ПАРАЗИТОЛОГИЯ, 53, 1, 2019
УДК 576.895.121
К ВОПРОСУ О ВЗАИМОРАСПОЛОЖЕНИИ ПОЛОВЫХ ЖЕЛЕЗ
У НЕКОТОРЫХ ТРЕХСЕМЕННИКОВЫХ ГИМЕНОЛЕПИДИД
© 2019 г. А. К. Галкин
Зоологический институт РАН Университетская наб., 1, СПб., 199034 Россия
е-mail: galkin_vermes@zin.ru
Поступила 03.08.2017 г.
Произведена верификации имеющихся в литературе (Скрябин, Матевосян, 1945; Спасский,
1959) схем топографических взаимоотношений гонад трехсеменниковых гименолепидид.
Схемы 9 из 15 модификаций в большей или меньшей степени не соответствуют анатомии
члеников своих «эталонных» видов - прежде всего, это касается положения центра женских
желез и среднего семенника относительно средней линии членика. Показано, что для II (вигисо-
идной) модификации характерна вариабельность расположения семенников - от острого угла
вершиной назад до линейного поперек членика. Восстановлена самостоятельность XI (прже-
вальскоидной) модификации по Скрябину и Матевосян, 1945. Дополнительно выделены 2 новые
модификации - XVIII (монопостоидная), с «эталонным» видом Diploposthe monoposthe (Dubini-
na, 1953), и XIX (скрябиноидная), с «эталонным» видом Microsomacanthus skrjabini Spasskaja,
1963.
Ключевые слова: цестоды, Hymenolepididae, половые железы, топография.
DOI: 10.1134/S0031184719010022
Семейство Hymenolepididae Ariola, 1899 насчитывает около 850 видов и является
самым богатым по видовому составу семейством цестод (Czaplinski, Vaucher, 1994).
Многочисленную группу составляют в нем трехсеменниковые виды, у которых от-
сутствуют дополнительные структуры копулятивного аппарата. Долгое время все эти
виды относили к роду Hymenolepis Weinland, 1858: О. Фурманн, ведущий специалист
по цестодам рубежа XIX-XX вв., считал его «неделимым». Для облегчения определе-
ния видов столь громоздкого рода он предложил учитывать взаиморасположение поло-
вых органов в членике (Fuhrmann, 1932).
В отечественной литературе используется классификация вариантов топографиче-
ских взаимоотношений половых желез трехсеменниковых гименолепидид, созданная
Скрябиным и Матевосян (1945). Одновременно авторы разработали в деталях типоло-
гию хоботковых крючьев - очень важного для систематики всего семейства признака.
Они даже составили таблицу для определения выделенных типов крючьев.
Современная система гименолепидид восходит к ревизии Спасского и Спасской
(1954). Ими выделено большое число новых родов, в диагнозы которых включены
как морфологические модификации хоботковых крючьев, так и модификации взаимо-
14
расположения гонад, согласно номенклатуре Скрябина и Матевосян (1945). Позднее
Спасский (1959) внес ряд изменений в диагностику, нумерацию и графическую схе-
му вариантов топографии гонад. В обновленной трактовке этот признак использован
в монографиях 1960-х гг. (Спасский, 1963; Спасская, 1966) - основополагающих оте-
чественных сводках по группе.
Оценивая на основе накопленного опыта систематическое значение топографии го-
над, Спасский (1959) подтвердил, что этот признак, в комплексе с другими, «должен
быть отражен в родовых и видовых диагнозах». Однако в диагнозах родов, даже не-
больших по объему (Drepanidotaenia Raillet, 1892; Hamatolepis Spassky, 1962), можно
встретить 2-3, а то и более вариантов расположения желез. Для рода Monosaccanthes
Czaplinski, 1967 их приведено 5 (Максимова, 1989). Нередко топографические взаимо-
отношения описывают «своими словами», без ссылки на их соответствие выделенным
типам (Спасский, Юрпалова, 1965; Максимова, 1989; Ryzhikov et al., 1985). Можно
напомнить и замечание Скрябина и Матевосян (1945) о том, что установленные ими
модификации взаиморасположения гонад «не являются совершенными».
Очевидно, что более широкое использование классификации вариантов топографии
гонад невозможно без ее пересмотра. Кратко рассмотрим этапы ее разработки.
История классификация взаиморасположения половых желез
Разнообразие топографических отношений гонад в члениках цестод рода Hymeno-
lepis впервые отобразил Фурманн (Fuhrmann, 1932. Fig. 111). Его «Схема расположе-
ния семенников у трехсеменниковых видов Hymenolepididae» включает 10 вариантов
(«диспозиций»), которые обозначены латинскими буквами (a-k). В члениках четырех
вариантов (a-d) семенники располагаются в форме треугольника. У видов, относящих-
ся к остальным вариантам, семенники лежат в поперечную линию, но их положение
относительно женских желез и экскреторных сосудов варьирует. Отмечено последова-
тельное смещение женских желез в апоральную сторону (варианты h, i, k). Для каждо-
го из вариантов автор указывает 1, реже 2 вида червей с соответствующим положением
половых желез.
К недостаткам «Схемы» Фурманна следует отнести отсутствие единообразия в изо-
бражении члеников, прежде всего их пропорций. Женские железы показаны только
у вариантов с расположением семенников в поперечную линию (при этом форма яич-
ника зачем-то детализирована), на половине схем не отмечена поральная сторона, т. к.
не показана половая бурса.
Скрябин и Матевосян (1945), классифицируя топографию гонад, различают уже
14 вариантов (модификаций), 10 из которых полностью соответствуют таковым Фур-
манна (таблица). За «эталон» той или иной топографической модификации гонад ав-
торы принимают конкретный вид паразита, в соответствии с принятой ими системой
группы. Из 10 «диспозиций» Фурмана для 4-х (a, c, f, k) «эталонными» оставлены ука-
занные им виды, для остальных они зачем-то заменены, далеко не всегда в лучшую
сторону. Каждый топографический вариант, кроме номера (римской цифры), получил
обозначение по своему «эталонному» виду (модификация I названа «коронулоид-
ной», по Dicranotaenia coronula (Dujardin, 1845); модификация II - «вигисоидной», по
D. vigisi Skrjabin et Mathevossian, 1943, и т. д.).
15
Сравнительное обозначение топографических отношений половых органов трехсеменниковых
гименолепидид
Скрябин, Матевосян, 1945
Fuhrmann, 1932
I коронулоидная
a
II вигисоидная
d
III коллароидная
b
IV эвансоидная
-
V мегалопсоидная
c
VI лонгивагиноидная
g
VII диминутоидная
e
VIII сетигероидная
h
IX микроцефалоидная
f
X нирокоидная
-
XI пржевальскоидная
i
XII ланцеолятоидная
k
XIII биакулеатоидная
-
XIV псевдоинфлятоидная
-
Авторы разработали оригинальные схемы члеников, которые формализованы значи-
тельно строже, чем у Фурманна. Для каждого из вариантов приведено положение всех
желез, как мужских, так и женских. При этом форма яичника передана условно, как бы
на ранних этапах развития (кроме модификации VI, где резко поперечно вытянутый
яичник срисован с варианта «g» Фурманна). Половые бурсы также показаны на схемах
всех модификаций, причем выделяющаяся по длине бурса у модификации IX копирует
вариант «f» Фурмана (остальные изображены очень короткими). Пропорции члеников
укладываются в 2 группы. Отношение длины члеников к их ширине для модификаций,
приуроченных к роду Dicranotaenia Railliet, 1892 (I-IV, с треугольным расположением
семенников), показано вдвое большим, чем для остальных.
Из четырех дополнительно предложенных модификаций у одной (эвансоидная -
IV) семенники образуют прямой угол, обращенный порально. У прочих они залегают
в линию поперек членика. Установлено поральное смещение женских желез (биакуле-
атоидная - XIII и псевдоинфлатоидная - XIV модификации). Расположение женских
желез в одной дорсо-вентральной плоскости с апоральным семенником обозначено как
нирокоидная (X) модификация.
Признавая валидными 3 рода трехсеменниковых гименолепидид, Скрябин и Мате-
восян ведут нумерацию топографических модификаций гонад от рода Dicranotaenia
(I-IV) через род Hymenolepis (V-IX) к роду Drepanidotaenia (X-XIV). Таким образом,
для каждого из родов предполагается наличие 4-5 разных модификаций.
Принятая Скрябиным и Матевосян система сем. Hymenolepididae не выдержала про-
верки временем: менее чем через 10 лет Спасский, Спасская (1954) разработали новую,
оригинальную систему, в которую продолжали вносить дополнения и в дальнейшем.
16
В отличие от взглядов Скрябина и Матевосян (1945) на систему гименолепидид, их
классификации крючьев и топографических взаимоотношений половых желез проч-
но вошли в научный обиход. Последняя была доработана Спасским (1959), который
привел все схемы члеников в единый вид, включил в них дополнительный элемент
- топографию вагины (или семяприемника), уточнил текстовые характеристики в мо-
дификациях и подчеркнул, что за положение яичника следует принимать «положение
анатомического центра этого органа, который имеет более постоянное положение, чем
/…/ орган в целом». Родовую принадлежность «эталонных» видов он привел в соот-
ветствие с собственной системой группы.
Два типа, XI (пржевальскоидный) и XII (ланцеолятоидный), объединены Спасским
под № XI и названием «ланцеолятоидный»: по его мнению, они «нередко переходят
один в другой в процессе онтогенеза». Дополнительно установлены два новых - для
паразитов млекопитающих XII (парвоидный, с треугольным расположением семенни-
ков и апоральным смещением женских желез) и для паразитов птиц - XV (арктикоид-
ный), по топографии желез соответствующий VIII (сетигероидному), но с прохождени-
ем вагины не впереди двух поральных семенников, а между ними. В итоге, порядковые
номера и названия модификаций топографических отношений гонад в монографии
Скрябина и Матевосян и в работах А.А. и Л.П. Спасских, опубликованных позже 1959
г., совпадают только до № X включительно.
Главным достоинством «Схемы» Фурмана является то, что варианты расположения
гонад сгруппированы с учетом их основных топографических особенностей: 4 вари-
анта (a-d) характеризуют виды с треугольным расположением семенников, 3 вариан-
та (e-g) - виды со срединным положением женских гонад и 3 варианта (h-k) - виды
с последовательным смещением их от средней линии членика.
Скрябин и Матевосян применили другой подход. Распределив модификации гонад
по родам, признаваемым ими валидными, они пронумеровали их в порядке возраста-
ния от рода к роду. Нумерация ставилась в зависимость от рода паразита. Когда же из
рода Hymenolepis было выделено множество новых родов, такая нумерация лишалась
внутреннего смысла. В 1959 г. вряд ли было оправдано сохранить ее неизменной для
первого десятка ранее выделенных модификаций с тем, чтобы нарушить на втором
десятке.
Что касается прохождения вагины, то оно определяется прежде всего положением
женских половых желез. Если поральнее женских гонад расположен всего 1 семенник,
вагина всегда проходит впереди него, что указано в характеристиках 6 модификаций
(II, III, V, VI, VII, IX). Если поральнее женских желез лежат 2 семенника, то у 3-х
модификаций (VIII, X, XII) она проходит впереди обоих семенников. Выделяя аркти-
коидную (XV) модификацию на основе прохождения вагины, Спасский никак не обо-
сновывает использование этого признака, формально не относящегося к топографии
половых желез.
Необходимость самого широкого использования признака топографических взаимо-
отношений гонад гименолепидид в систематических целях представляется несомнен-
ной. Но в опубликованных схемах и характеристиках модификаций накопились про-
тиворечия и упущения. Так, легко убедиться, что положение яичника и желточника по
Фурманну (диспозиции «e» и «f») и по Скрябину, Матевосян (VII и IX модификации)
заметно различается. Для X и XII модификаций положение среднего семенника по
17
Скрябину, Матевосян принципиально отлично от того, что показано Спасским. Требу-
ется верификация имеющихся схем путем сплошной их сверки с анатомией члеников
«эталонных» видов.
При подготовке работы сведения анатомического плана собраны из литературы -
сводок (Спасская, 1966; Yamaguti, 1959; Czaplinski, Vaucher, 1994), первоописаний (Ду-
бинина, 1953; Skrjabin, 1914) и переисследований (Галкин, 1997; Macko, 1959) видов.
Рисунки члеников, заимствованные у разных авторов, подвергнуты целенаправленной
переработке. Из них удалены не относящиеся к топографии половых желез подробно-
сти; при необходимости рисунки даны в зеркальном изображении. Для установления
«координат» той или иной половой железы в членике, что и определяет их взаиморас-
положение, на всех схемах обозначена средняя линия членика (ось симметрии, «нуле-
вой меридиан»).
Схемы всех модификаций взаиморасположения половых желез, как откорректиро-
ванные нами, так и не нуждающиеся в уточнении, будут приведены в следующей пу-
бликации. В ней предполагается дать обзор разнообразия топографических вариантов.
Результаты анализа взаиморасположения половых желез у «эталонных» видов
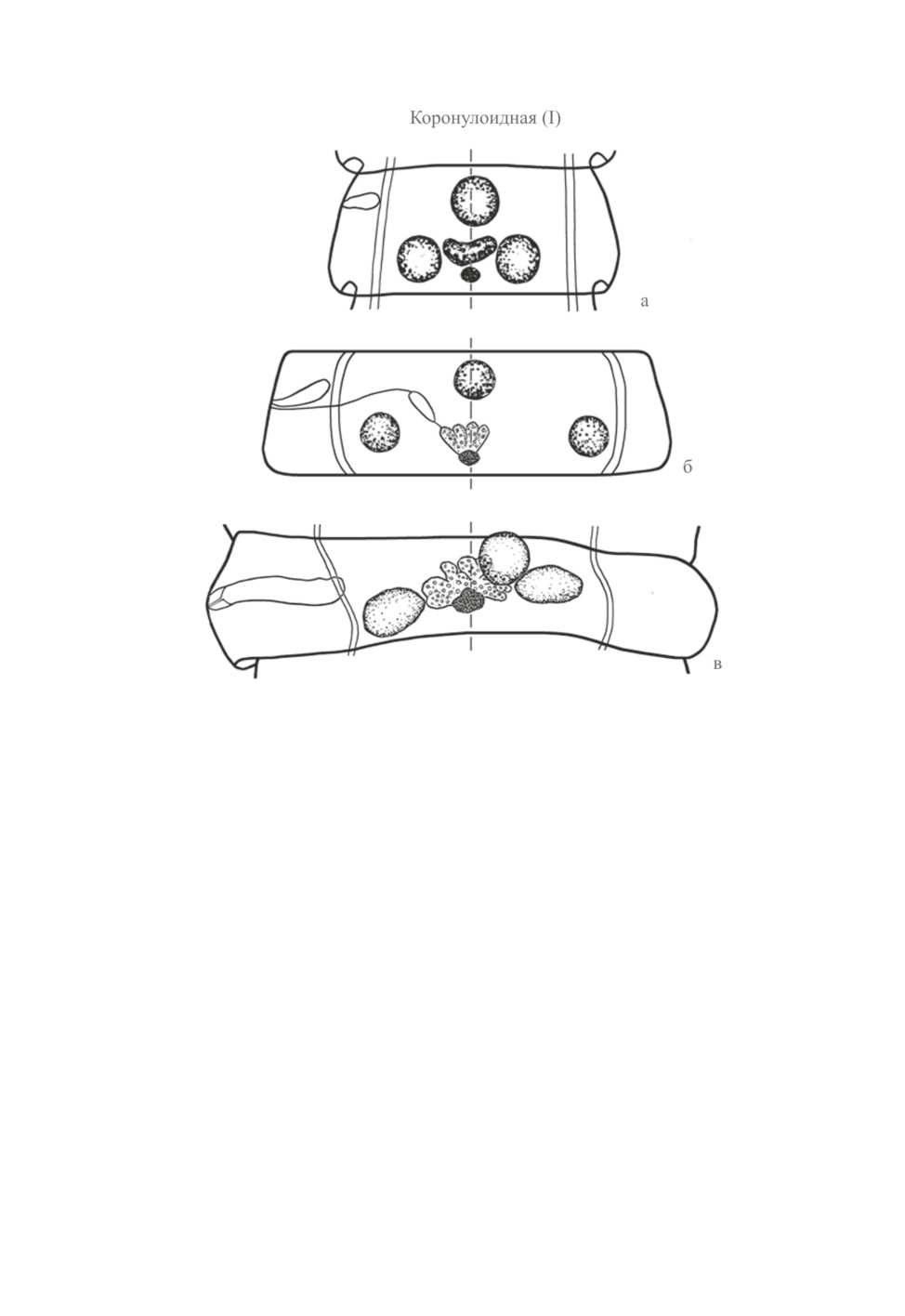
I (коронулоидная) модификация (рис. 1). Из схемы Фурманна следует, что у диспози-
ции «a» треугольник, образуемый семенниками, обращен вершиной вперед. Скрябин
и Матевосян (1945) приписали семенникам Dicranotaenia coronula, «эталонного» вида
модификации I, «характер равнобедренного треугольника», представив соответствую-
щую схему (рис. 1, а). Спасский (1959) заимствовал топографию желез из схемы 1945 г.
(рис. 1, б), но перефразировал описание, сохранив, однако, его смысл. Он подчеркнул,
что «анатомический центр яичника» лежит «позади среднего семенника». Впослед-
ствии Спасский (1963), не меняя графической схемы, заменил последние цитируемые
слова на: «позади (и порально) среднего семенника». Мало того, что с 1963 г. схема
коронулоидной модификации перестала соответствовать ее словесному описанию, по-
ложение анатомического центра яичника и среднего семенника оказалось неопреде-
ленным, не привязанным к средней линии членика (описание Скрябина и Матевосян,
1945 и Спасского, 1959 подразумевало, что средний семенник и центр яичника лежат
на этой линии). В действительности центр женских половых желез у D. coronula нахо-
дится на средней линии членика, а средний семенник смещен в апоральную сторону
на расстояние, не меньшее его диаметра - что наблюдается с самого начала закладки
семенников. Треугольник, образуемый семенниками, никак не может быть назван рав-
нобедренным - он неравнобедренный, близкий к прямоугольному (рис. 1, в).
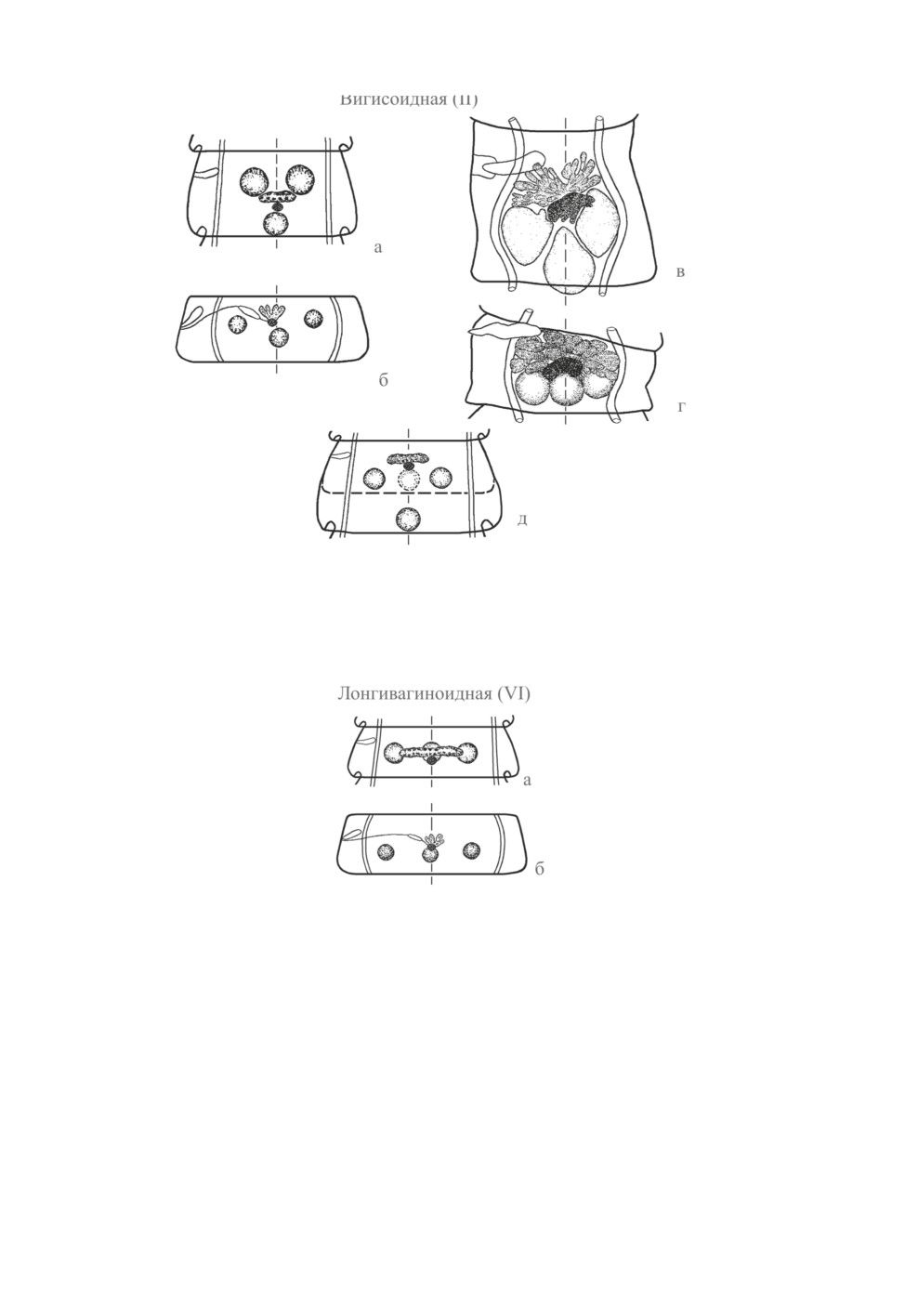
II (вигисоидная) (рис. 2). На ее схеме Скрябин и Матевосян (1945), в отличие от
Спасского (1959), отразили очень важную особенность члеников с расположени-
ем мужских желез в виде треугольника вершиной назад - их значительную длину
(рис. 2, а). «Эталонный» вид II модификации, Dicranotaenia vigisi, ныне обозначает-
ся как Microsomacanthus styloides (Fuhrmann, 1906) Spassky et Spasskaja, 1954. В гер-
мафродитных члениках вида семенники образуют равнобедренный прямоугольный
треугольник.
Хорошо исследованный типовой вид рода Retinometra Spassky, 1955, R. giranensis
(Sugimoto, 1934), дает наглядный пример непостоянства конфигурации семенников
при строго медианном положении женских желез и среднего семенника. В молодых
члениках R. giranensis зачатки семенников залегают в виде равнобедренного треуголь-
18
Рис. 1. Коронулоидная (I) модификация.
а - схема по Скрябину и Матевосян (1945); б - по Спасскому (1959),
в - половозрелый членик Dicranotaenia coronula (из Czaplinski, Vaucher, 1994, с изменениями).
ника, форма которого может быть разной - остроугольной, прямоугольной, тупоуголь-
ной. В половозрелых члениках боковые семенники, как правило, оттесняются кзади
яичником, и мужские железы принимают линейное расположение. При этом женские
железы находятся впереди всех трех семенников (Спасская, 1966) (рис. 2, в, г).
Таким образом, на схеме II модификации необходимо отразить варианты разнообра-
зия формы равнобедренного треугольника вершиной к заднему краю членика, обра-
зуемого семенниками. В члениках, длина которых не намного меньше ширины (или
квадратной формы, или поперечно вытянутых), средний семенник может значительно
отстоять назад - и мужские железы образуют острый угол. В коротких (или сократив-
шихся) члениках семенники выстраиваются под тупым углом или лежат в один попе-
речный ряд (что представляет собой развернутый угол) (рис. 2, д). Пропорции членика
меняются в ходе его созревания - в сторону уменьшения его относительной длины.
Спасский (1959) утверждает, что II модификация «легко переходит» в VI, лонгиваги-
ноидную (Рис. 3). Более осторожно пишет Спасская (1966): она отмечает, что в стро-
биле Retinometra giranensis половые железы в некоторых члениках соответствуют по
топографии не II-му, а «приближаются к VI (лонгивагиноидному) типу». Это «прибли-
жение», однако, ограничено топографией лишь мужских желез (см. ниже).
19
Рис. 2. Вигисоидная (II) модификация.
а, б - то же, что на Рис. 1; в, г - половозрелые членики стробилы Retinometra giranensis с различным
расположением семенников (из Спасской, 1966, с изменениями); д - схема, отражающая
варианты расположения семенников (от острого угла вершиной назад до линейного
поперек членика) (оригинал).
Рис. 3. Лонгивагиноидная (VI) модификация.
а, б - то же, что на Рис. 1.
Схемы III (коллароидной) и IV (эвансоидной) модификаций, у которых семенники
располагаются под прямым углом, не требуют корректировки.
Женские железы видов, относимых к III модификации, лежат на средней линии чле-
ника; вершина угла, образуемого семенниками, находится в апоральном заднем углу
членика. У IV модификации, крайне редкой (по Спасскому, 1959, найденной «один раз
у кроликов в Англии»), женские железы смещены апорально, а семенники занимают
положение, обратное III модификации (с вершиной в поральном переднем углу).
III модификация широко распространена среди гименолепидид, но ее «эталонный»
вид, Dicranotaenia collaris (Batsch, 1786), Скрябин и Матевосян выбрали крайне не-
20
удачно. Он относится к числу species inquirenda - скорее всего, при описании в XVIII
в. его собрали из фрагментов стробил разных видов. Соответственно, и род Myxolepis
Spassky, 1959, типовым видом которого Спасский (1959) избрал D. сollaris, не счи-
тается валидным (Czaplinski, Vaucher, 1994). В то же время текстовая и графическая
характеристика этой модификации очень четкие. У нее есть и «запасной» эталон -
вид Hymenolepis liguloides Gervais, 1847 (типовой вид рода Flamingolepis Spassky
et Spasskaja, 1954), принятый Фурманом в качестве образца для «диспозиции b», кото-
рая соответствует III модификации Скрябина и Матевосян (1945).
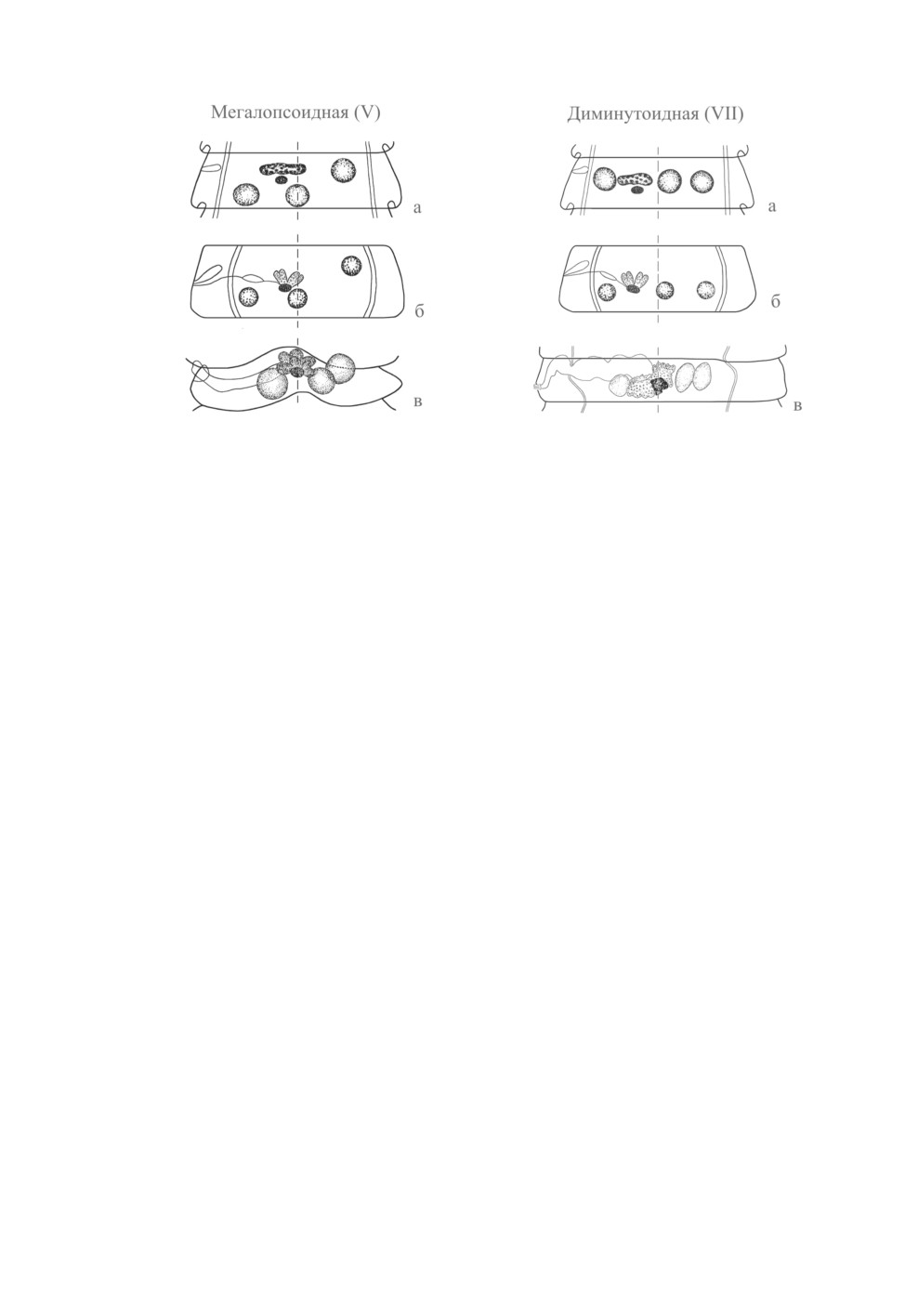
V (мегалопсоидная) модификация, как демонстрирует ее «эталонный» вид, Cloaco-
taenia megalops (Nitzsch in Creplin, 1829) Wolffhugel, 1938, характеризуется медиан-
ным положением центра женских желез. Подобная картина имеет место у и других
модификаций с треугольным расположением семенников - I, II и III. Напротив, сред-
ний семенник C. megalops смещен апорально (рис. 4, в). Т. о., топография женских
желез и среднего семенника на имеющихся в литературе схемах не отвечает реалиям
(рис. 4, а, б).
В отличие от модификации I, у модификаций III и V на одной поперечной линии
лежат не латеральные семенники, а поральный и средний. Апоральный семенник
у них сдвинут вперед. Модификации III и V объединяет смещение среднего семенни-
ка апорально и апорального - вперед. У модификации V такое смещение выражено
значительно меньше, чем у III: семенники здесь составляют тупой угол с вершиной,
занимающей апоральное положение. У модификации III апоральные семенники лежат
друг за другом на одной продольной линии - в результате чего мужские железы обра-
зуют прямой угол. Скрябин и Матевосян (1945) отмечают, что у семенников «прямой
угол, как правило, сохраняется весьма константно». Напротив, Спасский (1959) допу-
скает их переход в пределах одной стробилы от прямоугольного взаиморасположения
«в III или VII типы». О модификации VII речь будет идти ниже. Что касается располо-
жения семенников в члениках одной стробилы как в виде тупоугольного треугольни-
ка (V модификация), так и под прямым углом (III модификация), то подобные случаи
встречаются, например, у ряда видов родов Sobolevicanthus Spassky et Spasskaja, 1954
(S. gracilis (Zeder, 1803), S. dafilae (Polk, 1942)) и Passerilepis Spassky et Spasskaja, 1954
(Спасская, 1966). При этом непосредственной связи конфигурации семенников с про-
порциями членика не прослеживается, как это имеет место у модификации II. У других
видов взаиморасположение мужских желез различно уже с ранних стадий их развития,
и перехода от III модификации в V не происходит. Так, у Passerilepis passeris (Gmelin,
1790) зачатки семенников располагаются в виде тупоугольного треугольника (Спас-
ская, 1966. Рис. 346-3), тогда как у P. crenata их закладка сразу происходит под прямым
углом - 1 порально и 2 апорально, один позади другого (Галкин, 1979). Последняя кар-
тина наблюдается также в стробиле Echinolepis carioca (Magalhaes, 1898) - типичного
вида рода Echinolepis Spassky et Spasskaja, 1954.
VI (лонгивагиноидную) модификацию Скрябин и Матевосян (1945) характеризу-
ют так: «Семенники располагаются по одной горизонтали, а женские железы имеют
строго медианную локализацию». Ее «эталонным» видом они назначили Dicranotae-
nia longivaginata Fuhrmann, 1906, ныне обозначаемый как Retinometra (?) longivaginata
(Fuhrmann, 1906) Spasskaja, 1966. Спасский (1959) внес в характеристику важное уточ-
21
нение: желточник расположен «в одной сагиттальной плоскости со средним семенни-
ком».
Определяющим признаком VI модификации представляется не линейное располо-
жение семенников (боковые из них могут до ½ своего диаметра смещаться вперед от
среднего), а нахождение анатомического центра яичника и желточника в одной сагит-
тальной плоскости со средним семенником (рис. 3, а). Таким образом, женские железы
II и VI модификаций занимают принципиально разное положение: у II они лежат впе-
реди семенников, когда как у VI - вентрально от них, в той же сагиттальной плоскости.
К лонгивагиноидной модификации относится топография ряда видов родов Microso-
macanthus Lopez-Neyra, 1942 (M. acus Spassky et Jurpalova, 1964, M. compressa (Linton,
1892), M. paramicrosoma (Gasowska, 1931)), Retinometra (R. serrata (Fuhrmann, 1906))
и др.
VII (диминутоидная) модификация, согласно Скрябину и Матевосян (1945), харак-
теризуется следующим положением гонад: «женские железы отделяют поральный се-
менник от двух апоральных». Спасский (1959) справедливо заметил, что «у Hymeno-
lepis diminuta, имя которого положено в название типа [модификации - А. Г.], такое
расположение желез встречается редко» - т. е. «эталонный» вид не очень подходит
в качестве образца. О большой изменчивости в числе и положении семенников у это-
го паразита млекопитающих известно давно (Динник, Каменев, 1937; Мухин, 1967).
Все же их число, превышающее три, для H. diminuta представляется скорее ано-
малией. Когда семенников три, то 1 из них чаще расположен порально от яичника,
а 2 - апорально, но может встречаться и обратный порядок. У паразитов птиц с димину-
тоидным взаиморасположением желез число семенников постоянно (= 3) и они лежат
в поперечную линию, 1 порально и 2 апорально от женских гонад. Характеристику
этой модификации следует дополнить указанием на то, что центр женских гонад на-
ходится на средней линии членика (рис. 5, в). Эту картину безупречно отражает схе-
ма «диспозиции e» Фурманна: она основана на топографии желез Hymenolepis bisac-
cata Fuhrmann, 1906 - вида, который ныне относится к роду Sobolevicanthus Spassky
et Spasskaja, 1954 (Czaplinski, Vaucher, 1994).
Диминутоидная модификация встречается у многих видов рода Wardium Mayhew,
1925 (Спасская, 1966; Максимова, 1989). К ней же, на наш взгляд, следует относить
и случаи, когда семенники располагаются не строго поперек членика, а слегка диаго-
нально (Microsomacanthus pachycephala (Linstow, 1872)), или когда апоральный из них
«слегка сдвинут вперед». «Легким» смещением предлагаем назвать такое, когда оно не
превышает половины диаметра семенника, как, например, у рода Hamatolepis Spassky,
1962. Более значительное смещение соответствует V модификации. Диминутоидная
модификация обнаруживается и у представителей своеобразного рода Fimbriarioides
Fuhrmann, 1932, не имеющего наружной сегментации. Этот род относится к самостоя-
тельному подсемейству гименолепидил (или даже семейству Fimbriariidae Wolffhugel,
1899), на которое классификация топографических модификаций гонад ранее не рас-
пространялась.
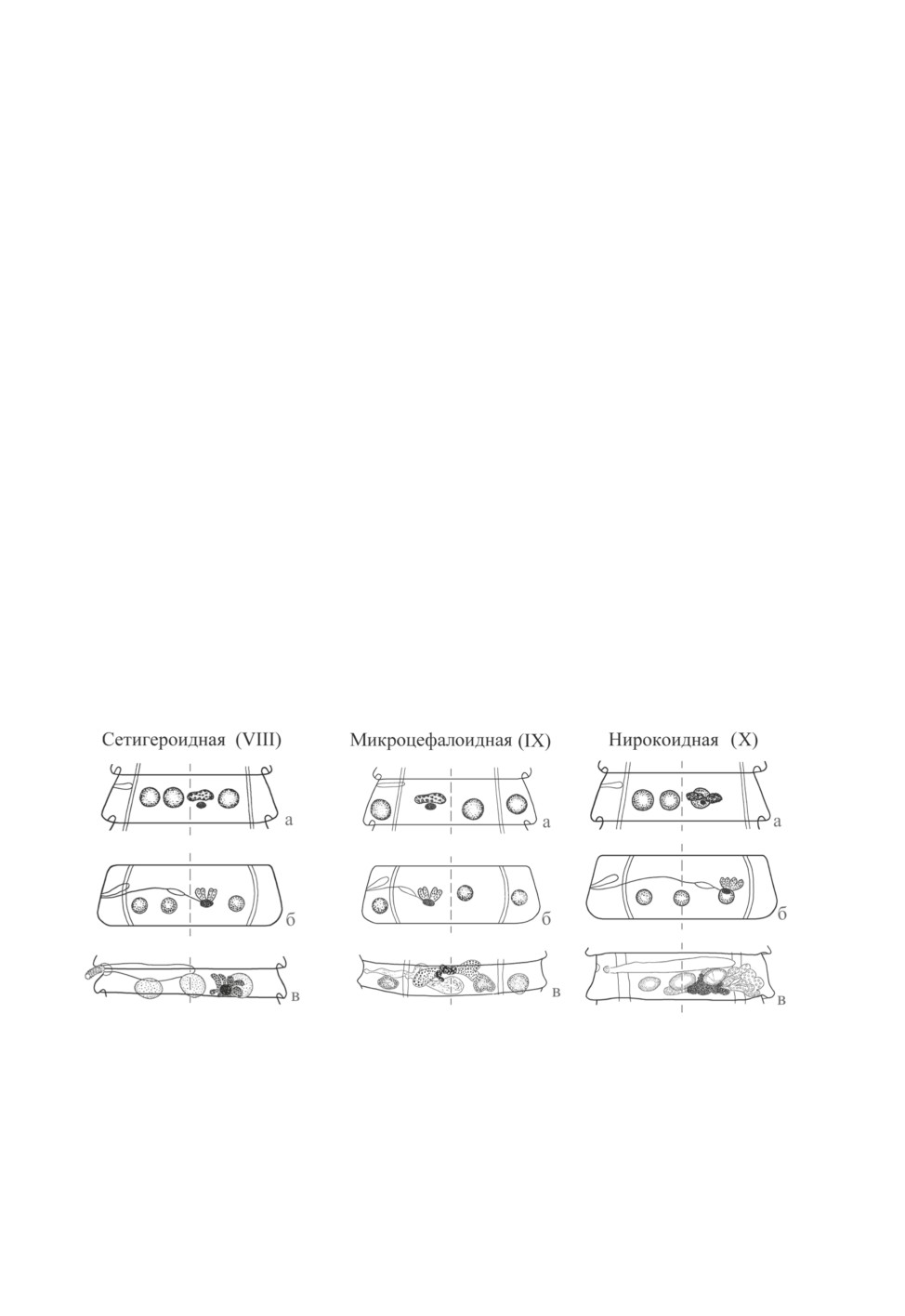
VIII (сетигероидная) модификация - схемы, представленные Скрябиным, Матево-
сян (1945) и Спасским (1959) (рис. 6, а, б), также не соответствуют действительно-
сти. Средний семенник ее «эталонного» вида, Tschertkovilepis setigera (Froelich, 1789)
Spassky et Spasskaja, 1954, расположен на средней линии членика. Женские железы
22
Рис. 4. Мегалопсоидная (V) модификация.
Рис. 5. Диминутоидная (VII) модификация.
а, б - то же, что на Рис. 1; в - половозрелый членик
а, б - то же, что на Рис. 1; в - половозрелый
Cloacotaenia megalops (из Спасской, 1966,
членик Hymenolepis diminuta (из Czaplinski,
с изменениями).
Vaucher, 1994, с изменениями).
лежат между средним и апоральным семенником так, что в половозрелых члениках
их анатомический центр может соприкасаться с поральным полюсом последнего
(рис. 6, в). Аналогичную топографию имеют половые железы Echinocotyle (Mariicoty-
le) brachicephala (Creplin, 1829) - типового вида подрода, выделенного Корнюшиным
(1983). Интересно, что этой же модификации соответствуют и половые железы у ви-
дов рода Hispaniolepis Lopez-Neyra, 1942, характеризующегося асимметрией членика
за счет разрастания его апорального края.
IX (микроцефалоидная) модификация, с «эталонным» видом Oschmarinolepis micro-
cephala (Rud., 1819), характеризуется тем, что латеральные семенники сдвинуты здесь
в латеральные поля членика, кнаружи от экскреторных сосудов. Однако положение
женских гонад и среднего семенника на имеющихся схемах передано неточно. Центр
женских гонад лежит на средней линии членика, тогда как средний семенник резко
сдвинут в сторону апорального экскреторного сосуда (рис. 7, в). В отдельных члениках
у Osch. microcephala встречаются 4 семенника (при этом «лишний» семенник залегает
в среднем поле), может наблюдаться непостоянство в положении среднего семенника
(иногда он оказывается сдвинутым поральнее женских желез) (Macko, 1959).
Схема X (нирокоидной) модификации по Скрябину, Матевосян (1945) заметно от-
личалась от действительной картины - ошибку исправил Спасский (1959) (рис. 8, б).
У «эталонного» вида, Wardoides nyrocae (Yamaguti, 1935), семенники расположены в
поперечную линию, средний из них лежит на средней линии членика, а центр женских
желез находится в одной сагиттальной плоскости с апоральным семенником. Распо-
ложение половых желез имеет здесь сходство с таковым у VIII (сетигероидной) моди-
фикации (рис. 6, в), с тем отличием, что женские железы несколько более сдвинуты в
апоральную сторону.
XI (пржевальскоидная) модификация. Ее «эталонный» вид был описан без сколекса
как Hymenolepis przewalskii Skrjabin, 1914; ныне он обозначается как Tschertkovilepis
krabbei (Kowalewski, 1895) Czaplinski, Jarecka, 1967. В диагнозе модификации сведе-
23
ний о расположении семенников нет, но из схемы (рис. 9, а) следует, что средняя линии
членика делит средний семенник на 2 неравных сегмента, как это изображено Скряби-
ным (Skrjabin, 1914) на рисунке членика H. przewalskii (рис. 9, А). Переописание Tsch.
krabbei показало, что средний семенник у вида обычно лежит на средней линии члени-
ка, а центр развивающихся женских желез расположен несколько апоральнее апораль-
ного семенника (желточник и апоральный семенник могут соприкасаться) (Czaplinski,
Jarecka, 1967) (рис. 9, Б). Такая же топография наблюдается у рода Chimaerolepis Spass-
ky et Spasskaja, 1972. По сравнению с VIII модификацией женские железы занимают
здесь зеркальное положение относительно апорального семенника, тогда как топогра-
фия семенников у модификаций VIII и XI одинакова.
Спасский (1959) объединил XI модификацию с XII (ланцеолятоидной). По нашему
мнению, оснований для этого нет: они различаются как по положению семенников от-
носительно средней линии членика, так и по положению женских желез относительно
апорального семенника.
XII (ланцеолятоидная) модификация с «эталонным» видом Drepanidotaenia lance-
olata (Bloch, 1782) Railliet, 1892 сохраняется в том понимании, которое было указано
Скрябиным и Матевосян (1945). Модификация отличается не только по топографии го-
над: расслабленные половозрелые членики этого вида имеют уникальные пропорции
- их ширина составляет более 8 мм и превосходит длину в 10-20 раз. У D. lanceolata
средний семенник полностью смещен в поральную сторону от средней линии члени-
ка, а женские половые желез даже при полном созревании располагаются значительно
апоральнее апорального семенника - на расстоянии, сопоставимом с диаметром зре-
лого семенника (рис. 10, в). Поральное смещение среднего семенника и существенный
промежуток между апоральным семенником и апорально удаленными женскими желе-
зами на всех этапах развития последних составляют главную черту XII модификации.
О ее самостоятельности и не сводимости к XI авторитетно высказался Чаплинский
Рис. 6. Сетигероидная (VIII)
Рис. 7. Микроцефалоидная (IX)
Рис. 8. Нирокоидная (X)
модификация.
модификация.
модификация.
а, б - то же, что на Рис. 1;
а, б - то же, что на Рис. 1;
а, б - то же, что на Рис. 1;
в - половозрелый членик
в - половозрелый членик
в - половозрелый членик
Tschertkovilepis setigera
Oschmarinolepis microcephala
Wardoides nyrocae
(по Галкин, 1997,
(по Macko, 1959,
(из Czaplinski, Vaucher, 1994,
с изменениями).
с изменениями).
с изменениями).
24
(Czaplinski, 1956), изучивший массовый материал по D. lanceolata: «У широких форм,
которые встречаются наиболее часто, фигурирует Тип [модификация - А. Г.] XII, тогда
как у узких экземпляров - тип, приближающийся к Типу XI». Как и в случае со II и VI
модификациями, здесь имеет место внешнее приближение, а не «слияние».
Выделенная Спасским (1959) для гименолепидид млекопитающих XII (парвоидная)
модификация, в связи с сохранением самостоятельности пржевальскоидной (XI) моди-
фикации, перенумеровывается в XV.
Эталонный» вид XIII (биакулеатоидной) модификации - кратко описанный под
названием Hymenolepis biaculeata Fuhrmann, 1909 паразит египетского гуся - имеет
резко отличную от всех других гименолепидид локализацию женских половых желез.
Яичник и желточник залегают поральнее семенников, средний из которых лежит на
средней линии стробилы. Спасский и Спасская, 1954, выделив H. biaculeata в осо-
бый род - Amphipetrovia Spassky et Spasskaja, 1954, отметили, что он имеет «обратный
(по сравнению с Drepanidotaenia) порядок расположения половых желез». С учетом
приведенных выше характеристик XI и XII модификаций, половые железы A. biac-
uleata выстроены в порядке, обратном в сравнении с H. przewalskii (= Tsch. krabbei),
а не с D. lanceolata.
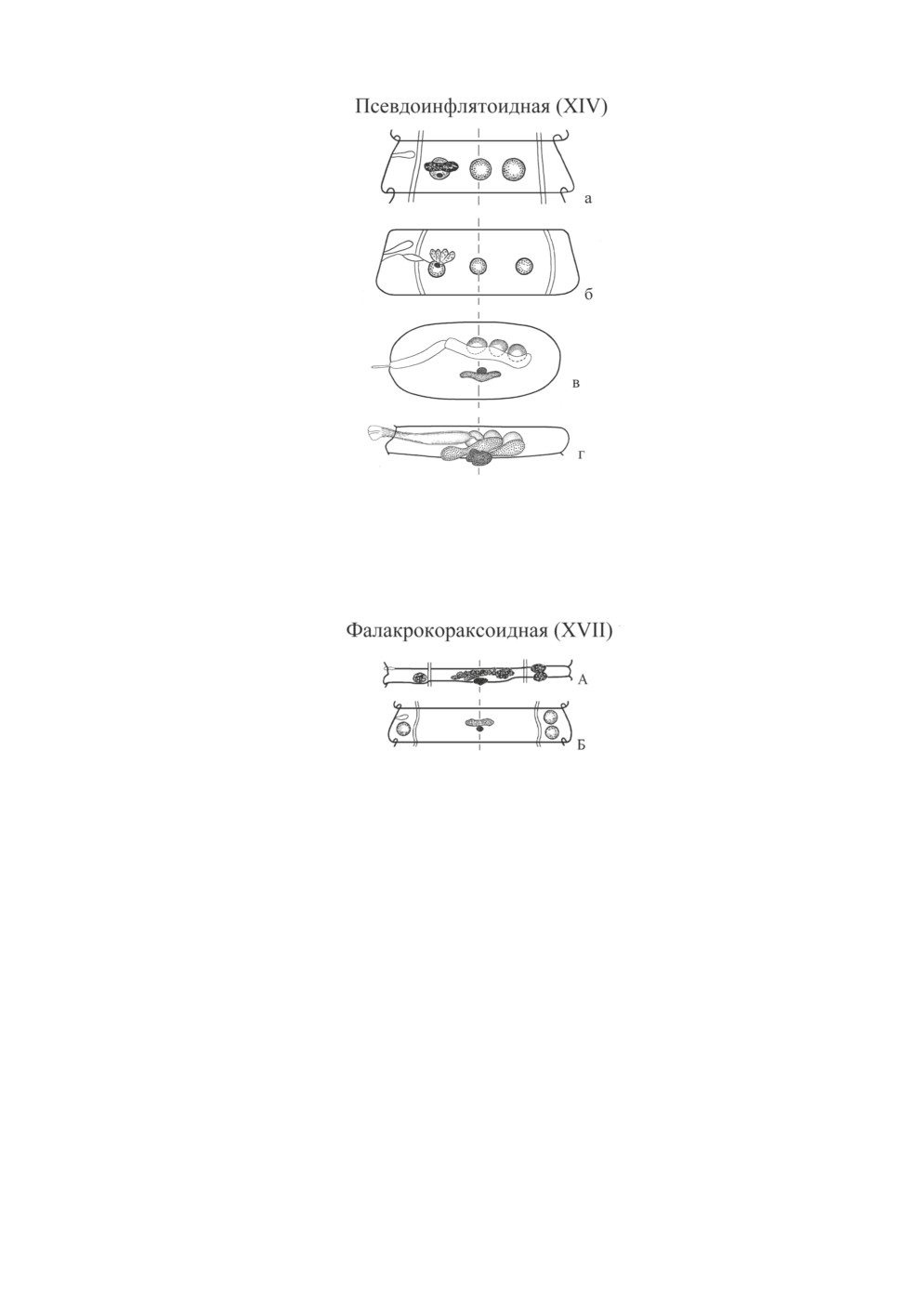
XIV (псевдоинфлятоидная) модификация «реконструирована» Скрябиным и Ма-
тевосян (1945) на основании литературных данных о виде, обозначенном ими как
Drepanidotaenia pseudoinflata Skrjabin et Mathevossian, 1942. На представленной схеме
средний семенник показан лежащим на средней линии членика, тогда как женские же-
лезы сдвинуты порально, «на одной дорсовентральной плоскости с поральным семен-
ником» (рис. 11, а). Сравнение этой схемы с помещенным в систематической части мо-
нографии авторским (Linstow, 1907) описанием и рисунком поперечного среза членика
«D. pseudoinflata» (рис. 11, в) показывает, что она не соответствует первоисточнику.
Не вдаваясь в вопрос о валидности вида и его систематическом положении, приведем
Рис. 9. Пржевальскоидная (XI) модификация.
Рис. 10. Ланцеолятоидная (XII)
модификация.
а - то же, что на Рис. 1; А - половозрелые членики
Drepanidotaenia (?) przewalskii (по Skrjabin, 1914,
а, б - то же, что на Рис. 1; в - половозрелый
с изменениями), Б - половозрелые членики
членики Drepanidotaenia lanceolata
Tschertkovilepis krabbei (по Czaplinski,
(из Czaplinski, Vaucher, 1994,
Jarecka, 1967, с изменениями).
с изменениями).
25
характеристику псевдоинфлятоидного типа в том виде, который бы соответствовал
материалам Линстова: яичник и маленький желточник расположены медианно, один
семенник (поральный) также залегает медианно, на уровне яичника, и два - апорально.
Интересно, что Ошмарин (1963) отнес к псевдоинфлятоидному типу в его правильном
понимании описанный им вид Amphipetrovia inflatocirrosa Oschmarin, 1963, у которого
медианная линия стробилы проходит через поральный край порального семенника,
против которого находится центр яичника. Однако к роду Amphipetrovia, типовой вид
которого, A. biaculeata, имеет принципиально иную топографию гонад (XIII модифи-
кация), вид никак не может быть отнесен. Регель, Бондаренко (1982) определили опи-
санный Ошмариным вид как Wardium retracta (Linstow, 1905), отметив, что женские
железы в большинстве случаев занимают положение между поральным и средним се-
менником, а на уровне порального семенника лежат «значительно реже».
Итак, все номера, присвоенные Скрябиным и Матевосян (1945) топографическим
модификациям половых желез трехсеменниковых гименолепидид, остаются без изме-
нений.
Модификации, выделенные после 1945 г., получают номера, начиная с XV-го. Пре-
жде всего, это 2 модификации, предложенные Спасским (1959):
XV модификация (парвоидная; по Спасскому, 1959 - XII) с «эталонным» видом Li-
neolepis parva (Rausch et Kuns, 1950). Положение семенников под тупым углом, верши-
ной назад и апорально, соответствует V модификации, но женские железы сдвинуты
в апоральную сторону.
XVI модификация (арктикоидная; по Спасскому, 1959 - XV) по взаиморасположе-
нию гонад напоминает VIII, но с другим прохождением вагины - впереди порального
и позади среднего семенника. Ее «эталонный» вид, Hymenolepis arctica Schiller, 1955,
Спасский (1959) перевел в род Wardium Mayhew, 1925. Нельзя не отметить, что новую
модификацию полностью проигнорировала Спасская (1966) - и в диагнозе рода Wardi-
um, и в определительной таблице видов, и тексте, посвященном W. arctica. Напротив,
Ямагути (Yamaguti, 1959) придал морфологическим особенностям H. arctica (включая
зигзагообразный ход вагины) важное систематическое значение, сделав его типовым
видом рода Decacanthus Yamaguti, 1959. Валидность этого рода признают Бондаренко,
Контримавичус (2006).
В первоописании H. arctica Шиллер (Schiller, 1955) приводит рисунки члеников
вида - молодого мужского и гермафродитного. Из них следует, что женские железы за-
кладываются и достигают зрелости не между средним и поральным семенниками, как
это имеет место у VIII модификации, а позади них. В связи с подобной локализацией
яичника половым протоками неизбежно приходится делать петлю.
XVII (фалакрокораксоидная) модификация предложена нами (Галкин, 1990) для обо-
значения взаимоположения гонад у монотипического рода Woodlandia Yamaguti, 1959.
Три семенника W. phalacrocorax (Woodland, 1929) лежат под прямым углом, но все -
в латеральные полях членика кнаружи от сосудов (1 порально и 2 апорально); женские
железы расположены на средней линии членика (рис. 12).
Дополнительно предлагаем две новые модификации.
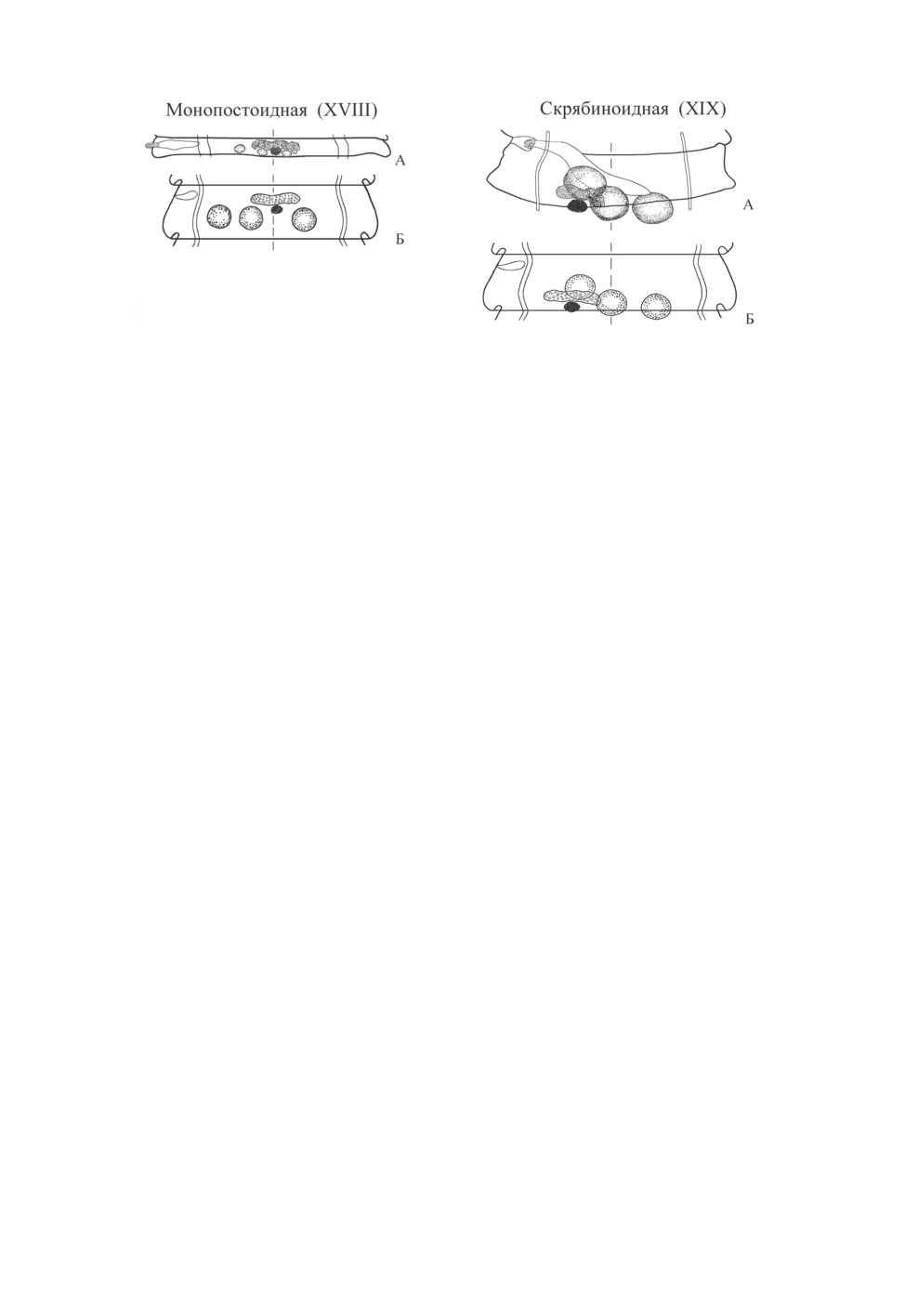
XVIII модификация (монопостоидная) с «эталонным» видом Diploposthe monoposthe
(Dubinina, 1953) Spasskaja, 1966 характеризуется развитием женских желез на средней
линии членика, тогда как средний семенник расположен порально от нее - случай,
26
Рис. 11. Псевдоинфлятоидная (XIV) модификация.
а, б - то же, что на Рис. 1; в - срез половозрелого членика Drepanidotaenia s. l. pseudoinflata
(из Скрябин, Матевосян, 1945, с изменениями); г - половозрелый членик Hymenolepis s. l. retracta
(из Спасской, 1966, с изменениями).
Рис. 12. Фалакрокораксоидная (XVII) модификация.
А - половозрелый членик Woodlandia phalacrocorax (из Yamaguti, 1959, с изменениями);
Б - схема по Галкин, 1990.
противоположный VII, диминутоидной, модификации, где средний семенник сме-
щен апорально. В половозрелых члениках D. monoposthe, которые становятся слегка
асимметричными из-за того, что «развившиеся циррусы» увеличивают их ширину с
поральной стороны, центр женских желез немного смещается в апоральную сторону
(рис. 13 А). Этой же модификации соответствует топография гонад рода Himantau-
rus Lopez-Neyra, 1942 - паразита сухопутных птиц со своеобразной асимметричной
стробилой. Спасская (1966) относит топографию Himantaurus к VIII (сетигероидному)
типу, что не соответствует действительности. Поверхностное сходство VIII и XVIII мо-
дификаций состоит в том, что женские гонады залегают между средним и апоральным
семенником. При этом у VIII модификации медианно расположен средний семенник,
а у XVIII - женские железы (рис. 12 Б), что составляет существенную разницу.
Наконец, XIX модификация (скрябиноидная) графически близка к XIV (псевдо-
инфлятоидной) по Скрябину, Матевосян (1945) (рис. 11, а), которая, как оказалась,
27
Рис. 13. Монопостоидная (XVIII)
Рис. 14. Скрябиноидная (XIX)
модификация.
модификация.
А - половозрелый членик Diploposthe monoposthe
А - половозрелый членик Microsomacanthus
(из Дубининой, 1953, с изменениями);
skrjabini (из Спасской, 1966, с изменениями);
Б - схема. Оригинал.
Б - схема. Оригинал.
не соответствует анатомии вида, избранного ее «эталоном» (см. выше). «Эталон-
ным» видом XIX модификации избран Microsomacanthus skrjabini Spasskaja, 1963
(рис. 14 А, Б). При всей изменчивости взаиморасположение половых желез, средний
семенник у этого вида чаще лежит на средней линии членика, а центр женских желез
в гермафродитных члениках сдвинут порально и совпадает с центром порального се-
менника (Спасская, 1966).
ВЫВОДЫ
Сокращение числа семенников у большой группы гименолепидидных цестод до
3-х привело к большому разнообразию вариантов взаиморасположения мужских и
женских половых желез как друг относительно друга, так и по отношению к сред-
ней линии членика. Это топографическое разнообразие укладывается в 19 модифи-
каций (типов). Как показано выше, положение гонад на схемах 9-ти из 15-ти ранее
установленных модификаций (Скрябин и Матевосян,1945; Спасский, 1959) в той или
иной степени оказалось отличным от реальной локализации органов в члениках их
«эталонных» видов. Выявленные несоответствия, по нашему мнению, связаны с тем,
что авторы классификации брали в расчет преимущественно положение отдельных же-
лез друг относительно друга, не всегда принимая во внимание местонахождение того
или органа по отношению к медиане членика. Такой подход привел Спасского (1959)
к исключению модификации XI (пржевальскоидной) по Скрябину, Матевосян (1945)
из числа самостоятельных и к объединению ее с XII (ланцеолятоидной). Но отличия в
положении как среднего семенника, так и женских желез относительно средней линии
членика у этих модификаций представляются настолько глубокими (см. рис. 9 и 10),
что эти модификации никак не могут быть сведены к одной.
Топографические взаимоотношения гонад, как и любой признак, обладают измен-
чивостью. Так, у II модификации поральный и апоральный семенники лежат на одной
поперечной линии, тогда как средний, в зависимости от длины членика, может быть
в той или иной степени смещенным назад. Залегание семенников в виде остроугольно-
го треугольника вершиной назад (крайнее состояние II модификации) возможно лишь
в члениках, имеющих значительную длину, т. е. близких по форме к квадрату. Напро-
тив, максимально выраженное смещение женских гонад в апоральную сторону, когда
они локализуются намного апоральнее апорального семенника (XII модификация),
приводит к резкому изменению пропорций членика в сторону его ширины. Остальные
28
модификации вполне «вписываются» в стандартные пропорции члеников, как они по-
казаны на схемах Спасского (1959).
Очевидно, местоположение семенников демонстрирует больший «размах изменчи-
вости», чем женских желез. Для Wardium freyei Mayhew, 1925 Бондаренко, Контри-
мавичус (2006) отмечают такую вариабельность в расположении мужских желез: «по
прямой линии, по косой, под углом, направленным вершиной вперед либо назад». При
этом, однако, средний семенник и центр женских желез почти всегда остаются на сред-
ней линии членика. Известны гименолепидиды, у которых апоральный семенник в зре-
лых члениках может почти целиком смещаться латеральнее апорального сосуда, тогда
как в молодых члениках все семенники залегают в среднем поле (Czaplinski, 1990). Но
последний пример не дает повода ни для выделения особой топографической моди-
фикации, ни для отрицания систематического значения тех модификаций, где 2 или
3 семенника постоянно локализуются экстраваскулярно.
Дивергенция по признаку топографии гонад, несомненно, имела большое значе-
ние в эволюции гименолепидид. С другой стороны, положение половых желез может
в определенной степени меняться в зависимости от степени их зрелости, а также при
мышечном сокращении стробилы. Уточнение известных и выявление новых типов то-
пографических взаимоотношений гонад трехсеменниковых гименолепидид необходи-
мо для создания новой, простой и удобной их классификации.
В перспективе крайне желательно провести изучение топографии гонад в ходе их
созревания у «эталонных» видов (на серии препаратов) - до сих пор топографические
модификации выделяли по отдельным рисункам члеников.
БЛАГОДАРНОСТИ
Работа выполнена по Государственному заданию «Разнообразие паразитарных
систем, адаптаций и путей эволюции паразитов» (номер темы: ААА-
А-А17-117030310322-3).
СПИСОК ЛИТЕРАТУРЫ
Бондаренко С. К., Контримавичус В. Л. 2006. Основы цестодологии. Т. XIV. Аплопараксиды диких и домаш-
них птиц. М., Наука, 444 с.
Галкин А. К. 1979. Морфогенез половой системы Passerilepis crenata (Cestoidea, Hymenolepididae). Парази-
тология 13 (6): 611-613.
Галкин А. К. 1990. Переописание Drepanidotaenia spinulosa и пересмотр положения вида в системе гимено-
лепидид. Паразитология 24 (5): 369-378.
Галкин А. К. 1997. Переопределение «Hymenolepis setigera» от гаг Мурманского побережья. Паразитология
31 (3): 223-230.
Динник Ю. А., Каменев В. П. 1937. Паразитические черви крыс и мышей гор. Краснодара. Труды Красно-
дарского гос. пед. ин-та 4: 175-182.
Дубинина М. Н. 1953. Ленточные черви птиц, гнездящихся в Западной Сибири. Паразитологический сбор-
ник ЗИН АН СССР 15: 117-233.
Корнюшин В. В. 1983. Анализ структуры рода Echinocotyle (Cestoda, Hymenolepididae). Паразитология 17
(4): 268-271.
Максимова А. П. 1989. Цестоды-гименолепидиды водных птиц Казахстана. Алма-Ата, Наука Казахской ССР,
220 с.
Мухин В. Н. 1967. Аномалии в строении цестод из тонкого кишечника рыжих крыс (Rattus norvegicus Berk.).
Паразитология 1 (4): 324.
Омарин П. Г. 1963. Паразитические черви млекопитающих и птиц Приморского края. М., Изд-во АН СССР,
324 с.
29
Регель К. В., Бондаренко С. К. 1982. Систематическое положение и жизненные циклы цестод Wardium re-
tracta (Linstow, 1905) comb. n. и W. pararetracta sp. n. (Cyclophyllidea, Hymenolepididae). Зоологиче-
ский журнал 61 (3): 325-335.
Скрябин К. И., Матевосян Е. М. 1945. Ленточные гельминты - гименолепидиды - домашних и охотни-
чье-промысловых птиц. М., Сельхозгиз, 488 с.
Спасская Л.П. 1966. Цестоды птиц СССР. Гименолепидиды. М., Наука, 698 с.
Спасский А. А. 1959. Уточнение классификации топографических отношений половых органов гименоле-
пидид. Зоологический журнал 38 (1): 31-37.
Спасский А. А. 1963. Основы цестодологии. Т. II, ч. 1. Гименолепидиды - ленточные черви диких и домаш-
них птиц. М., Изд-во АН СССР, 418 с.
Спасский А. А., Спасская Л. П. 1954. Построение системы гименолепидид, паразитирующих у птиц.
Тр. Гельминтол. лаб. АН СССР 7: 55-119.
Спасский А. А., Юрпалова Н. М. 1965. Echinatrium n. gen. - новый род гименолепидид гусиных птиц Чукот-
ки. Паразиты животных и растений. Кишинев: изд-во Картя молдовеняскэ 1: 104-112.
Czaplinski B. 1956. Hymenolepididae Fuhrmann, 1907 (Cestoda) parasites of some domestic and wild Anseri-
formes in Poland. Acta Parasitologica Polon 4: 175-373.
Czaplinski B., Jarecka L. 1967. Morphologie et cycle èvolutif de Tschertkovilepis krabbei (Kowalewski, 1893)
comb. n. - syn. Drepanidotaenia przewalskii (Skrjabin, 1914), nec Taenia tenuirostris Rud., 1819 (Cestoda,
Hymenolepididae). Acta Parasitologica Polon 15: 289-304.
Czaplinski B., Vaucher C. 1994. Family Hymenolepididae Ariola, 1899. In: L. F. Khalil, Jones, R. A. Bray (eds).
Keys to the Cestode Parasites of Vertebrates. Wallingford, U. K., CAB International, pp. 595-663.
Fuhrmann O. 1932. Les Tenias des oiseaux. Neuchâtel, 383 pp.
Linstow O. 1907. Neue und bekannte Helminthen. Zoologische Jahrbücher. Abt. Syst. 24: 1-18.
Macko J. K. 1959. K helmintofaune potapkovitych ptakov na Vychodnom Slovensku. Ceskoslov. parasitol. 7:
127-158.
Ryzhikov K. M., Ryšavý B., Khokhlova I. G., Tolkatcheva L. M., Kornyushin V. V. 1985. Helminths of fish-eating
birds of the Palaearctic Region II. Cestoda and Acanthocephales. M., Prague, 412 pp.
Schiller E, L, 1955. Studies on the helminths fauna of Alaska. XXIII. Some cestode parasites of eider ducks. Journal
of Parasitology 41 (1): 79-86.
Skrjabin K. I. 1914. Vogelcestoden aus Russisch Turkestan. Zoologische Jahrbücher. Abt. Syst. 37: 411-492.
Yamaguti S. 1959. Systema Helminthum. Vol. 2. The Cestodes of Vertebrates. New York & London, Interscience
Publishers, 860 pp.
CONCERNING MODIFICATIONS OF GONADS TOPOGRAPHY
IN SOME HYMENOLEPIDIDS WITH THREE TESTES
A. K. Galkin
Key words: Cestoda, Hymenolepididae, gonads, topography.
SUMMARY
Characteristics of known variations of gonads disposition in proglottids of hymenolepidids with
3 testes (Skrjabin, Mathevossian, 1945; Spassky, 1959) are checked up with the anatomy of their “mod-
el” species. Diagrams of 9 out of 15 modifications proved to be in this or that extent incorrect. II (Vigi-
soid) modification is shown to consist of a grade of variations of testes disposition - from an isosceles
acute angle with apex back up to a transversal line. Separate state of XI (Przewalskoid) modification
is confirmed. Two new modifications are distinguished, XVIII (Monoposthoid), with Diploposthe mo-
noposthe as “model” species, and XIX (Skrjabinoid) - with Microsomacanthus skrjabini.
30