ПАРАЗИТОЛОГИЯ, 53, 1, 2019
УДК 576.89/597
ФАУНА ПАРАЗИТОВ ОБЫКНОВЕННОГО ГОЛЬЯНА
PHOXINUS PHOXINUS НИЖНЕГО ТЕЧЕНИЯ
РЕК ПЕНЖИНА И ТАЛОВКА
© 2019 г. Т. Е. Буторина1*, М. В. Коваль2
1 Дальневосточный государственный технический рыбохозяйственный университет, кафедра
экологии и природопользования, ул. Луговая, 52-б, Владивосток, 690087 Россия
2 Камчатский научно-исследовательский институт рыбного хозяйства и океанографии, ул.
Набережная, 18, Петропавловск-Камчатский, 683000 Россия
*е-mail: boutorina@mail.ru
Поступила 02.07.2018 г.
Впервые проведено паразитологическое обследование речного гольяна бассейна р. Пенжина.
Найдено 32 вида паразитов, включая специфичных для гольянов Pellucidhaptor merus (Zaika,
1961) и Diplostomum phoxini (Faust, 1918), характерных для сем. Cyprinidae Trichodina intermedia
Lom, 1960, Myxobolus dujardini Thelohan, 1899, M. musculi Keysselitz, 1908, M. ellipsoides
Thelohan, 1892, Goussia carpelli (Leger et Stankovitch, 1921). У гольяна преобладают эктопарази-
ты (46.9 %), среди них доминируют инфузории (93.3 % или 43.8 % всей фауны), миксоспоридии
являются субдоминантами (21.9 %); паразиты, перешедшие на гольяна с представителей других
таксономических групп, составляют 37.5 % фауны. Большинство видов паразитов (59.3 %) от-
носится к бореальному равнинному комплексу. Рацион гольянов включает личинок амфибиоти-
ческих насекомых, хирономид, падающих в воду воздушных насекомых, нематод, нитчатые и
диатомовые водоросли.
Kлючевые слова: речной гольян, Phoxinus phoxinus, паразиты, рацион рыб, бореальный рав-
нинный комплекс, Пенжина, Таловка, Северо-Восток Азии.
DOI: 10.1134/S003118471901006X
Дальневосточные гольяны относятся к родам Phoxinus (Agassiz, 1835) и Rhynchocypris
Günther, 1889 (Ito et al., 2002; Никитин, 2010). Обыкновенный, или речной гольян
Phoxinus phoxinus (Linnaeus) является единственным представителем рода Phoxinus на
Дальнем Востоке и имеет обширный палеарктический ареал (Решетников, 2003). Па-
разитофауна этого вида изучена в реках Охота, Анадырь, верховье р. Колыма (Пугачев,
1983, 1984), Пойма и Комаровка в южном Приморье (Ермоленко, 1992), Унгра и Чуль-
ман в бассейне Алдана в южной Якутии (Буторина, Резник, 2015), гельминтофауна
и фауна паразитических ракообразных - в Вилюйском водохранилище и Индигирке
(Губанов и др., 1972; Однокурцев, 2010) и среднем течении р. Колыма (Губанов и др.,
1973) в Якутии.
61
В реках Пенжина и Таловка речной гольян обитает на всем протяжении от верховьев
до устьев и в большинстве притоков (Коваль и др., 2015а), в то время как озерный го-
льян ранее отмечался только в озерах верхнего течения р. Пенжина (Черешнев, 1996),
однако с 1975 г. не зафиксированы случаи его находок ихтиологами (Коваль и др.,
2015а). Речной гольян проник в Тихоокеанскую провинцию из Сибири через верховья
рек в послеледниковый период (Линдберг, 1955).
В бассейне р. Пенжина паразитологические исследования гольянов не проводились.
В связи с этим, цель данной работы - изучение паразитофауны обыкновенного гольяна
в нижнем течении рек Пенжина и Таловка.
МАТЕРИАЛ И МЕТОДИКА
Бассейн рек Пенжина и Таловка, включая общую устьевую часть, - самый большой
на Северо-Востоке Азии (Коваль и др., 2015а). Пенжина - третья по величине после
Амура и Анадыря река российского побережья Тихого океана длиной 713 км с пло-
щадью водосбора 73.5 тыс. км2 (Горин и др., 2015). Особенность эстуарной зоны рек
Пенжина и Таловка и Пенжинской губы - максимальные приливы (12-13 м), самые
большие из известных для рек России (Горин и др., 2015; Романенко, 2015). Ихтио-
фауна р. Пенжина представлена 21 видом рыб и рыбообразных (Коваль и др., 2015а).
Гольянов отлавливали закидным мальковым неводом размером 3х8 м в нижнем те-
чении р. Пенжина на расстоянии 30-75 км от морского побережья и в р. Таловка на
расстоянии около 40 км в июле 2015 г. Рыб в замороженном состоянии доставляли
в лабораторию, где проводили дальнейшее исследование.
Материалом для работы послужили результаты изучения 30 экз. рыб из рек Пенжина
и Таловка. В каждой из рек обследовано по 7 самок и 8 самцов. Паразитологический
анализ рыб проводили стандартными методами (Быховская-Павловская, 1985; Черны-
шева и др., 2009).
Рассчитывали стандартные показатели зараженности: экстенсивность инвазии, или
встречаемость (ЭИ) - доля зараженных рыб в выборке, в %, доверительный интервал
встречаемости (d) (Ройтман, Лобанов, 1985) и индекс обилия (ИО) - среднее число
паразитов, приходящееся на одну исследованную рыбу в выборке.
Для оценки степени сходства паразитофауны гольянов из разных водоемов исполь-
зовали попарное сравнение видовых списков паразитов с расчетом индекса общности
Чекановского -Сëренсена, который вычисляли по формуле:
КСS = 2 а/ [(а + b) + (а + с)],
где а - число общих видов при попарном сравнении, b - число видов, встречающих-
ся только в первом списке, с - число видов, встречающихся только во втором списке
(Песенко, 1982).
РЕЗУЛЬТАТЫ
В р. Пенжина самки имели длину 49-70 мм (средняя 60.14 r 2.89) и массу тела 1.10-
2.85 г (2.04 r 0.21); самцы были длиной 41-60 мм (52.00 r 2.84) с массой тела 0.65-1.90
г (1.32 r 0.17). В р. Таловка самки имели длину 46-67 мм (средняя 54.71 r 2.97) и массу
62
тела 0.65-2.30 г (1.34 r 0.25); самцы были длиной 40-55 мм (48.10 r 1.97) с массой тела
0.65-1.20 г (0.89 r 0.07). Все рыбы были половозрелыми.
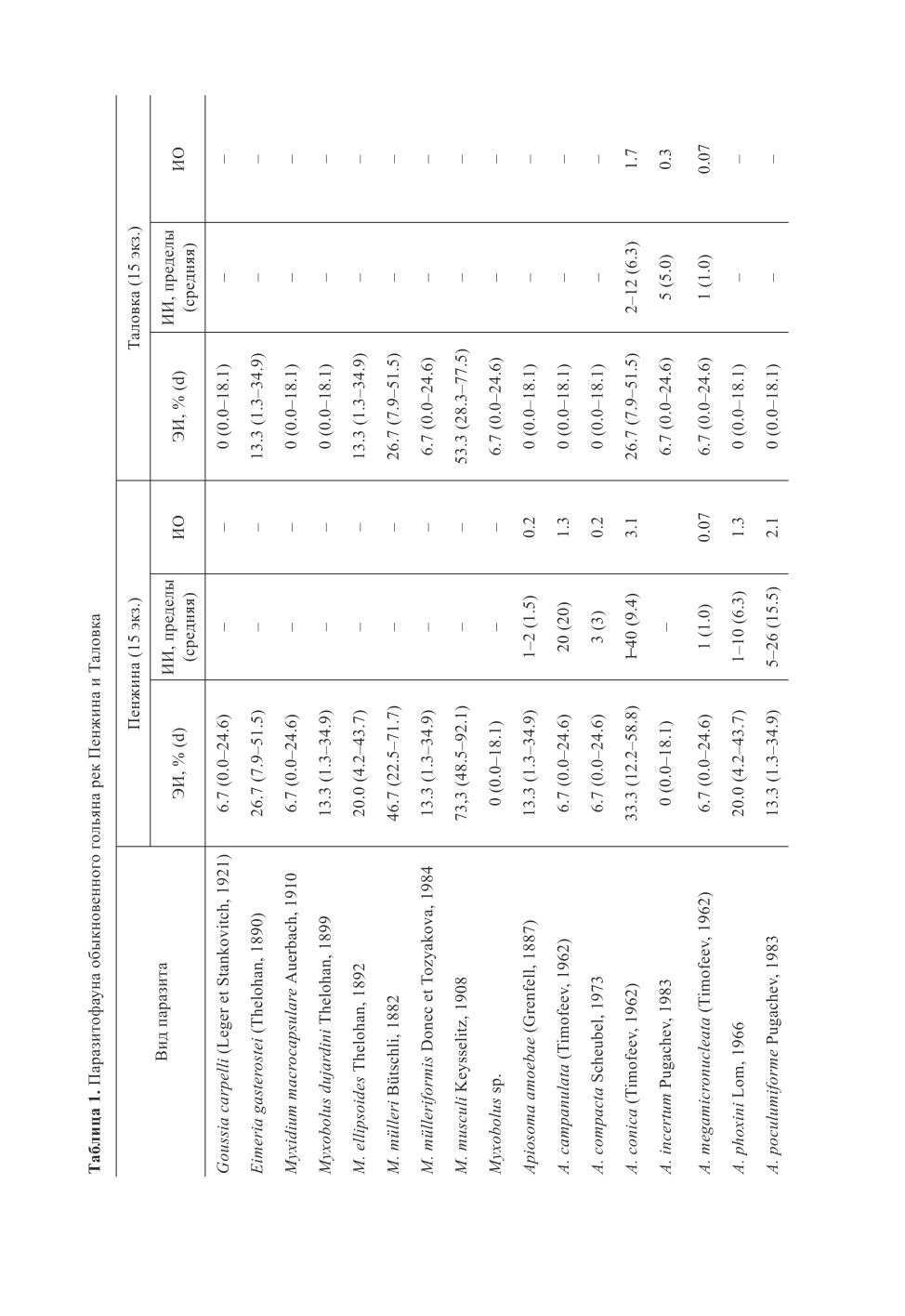
Паразитофауна речного гольяна - одна из самых разнообразных среди рыб бассейна
р. Пенжина. У речного гольяна рек Пенжина и Таловка обнаружено 32 вида паразитов
(табл. 1), в том числе 14 видов инфузорий, 7 видов миксоспоридий, 4 вида трематод,
по 2 вида моногеней и споровиков, по одному - цестод, нематод и скребней.
Практически все найденные паразиты являются пресноводными. Единственная на-
ходка трематоды Podocotyle atomon (Rudolphi, 1802) указывает на возможность зараже-
ния гольянов морскими паразитами при случайном поедании морских ракообразных
в период приливов, что связано с особенностями гидрологии рек Пенжина и Таловка
(Коваль и др., 2015б).
Почти половину фауны паразитов гольяна составляют эктопаразиты - 15 видов
(46.9 %), гельминты представлены 9 видами (28.1 %).
Из специфичных видов самым часто встречающимся был Diplostomum phoxini (Faust,
1918) (80 % рыб) и единично отмечен Pellucidhaptor merus (Zaika, 1961), специфичный
для речного гольяна (Пугачев, 2002, Русинек, 2005). В р. Пенжина у речного гольяна
найдена Apiosoma phoxini Lom, 1966 (20 %), которая «наиболее часто обнаруживалась
у гольянов» (Пугачев, 2001) и «очень редко… у других рыб» (Ермоленко, 1992). Для
гольянов бассейна р. Пенжины характерны Trichodina intermedia Lom, 1960 и Apiosoma
conica (Timofeev, 1962), отмеченные чаще других инфузорий (50 % рыб; ИИср. = 6.3
и 30 % рыб; ИИср.= 8.0 соответственно), Myxidium macrocapsulare Auerbach, 1910 -
паразит преимущественно гольянов и других карповых рыб. Найденная у гольянов
Apiosoma amoebae (Grenfell, 1887) встречается у представителей разных семейств, но
чаще всего - у колюшек и озерного гольяна (Банина, 1984).
С семейством карповых рыб связаны в своем распространении миксоспоридии
Myxobolus musculi Keysselitz, 1908, M. dujardini Thelohan, 1899, M. ellipsoides Thelohan,
1892 (Пугачев, 2001).
В целом, cпецифичные для гольянов и характерные на уровне семейства виды параз-
итов демонстрируют высокую встречаемость у речного гольяна в бассейне р. Пенжина
(21.9 %).
На первом месте по числу видов среди паразитов гольяна оказались инфузории
(14 видов, или 43.8 %), на втором - миксоспоридии (21.9 %).
Инфузории в обеих реках представлены как сидячими, так и подвижными формами
(табл. 1), которые локализовались на жабрах рыб, за исключением Trichodina urinaria
Dogiel, 1940, найденной у 2 рыб в почках (мочеточники).
Из подвижных форм чаще других отмечены T. intermedia и Paratrichodina incisa
(Lom, 1959), из сидячих - A. conica. Мы ориентировочно оценили общую численность
инфузорий у гольянов изученных выборок путем умножения индекса обилия паразита
на численность рыб. В р. Пенжина для сидячих форм она составила 159 экз., для под-
вижных форм - 131 экз., в р. Таловка - 31 экз. и 55 экз. соответственно.
Среди миксоспоридий самыми часто встречающимися оказались Myxobolus musculi
и M. mülleri Bütschli, 1882 (63.3 и 36.7 % соответственно). Споры первого вида обнару-
63
жены в мышцах, почках, жабрах, споры второго вида, помимо того, и в печени. Инвазия
гольянов остальными видами не превышает 13.3-20.0 % (табл. 1). На жабрах рыб отме-
чены цисты со спорами M. dujardini и M. ellipsoides (вид найден также в мышцах голо-
вы, мускулатуре сердца, почках). Споры Myxobolus mülleriformes Donec et Tozyakova,
1984 обнаружены в почках и печени гольянов. Жизненный цикл миксоспоридий вклю-
чает рыб (промежуточных хозяев) и олигохет (дефинитивных хозяев) (Okamura et al.,
2015). Как отмечалось нами ранее (Буторина и др., 2018), M. macrocapsulare при силь-
ной инвазии поражает протоки печени рыб (Протисты, 2007).
Кокцидии гольянов представлены специфичной для сем. Gasterosteidae Eimeria
gasterostei (Thelohan, 1890) Doflein, 1909 и Goussia carpelli (Leger et Stankovitch, 1921)
- обычным паразитом карповых рыб. Если первый вид не имеет паратенических или
промежуточных хозяев, то жизненный цикл G. сarpelli может протекать двояко: как
с участием олигохет в качестве таких хозяев, так и без них, напрямую (Steinhagen,
Korting, 1990). Ооцисты E. gasterostei найдены в печени рыб, второй вид отмечен на
стенках кишечника, в почках.
На жабрах и плавниках гольянов найдено два вида моногеней, один из которых
P. merus является специфичным палеарктическим паразитом речного гольяна (Пуга-
чев, 2002), второй представлен молодой особью рода Gyrodactylus Nordmann, 1832, не
идентифицированной до вида. Род Pellucidhaptor Price et Mizelle, 1964 связан с сем.
Catostomidae (Гусев, 1985), представители которого были доминирующей группой
пресноводной ихтиофауны в Северной Азии в палеогене и в дальнейшем были вытес-
нены карповыми рыбами (Сычевская, 1986).
Из гельминтов гольяны значительно инвазированы только метацеркариями трематод
рода Diplostomum Nordmann, 1832 (табл. 1). Массовыми являются специфичный па-
разит головного мозга гольянов D. phoxini (80 % рыб) и паразит колюшек Diplostomum
pungitii Shigin, 1965 (56.7 %), т.к. с возрастом происходит накопление инвазии (Шигин,
1986). Метацеркарии D. pungitii найдены в донной части глазного яблока.
Нематоды представлены у гольяна широко распространенным голарктическим
видом Raphidascaris acus (Bloch, 1779). Рыбы различных семейств, чаще карповые
(личинки, мальки) служат промежуточными хозяевами нематод, дефинитивными -
хищные рыбы (Moravec, 1980). Беспозвоночные (олигохеты, моллюски, планктонные
и бентосные ракообразные, личинки хирономид, мокрецов, ручейников, жуков) выпол-
няют роль паратенических хозяев (Енгашев, 1965; Косинова, Мозговой, 1965; Супря-
га, Мозговой, 1974; Smith, 1984). Инкапсулированные личинки обнаружены в ткани
печени гольянов (табл. 1), а также у колымского подкаменщика (Boutorina et al., 2017)
и единично у сиговых рыб (устное сообщение О. Ю. Бусаровой) в р. Пенжина.
Из скребней в кишечнике гольянов отмечен только Neoechinorhynchus beringianus
Mikhailowa et Atraschkevich, 2008, широко распространенный в водоемах Северо-Вос-
тока Азии (Mikhailowa, Atraschkevich, 2008). Основным окончательным хозяином это-
го вида является девятииглая колюшка, но «он найден еще у 13 видов рыб, обитающих
совместно с колюшками» (Михайлова, 2015). Промежуточными хозяевами паразита
служат пресноводные остракоды рода Candona Baird, 1845 (Атрашкевич, 2009). Дефи-
нитивные хозяева скребня в р. Пенжина - девятииглая колюшка и сопутствующие ей
66
колымский подкаменщик Cottus kolymensis Sideleva et Goto, 2012 (Boutorina et al., 2017;
Буторина и др., 2018) и речной гольян.
На основании общего анализа паразитофауны к наиболее часто встречающим-
ся видам паразитов речного гольяна бассейна р. Пенжина можно отнести трематод
D. phoxini, D. pungitii, миксоспоридий M. musculi, M. mülleri, M. ellipsoides, инфузорий
T. intermedia, A. conica, P. incisa, кокцидий E. gasterostei.
Изучение содержимого желудков показывает, что гольян бассейна р. Пенжина по-
требляет нитчатые зеленые водоросли, что характерно для этого вида (Никольский,
1956), реже - диатомовые, активно питается личинками амфибиотических насекомых
(особенно поденок), хирономид, падающими в воду имаго воздушных насекомых,
а также свободноживущими нематодами.
ОБСУЖДЕНИЕ
Паразитофауна обыкновенного гольяна бассейна р. Пенжина характеризуется соче-
танием специфичных (D. phoxini, P. merus) и широко распространенных видов параз-
итов (M. macrocapsulare, M. mülleri, M. musculi, A. phoxini, A. conica, Apiosoma robusta
(Zhukov, 1964), P. incisa, Trichodina esocis Lom, 1960, R. acus).
Подобно подкаменщикам и колюшкам, гольяны заражаются паразитами массовых
для р. Пенжина, обитающих совместно с ними видов рыб. В реках Пенжина и Таловка
у гольянов встречаются паразиты колюшек (E. gasterostei, N. beringianus), налимов -
Apiosoma megamicronucleata (Timofeev, 1962), окуней (T. urinaria), колюшек и окуня
(D. pungitii), гольяны участвуют в циркуляции паразитов щуки Triaenophorus nodulosus
(Pallas, 1781), R. аcus.
Для исследованных популяций речного гольяна характерно разнообразие и высокая
численность инфузорий, что указывает на его приуроченность к мелководным участ-
кам, галечным косам, речным заливам, пойменным проточным озерам и проточкам
(Коваль и др., 2015а). Для триходин и сидячих инфузорий большое значение имеет
обогащение воды рек Пенжина и Таловка органикой, связанное с выносом глинистых
донных осадков (Горин и др., 2015; Романенко, 2015) и поступлением большого коли-
чества аллохтонного растительного детрита преимущественно наземного происхож-
дения (Коваль и др., 2015в) в условиях периодических мощных приливов. Бытовые
органические отходы практически не загрязняют эти реки вследствие их труднодо-
ступности. Заметное обеднение фауны моногеней, по-видимому, также объясняется
отмеченной спецификой эстуарной зоны рек Пенжина и Таловка.
Разнообразие миксоспоридий и значительная инвазия гольяна диплостомидами
свидетельствуют о его контактах с олигохетами и моллюсками, а заражение R. аcus
связано с потреблением разнообразного бентоса. Планктон не играет заметной роли
в питании гольяна, о чем свидетельствуют единичные находки единственной цестоды
T. nodulosus (табл. 1).
Только два вида гельминтов локализовались в кишечниках гольянов, с учетом параз-
итов печени T. nodulosus и R. аcus, при питании в этих рыб попадает не более 12.5 %
всех видов паразитов, это сопоставимо с числом видов, найденных в почках, мышцах
и других внутренних органах.
67
Активный путь попадания в хозяина характерен для большинства паразитов гольяна
(81.2 %), соотношение видов с прямым и сложным циклом развития у него примерно
равное (56.2 и 43.8 %, соответственно) (табл. 2).
Сходство фауны паразитов гольяна рек Пенжина и Таловка составляет 63.6 %, это
ниже, чем у подкаменщиков тех же рек (78.6 %). По-видимому, на гольянах гидрологи-
ческие условия реки сказываются сильнее, чем на подкаменщиках.
Гольяны держатся примерно на тех же участках реки, что и подкаменщики, но состав
паразитов указывает на различия в их образе жизни. Если подкаменщики очень сла-
бо инвазированы сидячими инфузориями и миксоспоридиями (Boutorina et al., 2017),
то у гольянов эти паразиты составляют 50 % видов (табл. 1, 2). Гольяны активно мигри-
руют в пределах небольших участков реки или проток и, очевидно, сильнее подверга-
ются волновым и ветровым воздействиям и заражению разнообразными паразитами.
Покровы и жабры гольянов слабее защищены от нападения эктопаразитов, чем у под-
каменщиков. В отличие от гольянов - стайных рыб, образующих большие скопления
(Никольский, 1974), подкаменщики ведут оседлый, малоподвижный образ жизни, пря-
чутся под камнями, в укрытиях (Черешнев и др., 2001), что ограничивает возможности
заражения паразитами и их активное проникновение в рыб.
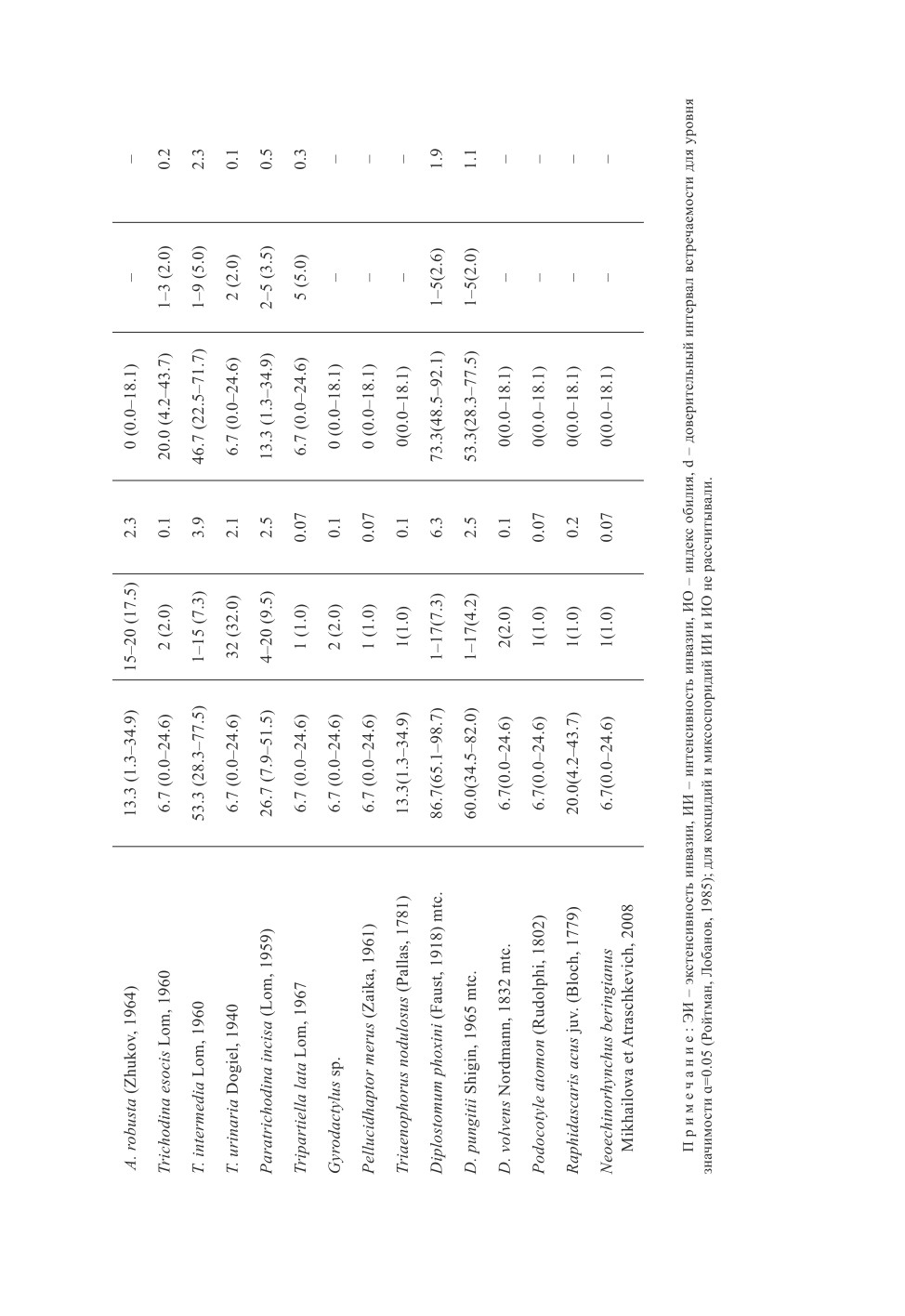
Сравнение фауны паразитов гольяна в реках Пенжина и Таловка показывает
(табл. 1), что видовое разнообразие паразитов в р. Пенжина почти вдвое больше
(30 и 16 видов соответственно), в первую очередь, за счет сидячих инфузорий и гель-
минтов. Так, только в р. Пенжина у него отмечены A. phoxini, A. amoebae, Apiosoma
poculumiforme Pugachev, 1983, A. robusta, T. nodulosus, Diplostomum volvens Nordmann,
1832, R. acus, N. beringianus и ряд других видов (табл. 1).
В р. Пенжина сильнее выражено заражение гольяна многими паразитами, в част-
ности, миксоспоридиями M. musculi, M. mülleri, кокцидии также отмечены чаще, чем
в Таловке. Индекс обилия P. incisа в р. Пенжина в 5 раз выше, чем в Таловке,
D. phoxini - в 3 раза, D. pungitii - более чем в 2 раза, больше разнообразие апиозом
(8 и 3 вида соответственно) и показатели инвазии ими, хотя заражение большинством
видов невысокое (табл. 1).
Шестнадцать из 27 видов паразитов гольяна (59.3%) представляют бореальный рав-
нинный комплекс. Полученные нами данные о паразитофауне речного гольяна, колым-
ского подкаменщика и девятииглой колюшки бассейна р. Пенжина (Boutorina et al.,
2017; Буторина и др., 2018) указывают на сильное влияние Арктического бассейна и
подтверждают существовавшую в прошлом связь Пенжины с реками Восточной Сиби-
ри (Пугачев, 1984), откуда в нее проникли как рыбы (обыкновенный гольян, тонкохво-
стый налим, сиговые, щука, подкаменщик и другие), так и паразиты (Sphaerospora
cristata Schulman, 1962, Chloromyxum dubium Auerbach, 1908, M. macrocapsulare,
M. dujardini и др.).
Cравнение паразитофауны речного гольяна бассейна р. Пенжина с таковой из рек
Унгра и Чульман в Южной Якутии (Буторина, Резник, 2015) выявило уровень сходства
31.6 %, из бассейна р. Енисей (Герман, Пронин, 2010) - 31.9 %, р. Колыма (Губанов
и др., 1973; Пугачев, 1983, 1984) - 15.8 %, из небольших рек южного Приморья Пойма
и Комаровка (Ермоленко, 1992) - 15.1 %. Разные популяции гольянов объединяет зара-
жение одними и теми же паразитами, специфичными для речного гольяна (D. phoxini,
68
Таблица 2. Характеристика паразитофауны гольяна рек Пенжина и Таловка
Группа паразитов
Пенжина
Река Пенжина
Река Таловка
(% от всех видов)
и Таловка
Всего видов паразитов,
32
30
16
в том числе:
Миксоспоридии/споровики
7 (21.9) / 2 (6.3)
6 (20.0) / 2 (6.7)
5 (31.3) / 1 (6.3)
Инфузории
14 (43.8)
13 (43.3)
8 (50.0)
Сидячие, % от всех инфузорий
9 (64.3)
8 (61.5)
3 (37.5)
Триходиниды, % от всех
5 (35.7)
5 (38.5)
5 (62.5)
инфузорий
Моногенеи
2 (6.3)
1 (6.7)
0
Цестоды
1 (3.1)
1 (3.4)
0
Трематоды
4 (12.5)
13.3
12.5
Нематоды
1 (3.1)
1 (3.4)
0
Скребни
1 (3.1)
1 (3.4)
0
Эктопаразиты / эндопаразиты
15 / 17
14 / 16
7 / 9
(46.9 / 53.1)
(46.7 / 53.3)
(43.8 / 56.2)
Специфичные
2 (6.3)
2 (6.7)
1 (6.3)
Активно заражающие
26 (81.2)
24 (80.0)
15 (93.8)
Половозрелые /личинки
5 / 4
4 / 4
0 / 2
С прямым / сложным циклом
18 / 14
16 / 14
9 / 7
(56.2 / 43.8)
(53.3 / 46.7)
(56.2 / 43.8)
P. merus), либо широко распространенными у карповых рыб (M. macrocapsulare,
M. musculi, M. mülleri, M. ellipsoides, R. acus, A. phoxini, A. campanulata (Timofeev,
1962), A. conica, A. robusta, P. incisa). Различия в числе и составе паразитов определя-
ются гидрологическими и другими особенностями водоемов, присутствием в них тех
или иных видов рыб, паразиты которых способны перейти на гольяна.
Особенности пенжинской популяции речного гольяна состоят в заражении фоновым
для водоемов Северо-Востока Азии видом N. beringianus и обеднении фауны моноге-
ней и гельминтов в целом.
Наиболее часто встречающиеся у гольяна бассейна р. Пенжина виды паразитов
(диплостомиды, апиозомы, миксоспоридии) выносливы к избыточному содержанию
органики в воде. Как было показано нами на примере рек южной Якутии (Буторина,
Резник, 2015) и на Северо-Востоке Европейской части России (Доровских, 2002), вы-
живание в условиях естественного или антропогенного загрязнения свидетельствует о
высоких биоиндикационных способностях речного гольяна и его паразитов.
Изучение линейно-весовых показателей популяций гольяна рек Пенжина и Талов-
ка и сравнение их с таковыми других популяций (Черешнев, 2008; Черешнев и др.,
2001; Буторина, Резник, 2015) показывает, что по длине и массе тела они ближе всего
к гольянам р. Унгра в Южной Якутии и чукотским гольянам. Большинство особей
69
(86.7 %) имеет длину тела в пределах 43.0-66.0 см и массу 0.65-2.45 г. Гольяны оби-
тают в реках с высоким видовым разнообразием хищников (щука, тонкохвостый
налим, камчатский хариус) и конкурентов (сиги, корюшки, колюшки, подкаменщи-
ки) и испытывают заметный пресс с их стороны, примерно такой же, как гольяны
в р. Унгра (Буторина, Резник, 2015). В последней жизненный цикл гольянов завершается
к 6-7 годам, его продолжительность определяется биотическими факторами, а также
низкой температурой воды в летний период (Черешнев и др., 2001; Буторина, Резник,
2015). Их популяционная стратегия направлена на относительно раннее созревание
и переход к размножению, чтобы компенсировать короткую продолжительность
жизни.
СПИСОК ЛИТЕРАТУРЫ
Атрашкевич Г.И. 2009. Скребни (Acanthocephala) в бассейне Охотского моря: таксономическое и экологиче-
ское разнообразие. Труды Зоологического института РАН 313 (3): 350-358.
Банина Н.Н. 1984. Подотряд Sessilina Kahl, 1933. Определитель паразитов пресноводных рыб фауны СССР.
Т. 1. Паразитические простейшие. Л., Наука, с. 281-321.
Быховская-Павловская И.Е. 1985. Паразиты рыб. Руководство по изучению. Л., Наука, 121 с.
Буторина Т.Е., Резник И.В. 2015. Фауна и структура сообществ паразитов гольяна Phoxinus phoxinus рек
южной Якутии. Паразитология 49 (3): 145-159.
Буторина Т.Е., Бусарова О.Ю., Коваль М.В. 2018. Паразитофауна полупроходной девятииглой колюшки ниж-
него течения реки Пенжина. Паразитология 52 (3): 214-223.
Герман Ю.К., Пронин Н.М. 2010. Паразитофауна и особенности сообщества паразитов гольяна обыкновен-
ного Phoxinus phoxinus (Linnaeus) рек Енисей и Кача. Паразитология 44 (1): 3-11.
Горин С.Л., Koваль M.B., Сазонов A.A., Teрский П.Н. 2015. Современный гидрологический режим нижнего
течения реки Пенжины и первые сведения о гидрологических процессах в ее эстуарии (по материалам
экспедиции 2014 г.). В кн.: Ю. П. Дьяков (гл. ред.). Исследования водных биологических ресурсов
Камчатки и северо-западной части Тихого океана. Река Пенжина и верхняя часть Пенжинской губы
(Северо-Западная Камчатка): результаты комплексных исследований 2014 г.: сб. науч. трудов. Петро-
павловск-Камчатский, КамчатНИРО, т. 37, с. 33-52.
Губанов Н.М., Находкина О.С., Однокурцев В.А. 1972. Паразитофауна рыб Колымо-Индигирской низмен-
ности. В кн.: Ф. Н. Кириллов (ред.). Рыбохозяйственное освоение озер бассейна Средней Колымы.
Якутск, ЯФ СО АН СССР, с. 140-148.
Губанов Н.М., Находкина О.С., Попов И.Е., Куличкин И.П. 1973. Паразитофауна рыб водоемов Колымской и
Индигирской низменностей. В кн.: М. В. Попов (ред.). Материалы по экологии и численности живот-
ных Якутии. Якутск, ЯФ СО АН СССР, с. 111-124.
Гусев А.В. 1985. Определитель паразитов пресноводных рыб фауны СССР. Т. 2. Паразитические многокле-
точные (первая часть). Л., Наука, 425 с.
Доровских Г.Н. 2002. Паразиты пресноводных рыб северо-востока Европейской части России (фауна, эко-
логия паразитарных сообществ, зоогеография). Автореф. дисс… д-ра биол. наук. Сыктывкар, 761 c.
Енгашев В.Г. 1965. Биология Raphidascaris acus (Bloch, 1779), эпизоотология и профилактика рафидаскари-
доза рыб. Автореф. дисс……канд. биол. наук. М., 18 с.
Ермоленко А.В. 1992. Паразиты рыб пресноводных водоемов континентальной части бассейна Японского
моря. Владивосток, ДВО РАН, 238 с.
Коваль М.В., Eсин E.В., Бугаев A.В., Kaрась В.A., Горин С.Л., Шатило И.В., Погодаев E.Г., Шубкин С.В.,
Заварина Л.O., Фролов O.В., Жаварин M.В., Коптев С.В. 2015а. Пресноводная ихтиофауна рек Пен-
жина и Taловка (Северо-Западная Камчатка). В кн.: Ю. П. Дьяков (гл. ред.). Исследования водных
биологических ресурсов Камчатки и северо-западной части Тихого океана. Река Пенжина и верхняя
часть Пенжинской губы (Северо-Западная Камчатка): результаты комплексных исследований 2014 г.:
сб. науч. трудов. Петропавловск-Камчатский, КамчатНИРО, т. 37, с. 53-145.
Коваль М.В., Горин С.Л., Калугин А.А. 2015б. Экологическая характеристика сообщества молоди рыб и
нектобентоса гиперприливного эстуария рек Пенжина и Таловка (Северо-Западная Каматка) в августе
2014 г. В кн.: Ю. П. Дьяков (гл. ред.). Исследования водных биологических ресурсов Камчатки и севе-
ро-западной части Тихого океана. Река Пенжина и верхняя часть Пенжинской губы (Северо-Западная
Камчатка): результаты комплексных исследований 2014 г.: сб. науч. трудов. Петропавловск-Камчат-
ский, КамчатНИРО, т. 37, с. 164-191.
70
Коваль М.В., Горин С.Л., Романенко Ф.А. 2015в. Условия среды и трофическая структура экосистемы гипер-
приливного эстуария (на примере устьев рек Пенжина и Таловка, Северо-Западная Камчатка). В кн.:
Материалы V Международной конференции памяти Г.Г. Винберга: «Функционирование и динамика
водных экосистем в условиях климатических изменений и антропогенных воздействий» (12-17 октя-
бря 2015 г., Санкт-Петербург). СПб, Зоологический институт РАН, с. 132-133.
Косинова В.Г., Мозговой А.А. 1965. Промежуточные хозяева Raphidascaris acus (Ascaridata: Anisakidae).
В кн.: Материалы зонального совещания «Работы по паразитофауне Юго-запада СССР». Кишинев,
с. 70-72.
Линдберг Г.У. 1955. Четвертичный период в свете биогеографических данных. Л., Изд-во АН СССР, 334 с.
Михайлова Е.И. 2015. Скребни рода Neoechinorhynchus (Acanthocephales: Neoechinorhynchidae) северо-вос-
точной Азии (таксономия, зоогеография, экология). Автореф. дис. … канд. биол. наук. CПб., 22 с.
Никитин В.Д. 2010. Гольяны острова Сахалин (систематика, распространение, экология). Автореф. дисс…
канд. биол. наук. М., 24 с.
Никольский Г.В. 1956. Рыбы бассейна Амура. M., Наука, 554 с.
Никольский Г.В. 1974. Экология рыб. M., Высшая школа, 357 с.
Однокурцев В.А. 2010. Паразитофауна рыб пресноводных водоемов Якутии. Новосибирск, Наука, 152 с.
Песенко Ю.А. 1982. Принципы и методы количествнного анализа в фаунистических исследованиях. М.,
Наука, 288 с.
Протисты: руководство по зоологии. 2007. СПб., Наука, Ч. 2, 1144 c.
Пугачев О.Н. 1983. Паразитические простейшие пресноводных рыб Северо-Востока СССР. Паразитологи-
ческий сборник 31: 158-177.
Пугачев О.Н. 1984. Паразиты пресноводных рыб Северо-Востока Азии. Л., Наука, 155 с.
Пугачев О.Н. 2001. Каталог паразитов пресноводных рыб Северной Азии. Простейшие. СПб., Изд-во
Зоологического института РАН, 242 c.
Пугачев О.Н. 2002. Каталог паразитов пресноводных рыб Северной Азии. Книдарии, моногенеи, цестоды.
Труды Зоологического института РАН 297: 1- 248.
Решетников Ю.С. (ред.). 2003. Атлас пресноводных рыб России. Т. 1. М., Наука, 379 с.
Ройтман В.А., Лобанов А.Л. 1985. Метод оценки численности гемипопуляций паразитов таксономии и био-
логии гельминтов птиц. В кн.: М. Д. Сонин (отв. ред.). Исследования по морфологии, таксономии и
биологии гельминтов птиц. Труды Гельминтологической лаборатории АН СССР 23: 102-123.
Романенко Ф.А. 2015. Рельеф и рыхлые отложения нижнего течения реки Пенжины и прилегающей ча-
сти Пенжинской губы. В кн.: Ю. П. Дьяков (гл. ред.). Исследования водных биологических ресурсов
Камчатки и северо-западной части Тихого океана. Река Пенжина и верхняя часть Пенжинской губы
(Северо-Западная Камчатка): результаты комплексных исследований 2014 г.: сб. науч. трудов. Петро-
павловск-Камчатский, КамчатНИРО, т. 37, с. 7-20.
Русинек О.Т. 2005. Паразиты рыб озера Байкал (фауна, сообщества, зоогеография). Автореф. дисс. …д-ра
биол. наук. СПб., 29 с.
Супряга В.Г., Мозговой А.А. 1974. Биологические особенности Raphidascaris acus (Ascaridata: Anisakidae) -
паразита пресноводных рыб. Паразитология 8 (6): 494-503.
Сычевская Е.К. 1986. Пресноводная палеогеновая ихтиофауна СССР и Монголии. М., Наука, 177 с.
Черешнев И.А. 1996. Биологическое разнообразие пресноводной ихтиофауны Северо-Востока России. Вла-
дивосток: Дальнаука. 197 с.
Черешнев И.А. 2008. Пресноводные рыбы Чукотки. Магадан, СВНЦ ДВО РАН, 324 с.
Черешнев И.А., Шестаков А.В., Скопец М.Б., Коротаев Ю.А., Макоедов А.Н. 2001. Пресноводные рыбы
Анадырского бассейна. Владивосток, Дальнаука, 336 с.
Чернышева Н.Б., Кузнецова Е.В., Воронин В.Н., Стрелков Ю.А. 2009. Паразитологическое исследование
рыб. СПб, ГосНИОРХ, 20 с.
Шигин А.А. 1986. Трематоды фауны СССР. Род Diplostomum. Метацеркарии. М., Наука, 253 с.
Boutorina T.E., Aseeva N.L., Koval M.V., Nguyen C.C. 2017. Parasite fauna of the bullhead Cottus kolymensis from
downstreams of the Penzhina and Talovka rivers (North-East Asia). Advances in Biology & Earth Sciences
2 (1): 92-102.
Ito Y., Sakai H., Shedko S.V., Jeon S.R. 2002. Genetic differentiations of the northern Far East Cyprinids, Phoxinus
and Rhynchocypris. Fisheries Science 68: 75-78.
Mikhailowa E. I., Atraschkevich G. I. 2008. Description and morphological variability of Neoechinorhynchus
beringianus n. sp. (Acanthocepha: Neoechinorhynchidae) from North-Eastern Asia. Systematic Parasitology
71: 41-48.
Moravec F. 1980. The lamprey Lampetra planeri as a natural intermediate host for the nematode Raphidascaris
acus. Folia parasitologica 27 (4): 347-348.
71
Okamura B., Gruhl A., Bartholomew J.L. (eds.). 2015. Myxozoan Evolution, Ecology and Development.
Switzerland, Springer Int. Publishing, 441 p.
Smith J.D. 1984. Development of Raphidascaris acus (Nematoda: Anisakidae) in paratenic, intermediate, and
definitive hosts. Canadian Journal of Zoology 62 (7): 1378-1386.
Steinhagen D., Korting W. 1990. The role of tubificid oligochaetes in the transmission of Goussia carpelli. Journal
of Parasitology 76 (1): 104-107.
PARASITE FAUNA IN MINNOW PHOXINUS PHOXINUS FROM
DOWNSTREAMS OF THE PENZHINA AND TALOVKA RIVERS
Т. Е. Boutorina, М. В. Коval
Keywords: minnow, Phoxinus phoxinus, parasites, fish ration, boreal plain complex,
Penzhina River, Talovka River, North-East of Asia.
SUMMARY
During the first parasitological survey of minnow Phoxinus phoxinus, inhabiting the Penzhina River
basin, 32 species of parasites were found, including specific to this fish: Pellucidhaptor merus (Zaika,
1961), Diplostomum phoxini (Faust, 1918) and the typical for cyprinid fishes Trichodina intermedia
Lom, 1960, Myxobolus dujardini Thelohan, 1899, M. musculi Keysselitz, 1908, M. ellipsoides
Thelohan, 1892, Goussia carpelli (Leger et Stankovitch, 1921). Ectoparasites were predominating
in the fish host (46.9 %), among them the majority were infusorians (93.3 or 43.8 % of the whole
fauna), myxosporeans served as subdominants. Parasites, that migrated to minnows from fish of other
taxonomic groups, made up 37.5 %. Most parasites (59.3 %) were referred to the boreal plain complex.
The minnows’ diet includes the larvae of amphibiotic insects, chironomids, insects falling in water,
nematodes, filamentous and diatoms algae.
72