ПАРАЗИТОЛОГИЯ, 53, 2, 2019
УДК 576.895.121:595.143.7(282.256.8)
К МОРФОЛОГИИ МЕТАЦЕСТОД РОДА KOWALEWSKIUS -
ПАРАЗИТОВ ПИЯВОК ERPOBDELLA OCTOCULATA
БАССЕЙНА КОЛЫМЫ
© 2019 г. К. В. Регель*, Н. А. Поспехова
Институт биологических проблем Севера ДВО РАН, Портовая ул., 18, Магадан, 685000 Россия
* е-mail: kire@ibpn.ru
Поступила 11.07.2018 г.
После доработки 27.08.2018 г.
Принята к публикации 27.08.2018 г.
Обобщены оригинальные данные о встречаемости трёх видов метацестод у пиявок Erpobdella
octoculata L. в озёрах Верхней Колымы. Приведено морфологическое описание двух видов ме-
тацестод рода Kowalewskius Yamaguti, 1959, локализованных в соединительной ткани хозяев.
Ранее высказанное предположение о сходстве метацестод Kowalewskius spp. с модификацией
циклоцерк (или циклоцистицеркоид) не нашло подтверждения при исследовании их тонкой мор-
фологии. Тем не менее, выявлен защитный барьер, который состоит из длинных микроворсинок
хвостового придатка и окружает полностью сформированную метацестоду.
Ключевые слова: метацестода, Kowalewskius, морфология, ультраструктура, промежуточный
хозяин, пиявка, Erpobdella, встречаемость, Верхняя Колыма, Магаданская область.
DOI: 10.1134/S0031184719020017
Метацестоды Kowalewskius parvulus (Kowalewski, 1904) Yamaguti, 1959 (Cyclo-
phyllidea: Hymenolepididae) - известные паразиты глоточных пиявок Erpobdella
octoculata L. (Hirudinea: Erpobdellidae) в Европе, от ее западных до восточных гра-
ниц (Joyeux, 1922; Soliman, 1955; Pike, 1968; Zajíček, Valenta, 1969; Баянов, Куссая,
1972; Гуров, Шапкин, 1972), а также E. punctata (Leidy) в Северной Америке (de Giusti,
Kingston, 1962). Видовую принадлежность метацестод неоднократно подтверждали
экспериментально - при скармливании зараженных пиявок опытным птенцам были
получены зрелые цестоды K. parvulus (Joyeux, 1922; de Giusti, Kingston, 1962; Pike,
1968; Zajíček, Valenta, 1969; Баянов, Куссая, 1972). Более того, в опубликованных тези-
сах доклада американских авторов (de Giusti, Kingston, 1962) приведены краткие све-
дения о сроках развития метацестод в экспериментально зараженных пиявках, авторам
удалось восстановить весь жизненный цикл K. parvulus.
Пайк (Pike, 1968) обнаружил у E. octoculata (из озера Линхейлин, Средний Уэльс, Ан-
глия), кроме K. parvulus, еще один вид метацестод - Haploparaksis (=Wardium) cirrosa
(Krabbe, 1869). Обсуждая форму хоботковых крючьев метацестод, изображенных Пай-
91
ком (Pike, 1968, fig. 2), Бондаренко и Контримавичус (2006) высказали сомнение в их
таксономической принадлежности. Одновременно авторы отметили, что описанные
Пайком метацестоды явно принадлежат одному из представителей сем. Aploparaksidae.
Исследование фауны метацестод хищных глоточных пиявок нами начаты в 2006 г.
после находки в желудке налима Lota lota (L.), пойманного в старице р. Кулу (пра-
вый образующий приток Верхней Колымы), пищевого комка из полупереваренных пи-
явок E. octoculata. Компрессорное исследование этих естественно релаксированных
пиявок позволило, ещё до вскрытия, обнаружить в них многочисленных метацестод
K. parvulus. Одновременно в старице были пойманы единичные живые пиявки E. octo-
culata, одна из которых также была заражена метацестодами K. parvulus. Сборы пи-
явок продолжили в других водоемах бассейна Колымы, в результате в озерах Сейм-
чано-Буюндинской впадины были впервые обнаружены метацестоды близкого вида
K. formosus (Dubinina, 1953) (Регель, 2010). Систематическому положению последнего
вида и восстановлению самостоятельности рода Kowalewskius Yamaguti, 1959 посвя-
щена отдельная публикация (Галкин, Регель, 2012).
Помимо метацестод Kowalewskius spp. в последнем районе у четырёх E. octoculata
обнаружены аплопараксидные метацестоды, по-видимому, близкие «цистицеркоидам
H. cirrosa», описанным Пайком (Pike, 1968) от пиявок Англии. Краткая характери-
стика метацестод, а также обсуждение их систематического положения представле-
ны в недавнем сообщении (Регель, 2016). Предварительно, до получения дополни-
тельных подтверждений, метацестод отнесли к виду Aploparaksis shigini Bondarenko,
Kontrimavichus, 2006, специфичному паразиту речных чайковых птиц.
В настоящей статье обобщены сведения о встречаемости перечисленных выше ви-
дов у пиявок бассейна Верхней Колымы, а также приведено морфологическое описа-
ние метацестод рода Kowalewskius.
МАТЕРИАЛ И МЕТОДИКА
Исследовано более 370 пиявок E. octoculata из 13 водоемов бассейна Колымы (сборы
2006-2017 гг.) - в семи из них (в бассейне Верхней Колымы) обнаружены метацесто-
ды трёх видов (табл. 1). В таблицу не включены отрицательные результаты вскрытий
пиявок E. octoculata (от 1 до 21 экз.) из четырех озер бассейна Верхней Колымы и двух
озер бассейна Нижней Колымы из окрестностей пос. Черского. Кроме того, небольшие
выборки пиявок Erpobdella spp. собраны в конце мая 2014 г. и начале июня 2017 г.
в окрестностях Санкт-Петербурга, в том числе в пруду парка г. Гатчина, где также об-
наружены метацестоды K. parvulus.
Вскрытие пиявок и изучение извлеченных метацестод проводили в 0.6 % растворе
NaCl. При полном вскрытии, удалив головной отдел, производили два латеральных
надреза кожно-мускульной стенки задней части тела пиявок, отделяли брюшную по-
ловину от спинной и с помощью оптики выискивали на дне чашки Петри выпавших
паразитов. Затем проводили осмотр поверхности средней кишки и толщи тела пия-
вок. В заключение делали соскоб мягких тканей (соединительной ткани) со стенок
кожно-мускульного мешка. При обнаружении осевших на дно чашки Петри мета-
цестод, все фрагменты пиявки просматривали компрессорно.
92
Частичное вскрытие пиявок (небольшой латеральный надрез в задней четверти на
расстоянии 1 см от задней присоски) проводили для последующего скармливания за-
раженных пиявок птенцам чаек. Летом 2010-2012 и 2015-2017 гг. сделано несколь-
ко безуспешных попыток экспериментально подтвердить таксономическую принад-
лежность метацестод, предварительно определенных как Aploparaksis shigini. Однако
трижды эти метацестоды были найдены в июне, до вылупления птенцов сизой чайки
(Larus canus L.) - потенциального дефинитивного хозяина (или других чаек), а од-
нажды - в сентябре. Напротив, в июле, при наличии птенцов для эксперимента, мы не
обнаруживали в пиявках аплопараксидных метацестод. В результате в 2010 и 2012 гг.
птенцам сизой чайки, в 2011 г. птенцу моевки (Rissa tridactyla L.), а в 2017 г. птенцу
тихоокеанской чайки (L. schistisagus Steineger) скормили пиявок, зараженных метаце-
стодами K. formosus. Но заражение чаек этим неспецифичным паразитом дало отрица-
тельный результат.
Измерение и фотографии живых объектов и препаратов сделаны с использованием
микроскопов Axio Imager. D1, Amplival, бинокуляра МБС-10 и фотоаппаратов Canon
PowerShot A95, A1100 IS. После прижизненного изучения, часть извлеченных метаце-
стод помещали в поливиниловый спирт или в среду Фора-Берлезе, часть фиксировали
в 70° этаноле, а фрагменты тел зараженных пиявок - в Буэне. Из последних фикса-
ций изготовлены парафиновые срезы, окрашенные гематоксилин-эозином. Фрагменты
соединительной ткани, содержащие живых метацестод K. formosus, а также отдель-
ные метацестоды были зафиксированы в 2 % растворе глутарового альдегида на 0.1М
фосфатном буфере, дофиксированы в 1 % растворе ОsО4 на 0.2М фосфатном буфере,
окрашены в насыщенном растворе уранилацетата и залиты в смесь ЭПОН-Аралдит.
Ультратонкие срезы (около 90 нм) контрастировали цитратом свинца, затем изучали
в трансмиссионных электронных микроскопах JEM-1011 и JEM-1400Plus (JEOL, Япо-
ния).
Измерения в статье приведены в мм, если не указано иначе.
РЕЗУЛЬТАТЫ
Глоточные или малые ложноконские пиявки (Erpobdellidae) в исследованных озерах
Северного Приохотья, по-видимому, чрезвычайно редки. Лишь в одной пробе из озе-
ра низовья р. Гижига обнаружены единичные E. octoculata. Не многочисленны они и
в большинстве исследованных нами водоемов бассейна Колымы, многие из которых
ещё и труднодоступны. Относительно объемные выборки E. octoculata получены лишь
из шести близлежащих озер в окрестности пос. Сеймчан (Среднеканский район Ма-
гаданской области, 500 км от Магадана). Результаты вскрытий представлены в табл. 1.
Обнаруженные нами места обитания пиявок E. octoculata сосредоточены в пой-
мах рек. В настоящее время очаг заражения пиявок метацестодами K. parvulus
(рис. 1) достоверно установлен только в старице среднего течения р. Кулу (Тенькинский
район Магаданской области, 430 км от Магадана). Метацестоды K. formosus (рис. 2е-
2ж; 3) встречены в трех озерах Сеймчано-Буюндинской впадины. В двух водоемах
близ пос. Сеймчан («Лягушачье» и «Карьер») обнаружены метацестоды Kowalewskius
sp. на ранних стадиях сколексогенеза (до начала формирования хоботковых крючьев)
(рис. 2а-2г), что не позволило определить их вид. Здесь следует отметить, что по-
ловозрелые цестоды обоих видов рода Kowalewskius обнаружены в начале сентября
93
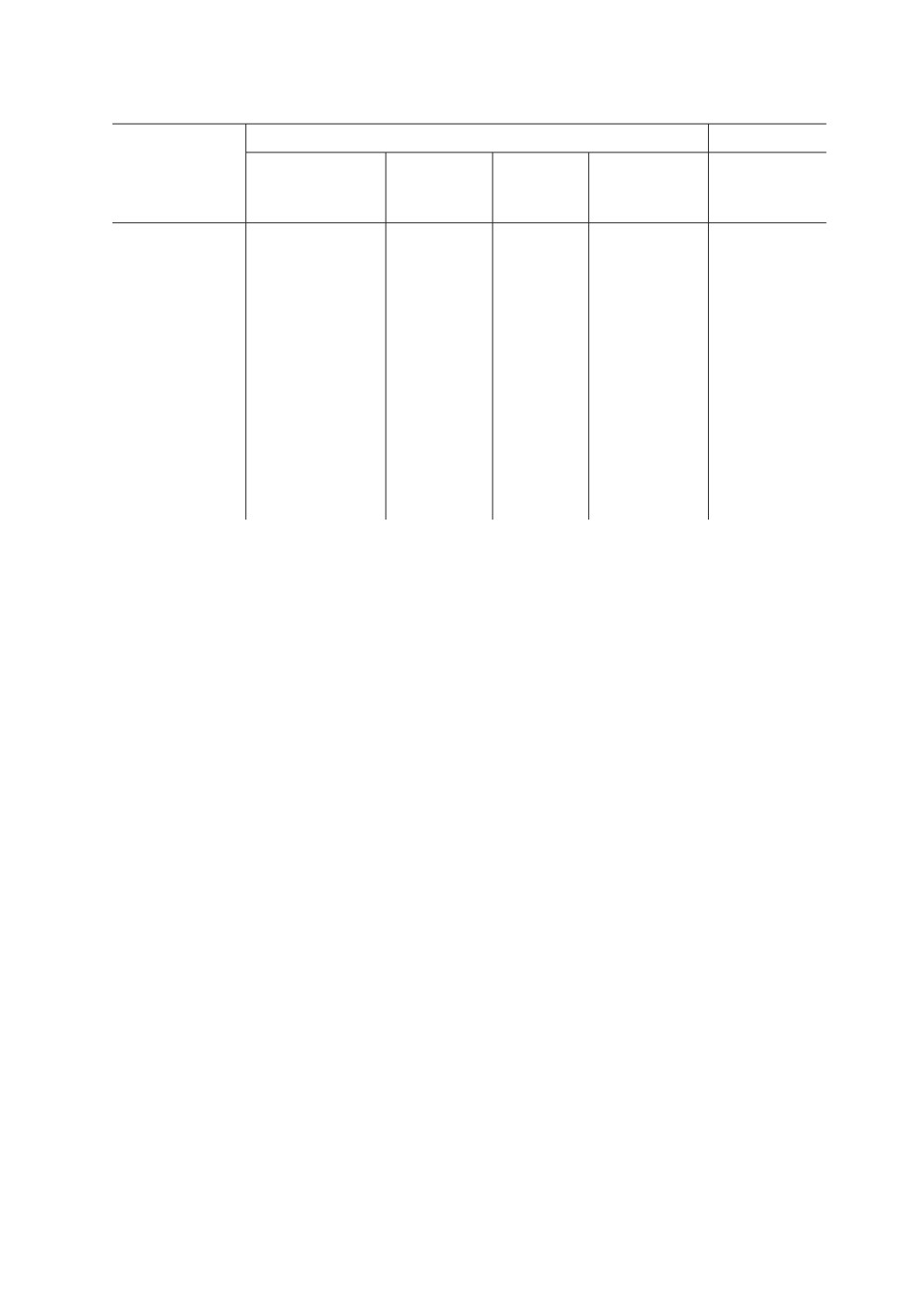
Таблица 1. Встречаемость метацестод Kowalewskius spp. и Aploparaksis shigini у пиявок
Erpobdella octoculata в водоемах бассейна Верхней Колымы
Озера,
Даты
Вскрыто,
Заражено метацестодами, экз. (w ± %) [ИИ]
координаты
сборов
экз.
K. parvulus
K. formosus
A. shigini
«Старица-Кулу»
27.08.2006,
23*
6**
-
-
61°52ʹ N, 147°26ʹ E
24.06.2014
(26.1 ± 9.2)
[40 - 136]
«Формоза»,
09.06.2010-
9
-
2 [50; 78]
-
62°33ʹ N, 153°37ʹ E
13.06.2012
«Тайвань»
10.06.2011
3
-
-
1 [41]
62°33ʹ N, 153°36ʹ E
«Утиное»
19.06.2009-
140
-
4 (2.1 ± 1.2)
2 (1.4 ± 1.0)
62°49ʹ N, 152°24ʹ E
26.06.2017
[61 - 100]
[18; 20]
«Длинное»
17.09.2011,
22
-
-
1 (4.5 ± 4.4)
62°48ʹ N, 152°25ʹ E
01.07.2015
[18]
«Лягушачье»
13.06.2010,
83
-
5 (6.0 ± 2.6)
-
62°50ʹ N, 152°24ʹ E
15.06.2011,
[23 - 100]
26.06.2017
1*** (1.5 ± 1.5) [c. 50]
«Карьер»
08.06.2016
29
1*** (3.4 ± 3.4) [более 14]
-
62°54ʹ N, 152°24ʹ E
П р и м е ч а н и е : * В том числе 15 полупереваренных пиявок из желудка налима и 3 живые пиявки от
27.08.2006 и 5 живых пиявок от 24.06.2014.
** Заражены 4 пиявки из желудка налима, и по одной живой из сборов 27.08.2006 и 24.06.2014.
*** Заражены метацестодами Kowalewskius sp. на стадии раннего сколексогенеза.
2009 г. у хлопунца хохлатой чернети (Aythya fuligula L.), добытого на одном из
«сеймчанских» озер (оз. «Копыто», 62°51ʹ N, 152°24ʹ E - с отрицательным результатом
вскрытий пиявок). Тогда же, на берегу озера Утиное был найден погибший птенец хох-
латой чернети, из кишечника которого извлечены зрелые цестоды и многочисленные
пакеты яиц K. formosus (рис. 2д), размер которых составил в среднем 0.396 ± 0.047 ×
0.454 ± 0.059, т.е. был несколько меньше значения 0.601 × 0.470, приведённого для
K. parvulus (Pike, 1968). Находки зрелых цестод у нелетных хлопунцов позволяют счи-
тать группу «сеймчанских» озер потенциальным очагом заражения обоими видами
рода Kowalewskius.
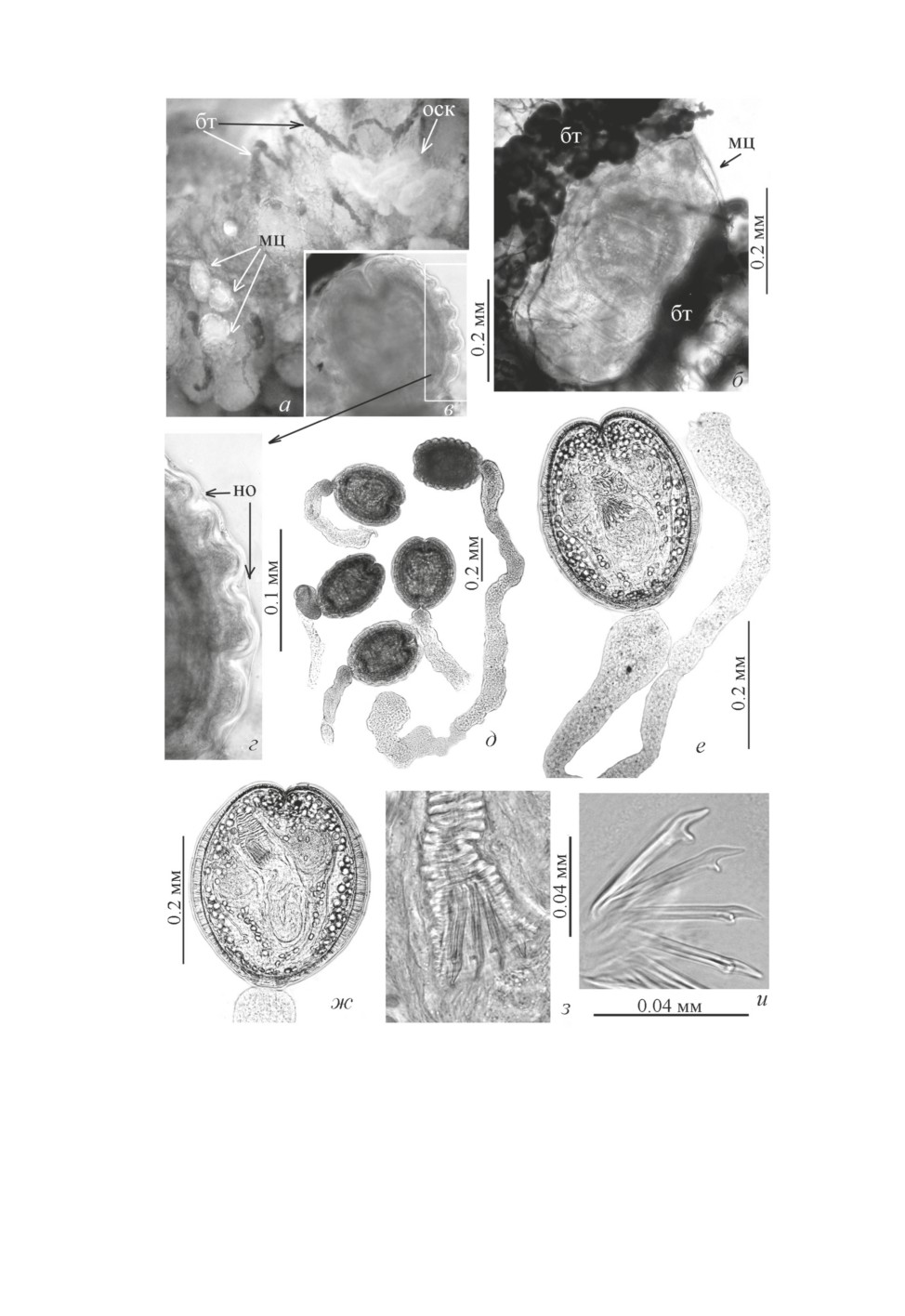
Метацестоды Kowalewskius spp. локализуются в задней трети тела пиявок вокруг
кишки в толще соединительной (ботриоидной) ткани (рис. 1б, 1в, 1д; 3а-3в). Полно-
стью сформированные (живые) метацестоды выглядят опалесцирующими овальны-
ми тельцами молочно-бирюзового цвета (рис. 3а). Они прочно «спаяны» с окружа-
ющими их элементами соединительной ткани пиявки. Прижизненное компрессорное
исследование метацестод позволило предположить, что они снабжены дополнитель-
ной защитой оболочкой - тонкостенной наружной цистой (рис. 3г, стрелки). На ги-
стологическом срезе (рис. 1д) видны две свободные от соединительной ткани хозяина
«полости» - ниши, занятые цистами и оплетающими их хвостовыми придатками, при-
чём эти полости имеют чётко выраженные границы. Очевидно, морфогенез метацестод
94
Таблица 2. Размеры метацестод Kowalewskius parvulus и K. formosus
K. parvulus
K. formosus
Баянов,
Показатель
Joyeux, 1922
Pike, 19681
Куссая,
Наши данные
Наши данные
1972
Длина цисты
0.220*
0.278-0.313
0.260-0.340
0.256-0.300
0.281-0.371
(0.292)
(0.274 ± 0.01)
(0.323 ± 0.011)
Ширина цисты
0.198*
0.244-0.278
0.200-0.300
0.192-0.255
0.211-0.299
(0.264)
(0.232 ± 0.017)
(0.263 ± 0.013)
Длина / диаметр
– / 0.020-0.070*
0.348-1.175 /
1.120 / -
До 1.250 /
До 1.903 /
хвостового
0.010-0.124*
0.020-0.170
0.020-0.150
придатка
(0.638 / -)
Толщина стенки
0.012-
0.017-0.031
0.020
0.010-0.031
0.020-0.044
цисты
-0.015*
(0.025)
(0.017)
Длина крючьев
0.038-
0.038-0.039
0.039-0.044
0.036-0.040
0.037-0.042
хоботка
-0.039
Длина лезвия
-
-
0.008-0.009
0.008-0.009
0.007-0.008
П р и м е ч а н и е : *Помечены размеры, внесенные нами, согласно масштабу на рисунках авторов
(Joyeux, 1922, fig. 1; Pike, 1968, fig. 4).
1 В таблицу включены лимиты размеров живых метацестод и (в скобках) среднее значение, хотя автор
(Pike, 1968) в своей работе привёл также параметры фиксированного материала.
сопровождается формированием некоего защитного барьера - аналога неклеточной на-
ружной цисты. Препарирование и извлечение метацестод из тканей пиявки приводит
к разрушению не только этой наружной оболочки (барьера), но часто и хвостового
придатка, оплетающего цисту. Исключительно редко хвостовой придаток сохранял це-
лостность (рис. 3д).
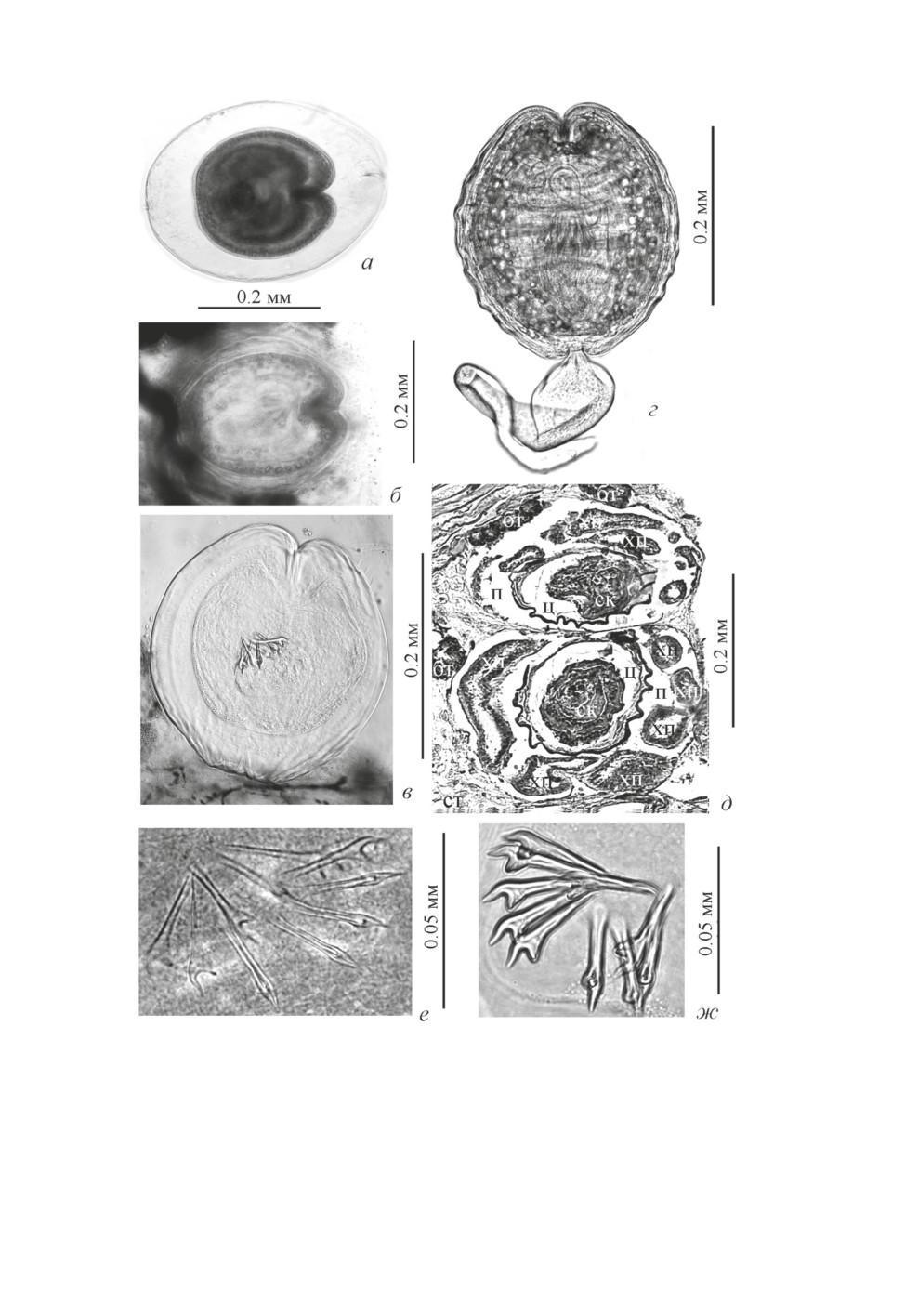
Метацестоды K. parvulus, извлечённые из первой полупереваренной пиявки (из же-
лудка налима), были полностью лишены хвостовых придатков (рис. 1а). У большин-
ства особей произошло частичное или полное отслоение наружного слоя стенки цисты
(вероятно, гликокаликса или «гиалинового» слоя, в терминах световой микроскопии).
Остальные слои стенки цисты (тегумент, наружный фиброзный, клеточный, внутрен-
ний фиброзный) плотно облегали проспективное тело.
Помимо завершивших морфогенез метацестод K. parvulus в одной пиявке из же-
лудка налима (после просветления её тела в поливиниле) выявлены три особи на ста-
дии позднего сколексогенеза с развивающимися хоботковыми крючьями (рис. 1е).
Полностью сформированные метацестоды K. parvulus из живых пиявок (рис. 1б- 1г)
имели характерные для вида крючья хоботка с широким корневым отростком 0.0045-
0.005 × 0.004-0.005 и уплощенным концом рукоятки (рис. 1ж). Размеры метацестод
Kowalewskius spp. по литературным и нашим данным сведены в табл. 2.
Метацестоды Kowalewskius sp. на стадии раннего сколексогенеза встречены дважды,
здесь приведем их описание по первой находке (из пиявки, пойманной в оз. «Лягу-
шачье» 13.06.2010 и вскрытой 20.06.2010). Общая длина метацестод, очевидно, одно-
го возраста (без учета хвостового придатка) варьирует от 0.266 до 0.843 (рис. 2а-2г).
95
Рис. 1. Метацестоды Kowalewskius parvulus.
а - из полупереваренной пиявки, б - в ткани пиявки, в - в поливиниле, г - извлечённая из тканей пиявки,
д - гистологический срез тела пиявки с двумя метацестодами, е - не завершившие развитие крючья
на стадии позднего сколексогенеза, ж - сформированные хоботковые крючья.
Обозначения: бт - ботриоидная ткань, п - полость с метацестодой, ск - сколекс, ст - соединительная ткань,
хп - хвостовой придаток, ц - циста.
96
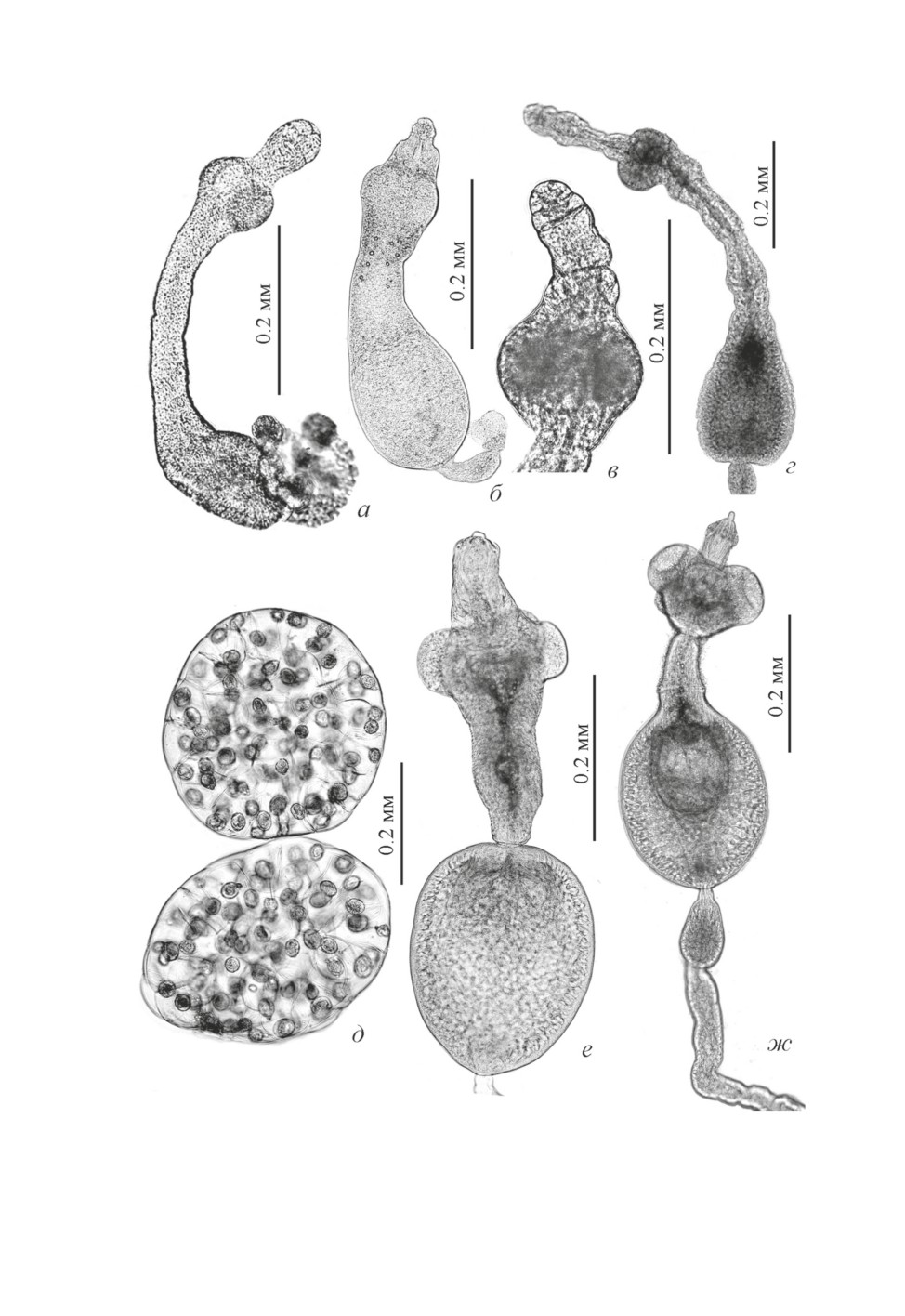
Рис. 2. Метацестоды Kowalewskius sp. на стадии раннего сколексогенеза (а - г); пакеты яиц (д)
и метацестоды K. formosus на стадии позднего сколексогенеза и перед инцистированием (е-ж).
97
Размер будущей цисты у самой мелкой особи 0.130 × 0.082, крупной - 0.280 × 0.205.
Длина проспективного отдела также варьирует в больших пределах (0.140-0.564). Ди-
аметр шейки 0.043-0.060, в ней заметны первые известковые тельца. Ширина сколекса
0.092-0.127, диаметр присосок 0.046-0.075. Хоботок нередко превышает длину ско-
лекса, его диаметр в средней части 0.040-0.055, в апикальной части уже намечена бо-
розда, зона формирования зачатков хоботковых крючьев (рис. 2в).
Единичные метацестоды K. formosus на стадии позднего сколексогенеза и инваги-
нации (рис. 2е, 2ж) были обнаружены (в пиявке, пойманной в оз. «Формоза» 9.06.
2010, вскрытой 15.06.2010) среди многочисленных особей, недавно завершивших ин-
цистирование (рис. 3е, 3ж). Параметры метацестод на стадии позднего сколексоге-
неза (рис. 2е) и в процессе инвагинации (рис. 2ж), соответственно: длина (без учета
хвостового придатка) 0.556 и 0.551, размер цисты 0.279 × 0.213 и 0.286 × 0.218, дли-
на проспективной части 0.377 и 0.265, диаметр шейки 0.047-0.084, сколекса - 0.170
и 0.176, присосок - 0.070-0.075 и 0.078-0.081, хоботка - 0.056-0.075 и 0.039-0.047.
Длина поврежденного хвостового придатка превышает 0.5, в проксимальной части его
ширина 0.064-0.065, в самой узкой - 0.028. Метацестоды из этой же пиявки, по-види-
мому, недавно завершившие инцистирование (рис. 3е, 3ж) отличались прозрачностью
и гладкими стенками цисты. По мере «дозревания» и сокращения мышечных воло-
кон стенки цисты, ее поверхность становится ребристой (рис. 3в, 3г). Стенка цисты
образована пятью хорошо выраженными слоями общей толщиной от 0.020 до 0.044.
Толщина пристеночной части шейки 0.008-0.022, ширина сколекса 0.138-0.185, хо-
ботка 0.030-0.072, хоботкового влагалища 0.070-0.085, диаметр присосок 0.070-0.080.
Длина хоботковых крючьев (см. табл. 2) варьирует в больших пределах, чем отмечено
у взрослых цестод. По форме крючья K. formosus (рис. 3з-3и) более стройные (изящ-
ные), чем у K. parvulus (рис. 1е). При сходной длине крючка и лезвия они отличаются
меньшим размером корневого отростка (0.003 × 0.0025-0.003).
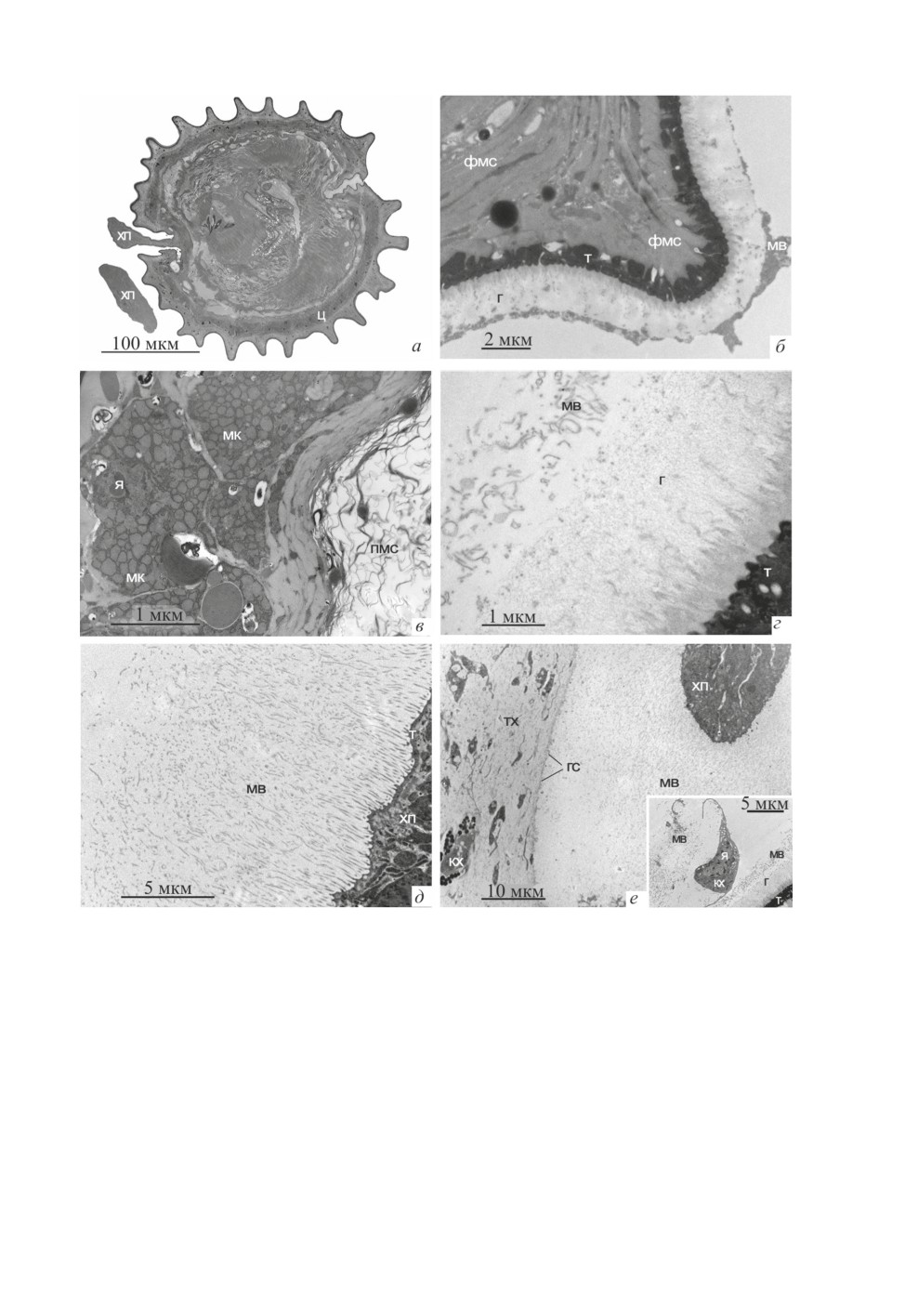
Электронно-микроскопическое изучение метацестод K. formosus в ткани хозяина и
свободных метацестод позволило более подробно ознакомиться с их морфологией и
паразито-хозяинными отношениями на клеточном уровне. Ультраструктура стенки ци-
сты K. formosus (рис. 4а-4в) демонстрирует следующие особенности: поверхностная
цитоплазма тегумента цисты толщиной 1-1.5 мкм заполнена отложениями плотного
материала. В области заднего полюса наблюдается постепенная замена плотной по-
верхностной цитоплазмы на более рыхлую, несущую микроворсинки, цитоплазму, ко-
торая покрывает отходящий от заднего полюса хвостовой придаток.
Под базальной пластинкой тегумента цисты располагаются фиброзные слои (наруж-
ный - кольцевой и внутренний - продольный), в которых заключены мышечные во-
локна той же ориентации. Характерные выступы (рёбра) наружной поверхности цисты
сформированной метацестоды (рис. 4а, 4б) образованы именно этими фиброзно-мы-
шечными слоями, тогда как подлежащие слои имеют примерно равную толщину, су-
жаясь только возле полюсов цисты. В клеточном слое наиболее заметны мышечные
клетки, у которых основная часть цитоплазмы заполнена расширенными профилями
гранулярного эндоплазматического ретикулюма (ГЭР), а также цитоны тегумента.
Глубже располагаются экскреторные каналы и известковые тельца, более многочислен-
ные в области полюсов. Экскреторные каналы у заднего полюса впадают в экскретор-
ный атриум. Хорошо развитый т.н. псевдомиелиновый слой, образованный отростками
98
Рис. 3. Полностью сформированные метацестоды Kowalewskius formosus.
а - на брюшной стенке тела пиявки; б - в ткани пиявки; в, г - ребристая стенка цисты; д - извлеченные
из ткани пиявки; е, ж - очевидно, недавно инцистированные; з -хоботковые крючья живой метацестоды;
и - они же в поливиниле.
Обозначения: бт - ботриоидная ткань (стрелки), мц - метацестоды (стрелки), но - наружная оболочка
(стрелки), оск - остаток стенки кишки (стрелка).
99
Рис. 4. Тонкая морфология метацестоды K. formosus.
а - продольный срез цисты и фрагмента хвостового придатка, б - выступ наружной поверхности цисты,
в - клеточный и псевдомиелиновый слои стенки цисты; г - гликокаликс цисты, д - микроворсинки
хвостового придатка, е - фрагмент полости с метацестодой в тканях хозяина, на врезке - клетка
хозяина возле цисты.
Обозначения: г - гликокаликс, гс - гранулярный слой, кх - клетка хозяина, мв - микроворсинки хвостового
придатка, мк - мышечная клетка, пмс - псевдомиелиновый слой цисты, т - тегумент цисты и хвостового
придатка, тх - ткань хозяина, фмс - фиброзно-мышечный слой цисты, хп - хвостовой придаток, я - ядро.
100
клеток, ограничивает внутреннюю поверхность стенки цисты (рис. 4в) и отделяет её
от пристеночной части инвагинированной шейки, выстилающей внутреннюю полость,
занятую сколексом с присосками и втянутым хоботком. Шейка и сколекс покрыты те-
гументом с микротрихиями; тегумент присосок несёт конусовидные микротрихии, бо-
лее крупные по сравнению с таковыми остальной части тегумента сколекса.
Гликокаликс тегумента цисты имеет толщину около 3 мкм и характерную сетчатую
структуру, т.е. состоит из переплетённых фибрилл, более плотных у поверхности тегу-
мента (рис. 4г). Наружный слой гликокаликса менее плотен; по его наружной границе
нередки везикулы и фрагменты микроворсинок хвостового придатка. Последний об-
разован цитонами тегумента и мышечными клетками; их сократительная часть рас-
полагается под дистальной цитоплазмой в виде коротких тяжей небольшого сечения.
У заднего полюса метацестоды микроворсинки хвостового придатка имеют неболь-
шую длину, на некотором удалении она превышает 10 мкм (рис. 4д).
При анализе срезов ботриоидной ткани хозяина, содержащей цистицеркоиды
K. formosus, обращает на себя внимание следующее обстоятельство: в полостях с ме-
тацестодами почти не отмечено участков, свободных от микроворсинок. Создаётся
впечатление, что именно микроворсинки определяют размеры зоны, в которой лежит
циста с хвостовым придатком. Граница этой зоны и тканей хозяина на срезах имеет
вид тонкого (0.2-0.3 мкм) слоя гранулярного материала и везикул (рис. 4е), оконту-
ривающего рыхлую соединительную ткань хозяина, в которой располагаются имму-
нокомпетентные и ботриоидные клетки пиявки. В редких случаях, при повреждении
этого барьера, регистрируется проникновение клеток хозяина в полость, занятую мета-
цестодой, но они всегда задерживаются у наружной поверхности гликокаликса цисты
(рис. 4е, врезка). При этом наблюдаются адгезия микроворсинок на поверхности кле-
ток хозяина, а также фрагменты микроворсинок в фагоцитарных вакуолях клеток хо-
зяина.
ОБСУЖДЕНИЕ
Большинство исследователей паразитов пиявок признавали их промежуточными хо-
зяевами метацестод K. parvulus (Joyeux, 1922; Soliman, 1955; de Giusti, Kingston, 1962;
Pike, 1968; Баянов, Куссая, 1972; Гуров, Шапкин, 1972). В отличие от перечисленных
авторов, чешские гельминтологи (Zajíček, Valenta, 1969) отводят пиявкам в жизненном
цикле K. parvulus иную роль. Авторы нашли в желудках двадцати восьми (из 542) пи-
явок E. octoculata по 3-6 метацестод, лишенных хвостового придатка и лежащих сво-
бодно в «мембранных цистах, либо без них». Не получив положительного результата
при экспериментальном заражении пиявок пакетами яиц K. parvulus, они пришли к вы-
воду о «резервуарном» статусе этого хозяина. Баянов и Куссая (1972) критикуют чеш-
ских авторов, отмечая обнаружение метацестод не в кишке, а в «паренхиме» пиявок по
всему телу. Мы уже высказывали предположение, что находка метацестод в кишечнике
пиявки возможна при каннибализме (Регель, 2010) (однако трудно представить, что все
28 пиявок заразились таким путем). Здесь следует отметить внешнее сходство метаце-
стод K. parvulus на рисунке чешских авторов (Zajíček, Valenta, 1969, fig. 2) и на нашей
фотографии (рис. 1 а), где представлена ларвоциста (из полупереваренных в желудке
налима пиявок) с отделившимся наружным слоем стенки цисты. Возможно, чешские
авторы вскрывали пиявок после длительной анестезии (или уже мертвых). У таких
101
червей целостность тканей нарушается, так что при любом нажатии на тело пиявки
часть паразитов может переместиться в просвет кишки (желудка).
Расположение хвостового придатка, плотно окружающего цисту (эндоцисту), соз-
дает впечатление, что развитие личинки проходит под защитой тонкой неклеточной
«экзоцисты», то есть метацестоды напоминают по форме морфологический тип цисти-
церкоида - циклоцерк (Котельников, 1971). Постэмбриональное развитие последних
метацестод проходит под защитой неклеточной наружной оболочки в полости тела ра-
кообразных. Данная модификация цистицеркоида характерна для представителей рода
Fimbriaria и типичного подрода Microsomacanthus. Линейный рост цистицеркоида на
стадии метамеры и сколексогенеза, скованный сферической наружной цистой, сопря-
жен с цикличной укладкой проспективного отдела (сколекса и шейки), эндоцисты и
хвостового придатка. После инвагинации шейки и втягивания сколекса внутренняя
циста остается оплетенной хвостовым придатком (размер которого зависит от интен-
сивности инвазии промежуточного хозяина - максимальной длины хвостовой при-
даток достигает при низкой интенсивности инвазии) (Регель, 1986). При аккуратном
вскрытии рачков в физиологическом растворе циклоцерки (или циклоцистицеркоиды)
сохраняют целостность неклеточной наружной оболочки (экзоцисты).
Однако изучение тонкой морфологии метацестод K. formosus не выявило существо-
вания у них такой «экзоцисты», хотя и позволило установить существование тонкого
гранулярного слоя, ограничивающего соединительную ткань хозяина (Поспехова, Ре-
гель, 2013). Происхождение гранулярного слоя при изучении спонтанно заражённых
пиявок остаётся неясным. Предположим, метацестоды рода Kowalewskius близки мор-
фологической модификации циклоцерк, что вполне вероятно, учитывая характерное
расположение хвостового придатка, обвивающего эндоцисту. Тогда уже в процессе
проникновения через стенку кишечника хозяина они должны сформировать тонкую
неклеточную оболочку, под защитой которой будет происходить морфогенез метаце-
стоды хотя бы на ранних стадиях - до формирования эндоцисты и хвостового придатка
(Краснощеков, Томиловская, 1978; Краснощеков, 1980). В дальнейшем, когда защит-
ные функции берут на себя длинные микроворсинки хвостового придатка, наружная
неклеточная оболочка либо исчезает, либо становится плохо различимой, прилегая
к тканям хозяина. С целью подтверждения либо опровержения принадлежности мета-
цестод Kowalewskius к модификации циклоцерк необходимо либо экспериментальное
заражение, либо новая находка пиявок с метацестодами на ранних стадиях лярвогенеза
для последующего электронно-микроскопического изучения.
Клеточные реакции организма на чужеродные объекты среди Annelida были изуче-
ны, в основном, на примере олигохет, тогда как пиявки стали объектами наблюдения
относительно недавно (De Eguileor et al., 2000), причём в качестве объекта использова-
лись медицинские пиявки, имеющие другой тип питания. Из наблюдений этих авторов
следует, что паразиты в организме медицинской пиявки инкапсулируются путём отло-
жения на их поверхности меланина. В случае E. octoculata отложения тёмного матери-
ала (возможно, меланина) изредка отмечали на поверхности метацеркарий, тогда как
поверхность метацестод была от них свободна.
Итак, география встречаемости метацестод у глоточных пиявок рода Erpobdella рас-
ширена и ныне включает бассейн верхнего течения реки Колыма на северо-востоке
Азии. Очевидно, при наличии интереса к подобным исследованиям, очаги инвазии
102
(и, возможно, иные виды метацестод) будут обнаружены у хищных пиявок и в других
регионах Евразии (например, в Западной Сибири, типовом месте нахождения взрос-
лых цестод K. formosus). Одновременно надеемся, будет решён и вопрос, какой барьер
защищает ранние стадии развития метацестод Kowalewskius spp. (до дифференциации
хвостового придатка) от реакций тканей хозяина.
БЛАГОДАРНОСТИ
Авторы выражают искреннюю благодарность сотруднице института Кусенко К.В. за
помощь в изготовлении гистологических препаратов, а также рецензентам статьи за
конструктивный и доброжелательный отзыв.
СПИСОК ЛИТЕРАТУРЫ
Баянов М.Г., Куссая Н.А. 1972. Пиявки в Башкирии как промежуточные хозяева гельминтов птиц. Труды
Башкирского сельскохозяйственного института. Уфа, т. 17, с. 33-45.
Бондаренко С.К., Контримавичус В.Л. 2006. Аплопараксиды диких и домашних птиц. Основы цестодологии,
т. 14. М., Наука, 443 с.
Галкин А.К., Регель К.В. 2012. О систематическом положении Hymenolepis formosa Dubinina, 1953 и
о валидности рода Kowalewskius Yamaguti, 1959. В кн.: Проблемы цестодологии, вып. 4, СПб, «ЭЛ-
МОР», с. 35-47.
Гуров В.Н., Шапкин В.А. 1972. О спонтанном заражении пиявок личинками гельминтов в Башкирии. Ма-
териалы научных исследований членов Всесоюзного общества гельминтологов, 1970-1971 гг., М., т.
24, с. 45-46.
Котельников Г.А. 1971. Типология личиночных форм у цестод семейства гименолепидид. Материалы науч-
ной конференции Всесоюзного общества гельминтологов, М., т. 22, с. 116-126.
Краснощеков Г. П. 1980. Церкомер - личиночный орган цестод. Журнал общей биологии 41 (4): 615-627.
Краснощеков Г. П., Томиловская Н. С. 1978. Морфология и развитие цистицеркоидов Paricterotaenia porosa
(Cestoda: Dilepididae). Паразитология 12 (2): 108-115.
Поспехова Н.А., Регель К.В. 2013. Ультраструктура метацестоды Kowalewskius formosus (Dubinina, 1953)
Regel, 2010 (Hymenolepididae) в промежуточном хозяине. Чтения памяти академика К.В. Симакова:
Материалы докладов Всероссийской научной конференции. Магадан, СВНЦ ДВО РАН, с. 160-162.
Регель К.В. 1986. Развитие лярвоцист типа циклоцерк. Паразитология 20 (3): 188-194.
Регель К.В. 2010. Пиявки Erpobdella octoculata L. - промежуточные хозяева метацестод Kowalewskius
parvula (Kowalewski, 1904) и Kowalewskius formosa (Dubinina, 1953) comb. nov. в бассейне Колымы.
В кн.: Паразиты Голарктики. Материалы Международного симпозиума. Петрозаводск, Институт био-
логии Карельского научного центра РАН, т. 2, с. 70-73.
Регель К.В. 2016. О таксономическом положении аплопараксидных метацестод, обнаруженных у пиявок
Erpobdella octoculata в бассейне Верхней Колымы. Труды Центра паразитологии. T. XLIX: материалы
Международной научной конференции «Фауна и экология паразитов», М., с. 121-123.
De Eguileor M., Grimaldi A., Tettamanti G., Valvassori R., Lanzavecchia G. 2000. Different types of response to
De Giusti D. L., Kingston N. 1962. A preliminary account of the life cycle of Kowalewskius parvula (Kowalewski,
1904; Yamaguti, 1959). Cestoda: Hymenolepididae. American Zoologist 2 (4): 517. (Abstr.)
Joyeux C. 1922. Recherches sur les ténias des Ansériformes. Développement larvaire d’Hymenolepis parvulus Kow.
chez Herpobdella octoculata (L.) (Hirudinée). Bulletin de la Société de pathologie exotique 15 (1): 46-51.
Soliman K.N. 1955. Observations on some Helminth Parasites from Ducks in Southern England. Journal of
Helminthology 29 (1/2): 17-26.
Pike A.W. 1968. Notes on some Cysticercoids from Pulmonate Molluscs and Leeches in British Freshwaters. Journal
of Helminthology 42 (1/2): 131-138.
Zajíček D., Valenta Z. 1969. Erpobdella octoculata L. (Hirudinea), the reservoir host of Microsomacanthus parvulus
(Kowalewski, 1904) in Chechoslovakia. Věstník Československé společnosti zoologické (and Acta Societatis
Zoologicae Bohemoslovacae) 33 (3): 272-277.
103
ON THE MORPHOLOGY OF METACESTODES OF THE GENUS
KOWALEWSKIUS - PARASITES OF THE LEECHES ERPOBDELLA OCTOCULATA
FROM THE KOLYMA RIVER BASIN
K. V. Regel, N. A. Pospekhova
Key words: metacestode, Kowalewskius, morphology, ultrastructure, intermediate host,
leech, Erpobdella, occurrence, Upper Kolyma, Magadan region.
SUMMARY
The original data on the occurrence of three metacestode species in the leeches Erpobdella octo-
culata L. from the lakes of the Upper Kolyma River are summarized. Morphological description of
two metacestodes of the genus Kowalewskius Yamaguti, 1959, localized in the connective tissue of the
host, is given.
The previously suggested assumption on the similarity of the metacestodes of Kowalewskius spp. and
the modification of the cyсlocercus (or cycloсysticercoid) was not confirmed by the study of their fine
morphology. Nevertheless, a protective barrier was discovered, consisting of long microvilli of the tail
appendage surrounding the fully formed metacestode.
104