ВВЕДЕНИЕ
Рукокрылые – одна из самых успешных в эволюционном отношении групп млекопитающих с почти повсеместным распространением. Эти единственные активно летающие млекопитающие играют исключительно важную роль в биоценозах и экосистемах планеты. Особенно богаты и разнообразны ассоциации рукокрылых, обитающих в тропических регионах Палеарктики и Неарктики.
Позднеплейстоценовые и голоценовые позвоночные из местонахождений Кубы и других Антильских островов представляют особый интерес с позиций изучения эволюции биоразнообразия Южной и Центральной Америки, особенностей видообразования и феноменов вымираний в голоценовое время. Особенно актуальны исследования рукокрылых – одной из самых разнообразных групп млекопитающих, в составе которой много как эндемиков, так и видов с широким ареалом обитания в современности.
Предшествующие исследования показали, что развитие сообществ рукокрылых в четвертичный период было тесно связано с историей древних людей. В частности, это установлено для палеолитических памятников Горного Алтая (Росина, 2005; Rossina, 2006). Разностороннее влияние антропогенного фактора на современные популяции рукокрылых также хорошо известно (Mancina et al., 2007). Изучение четвертичных фаун рукокрылых позволяет подойти к решению многих вопросов палеобиогеографии и палеоэкологии Кубы и всего Карибского региона.
В указанном отношении весьма перспективно позднеплейстоценовое местонахождение Эль-Аброн (22°40′ с.ш., 83°28′ з.д.) в провинции Пинар-дель-Рио на западе о. Куба. Отложения пещеры Эль-Аброн богаты остатками представителей четвертичной фауны позвоночных, накопление которых происходило в основном из погадок хищных птиц (Suárez, Díaz-Franco, 2003, 2011; Suárez, 2004). При этом тафоценоз включает в себя многочисленные остатки как самих хищных птиц, которые гнездились в пещере, так и их жертв – преимущественно, грызунов, насекомоядных, мелких птиц, рептилий и земноводных (Díaz-Franco, 2001; Suárez, Díaz-Franco, 2003, 2011; Suárez, 2004; Fiol González, 2015). Находки некоторых из них представляют особый интерес с точки зрения изучения палеобиоразнообразия, палеобиогеографии, палеоэкологии и палеобиологии вымерших животных Кубы (Зеленков, Гонсалес, 2020; Syromyatnikova et al., 2020, 2021; Лопатин, 2021а, б).
Ископаемые остатки рукокрылых в местонахождении Эль-Аброн также очень многочисленны (Suárez, Díaz-Franco, 2003). Из этого местонахождения был описан один из вымерших видов американских листоносов – Phyllops silvai Suárez et Díaz-Franco, 2003.
В 2018–2019 гг. в ходе работ Совместной российско-кубинской палеонтологической экспедиции были проведены раскопки четвертичных отложений пещеры Эль-Аброн и собран новый костный материал, в т.ч. по рукокрылым (Зеленков и др., 2021). Особенно богатым оказался комплекс позвоночных из верхнеплейстоценового слоя VII, имеющего радиоуглеродную датировку 17 406 + 161 лет (калиброванный возраст 20 050–21 474 лет) (Suárez, Díaz-Franco, 2003, 2011). В настоящей работе представлены результаты изучения остатков рукокрылых из слоя VII местонахождения Эль-Аброн.
МАТЕРИАЛ И МЕТОДЫ
Геологическое, седиментологическое и биостратиграфическое описание местонахождения Эль-Аброн приведены в более ранних публикациях (Suárez, Díaz-Franco, 2003; Fiol González, 2015).
Всего было изучено 105 костных фрагментов рукокрылых, собранных из слоя VII. До видового уровня удалось определить 68 костных остатков, представленных черепами или их ростральными частями с зубами и без зубов, фрагментами верхнечелюстных и нижнечелюстных костей и изолированными зубами (табл. 1; рис. 1–3). В работе использована современная таксономическая номенклатура (Solari et al., 2019) и традиционная терминология зубных рядов (Miller, 1907).
Таблица 1.
Плейстоценовые рукокрылые из слоя VII местонахождения Эль-Аброн (n – число остатков) и их находки в других местонахождениях Кубы. Обозначения: * – вымерший или исчезающий вид; П – вид-пещерник, преимущественно населяющий убежища пещерного типа; Д – вид, преимущественно населяющий убежища древесного типа (см. Solari et al., 2019)
| Рукокрылые | n (%) | Экология | Другие ископаемые находки на Кубе |
|---|---|---|---|
| Phyllostomidae | 61 (89.7%) | ||
| Macrotus waterhousii Gray, 1843 | 40 (58.8%) | П | Antony, 1919; Silva Taboada, 1974; Orihuela, 2012; Orihuela et al., 2020а |
| Artibeus anthonyi (Wołoszyn et Silva Taboada, 1977)* | 9 (13.2%) | Д/П? | Orr, Silva Taboada, 1960; Balseiro et al., 2009; Orihuela et al., 2020а |
| Brachyphylla nana Miller, 1902 | 8 (11.8%) | П | Antony, 1919; Arredondo, 1970; Silva Taboada, 1974; Wołoszyn, Silva Taboada, 1977; Orihuela, 2012; Orihuela et al., 2020а |
| Phyllonycteris poeyi Gundlach, 1861 | 2 (2.95%) | П | Martin, 1972; Orihuela et al., 2020a, b |
| Monophyllus redmani Leach, 1821 | 2 (2.95%) | П | Silva Taboada, 1974; Orihuela et al., 2020а |
| Vespertilionidae | 5 (7.4%) | ||
| Antrozous koopmani Orr et Silva Taboada, 1960* | 5 (7.4%) | Д/П? | Viera, 2004; Orihuela et al., 2020а |
| Mormoopidae | 1 (1.45%) | ||
| Pteronotus parnellii (Gray, 1843) | 1 (1.45%) | П | Martin, 1972; Orihuela et al., 2020b |
| Molossidae | 1 (1.45%) | ||
| Rhizomops brasiliensis (Geoffroy, 1824) | 1 (1.45%) | П | Martin, 1972; Orihuela et al., 2020b |
| Всего | 68 (100%) |
Рис. 1.
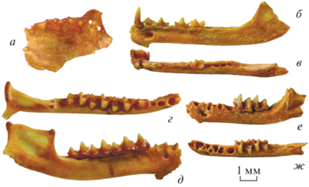
Рукокрылые из слоя VII местонахождения Эль-Аброн, Куба, верхний плейстоцен: а–к – Artibeus anthonyi (Wołoszyn et Silva Taboada, 1977): а–в – экз. ПИН, № 5807/1022, череп c обоими P4: а – с вентральной стороны, б – с правой латеральной стороны, в – с дорсальной стороны; г–е – экз. ПИН, № 5807/1023, ростральная часть черепа c обоими P4: г – с вентральной стороны, д – с правой латеральной стороны, е – с дорсальной стороны; ж, з – экз. ПИН, № 5807/1034, левая нижнечелюстная кость с p4: ж – с окклюзиальной стороны, з – с лабиальной стороны; и, к – экз. ПИН, № 5807/1036, неполная нижняя челюсть без зубов: и – с правой лабиальной стороны, к – с окклюзиальной стороны; л, м – Antrozous koopmani Orr et Silva Taboada, 1960, экз. ПИН, № 5807/1026, неполная нижняя челюсть с обоими р4: л – с окклюзиальной стороны, м – с левой лабиальной стороны; н–у – Brachyphylla nana Miller, 1902: н‒п – экз. ПИН, № 5807/1038, ростральная часть черепа без зубов: н – с вентральной стороны, о – с правой латеральной стороны, п – с дорсальной стороны; р, с – экз. ПИН, № 5807/1040, фрагмент нижней челюсти с правыми р4 и m3: р – с правой лабиальной стороны, с – с окклюзиальной стороны; т, у – экз. ПИН, № 5807/1042, неполная нижняя челюсть с обоими р4: т – с левой лабиальной стороны, у – с окклюзиальной стороны; ф–э – Macrotus waterhousii Gray, 1843: ф‒ц – экз. ПИН, № 5807/1001, череп с поврежденной мозговой коробкой и обоими рядами Р2–М3: ф – с вентральной стороны, х – с правой латеральной стороны, ц – с дорсальной стороны; ч, ш – экз. ПИН, № 5807/1002, череп с поврежденной мозговой коробкой и обоими рядами Р4–М3: ч – с вентральной стороны, ш – с правой латеральной стороны; щ, э – экз. ПИН, № 5807/1009, правая нижнечелюстная кость с р3–m3: щ – с окклюзиальной стороны, э – с лабиальной стороны.

Рис. 2.
Зубные ряды рукокрылых из слоя VII местонахождения Эль-Аброн, Куба, верхний плейстоцен: а‒в – Macrotus waterhousii Gray, 1843: а – экз. ПИН, № 5807/1001, правый верхний зубной ряд с окклюзиальной стороны; б, в – экз. ПИН, № 5807/1009, правая нижнечелюстная кость с р3–m3: б – с лабиальной стороны, в – с окклюзиальной стороны; г–е – Brachyphylla nana Miller, 1902: г – экз. ПИН, № 5807/1024, ростральная часть черепа с альвеолами зубов с окклюзиальной стороны; д, е – экз. ПИН, № 5807/1040, фрагмент нижней челюсти с правыми р4 и m3: д – с окклюзиальной стороны, е – с правой лабиальной стороны; ж–и – Phyllonycteris poeyi Gundlach, 1861: ж – экз. ПИН, № 5807/1052, фрагмент черепа без зубов с вентральной стороны; з, и – экз. ПИН, № 5807/229, неполная нижняя челюсть без зубов: з – с левой лабиальной стороны, и – с окклюзиальной стороны; к, л – Antrozous koopmani Orr et Silva Taboada, 1960, экз. ПИН, № 5807/1025, фрагмент нижней челюсти с правыми с1–m2 и левыми c1–p4: к – с правой лабиальной стороны; л – с окклюзиальной стороны.
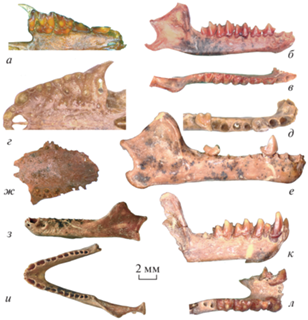
Рис. 3.
Мелкие рукокрылые из слоя VII местонахождения Эль-Аброн, Куба, верхний плейстоцен: а–в – Monophyllus redmani Leach, 1821: a – экз. ПИН, № 5807/1051, фрагмент левой верхнечелюстной кости с M1–M2 с окклюзиальной стороны; б, в – экз. ПИН, № 5807/227, фрагмент нижней челюсти с левыми с1–m1 и неполной восходящей ветвью: б – с левой лабиальной стороны, в – с окклюзиальной стороны; г, д – Pteronotus parnellii (Gray, 1843), экз. ПИН, № 5807/242, правая нижнечелюстная кость с p4–m3: г – с окклюзиальной стороны, д – с лабиальной стороны; е, ж – Rhizomops brasiliensis (Geoffroy, 1824), экз. ПИН, № 5807/232, фрагмент левой нижнечелюстной кости с m2–m3 и неполной восходящей ветвью: е – с лабиальной стороны, ж – с окклюзиальной стороны.
Изученный материал хранится в Палеонтологическом ин-те им. А.А. Борисяка РАН (ПИН; колл. № 5807) в Москве. Фотографии изготовлены с использованием электронного стереоскопического микроскопа Di-Li D-67659 с камерой HDMI-AF 16:9 и цифрового зеркального фотоаппарата Nikon D800 с объективом AF-S Micro NIKKOR 60mm f/2.8G ED.
РЕЗУЛЬТАТЫ
В тафоценозе слоя VII местонахождения Эль-Аброн установлено присутствие восьми видов рукокрылых (табл. 1), принадлежащих к четырем семействам: американских листоносов Phyllostomidae Gray, 1825 (подсемейства Macrotinae Van Den Bussche, 1992, Glossophaginae Bonaparte, 1845 и Stenodermatinae Gervais, 1856), гладконосов Vespertilionidae Gray, 1821, подбородколистых листоносовых Mormoopidae de Saussure, 1860 и бульдоговых летучих мышей Molossidae Gervais, 1856.
Подсемейство Macrotinae семейства Phyllostomidae представлено видом Macrotus waterhousii Gray, 1843 (рис. 1, ф–э). Материалы по листоносу Уотерхауса включают в себя 40 образцов: черепные фрагменты разной сохранности, верхнечелюстные и нижнечелюстные остатки (табл. 1). Изученная плейстоценовая форма морфологически сходна с современными представителями вида (Anderson, Nelson, 1965, рис. 3), которые являются типичными энтомофагами и демонстрируют характерное для этого типа питания строение зубной системы (рис. 2, а–в): удлиненную за счет крупных премоляров переднюю часть зубного ряда при значительной редукции третьих моляров. Диета современных кубинских популяций данного вида включает в себя крупных, зачастую нелетающих, насекомых Lepidoptera, Orthoptera и Odonata (см. ссылки в: Solari et al., 2019).
Подсемейство Glossophaginae представлено в тафоценозе видами Monophyllus redmani Leach, 1821 (триба Glossophagini Bonaparte, 1845; рис. 3, а–в), Brachyphylla nana Miller, 1902 (рис. 1, н–у; 2, г–е) и Phyllonycteris poeyi Gundlach, 1861 (триба Brachyphyllini Gray, 1866; рис. 2, ж–и).
Cо времен Г. Миллера (Miller, 1907) роды Phyllonycteris и Erophylla выделяют в самостоятельное подсемейство Phyllonycterinae (напр., Wetterer et al., 2000). Однако новейшие молекулярные и морфологические исследования свидетельствуют о близком филогенетическом родстве между Brachyphylla, Phyllonycteris и Erophylla, что позволяет объединять эти роды в трибу Brachyphyllini в составе подсемейства Glossophaginae (Davalos et al., 2014; Rojas et al., 2016; Solari et al., 2019).
Среди представителей подсемейства Glossophaginae самым многочисленным видом в изученном тафоценозе оказался американский листонос Brachyphylla nana (рис. 1, ж–и; 2, г–е), морфологически неотличимый от современных популяций (Swanepoel, Genoways, 1983, рис. 1). Верхние моляры этого вида утратили типичную W-образную форму стилярной полки за счет буккального смещения паракона и метакона, формирующих высокий буккальный гребень. Сильно расширенная лингвальная часть коронок верхних моляров формирует выраженный лингвальный гребень, образованный элементами, которые трудно гомологизировать с элементами коронок типично насекомоядных рукокрылых (Siles, Rios, 2019, рис. 2). Современный вид Brachyphylla nana, обитающий на Кубе, является всеядной формой, питающейся преимущественно нектаром и пыльцой, но его пища часто включает в себя фрукты и насекомых (см. ссылки в: Solari et al., 2019).
Костные остатки более мелких видов, Phyllo-nycteris poeyi (рис. 2, ж–и) и Monophyllus redmani (рис. 3, а–в), встречаются в тафоценозе значительно реже (табл. 1). Листонос Лича, Monophyllus redmani, представлен в материале фрагментом верхнечелюстной кости с M1–М2 (экз. ПИН, № 5807/1051, рис. 3, а) и нижнечелюстным фрагментом с с1–m1 и неполной восходящей ветвью (экз. ПИН, № 5807/227; рис. 3, б, в), по строению идентичными соответствующим скелетным элементам современных представителей вида (Freeman, 1998, рис. 9.5 ). Эта мелкая форма филлостомид питается нектаром и имеет очень мелкие зубы.
Остатки кубинского цветочного листоноса Phyllonycteris poeyi представлены фрагментом верхнечелюстной кости без зубов (экз. ПИН, № 5807/1052; рис. 2, ж) и лишенной зубов нижней челюстью (экз. ПИН, № 5807/229; рис. 2, з, и). Нижняя челюсть Phyllonycteris poeyi отличается от очень сходного вида Erophylla sezekorni Gundlach, 1861 наличием альвеол двух корней нижнего малого премоляра (р2), которые по длине практически равны альвеолам корней р4 (рис. 2, и). Расположение сохранившихся альвеол корней премоляров и моляров верхнечелюстного фрагмента (экз. ПИН, № 5807/1052; рис. 2, ж) свидетельствует о свободном (без соприкосновения с соседними зубами) расположении коронок верхних премоляров, как у Phyllonycteris poeyi (Mancina, 2010, рис. 2). У E. sezekorni верхний малый премоляр хоть и близок по размеру к таковому P. poeyi, но заметно сильнее зажат между коронками клыка и большого премоляра, что соответствует расположению нижних премоляров этого же вида (Baker et al., 1978, рис. 2). Известно, что P. poeyi питается нектаром и пыльцой, но также, в существенной степени, фруктами и личинками насекомых в них (см. ссылки в: Solari et al., 2019). Наблюдаемое удлинение челюстей и наличие промежутков между зубами у Phyllonycteris, по-видимому, связаны с большей специализацией данного вида к питанию нектаром, требующему соответствующего удлинения языка и вмещающего его пространства ротовой полости (Freeman, 1995).
Подсемейство Stenodermatinae представлено нередко встречающимся в тафоценозе Artibeus anthonyi (Wołoszyn et Silva Taboada, 1977) – одним из самых крупных вымерших рукокрылых Кубы (табл. 1). Морфологически форма из пещеры Эль-Аброн (рис. 1, а–к) соответствует типовой выборке вида (Wołoszyn, Silva Taboada, 1977; Balseiro et al., 2009) и находкам из других местонахождений Кубы (Orihuela et al., 2020c, рис. 10.1 ). Это типичная фруктоядная форма, на ее верхних молярах утрачена W-образная форма стилярной полки и за счет буккального смещения паракона и метакона сформирован высокий буккальный гребень (Freeman, 1998, рис. 9.5 ). Лингвальная часть коронок верхних моляров сильно расширена и формирует выраженный лингвальный гребень. Современные представители рода Artibeus, обитающие на Кубе, питаются различными фруктами, но предпочитают плоды фиговых деревьев (см. ссылки в: Solari et al., 2019).
Материалы по Vespertilionidae представлены немногочисленными остатками эндемичного для Кубы вида Antrozous koopmani Orr et Silva Taboada, 1960. Морфологически ископаемая форма (рис. 1, л, м; 2, к, л) идентична более поздним материалам по гладконосу Купмана и близка современному широко распространенному американскому виду Antrozous pallidus (Le Conte, 1856), от которого тем не менее отличается формой черепа и нижней челюсти (Orihuela et al., 2020а, рис. 2). Зубная система A. koopmani несет многие характерные признаки представителей Vespertilionidae, такие как W-образная форма буккальной части верхних моляров без выраженных дополнительных бугорков и конулей, а также миотодонтия нижних моляров. Массивные зубы при упрощении структуры их коронок, редуцированные малые премоляры и третьи моляры характерны для рукокрылых, питающихся сравнительно крупными жертвами. Рацион современных видов рода Antrozous включает разнообразных крупных членистоногих, в т.ч. скорпионов, и даже мелких позвоночных (мелкие виды грызунов, ящериц и рукокрылых; Solari et al., 2019).
В местонахождении Эль-Аброн найдены единичные остатки представителей Mormoopidae и Molossidae (табл. 1).
Семейство Mormoopidae представлено видом Pteronotus parnellii (Gray, 1843), которому принадлежит фрагмент нижнечелюстной кости с p4–m3 и восходящей ветвью с поврежденным угловым отростком (экз. ПИН, № 5807/242; рис. 3, г, д). Это типичные насекомоядные рукокрылые, верхние моляры которых имеют W-образную форму, хорошо развитые метаконуль, паралоф и металоф (Clare et al., 2013, рис. 1). Нижний зубной ряд характеризуется удлиненным рядом премоляров и нередуцированными третьими молярами (рис. 3, г, д). Пищевой рацион современных Pteronotus parnellii состоит преимущественно из мелких и крупных Lepidoptera, Diptera и Coleoptera, а также тараканов и термитов (см. ссылки в: Solari et al., 2019).
Самым мелким видом рукокрылых в тафоценозе пещеры Эль-Аброн оказался представитель бульдоговых летучих мышей – Rhizomops brasiliensis (Geoffroy, 1824), от которого обнаружен фрагмент нижнечелюстной кости с m2–m3, альвеолами i1, i2, c1, p3, p4, m1 и поврежденной восходящей ветвью (экз. ПИН, № 5807/232; рис. 3, е, ж). На основании особенностей морфологии (в частности, зубной системы) вид Tadarida brasiliensis Geoffroy, 1824 был выделен в отдельный род Rhizomops (Legendre, 1984; Hand, 1990). В Европе известны ископаемые формы молоссид, отнесенные к этому роду (Legendre, 1985). По всем морфологическим признакам, включая форму горизонтальной ветви нижней челюсти и морфологию зубного ряда, ископаемый образец из местонахождения Эль-Аброн наиболее близок к современному R. brasiliensis, в т.ч. и к некоторым ископаемым находкам этого вида (Hadler et al., 2010, рис. 2, I). В общем случае для R. brasiliensis характерно наличие трех нижних резцов (на этом настаивает ряд авторов, в том числе и Silva Taboada, 1979), однако известны популяции и с двумя нижними резцами (Legendre, 1984; Taylor et al., 2019). От Eumops nanus (Miller, 1900) ископаемый образец из пещеры Эль-Аброн отличается более удлиненной формой передней части нижней челюсти и менее редуцированными малыми премолярами (суммарная длина альвеол р2 у ископаемого образца лишь несколько уступает таковой альвеол корней р4; рис. 3, е, ж). От Nyctinomops laticaudatus Geoffroy, 1805 R. brasiliensis из местонахождения Эль-Аброн отличается диагональным расположением корней p4 (рис. 3, е, ж). В целом зубная система R. brasiliensis характерна для рукокрылых насекомоядного типа. При этом преимущественно в их рацион входят мелкие Lepidoptera и Diptera, но также жуки и муравьи (см. ссылки в: Solari et al., 2019).
ОБСУЖДЕНИЕ
Современная фауна рукокрылых Кубы – тропического типа, характеризуется большим таксономическим разнообразием, высокой популяционной численностью отдельных видов и широким спектром трофических специализаций. Куба считается центром разнообразия рукокрылых Карибского архипелага, в современности насчитывается 27 видов, принадлежащих семействам Vespertilionidae, Natalidae, Phyllostomidae, Molossidae, Noctilionidae и Mormoopidae (Mancina et al., 2017: прил. 21.1). Это составляет более 45% всех видов рукокрылых, населяющих острова Карибского бассейна (Silva Taboada, 1979; Mancina et al., 2007, 2017; Orihuela et al., 2020a, b).
Богатство фауны рукокрылых Кубы связано не только с размерами острова, но и, по-видимому, с геологическими особенностями территории, на которой очень широко распространены карстующиеся породы, что обеспечивает высокое разнообразие соответствующих биотопов и убежищ, в частности, наличие множества пещер. Действительно, более половины видов рукокрылых Кубы (61% видового разнообразия) используют карстовые полости в различные периоды жизни, а десять видов являются облигатными видами-пещерниками (Silva Taboada, 1979; Orihuela et al., 2020a, b). Семь современных видов рукокрылых – эндемики острова (см. Orihuela et al., 2020a, b).
В конце четвертичного периода фауна рукокрылых Кубы включала в себя еще как минимум шесть видов, вымерших в различные периоды на протяжении позднего плейстоцена и голоцена: Artibeus anthonyi (Wołoszyn et Silva Taboada), Phyllops vetus Anthony, P. silvai Suárez et Díaz-Franco, Mormoops magna Silva Taboada, Pteronotus pristinus Silva Taboada и Cubanycteris silvai Mancina et Garcia-Rivera. Остатки этих ископаемых видов найдены в многочисленных позднечетвертичных местонахождениях Кубы (Silva Taboada, 1974; Wołoszyn, Silva Taboada, 1977; Suárez, Díaz-Franco, 2003; Mancina, Garcia-Rivera, 2005; Orihuela et al., 2020a, b; см. табл. 1).
В послеколумбовый период из фауны Кубы исчезли подбородколист Петерса Mormoops me-galophylla Peters, 1864 и обыкновенный вампир Desmodus rotundus Geoffroy, 1810, в современности широко распространенные в Центральной и Южной Америке (Orihuela et al., 2020a, b). К вымершим видам, по-видимому, можно причислить и Antrozous koopmani – чрезвычайно редкий вид Vespertilionidae, последняя находка которого на Кубе датирована началом XX в. (Silva Taboada, 1979; Orihuela et al., 2020b).
Анализ новых ископаемых материалов из местонахождения Эль-Аброн выявил значительное таксономическое разнообразие рукокрылых (табл. 1), несмотря на сравнительно небольшой объем изученной выборки.
Самыми многочисленными оказались находки Macrotus waterhousii – мелкой насекомоядной формы американских листоносов, населяющих преимущественно пещеры и пещерные полости. В настоящее время этот вид представлен шестью подвидами, широко распространенными на территории Мексики и островов Карибского архипелага. Современные виды живут колониями до 500 особей (Solari et al., 2019). В ископаемом состоянии вид хорошо известен из плейстоценовых и голоценовых пещерных отложений Кубы (табл. 1).
Растительноядные формы американских листоносов – Artibeus anthonyi и Brachyphylla nana – представлены в тафоценозе почти одинаковым числом остатков. Современный вид B. nana – эндемик Больших Антильских о-вов, широко распространен на Кубе, причем многочисленные колонии этого вида селятся исключительно в пещерах (Solari et al., 2019). Самый крупный вид из тафоценоза пещеры Эль-Аброн – Artibeus anthonyi – является эндемичным элементом ископаемой фауны Кубы. Остатки этого вымершего вида широко встречаются в позднечетвертичных карстовых местонахождениях Кубы (табл. 1), нередко совместно с костными остатками современного вида Artibeus jamaicensis Leach, 1821 (Orihuela et al., 2020a). Видовое разнообразие рода Artibeus довольно высоко и требует таксономической ревизии. Обитающий ныне на Кубе A. jamaicensis населяет разнообразные убежища, в том числе и пещеры, где может образовывать скопления более чем в 500 особей (Solari et al., 2019). Позднечетвертичная история взаимоотношений A. anthonyi и A. jamaicensis – родственных таксонов, по-видимому, сходных по экологии и населяющих одни и те же территории, представляется очень перспективной темой палеоэкологических исследований.
Остатки еще двух представителей Phyllostomidae – Phyllonycteris poeyi и Monophyllus redmani – сравнительно редки в тафоценозе пещеры Эль-Аброн (табл. 1). В современности оба вида довольно широко распространены на территории Кубы. Распространение в современной фауне Кубы колоний вида P. poeyi тесно связано с распределением на территории так называемых “горячих пещер”, которые данный вид населяет. Этот эндемичный для Кубы вид обычно селится во внутренних частях слепых пещерных галерей, которые имеют геоморфологические особенности (такие как узкий и единственный вход), определяющие очень низкий уровень вентиляции. Внутри этих галерей P. poeyi образует многотысячные колонии, покрывая весь потолок и стены полости (Silva Taboada, 1979). Метаболическое тепло, производимое телами многочисленных зверьков, создает повышенную температуру (28–40°C) и влажность (>90%) в таких “горячих пещерах” (Mancina, 2010).
Листонос Лича Monophyllus redmani – обычный на Кубе вид, широко распространенный на Антильских и Багамских о-вах. Он населяет пещеры различного типа, в которых образует колонии до 500 тыс. особей. Единичные находки в тафоценозе пещеры Эль-Аброн едва ли отражают реальную долю данного вида в плейстоценовом сообществе; скорее всего, их немногочисленность связана с мелкими размерами и хрупкостью костей и зубов, фрагменты которых могли быть пропущены при сборах. На Кубе M. redmani часто встречается в ископаемом и полуископаемом состоянии, в том числе в погадочном материале (Orihuela et al., 2020a).
Остатки гладконоса Купмана Antrozous koopmani нередки в анализируемом тафоценозе (табл. 1). Примечательно, что этот вид обнаруживают почти исключительно в ископаемом и субфоссильном состоянии, причем самые молодые полуископаемые находки этого вида имеют позднеголоценовый возраст (Orihuela et al., 2020a, b). Единственная современная находка A. koopmani была датирована началом XX в. (две самки на территории западной Кубы; Silva Taboada, 1976). В некоторых систематических работах A. koopmani рассматривается в качестве подвида бледного гладконоса A. pallidus (Le Conte, 1856), широко распространенного на Кубе, в Мексике и западной части Южной Америки (Solari et al., 2019). Крупный современный вид A. pallidus питается в основном крупными членистоногими, но охотится и на мелких позвоночных, включая других рукокрылых. Он населяет самые разнообразные убежища, в т.ч. пещеры, где формирует небольшие колонии, как правило, до 100 особей (Solari et al., 2019; Orihuela et al., 2020a).
Все виды, установленные нами в тафоценозе слоя VII, были ранее определены из местонахождения Эль-Аброн (без морфологической характеристики и изображения остатков; Suárez, Díaz-Franco, 2003; Fiol González, 2015). Отсутствие в выборке ископаемых видов Phyllops vetus и P. silvai, а также сохранившихся до современности Erophylla sezekorni и Eptesicus fuscus (Palisot de Beauvois, 1796), вероятнее всего, связано с ограниченным объемом проанализированного материала. Все идентифицированные из слоя VII рецентные виды рукокрылых, в т.ч. представленные единичными находками Pteronotus parnellii и Rhizomops brasiliensis, и ныне широко распространены на территории провинции Пинар-дель-Рио, включая окрестности пещеры Эль-Аброн (Silva Taboada, 1979; Fiol González, 2015, рис. 7 ).
Обнаружение в тафоценозе остатков вымершего вида Artibeus anthonyi и вероятно вымершего Antrozous koopmani свидетельствует в пользу плейстоценового возраста фауны, подтвержденного данными радиоуглеродного анализа (Suárez, Díaz-Franco, 2003, 2011).
Кости рукокрылых в плейстоценовых отложениях местонахождения Эль-Аброн многочисленны и, вероятно, могли поступать в тафоценоз как из погадок хищных птиц, так и в результате естественной гибели зверьков из состава населявших пещеру колоний. Однако преобладание в тафоценозе остатков рукокрылых крупного размерного класса, которые зачастую становятся жертвами пернатых хищников, скорее свидетельствует о погадочном генезисе ископаемого материала. Данное тафономическое наблюдение согласуется с предположениями, полученными в ходе анализа остатков других мелких млекопитающих из тафоценоза местонахождения Эль-Аброн (Suárez, Díaz-Franco, 2003, 2011; Suárez, 2004).
Позднечетвертичная динамика фаун рукокрылых Кубы, как и феномены их голоценовых вымираний, связаны с уникальным сочетанием воздействия естественных факторов, таких как ураганы, цунами и изменения климата, и очевидным антропогенным влиянием, тоже влекущим существенное изменение ландшафтов и биотопов, а также исчезновение отдельных популяций и видов (Mancina et al., 2007). Всестороннее изучение данных событий прошлого может обеспечить рациональное природопользование в настоящем и будущем. Богатые позднечетвертичные материалы по позвоночным, в т.ч. рукокрылым, из карстового местонахождения Эль-Аброн – надежная и перспективная основа для таких исследований.
* * *
Исследование выполнено при финансовой поддержке Российского фонда фундаментальных исследований (РФФИ) и Министерства науки, технологий и окружающей среды Республики Куба (CITMA) в рамках проекта РФФИ и CITMA № 18-54-34004 (“Позднечетвертичные позвоночные Кубы: реликтовые фауны на пороге голоценового вымирания”). Ископаемые материалы собраны в ходе работы Совместной российско-кубинской палеонтологической экспедиции, созданной на основании соглашения между ПИН и Национальным музеем естественной истории Кубы (НМЕИ). Мы благодарим всех кубинских исследователей, принявших участие в работе экспедиции – Э. Перес Лоренсо, Х.М. Пахона Морехона (НМЕИ), Р. Рохаса-Консуэгра (Центр исследований нефти Кубы) и их коллег, и выражаем особую благодарность Н.В. Зеленкову (ПИН) за участие в сборе материалов и организационную помощь. Также мы признательны С.В. Багирову (ПИН) за помощь в фотографировании части материала.