ПАРАЗИТОЛОГИЯ, 53, 2, 2019
УДК 595.775
СПОСОБНОСТЬ БЛОХ CITELLOPHILUS TESQUORUM
И FRONTOPSYLLA LUCULENTA LUCULENTA
(SIPHONAPTERA, CERATOPHYLLIDAE) К ПЕРЕЖИВАНИЮ
ОТРИЦАТЕЛЬНЫХ ТЕМПЕРАТУР В ЗИМНИЙ ПЕРИОД
© 2019 г. Е. Г. Токмакова*, Л. П. Базанова, Е. А. Вершинин
Иркутский научно-исследовательский противочумный институт Роспотребнадзора,
ул. Трилиссера, 78, Иркутск, 664047 Россия
* е-mail: flea98@mail.ru
Поступила 17.10.2018 г.
После доработки 14.11.2018 г.
Принята к публикации 14.11.2018 г.
Проведено сравнительное изучение выживаемости блох Citellophilus tesquorum и Frontopsylla
luculenta из пяти природных популяций Сибири при отрицательных температурах в зимний пе-
риод. Выявлены межпопуляционные и межполовые различия внутри каждого из этих видов,
а также различия между популяциями двух видов, связанных с одной и той же популяцией хо-
зяина. Абсолютные значения отрицательных температур в пределах климатической нормы не
являлись критическим фактором для выживаемости блох в течение февраля, когда проводили
наблюдения.
Ключевые слова: блохи, Citellophilus tesquorum, Frontopsylla luculenta, зимовка, холодостой-
кость.
DOI: 10.1134/S0031184719020030
Блохи Citellophilus tesquorum и Frontopsylla luculenta в Сибири являются массо-
выми видами. Citellophilus tesquorum - специфический паразит сусликов, а F. lucu-
lenta - эвризоидный вид, встречается на грызунах многих видов, даурской пищухе
(Иофф, Скалон, 1954; Васильев, 1966; Жовтый, Вершинин, Ромашева, 1984; Жовтый,
Плеснивцева, 1986), в гнездах птиц (Дубинин, 1949), отмечена на человеке и домашних
животных, в частности на собаке (Федорова, 1953). С юга ареал этих видов ограничен
среднегодовой изотермой +2 °С, которая в азиатской части проходит за пределами на-
шей страны (Kiefer et al., 2006). Распространение в северном направлении привязано
к ареалу длиннохвостого суслика, крайние находки зарегистрированы в долинах рек
Яны и Индигирки (Жовтый, Плеснивцева, 1986). Протяжённая территория обитания
характеризуется значительным разнообразием климатических условий, которые на-
прямую (общее понижение температуры) или опосредованно (длительность спячки
хозяев) определяют холодостойкость блох.
120
Температурный режим жизни блох на млекопитающих Сибири (Жовтый, Васильев,
1962; Олькова, 1972) и в их убежищах изучены достаточно подробно (Пауллер, 1959;
Пауллер, Чипизубова, 1961; Жовтый, 1967; Васильев, Жовтый, 1974), что позволяет
косвенно судить об отношении этих насекомых к температуре. Специальными иссле-
дованиями установлена неспособность южной крысиной блохи Xenopsylla cheopis
к выживанию в условиях заполярной зимы (Аксёненко, Сперанская, 1966), показаны
устойчивость блох C. tesquorum и Ctenophthalmus breviatus к отрицательной темпе-
ратуре до -28.5 °С (Тифлов, Фаворисова, 1940) и разная реакция на такую темпера-
туру у представителей различных подсемейств блох (Гусева, Косминский, 1978).
В двух последних упомянутых работах блохи, использованные в опытах, происходили
не из сибирских популяций, а холодостойкость рассматривалась как видовая харак-
теристика, что, конечно, необходимо уточнять, учитывая масштабы распространения
многих видов. С практической точки зрения, способность блох сохранять имагиналь-
ную «прослойку» зимой - одна из составляющих их эпизоотологического значения.
Цель нашего исследования - сравнительное изучение выживаемости блох C. tesquorum
и F. luculenta из разных популяций Сибири при отрицательных температурах в зимний
период.
МАТЕРИАЛ И МЕТОДИКА
Опыты проводили в течение пяти последовательных лет (2007-2011 гг.) с конца ян-
варя по начало марта. В качестве объекта исследования взяты блохи, происходящие
из пяти природных популяций: C. tesquorum altaicus из Монгун-Тайгинского рай-
она Республики Тыва (650 особей), C. tesquorum sungaris и F. luculenta luculenta из
Борзинского района Забайкальского края (703 и 200 особей), C. tesquorum altaicus и
F. luculenta luculenta из Ольхонского района Иркутской области (150 и 250 особей).
Необходимо сказать, что популяционная дифференциация блох, которая основывается
на таковой их хозяев, в этих местах изучена в разной степени. В Забайкалье такие ис-
следования не проводились. Длиннохвостые суслики Ольхонского района объединены
в Приморско-Байкальский класс популяций, более подробного описания структуры их
населения здесь не приводится (Холин, 2013). В Юго-Западной Туве выделено восемь
популяций длиннохвостого суслика и соответствующих им популяций C. tesquorum
(Вержуцкий, 2012). В наших опытах блохи были добыты в Каргинской популяции, мы
условно обозначим их как «тувинские». Популяционный, половой состав и количество
использованных эктопаразитов в конкретный год ограничивались их наличием в ин-
сектарии на момент опыта. Блох помещали в пробирки группами по 50 или 100 особей.
Соотношение самок и самцов в пробирках составляло: C. tesquorum 1 : 1 либо 3 : 2,
F. luculenta 3 : 2. Пробирки заворачивали в вату и помещали в металлический пенал,
который закапывали в снег на глубину около 50 см. В 2009 г. блох распределили по-
ровну в два пенала, один из которых поместили в морозильную камеру бытового холо-
дильника (1 опыт), а другой - поместили в снег (2 опыт). Температура в морозильной
камере поддерживалась в пределах от -1 °С до -3 °С. Температуру в снегу измеряли
ежедневно в утренние часы на глубине 40 см. За 4 недели опыта она колебалась от
-10 °С до -15 °С. По окончании опытов, через четыре недели, блох в пробирках отогре-
вали при комнатной температуре в течение 2 ч, после чего учитывали живых и мерт-
вых насекомых. В 2007-2008 гг. насекомых из половины пробирок извлекали через две
121
недели для промежуточного учёта. Статистическая обработка результатов проведена
общепринятыми методами (Рокицкий, 1973).
РЕЗУЛЬТАТЫ
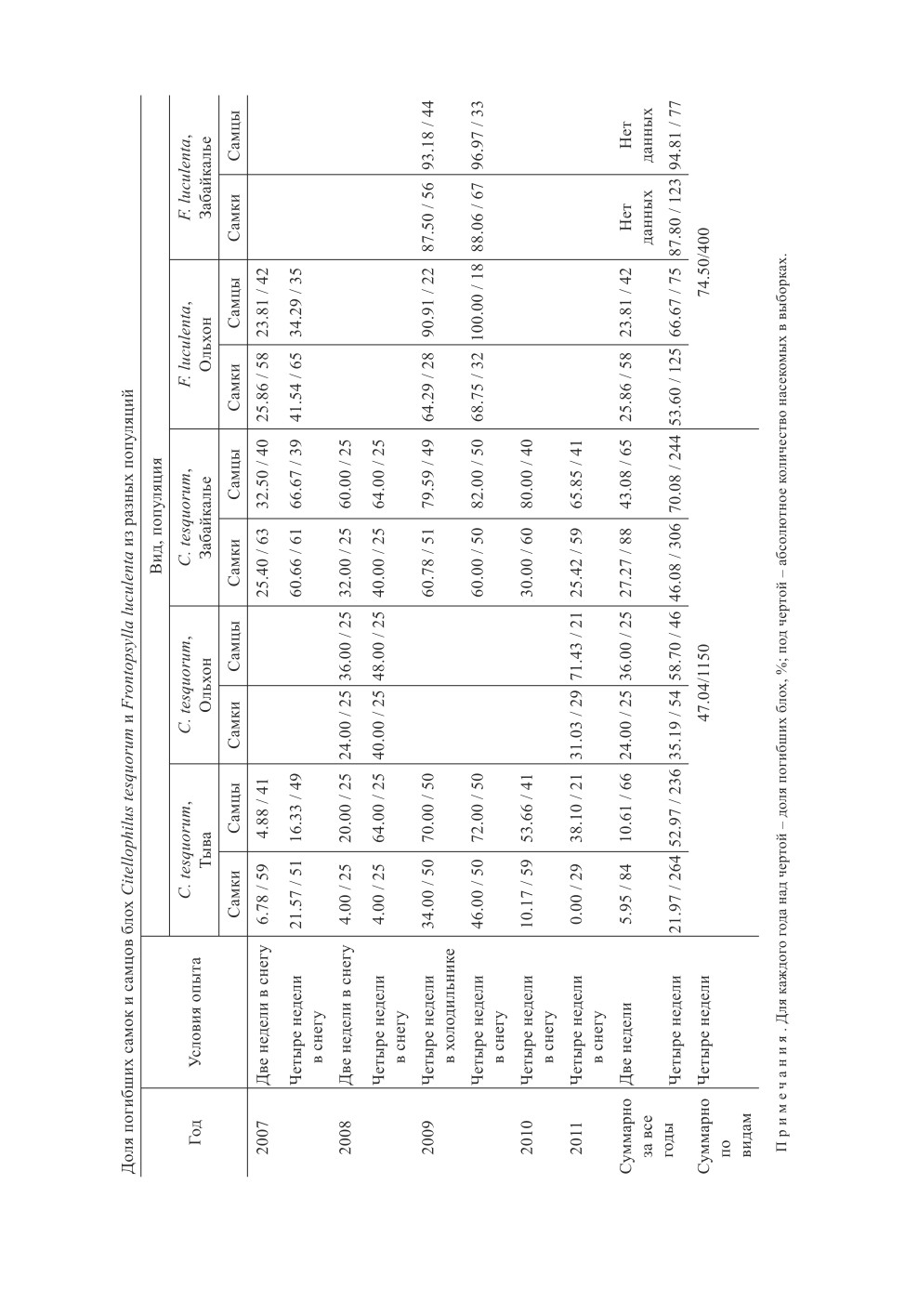
Результаты всех опытов приведены в таблице.
Устойчивость к зимним температурам блох из разных популяций
Самым большим массивом данных мы располагаем по C. tesquorum. Через две
недели в 2007 г. погибло 6.8 % самок и 4.9 % самцов из Тувы, и 25.4 % самок и
32.5 % самцов из Забайкалья (для самок t = 2.78, P < 0.01; для самцов t = 3.20, P < 0.01).
В 2008 г. за тот же срок доля мёртвых блох из Тувы составила 4.0 % самок и
20.0 % самцов, из Забайкалья - 32.0 % самок и 60.0 % самцов (соответственно t = 2.58
для самок и 2.89 для самцов, в обоих случаях P < 0.01). Эктопаразиты из Ольхонского
района по смертности занимали промежуточное положение - 24.0 % самок и 36.0 %
самцов, установлены достоверные различия в количестве погибших самок от тувин-
ских блох t = 2.04, P < 0.05; для самцов t = 1.26, Р > 0.05. Различий в количестве мерт-
вых особей между ними и забайкальскими C. tesquorum не выявили: для самок t = 0.63,
для самцов t = 1.70, P > 0.05.
За пять лет после 4-недельного пребывания в снегу погибли 0-46.0 % самок и 16.3-
72.0 % самцов из Тувы. Забайкальские C. tesquorum потеряли 25.4-60.8 % самок и
64.0-82.0 % самцов. У ольхонских C. tesquorum за два года наблюдений учли мёртвы-
ми 31.0 % и 40.0 % самок и 48.0 % и 71.4 % самцов. Несмотря на значительный раз-
брос значений по годам для блох из одной популяции, с увеличением продолжитель-
ности экспериментов наименьшие показатели смертности также принадлежат блохам
из Тувы. За четыре года из пяти различия по доле замёрзших самок между тувинскими
и забайкальскими эктопаразитами были достоверны, величина t-критерия составила:
в 2007 г. 4.16 (Р < 0.001), в 2008 г. 3.07 (P < 0.01), в 2009 г. 1.40 (Р > 0.05), в 2010 г. 2.70
(P < 0.01), в 2011 г. 2.98 (P < 0.01). По самцам обнаружили значимые различия за три
года из пяти: в 2007 г. 4.82 (Р < 0.001), в 2008 г. 0 (Р > 0.05), в 2009 г. 1.19 (Р > 0.05),
в 2010 г. 2.51 (P < 0.05), в 2011 г. 2.09 (P < 0.05). Самки блох ольхонской популяции
по количеству утративших жизнеспособность достоверно опережали самок тувинской
(в 2008 г. t = 3.07, в 2011 г. t = 3.26, P < 0.01), не отличаясь от забайкальских (в 2008 г.
t = 0, в 2011 г. t = 0.56, Р > 0.05). Погибших самцов из этой популяции в 2011 г. отмечено
почти в два раза больше, чем из Тувы (t = 2.17, P < 0.05). В других случаях различия
не выявлены: в 2008 г. при сравнении с забайкальскими и тувинскими - 1.14; в 2011 г.
с забайкальскими - 0.44.
У F. luculenta также отметили значительный разброс значений по годам: в 2007 г.
за 4 недели замёрзло меньше и самок (41.5 %), и самцов (34.3 %), чем в 2009 (соответ-
ственно, 68.8 % и 100 %). В 2009 г. после содержания в снегу в течение 4 недель самки
ольхонских блох убыли в достоверно меньшем количестве, чем таковые из Забайкалья
(t = 2.33, P < 0.05), самцы с Ольхонского района погибли все, из Забайкалья выжил
один самец (t = 0.75, P > 0.05).
Очевидно, блохи из разных популяций могут различаться по зимостойкости. В на-
ших опытах выявлены существенные различия в зимостойкости C. tesquorum из Тувы
и Забайкалья, F. luculenta из Ольхонской и Забайкальской популяций.
122
Устойчивость к зимним температурам у C. tesquorum и F. luculenta,
происходящих из Борзинского района Забайкальского края
Поскольку блохи, доставленные в инсектарий из Борзинского района, были выбраны
из одних и тех же гнёзд, есть основания предполагать сходство их реакций на дей-
ствие низкой температуры. Через месяц после закладки в снег среди блох, извлечён-
ных из пробирок, обнаружили 88.1 % мёртвых самок F. luculenta и 60 % - С. tesquorum
(t = 3.52; P < 0.001), 97.0 % мёртвых самцов F. luculenta и 82.0 % - С. tesquorum
(t = 2.05; P < 0.05). Без учёта пола доля погибших блох F. luculenta составила 91.0 %,
а C. tesquorum достигла 71.0 % (t = 3.60, P < 0.001). Таким образом, блохи С. tesquorum
достоверно превосходили F. luculenta по способности к выживанию при отрицатель-
ных температурах, несмотря на территориальную общность их популяций.
Различия блох в устойчивости к зимним температурам,
обусловленные половыми особенностями
В 2007 г. различий в смертности самок и самцов ни в одной группе не отмечено.
По результатам остальной части опытов самцы погибали чаще самок. В 2008 г. досто-
верность этого факта установлена через две недели для забайкальских С. tesquorum
(t = 1.99, P < 0.05), через четыре недели для тувинских (t = 4.48, P < 0.001). В 2009 г. раз-
личия между самцами и самками достигли значимого уровня у С. tesquorum из Тувы
(t = 2.64, P < 0.01) и Забайкалья (t = 2.42, P < 0.05) и F. luculenta из Ольхонского района
(t = 2.65, P < 0.01). В 2010-2011 гг. различия статистически доказаны во всех опытных
группах, начиная с ольхонских С. tesquorum (t = 2.82, P < 0.01 и выше).
Устойчивость блох к разным отрицательным температурам
Состояние эктопаразитов, только что извлечённых из морозильной камеры и из-под
снега, при визуальной оценке различалось: у блох из холодильника наблюдались вялые
движения конечностей, блохи из снега были совершенно оцепеневшие и перемеща-
лись в пробирке, как сухие песчинки. Доля погибших блох в условиях разных темпе-
ратур показана на рисунке.
Как следует из приведённых данных, при неодинаковых абсолютных величинах от-
рицательной температуры доля погибших особей не различалась. Причём это просле-
живалось у насекомых разного пола, разных видов, из разных популяций. По всей ви-
димости, блохи в состоянии холодового оцепенения малочувствительны к изменениям
температуры. Во всяком случае, срока в один месяц недостаточно для того, чтобы фак-
тор температуры «сработал». Другой вывод состоит в том, что изначально одна часть
имаго предрасположена к переживанию отрицательных температур, а другая часть
имаго - нет. Каждая выборка располагает определённым соотношением тех и других
особей, которое обусловливает свойство популяции перезимовывать в имагинальной
фазе.
ОБСУЖДЕНИЕ
По многолетним наблюдениям, в Туве в холодное время года среднее число блох
С. tesquorum в летних необитаемых гнёздах в 40 раз превышает таковое в зимовочных
(Вержуцкий, 2012). В феврале 1983 г. были добыты шесть нежилых гнёзд, в пяти из
124
Доля погибших блох из разных популяций после 4 недель содержания при температуре от
-1 до -3 °С и от -10 до -15 °С.
Белые столбцы гистограммы - доля погибших блох из разных популяций после 4 недель содержания
при температуре от -1 до -3 °С, чёрные столбцы - при температуре от -10 до -15 °С; над каждой парой
столбцов приведены значения t-критерия. Во всех случаях различия недостоверны.
которых почти половину имаго (104 экз. из 217) составляли С. tesquorum . Температура
в гнездовых камерах опускалась до -12 °С (Ткаченко и др., 1983). В Забайкалье, по дан-
ным О.Ф. Пауллер (1980), С. tesquorum - один из преобладающих видов в зимовочных
гнёздах даурского суслика, среди всех имаго блох 59 % принадлежали к этому виду.
То же самое наблюдал один из авторов данной работы, раскопавший в декабре 1981 г.
гнездо со спящим сусликом. В гнезде находились 94 блохи разных видов, из которых
43 С. tesquorum преимущественно I физиологического возраста. О.Ф. Пауллер отме-
чает, что самки даурского суслика после пробуждения используют зимовочные гнёзда
как выводковые, либо оставляя гнездо в той же норе, либо перенося его на новое место.
Осенью наблюдается обратный процесс, когда суслики используют часть подстилки из
летних нор для устройства зимних гнёзд. Вместе с гнездовым субстратом переезжают
и блохи. В искусственном зимовочном гнезде в присутствии спящего суслика с ноября
по март температура ни разу не опускалась ниже 0.5 °С (Пауллер, 1959). Таким обра-
зом, зимовка С. tesquorum в Забайкалье проходит в более мягких условиях, создавае-
мых присутствием зверька, чем в Туве, в заброшенных летних гнёздах. Не удивитель-
но, что блохи этого вида из Тувы лучше переносят отрицательные температуры, чем
его представители из Забайкалья.
О причинах различий в зимостойкости F. luculenta из разных популяций судить слож-
но, поскольку данных об экологических особенностях этих блох в Ольхонском районе
немного. По данным Г.И. Васильева (1970), по численности на суслике блоха занимает
второе место в Забайкалье (на даурском) и пятое - в Прибайкалье (на длиннохвостом).
125
Зимует в летних гнёздах без хозяина: в Прибайкалье в фазе имаго и куколки (Васильев,
1970), в Забайкалье в фазе имаго и яйца (Вершинин, 1993). Отмечено, что в Прибай-
калье даже в неглубоко расположенных (до 120 см) нежилых гнёздах длиннохвостого
суслика температура зимой не опускается ниже -4 °С, имаго зимуют даже в устье норы
(Васильев, 1970). По наблюдениям О.Ф. Пауллер и П.А. Чипизубовой (1961), наиболь-
шее количество гнёзд даурского суслика устроены на глубине от 50 до 110 см. Сред-
немесячная температура почвы в феврале на глубине 80 см в пос. Борзя опускалась
до -12.8 °С (Жовтый, 1967). Разница в температуре гнездовой подстилки покинутого
гнезда и окружающей почвы не превышает 4 °С (Пауллер, Чипизубова, 1961). Как сле-
дует из приведённых данных, в Забайкалье температурные условия гнезда в отсут-
ствие суслика менее благоприятны. Поэтому особенно холодные и малоснежные зимы
F. luculenta переживают здесь преимущественно в стадии яйца (Вершинин, 1993).
В Забайкалье зимующие имаго обнаруживаются единично в гнёздах даурского сус-
лика (Пауллер, 1980; Вершинин, 1993), незимоспящих грызунов: узкочерепной по-
лёвки (Вершинина, 1971), полёвки Брандта (Вахрушева, 1979), и даурской пищухи
(Акиншина, 1970). Естественно, что в гнёздах, обогретых хозяином, гораздо теплее.
Однако все исследователи указывают сходное состояние блох: голодные неподвижные
I-II физиологических возрастов с развитым жировым телом (Акиншина, 1970; Васи-
льев, 1970; Вершинина, 1971; Вахрушева, 1979; Вершинин, 1993), при этом у ранее
размножавшихся самок происходит резорбция яиц (Вахрушева, 1979). Эти данные
отчасти напоминают наш опыт по сохранению блох при разных по абсолютному зна-
чению отрицательных температурах - количество зимующих имаго не увеличивается
в более тёплых гнёздах. Кроме того, здесь уместно вспомнить о половых различиях,
которые в одних наших экспериментах обнаруживались, а в других нивелировались.
В инсектарии годовые циклы блох нарушаются, поэтому подопытные насекомые име-
ли разный возраст, а также возможность спариваться и размножаться. Эти факторы,
безусловно, имели влияние и на различную выживаемость у самок и самцов, и на боль-
шой разброс в количестве замёрзших особей между опытами разных лет.
Результаты лабораторных испытаний холодостойкости блох С. tesquorum согласуют-
ся с данными полевых наблюдений. Многочисленные зимующие имаго из тувинской
и забайкальской популяций используют для переживания неблагоприятного периода
убежища разного типа. В Туве это, как правило, летние выводковые гнезда, в Забай-
калье - зимовочные. Однако у F. luculenta не наблюдается такого соответствия между
устойчивостью имаго к отрицательным температурам и условиями перезимовки, бо-
лее мягкими в Прибайкалье. Очевидно, основная часть блох этого вида как здесь, так
и в Забайкалье зимует в преимагинальных фазах, а небольшое количество взрослых
особей даже в случае успешной перезимовки не оказывает решающего влияния на по-
пуляцию.
Интересно, что виды С. tesquorum и F. luculenta, занимающие по численности в так-
соценозе блох даурского суслика соответственно первое и второе места, значительно
различаются по способности переживать отрицательные температуры. Забайкальские
F. luculenta в этом отношении оказались самыми уязвимыми среди всех, взятых в опы-
ты. Обращаясь к работе Гусевой и Косминского (1978), обнаруживаем, что нижний
температурный порог жизни у представителей подсемейства Leptopsyllinae составлял
всего -12-14 qС в противоположность Ceratophyllinae, у которых он мог опускать-
ся до -26-28 qС. Хотя F. luculenta относятся к другому подсемейству - Paradoxo-
126
psyllinae - семейства Leptopsyllidae, можно предположить, что плохая переносимость
низких температур у этих блох наследственно обусловлена.
СПИСОК ЛИТЕРАТУРЫ
Акиншина Т. В. 1970. Экология блох даурской пищухи (Ochotona dauurica Pallas, 1776) Забайкальского при-
родного очага чумы. Дис. …канд. биол. наук. Иркутск, 202 с.
Аксёненко Г.Р., Сперанская В.Н. 1966. К вопросу о выживаемости блох Xenopsylla cheopis Roths., 1903
в условиях лесотундровой зоны. В сб.: Степанченко А.И. (ред.). Особо опасные инфекции в Сибири
и на Дальнем Востоке: Доклады Иркутского противочумного института (Кызыл), вып. 7, с. 243-245.
Васильев Г.И. 1966. О спячке длиннохвостого суслика в Прибайкальской лесостепи. В сб.: Степанчен-
ко А.И. (ред.). Особо опасные инфекции в Сибири и на Дальнем Востоке: Доклады Иркутского про-
тивочумного института (Кызыл), вып. 7, с. 176-178.
Васильев Г.И. 1970. Блохи длиннохвостого суслика (видовой состав, экология, эпизоотологическое значение
при чуме). Дис. …канд. биол. наук. Иркутск, 212 с.
Васильев Г.И., Жовтый И.Ф. 1974. Наблюдения за условиями перезимовки длиннохвостого суслика и его
эктопаразитов. В сб.: Лемешева Л.Б. (ред.). Доклады Иркутского противочумного института. Чита,
вып. 10, с. 218-221.
Вахрушева З.П. 1979. Очерк популяционной экологии полёвки Брандта в Забайкальском природном очаге
чумы. Дис. … канд. биол. наук. Чита, 232 с.
Вержуцкий Д.Б. 2012. Пространственная организация населения хозяина и его эктопаразитов. Теорети-
ческие и практические аспекты. Saarbrücken: Palmarium Academic Publishing, 351 c.
Вершинин Е.А. 1993. Экологические особенности блох даурского суслика в Забайкальском природном очаге
чумы. Дис. … канд. биол. наук. Иркутск, 243 с.
Вершинина О.Н. 1971. Эктопаразиты стадной полёвки Забайкальского природного очага чумы. Дис. … канд.
биол. наук. Чита, 265 с.
Гусева А.А., Косминский Р.Б. 1978. О нижней температурной границе жизни некоторых видов блох.
В сб.: Пилипенко В.Г. (ред.). Особо опасные инфекции на Кавказе: Тезисы докладов IV краевой на-
учно-практической конференции по природной очаговости, эпидемиологии и профилактике особо
опасных инфекционных болезней (20-22 декабря 1978 г.). Ставрополь, с. 201-204.
Дубинин В.Б. 1949. Птицы даурской степи и их роль в распространении блох. В сб.: Алтарева Н. Д. (ред.).
Известия Иркутского государственного научно-исследовательского противочумного института Сиби-
ри и Дальнего Востока. Иркутск: Иркутское областное государственное издательство, т. 7, с. 237-253.
Жовтый И.Ф., Васильев Г.И. 1962. Температурные условия обитания блох в шерсти грызунов. В сб.: Некипе-
лов Н. В. (ред.). Доклады Иркутского противочумного института, вып. 4, с. 152-155.
Жовтый И.Ф. 1968. Изучение условий обитания блох в норах грызунов Сибири и Дальнего Востока. В сб.:
Сафонова А.Д. (ред.). Носители и переносчики возбудителей особо опасных инфекций Сибири и
Дальнего Востока: Известия Иркутского государственного научно-исследовательского противочум-
ного института Сибири и Дальнего Востока. Кызыл, т. 27, с. 212-230.
Жовтый И.Ф., Вершинин Е.А., Ромашева Т.П. 1984. О свойствах популяции эвризоидной блохи Frontopsylla
luculenta luculenta I. et R. 1923 (Siphonaptera) в Юго-Восточном Забайкалье. В сб.: Голубинский Е.П.
(ред.). Современные аспекты профилактики зоонозных инфекций: Тезисы докладов к Всесоюзной
научной конференции специалистов противочумных учреждений. Иркутск, т. 1, с. 72-74.
Жовтый И.Ф., Плеснивцева Г.Г. 1986. Эктопаразиты млекопитающих Якутии. Якутск: Якутский филиал СО
АН СССР, 168 с.
Иофф И.Г., Скалон О.И. 1954. Определитель блох Восточной Сибири, Дальнего Востока и прилегающих
районов. М.: Медгиз, с. 84-86.
Олькова Н.В. 1972. Температура тела у грызунов и зайцеобразных с различной инфекционной чувствитель-
ностью к чуме. Проблемы особо опасных инфекций, Саратов, т. 3, с. 46-50.
Пауллер О.Ф. 1959. О зимней спячке длиннохвостого суслика и условиях перезимовки блох в его гнезде.
В сб.: Домарадский И.В. (ред.). Известия Иркутского государственного научно-исследовательского
противочумного института Сибири и Дальнего Востока, Иркутск, т. 21, с. 334-339.
Пауллер О.Ф., Чипизубова П.А. 1961. Условия зимовки и состояние блох в жилых и нежилых гнёздах ран-
ней весной. В сб.: Степанченко А.И. (ред.). Доклады Иркутского противочумного института, Чита,
вып. 2, с. 89-91.
Пауллер О.Ф. 1980. Особенности строения зимовочных нор даурского суслика и состояние блох в них
в зимний период в Забайкальском очаге чумы. В сб.: Лемешева Л.Б. (ред.). Проблемы природной оча-
говости чумы. Иркутск, ч. 1, с. 80-82.
127
Рокицкий П.Ф. 1973. Биологическая статистика. Минск: Вышэйшая школа, 320 с.
Тифлов В.Е., Фаворисова Б.Ю. 1940. Холодостойкость блох. Вестник микробиологии, эпидемиологии
и паразитологии 19 (2): 295-302.
Ткаченко В.А., Вержуцкий Д.Б., Верзилова Т.Н., Чарная Т.Г., Колосов В.М. 1983. О сохранении возбудите-
ля чумы в межэпизоотический сезон в Тувинском природном очаге. В сб.: Задорожный В.Ф. (ред.).
Природно-очаговые инфекции в Забайкалье: Тезисы докладов научной конференции. Чита, с. 17-19.
Федорова Л.В. 1953. Блохи жилищ человека и домашних животных Юго-Восточного Забайкалья. В сб.: Ал-
тарева Н.Д. (ред.). Известия Иркутского государственного научно-исследовательского противочумно-
го института Сибири и Дальнего Востока, Иркутск, т. 11, с. 198-202.
Холин А.В. 2013. Субвидовые группировки длиннохвостого суслика (Spermophilus undulatus Pallas, 1778)
в Южной Сибири (на примере Юго-Западной Тувы и Прибайкалья). Автореф. дис. … канд. биол.
наук. Иркутск, 25 с.
Kiefer D., Kiefer M.S., Lobachev V.S., Samiya R., Samiya D., Tserennorov D. 2006. Distribution of fleas in
dependence of climatical factors in Mongolia. Scientific Journal (Ulaanbaatar) 14: 111-115.
ABILITY OF FLEAS CITELLOPHILUS TESQUORUM AND FRONTOPSYLLA
LUCULENTA LUCULENTA (SIPHONAPTERA, CERATOPHYLLIDAE)
TO SURVIVE AT NEGATIVE TEMPERATURES DURING WINTER
E. G. Tokmakova, L. P. Bazanova, E. A. Vershinin
Key words: flea, Citellophilus tesquorum, Frontopsylla luculenta, wintering, cold resistance.
SUMMARY
Comparative research of the ability of two species fleas from different populations to survive at negative
temperatures during winter time is conducted. It is shown that the share of Imago fallen considerably differs
depending both on its species and population and sex belonging. Сitellophilus tesquorum significantly
exceeded Frontopsylla luculenta fleas by the ability to survive at negative temperatures even at the territorial
commonality of its populations. Essential interpopulation distinctions in winter resistance were revealed in
C. tesquorum from Tuva and Transbaikalia, F. luculenta from Olkhon area of the Irkutsk region and
Transbaikalia. Males died more often than females in the majority of experiments. At the same time
distinctions in number of the fallen fleas being at unequal absolute negative temperatures during the
experiment were not revealed. It is suggested that the population ability to wintering in Imago phase
is caused by the correlation of individuals that ancestrally and physiologically predisposed to survive
at negative temperatures. Results of the experiments are compared with the data of field observations.
128