соотношение потребностей строматолитообразователя и направлений эволюции основных черт строения ископаемых строматолитов
| Строматолитообразователь | Строматолит |
| Уход из зоны взмучивания осадка | Увеличение высоты постройки (высоты слоя) |
| Удаление осадка с поверхности | Увеличение крутизны арки |
| Сохранение и увеличение относительной поверхности фотосинтеза | Возникновение ветвления, в том числе активного |
| Поддержание оптимальной формы | Упрочнение постройки, определяемое расположением агрегатов карбонатных зерен в слое (усложнение микроструктуры) |
В случае образования строматолита как твердого опорного минерального тела жизненные потребности строматолитообразователя будут выражаться в следующем: общем увеличении высоты постройки (высоты слоя), увеличении крутизны арки, возникновении ветвления, упрочении постройки путем определенного расположения агрегатов карбонатных зерен в слое.
ОБЩИЕ ТЕНДЕНЦИИ ИЗМЕНЕНИЯ ОСНОВНЫХ МОРФОЛОГИЧЕСКИХ ЧЕРТ СТРОЕНИЯ СТРОМАТОЛИТОВ ВО ВРЕМЕНИ
Тренд направленного изменения морфологических черт строматолитов, который обеспечивает потребности строматолитообразователя, может быть прослежен, начиная с самых древних, палеоархейских, находок (3.5 млрд лет).
В архее (3.5–2.5 млрд лет назад) одним из наиболее распространенных морфотипов являются псевдостолбчатые постройки, которые встречаются и в более поздних отложениях.
Наряду с ними отмечены постройки, представляющие собой, по-видимому, полный переходный ряд от псевдостолбчатых к собственно столбчатым постройкам. Средние члены этого ряда названы Хофманном “partly linked” (“частично соединенные”) (Hofmann, 2000). Вероятно, такой набор форм в древнейших строматолитовых комплексах отражал процесс становления морфотипа столбчатых строматолитов на базе построек пластового типа.
В биогермных рядах (Крылов, 1975) рифейских строматолитов (1650–600 млн лет) образованию столбчатой части биогерма предшествует образование пластовой постройки. Ведущим механизмом эволюционных преобразований при переходе от пластового морфотипа к столбчатому было выделение в пределах единой минерализующейся пленки, образующей пластовый строматолит, таких элементов ее строения, которые повышали устойчивость к воздействию неблагоприятных факторов среды, в частности засыпанию осадком (Маслов, 1960; Власов, 1970). Такими элементами могут быть выпуклые структуры рельефа, которые позволяют разгружать осадок в промежутках между ними. Впоследствии чередование выпуклых и вогнутых структур приобретает унаследованный характер (рис. 1а). Одним из факторов морфологического прогресса будет, очевидно, увеличение крутизны выпуклых структур рельефа и расстояний между ними (рис. 1б). Развитие такой организации поверхности позволяет эффективно предотвращать засыпание осадком одних участков, но делает невозможным обитание пленки цианобактерий на других.
В дальнейшем эволюционные преобразования могут быть связаны с разделением первоначально единой пленки на отдельные участки, которые занимают только возвышенные части поверхности при ограничении роста пленки в понижениях (рис. 1в). Результатом этого процесса должно быть появление столбчатого морфотипа (рис. 1г). Эти преобразования являются следствием морфогенетического контроля отдельных участков пленки над своей формой и своими границами. Дальнейшим развитием такого контроля будет появление ветвящихся строматолитов (рис. 1д). Деление их арки ограничивает размер отдельных столбиков при их росте и позволяет пленке сохранять достаточно большую относительную поверхность (Сумина, 2002). Морфогенетический контроль формы построек в этом ряду проявляется раньше, чем морфогенетический контроль размеров.
Для архея (3.5–2.5 млрд лет назад) отмечаются также постройки с бугорчатой поверхностью, на срезе имеющие волнистую форму слоя с изгибами сантиметровой размерности. В более поздних отложениях, например, рифейских (1650–600 млн лет), они не встречаются, там развиты строматолиты с гладкой формой слоя. Это также указывает на увеличение степени морфогенетического контроля строматолитообразователей над своей формой и, соответственно, постройкой, хотя и для архея отмечено наличие форм с гладкими слоями.
Эволюционную направленность изменений строматолитов и соподчиненность изменения их морфологических признаков можно четко проследить на примере столбчатых строматолитов рифея, в частности ветвящихся строматолитов Анабарского массива (Комар, 1964). Здесь можно выявить следующие закономерности (рис. 2):
1. Усложнение формы построек, связанное с изменением характера ветвления и формы строматолитовых столбиков – от простых по форме пассивно ветвящихся построек группы Kussiella Kryl. до активно ветвящихся со сложными столбиками построек Baicalia Kryl. и Anabaria Kom.
2. Изменение характера боковой поверхности столбиков. Неровная, бахромчатая поверхность столбиков (Kussiella Kryl.) сменилась на столбики сначала с более ровной поверхностью (Anabaria Kom.), а затем на столбики с полным многократным облеканием (Boxonia Kor.).
3. Изменение микроструктур строматолитовых слоев от простых пластинчатых до сложных – сгустковых и шестоватых. Эти закономерные изменения авторы также интерпретируют как “отражение естественной эволюции водорослей-строматолитообразователей” (Комар, 1964, с. 100). Важным является то, что это усложнение построек прослеживается в пределах литологически однородной толщи доломитов, накопление которой происходило в течение примерно 800 млн лет (Комар, 1964, 1966). По этой причине усложнение нельзя связывать с фациальной изменчивостью. Это можно рассматривать как пример “чистой” эволюции, когда организмы в течение длительного времени находились в практически неизменяемых условиях обитания.
Приведенные данные показывают не только направленность эволюции, но и взаимосвязь изменений разных групп признаков. Одновременно и согласованно меняются общая морфология постройки, отдельных ее частей и микроструктура. Здесь возможно провести аналогию с тремя различными иерархическими уровнями организации индивидуумов многоклеточных эукариот – организменным, “органным” и тканевым (Кремянский, 1969). Организменному уровню может соответствовать общая форма постройки, “органному” – особенности строения отдельных ее частей, тканевому – микроструктурные особенности слоев. Согласованность в усложнении структур, принадлежащих различным уровням организации строматолитовой постройки, может быть показателем высокой целостности строматолитообразователя.
Рассмотрим в контексте приведенной аналогии изменение четырех наиболее хорошо прослеживаемых на ископаемом материале признаков строматолитов. Они могут быть сгруппированы следующим образом: к организменному (наиболее общему) уровню могут быть отнесены тип ветвления и форма столбиков, “органному” (частному) – характер бокового ограничения, к тканевому (элементарному) – микроструктура.
Тип ветвления. Сравнение типа ветвления столбчатых строматолитов Южного Урала выявило следующую закономерность: тип ветвления направленно усложняется от нижнего рифея (1650–1350 млн лет) к верхнему (1030–600 млн лет). Так, для строматолитов группы Kussiella из нижнего рифея характерно простое деление столбика на два более узких. Для среднерифейских (1350–1030 млн лет) построек группы Baicalia характерно ветвление на два столбика или несколько, с резким пережимом в месте ветвления. У верхнерифейских Minjaria u Gymnosolen наблюдается сложное кустистое или древовидное ветвление столбиков (Крылов, 1963). Изменение характера ветвления во времени подтвердилось при изучении рифейских строматолитов из других регионов (Семихатов, 1960; Комар, 1964). М.Е. Раабен (1964) на материале Полюдова кряжа также отметила ряд закономерностей, связанных с ветвлением строматолитовых столбиков. Пассивное ветвление, при котором столбики в процессе роста не увеличивают свою площадь, характерно для нижнего рифея, хотя может быть встречено и в более молодых отложениях. Активное ветвление столбиков встречается в среднем рифее и выше, а для верхнего рифея характерны столбики с активным ветвлением, т.е. расширяющиеся в процессе роста.
Форма столбиков. Форма столбика изменяется также направленно. Для построек нижнего рифея характерны столбики субцилиндрические, прямоосные и субвертикальные. Средне- и верхнерифейские строматолиты, наряду с субцилиндрическими, имеют и столбики неправильной формы (Комар, 1966).
Характер бокового ограничения. И.К. Королюк (1960) при изучении строматолитов Иркутского амфитеатра отметила, что строматолиты из нижне- и среднерифейских отложений характеризуются “бахромчатой” боковой поверхностью столбиков. В верхнем рифее появляются формы с гладкими столбиками. Из всего разнообразия выделенных ею типов боковой поверхности столбиков (без стенки, с однослойной стенкой, с многослойной стенкой, со стенкой, образованной “особой тканью”) формы с отчетливыми стенками появляются в самых верхах докембрия – в верхнем рифее и в кембрии (600–535 млн лет назад) (группа Boxonia Kor.). В рифее большинство строматолитов имеют стенку только на отдельных участках столбиков. Эти наблюдения полностью подтверждаются выводами, сделанными по строматолитам Южного Урала И.Н. Крыловым (1963). В целом аналогичные наблюдения приводятся и В.А. Комаром (1966) для строматолитов севера Сибирской платформы – в нижнем рифее присутствуют формы с неровной или бахромчатой поверхностью, без облекания. В среднем рифее появляются строматолиты с гладкой боковой поверхностью, с нечетко выраженным облеканием. Верхний рифей характеризуют и “определяют лицо строматолитового комплекса” (Комар, 1966, с. 53) формы с гладкой боковой поверхностью, часто с многослойным облеканием. Наряду с этим продолжают развиваться и формы без облекания.
Микроструктура. Этот признак, характеризующий внутреннее строение слоев, также имеет определенную приуроченность к четким стратиграфическим уровням. В.А. Комар (1964) при изучении столбчатых строматолитов рифея Анабарского массива установил следующую закономерность: микроструктура изменяется во времени от простой – пластинчатой и линзовато-пластинчатой в раннем рифее – до сложной – сгустковой и шестоватой, которая появляется в более поздних горизонтах рифея. Такая закономерная смена типов микроструктуры на определенных стратиграфических уровнях позволила сделать вывод о естественной эволюции организмов-строматолитообразователей. Эта точка зрения нашла подтверждение и при использовании биометрических методов (Комар и др., 1965). Постоянство типов микроструктур на определенных стратиграфических уровнях отмечалось для Енисейского кряжа и Туруханского района (Семихатов, 1962), Учуро-Майского района (Нужнов, 1960), Полюдова кряжа (Раабен, 1964), Южного Урала (Крылов, 1963, 1975).
В.А. Комар (1966) для каждого подразделения рифея в качестве примера приводит определенный тип микроструктуры из разрезов различных регионов: зонально-пластинчатая характерна для нижнего рифея, линзовидно-штриховатая – для среднего рифея, сгустково-слоистая – для верхнего рифея. Выдержанность микроструктур отмечается не только для столбчатых построек, но и для других морфотипов, например, пластовых.
ПРИСПОСОБИТЕЛЬНОЕ ЗНАЧЕНИЕ МОРФОЛОГИЧЕСКИХ ПРИЗНАКОВ СТРОМАТОЛИТОВ
Рассмотрим функциональный смысл описанных выше изменений.
Эволюция строматолитов как направленное изменение целостных образований может считаться наблюдаемым, эмпирически установленным фактом. Это изменение должно иметь постоянно действующие внутренние причины, поскольку существенных колебаний условий среды за это время (по крайней мере в течение рифея) не происходило – принципиально не менялись темпы осадконакопления и состав пород (Раабен, 1981). Увеличение прижизненной высоты строматолитов над осадком могло быть достигнуто только благодаря повышению скорости отложения минерального материала строматолитообразователями. Как указывают М.А. Семихатов и М.Е. Раабен (1996), ряд перестроек палеогеографической обстановки на различных этапах формирования строматолитов создавал предпосылки для увеличения их разнообразия, чего, однако, не происходило. Единственным фактором, который существенно изменял свои параметры, был состав гидросферы, однако он не мог оказать прямого влияния на изменение форм построек и если и влиял, то опосредованно, через биохимические свойства строматолитообразователей.
Общей тенденцией эволюционного преобразования построек на протяжении рифея явилось увеличение крутизны арки и высоты слоя, что позволяло строматолитообразователям, как бентосным сообществам, выходить из зоны придонного волнения и взмучивания. Следствием увеличения арки является облекание боковой поверхности столбика и образование стенки. Облекание само по себе является важным морфогенетическим приобретением, поскольку резко упрочняет постройку и дает возможность образования сложных ветвящихся форм. Однако в отношении увеличения высоты арки оно явно вторично и является его следствием.
Увеличение крутизны арки является показателем увеличения удельной фотосинтетической продукции, поскольку выпадение биогенного карбоната кальция является побочным продуктом фотосинтеза. Наиболее интересным в наблюдаемом эволюционном ряду является то, что при приобретении строматолитообразователями избыточного материала, а именно карбоната, который может быть использован в дальнейших морфогенетических построениях, в морфогенез он вовлекается спустя некоторое время (рис. 3).
Рис. 3.
Морфогенетические преобразования рифейских столбчатых строматолитов (временная шкала дана вне масштаба) (по: Сумина и др., 2020). R – рифей, V – венд.
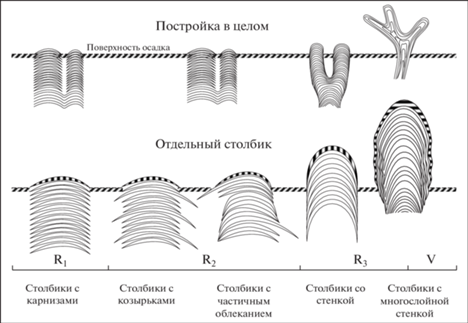
На ископаемом материале можно наблюдать становление данной морфогенетической реакции в ее развитии (рис. 1). Вероятно, наиболее простым типом столбчатого строматолита является постройка с практически плоской или слабо выпуклой аркой и без облекания. Несмотря на то, что в породе на продольном срезе такая постройка может иметь достаточно сложную морфологию и достигать нескольких метров в высоту, прижизненно такой строматолит возвышался над осадком не более чем на высоту одного слоя, т.е. около 5 мм. Примером такого строматолита могут служить постройки Kussiella kussiensis, характерные для отложений нижнего рифея. Можно предполагать, что в этом случае продукции карбоната хватало лишь на то, чтобы обеспечить только небольшое возвышение над поверхностью осадка, и скорость роста строматолита не превышала скорости фонового осадконакопления.
В верхних горизонтах нижнего рифея появляются строматолиты с несколько отличной морфологией, такие как Baicalia, а в среднем и верхнем рифее – Tungussia. С одной стороны, они проявляют принципиальное сходство с Kussiella, поскольку имеют достаточно низкие слои. Но отличаются от последних тем, что отдельные слои выходят за пределы боковой поверхности столбиков (особенно у Tungussia). Это может указывать на повышение удельной продукции фотосинтеза и скорости роста самих строматолитообразователей – в отдельные благоприятные периоды, например, связанные с локальным уменьшением скорости осадконакопления, они захватывали близлежащие поверхности осадка. Такое изменение формы боковой поверхности столбика не было связано с изменением формы слоев, т.е. фактически происходило без морфологического усложнения всей постройки. Строматолитообразователю удавалось лишь на некоторое время увеличить свою поверхность. Это приводило к образованию столбика, из которого в разные стороны выступали редкие карнизы.
У более поздних форм появляется такая особенность строения столбика, как облекание, т.е. охватывание вышележащими слоями нижележащих. В этом случае происходит принципиальное изменение формы слоя – избыточный карбонат расходуется не на увеличение горизонтальной поверхности (карнизов), а на организацию вертикальной, т.е. боковой, поверхности. Это приводит к подъему постройки над осадком, позволяет строматолиту увеличивать высоту со скоростями, более высокими, чем темпы осадконакопления, и обеспечивает постройку с достаточной внутренней прочностью. Все перечисленные морфологические преобразования на базе облекания позволяют образовывать высокие (высота выступающей над осадком части постройки), сложно разветвленные постройки. Такие постройки развиты в верхнем рифее, например, постройки р. Gymnosolen.
Процесс преобразования формы слоя показывает, как в рамках предыдущих, более простых морфотипов появляется избыточный ресурс. Он инициирует следующий этап морфологического усложнения построек, который повышает в конечном итоге степень независимости строматолитообразователя от неблагоприятных факторов среды.
Описанная выше тенденция отражает общие черты развития и смены морфотипов строматолитов. Однако имеется целый ряд исключений, которые, на первый взгляд, противоречат общей наблюдаемой картине. Так, из ятулийских отложений Карелии (нижний рифей, 1650–1350 млн лет) известны находки четких гимносоленоподобных построек. Для строматолитов группы Sundia из тех же отложений характерно отчетливое (“верхнерифейское”) боковое ограничение (Крылов, 1975). Однако “отдельные находки стеночных или кустистых строматолитов в глубоком докембрии не изменяют общего представления о смене строматолитовых комплексов в разных горизонтах рифея, а опровергают только упрощенные схематические представления об этих комплексах” (Крылов, 1975, с. 154). Этот же автор подчеркивает, что попытки устанавливать возраст по “первому появлению” построек какой-либо группы привело бы к резкому и неоправданному смещению возрастных границ комплексов.
Появление таких “противоречивых”, не укладывающихся в общий эволюционный тренд находок, вероятнее всего, связано с тем, что на общий ход строматолитообразования оказывал влияние не некий постоянно изменяющийся внешний фактор или группа факторов. Это могло быть результатом однонаправленного, но идущего у разных групп не одновременно процесса морфологического усложнения, вызванного внутренними причинами (Бернар, 1878). Возможно, именно это обстоятельство приводило к некоторым несоответствиям. Реконструируемые изменения палеогеографических обстановок предполагали определенный ожидаемый ответ в виде падения или увеличения разнообразия строматолитов, а реально наблюдаемая картина иногда имела прямо противоположный облик. Так, при трансгрессии в конце раннего рифея на территории современного Китая, когда следовало ожидать увеличения разнообразия строматолитов, происходит его сильное сокращение. Подобная картина наблюдалась и на территории Сибири и Южного Китая в венде (600–535 млн лет назад) (Семихатов, Раабен, 1996).
Вероятно, рассматриваемое эволюционное изменение строматолитов было вызвано усилением метаболизма строматолитообразователей. Оно позволяло им, с одной стороны, наращивать постройку со скоростью, значительно превышающей скорость фонового осадконакопления, с другой стороны, развивать внутреннюю среду, что выражается в возрастающих возможностях перераспределения минерального компонента в слое. По ископаемым остаткам, в частности по характеру ветвления арки и способу образования построек сложной формы, такое явление можно предполагать (Раабен, 1964; Власов, 1970; Сумина и др., 2020). В наших экспериментах перераспределение минеральных компонентов в культуре нитчатых цианобактерий обнаружено не было, однако удалось установить перераспределение внутри пленок газообразных продуктов жизнедеятельности (Сумина, Сумин, 2013).
Эти изменения макроскопических черт строения строматолитов обнаруживают коррелятивную связь с изменениями на микроструктурном уровне, определяющем прочность постройки и, таким образом, ее морфогенетические возможности.
По-видимому, тип микроструктуры характеризует не строение элементарных частей строматолитообразователя, а его способность воздействовать на перераспределение выпадающего осадка. В широком смысле – характеризует эволюционный уровень развития организма. В.А. Комар (1964) для ветвящихся строматолитов и группа авторов (Комар и др., 1965) для неветвящихся форм установили сходное эволюционное развитие во времени микроструктур этих типов построек. Общей тенденцией является нарушение сплошности темных слоев и увеличение их толщины. Для неветвящихся строматолитов этот процесс коррелятивно связан с уменьшением размеров постройки и увеличением упорядоченности в ее строении. Прочность постройки вначале обеспечивалась ее величиной, а впоследствии – структурой ее материала, что позволило увеличить относительную поверхность за счет уменьшения размеров.
В.А. Комар (1966) приводит данные, что не все слои принимают участие в образовании стенки. Обычным является выклинивание темных слоев при приближении к краевой части столбика. Светлые слои становятся тоньше и, сливаясь между собой, образуют гладкую боковую поверхность столбика. Это говорит в пользу того, что именно светлые слои, состоящие из более крупных кристаллов карбоната, образуются в результате жизнедеятельности цианобактерий, поскольку они распространяются на всю поверхность столбика – субгоризонтальные и вертикальные ее части. Темные слои занимают только субгоризонтальные части постройки (арки), т.е. состоят из осадочного материала, который не удерживался на вертикальных поверхностях.
По всей видимости, смена во времени непрерывных ленточных микроструктур штриховатыми, а затем сгустковыми, может объясняться развивающейся способностью цианобактерий к перераспределению выпадающего на их поверхность осадка. При образовании наиболее простого ленточного типа осадочный материал выпадал непрерывным ровным слоем, который в таком виде и сохранялся в структуре строматолита, не подвергаясь какой-либо переработке. Этот слой мог отражать рельеф пленки цианобактерий, отвечая неравномерно-ленточному подтипу в классификации (Комар и др., 1965). Пленка возобновляла фотосинтез после того, как нитчатые цианобактерии проползали через слой выпавшего осадка и вновь располагались на его поверхности. На этой стадии сообщество уязвимо, так как отдельные нити не связаны друг с другом и могут быть легко смыты вместе с рыхлым осадком. По этой причине, вероятно, отбор происходил в направлении тех форм, у которых эта стадия занимала, возможно, меньшее время. Вероятно, более эволюционно продвинутые формы цианобактерий приобретали способность к перераспределению осадочного материала по своей поверхности. На начальных стадиях становления этой способности в едином слое выпавшего осадка образовывались разрывы, позволявшие пленке продолжать фотосинтез без стадии “проползания”. Этому эволюционному этапу может соответствовать штриховатый тип микроструктуры. Дальнейшее развитие этой способности могло привести к образованию из материала нескольких слоев выпавшего осадка компактных скоплений, что отвечает сгустковому типу микроструктуры. При таком распределении осадка стадия “проползания” занимала минимальное время, поскольку “проползание” могло осуществляться не индивидуальными нитями, а их группами.
Таким образом, направление вертикальной смены типов микроструктур строматолитов на протяжении рифея и эволюционные преобразования организмов-строматолитообразователей могут быть соотнесены друг с другом. Эволюционные преобразования, вероятно, касаются развития информационной согласованности (Греченко и др., 2015; Grechenko, Kharitonov, 2021) и становления внутренней среды внутри сообщества цианобактерий.
Коррелятивная связь между усложнением микроструктуры и типом ветвления, вероятно, также объясняется развитием способности цианобактерий к перераспределению минерального материала. Наиболее сложные активно-ветвящиеся постройки с тонкими наклонными столбиками обладают наиболее сложными типами микроструктуры. При их образовании светлый органогенный карбонат располагается не отдельными не связанными друг с другом слоями, а образует гораздо более прочную непрерывную пространственную решетку.
Другой аспект регуляции скорости осаждения биогенного карбоната проявляется при ветвлении столбиков. Ветвлению предшествует деление арки (самой выпуклой части слоя) строматолитового столбика и возникновение вместо одной вершины двух, с разделяющей их седловиной (рис. 4). Это деление происходит за счет изменения толщины светлого слоя в центре и по краям: в центре слой становится тоньше, а по краям, наоборот, толщина его увеличивается (Раабен, 1964). Это может означать, что интенсивность выпадения биогенного карбоната, образующего светлый слой, регулируется организмом-строматолитообразователем и является проявлением его формообразующих функций. Ф.Я. Власов (1970) на основе серии шлифов ископаемых строматолитов приводит следующие стадии процесса ветвления арки – наиболее выпуклой части слоя. В апикальной части слоя перед началом ветвления возникают микроскопические бугорки – стигмы. При образовании нескольких последующих слоев бугорки каждого нового слоя сохраняют свое местоположение. Затем области расположения бугорков – полистигмы – разделяются и в дальнейшем на этих местах образуются вершины двух новых столбиков.
Рис. 4.
Деление арки, предшествующее ветвлению столбика: а – единая вершина арки в промежутках между делениями (по: Крылов, 1975), б – разделение вершины арки и последующее деление столбика.

М.Е. Раабен (1964) описывает явление срастания строматолитовых столбиков, выделяя три его типа. Первый тип – срастание столбиков, происходящее с полным объединением слоев. Второй тип – срастание с образованием специальной ткани. Третий – сближение столбиков без объединения слоев. В этом ряду, по всей видимости, просматривается становление процессов, обеспечивающих поддержание границ, определяющих целостность и индивидуальность организмов-строматолитообразователей.
Образование сложных типов микроструктур, особых типов биогенной ткани и изменение толщины биогенных слоев перед ветвлением предполагают наличие у строматолитообразователя ферментативных механизмов, позволяющих ему осуществлять тонкое управление биоминерализацией. Это предположение подтверждается обнаружением у нитчатой цианобактерии Мicrocoleus chthonoplastes фермента карбоангидразы, управляющего карбонатной биоминерализацией при образовании скелетов одноклеточных и многоклеточных эукариот, в частности у кокколитофорид и кораллов (Куприянова, Пронина, 2011).
ЗАКЛЮЧЕНИЕ
Таким образом, строматолиты образуются в результате жизнедеятельности макроскопического объекта, обладающего морфогенетической целостностью и механизмами управления биоминерализацией, и могут рассматриваться в качестве опорных минеральных структур. На это указывает наличие собственной пространственной организации и иерархической соподчиненности его частей. Изменение строматолитов во времени происходит в рамках приспособительных реакций строматолитообразователя. Наблюдаемые особенности эволюции строматолитов свидетельствуют об определенной структурной и функциональной преемственности черт строения, предполагающей наследственный контроль. Факторы среды, безусловно, оказывают влияние на этот процесс, но не непосредственно определяют форму строматолита, а через целостную реакцию строматолитообразователя, проявляющуюся на всех видимых иерархических уровнях его организации.