ПАРАЗИТОЛОГИЯ, 2019, том 53, № 3, с. 179-197.
УДК 576.895.775:574.9/579.842.23
РАЗНООБРАЗИЕ БЛОХ (SIPHONAPTERA) - ПЕРЕНОСЧИКОВ
ВОЗБУДИТЕЛЕЙ ЧУМЫ: ПАРАЗИТ СУСЛИКОВ - БЛОХА
CITELLOPHILUS TESQUORUM (WAGNER, 1898)
© 2019 г. С. Г. Медведевa, *, Б. К. Коттиb, **, Д. Б. Вержуцкийc, ***
a Зоологический институт РАН,
Университетская наб., 1, Санкт-Петербург, 199034 Россия
b Северо-Кавказский федеральный университет,
ул. Пушкина, 1, Ставрополь, 355009 Россия
c Иркутский научно-исследовательский противочумный институт,
Трилиссера 78, Иркутск, 664047 Россия
* e-mail: smedvedev@zin.ru, sgmed@mail.ru
** e-mail: boris_kotti@mail.ru
*** e-mail: verzh58@rambler.ru
Поступила в редакцию 17.02.2019 г.
После доработки 05.03.2019 г.
Принята к публикации 05.03.2019 г.
Проанализированы таксономическое разнообразие, характер распространения и паразито-хо-
зяинные связи видов блох, отмеченных в качестве переносчиков возбудителя чумы. Рассмо-
трены экологические особенности и эпизоотологическое значение паразита сусликов - блохи
Citellophilus tesquorum (Wagner, 1898).
Ключевые слова: блохи, Siphonaptera, виды-переносчики возбудителя чумы, таксономиче-
ское разнообразие, Citellophilus tesquorum.
DOI: 10.1134/S0031184719030013
Данная работа является первой, посвященной анализу и обобщению некоторых све-
дений, касающихся особенностей распространения и паразито-хозяинных связей ви-
дов блох - активных переносчиков возбудителя чумы. Рассмотрено таксономическое
разнообразие видов блох и их хозяев, из которых в естественных условиях различных
регионов мира выделяли возбудитель чумы, обобщены данные по распространению
и эпизоотологическому значению блох рода Citellophilus Wagner, 1934. Особое внима-
ние уделено паразиту сусликов - блохе C. tesquorum (Wagner, 1898).
Чума - одно из наиболее опасных инфекционных заболеваний за всю историю че-
ловечества. В эпидемиологическом плане высокие риски быстрого распространения
этого патогена среди людей по планете сохраняются и в настоящее время. Природные
очаги чумы расположены на обширных территориях всех материков, кроме Австралии
и Антарктиды. Особенно настораживает наблюдаемая в последние годы необычная
активизация ряда природных очагов чумы в Центральной Азии (Балахонов и др., 2014;
Вержуцкий, 2018), на Мадагаскаре и в ряде других регионов мира (Bertherat, 2016).
179
В степных и пустынных зонах равнинных и горных районов страны, от Кавказа до
Забайкалья, а также на территории ряда стран, примыкающих к южной границе Рос-
сии, расположены многочисленные очаги чумы разных типов. Основными носителями
чумы являются различные виды мелких норовых млекопитающих, основными перено-
счиками и хранителями - блохи. При оценке роли тех или иных видов блох в качестве
переносчиков возбудителя необходимо принимать во внимание то, что чумной микроб
хорошо адаптирован только к небольшой группе видов, которые способны его сохра-
нять продолжительное время, включая межэпизоотические сезоны (Ващенок, 1988,
1999). Так, на территории Российской Федерации и соседних стран только 19 видов
считаются основными переносчиками чумы (Природные очаги чумы…, 2004; Гонча-
ров и др., 2013). Несколько большее число видов блох отмечалось как дополнительные
переносчики, которые способны участвовать в передаче возбудителя, заражаясь им
при питании на инфицированном хозяине.
В основу настоящей работы положен анализ литературных данных. Значительная
часть из них унифицирована и обобщена средствами электронных таблиц Excel. Так,
были проанализированы обширные сведения, приведенные в главе 5 сводки «Yersinia
pestis: Retrospective and Perspective Ecology of Yersinia pestis (Advances in Experimental
Medicine and Biology)». Обобщены и проанализированы сведения, почерпнутые из
таблиц 5.6 «Виды млекопитающих - носителей чумы в различных странах мира» и
5.7 «Виды и подвиды блох, инфицированные в очагах чумы различных стран мира»
(Dubyanskiy, Yeszhanov, 2016). Все собранные материалы сопоставлены с данными по
мировой фауне блох из информационно-аналитической системы (ИАС) PARHOST1,
разрабатываемой в Зоологическом институте РАН с 2001 г.
Дополнительно рассмотрены материалы приложения 3 к методическому пособию
«Список видов и подвидов блох, обнаруженных зараженными возбудителем чумы
в естественных условиях» (Гончаров и др., 2013). Данная таблица содержит сведения
о видовом составе блох, инфицированных возбудителем чумы, в 45 очагах на терри-
тории России и сопредельных стран. Сводка сведений о значении блохи C. tesquorum
в природных очагах чумы Евразии представлена в табл. 4 настоящей публикации.
Блохи и их хозяева - переносчики и носители возбудителей чумы
Большинство представителей теплокровных и пойкилотермных животных вовлека-
ется в эпизоотический процесс случайно и существенной роли в поддержании энзоот-
ии чумы не играет. Число видов-носителей и переносчиков, которые реально обеспе-
чивают жизнедеятельность возбудителя и его успешное выживание, намного меньше
и в конкретных природных очагах, как правило, ограничивается несколькими видами.
В некоторых случаях в поддержании эпизоотического процесса или сохранении чум-
ного микроба могут принимать участие дополнительные или второстепенные виды,
включающиеся в циркуляцию возбудителя в разные фенологические сезоны или на
разных фазах эпизоотического цикла (Ралль, 1965).
Естественная зараженность чумой установлена у 233 видов млекопитающих (Кари-
мова и др., 2010) и, как минимум, у 244 видов и подвидов блох мировой фауны (Гон-
чаров, Плотникова, 2010). Наш анализ данных, опубликованных позднее (Dubyanskiy,
Yeszhanov, 2016), показывает, что виды блох, обнаруженные инфицированными чумой
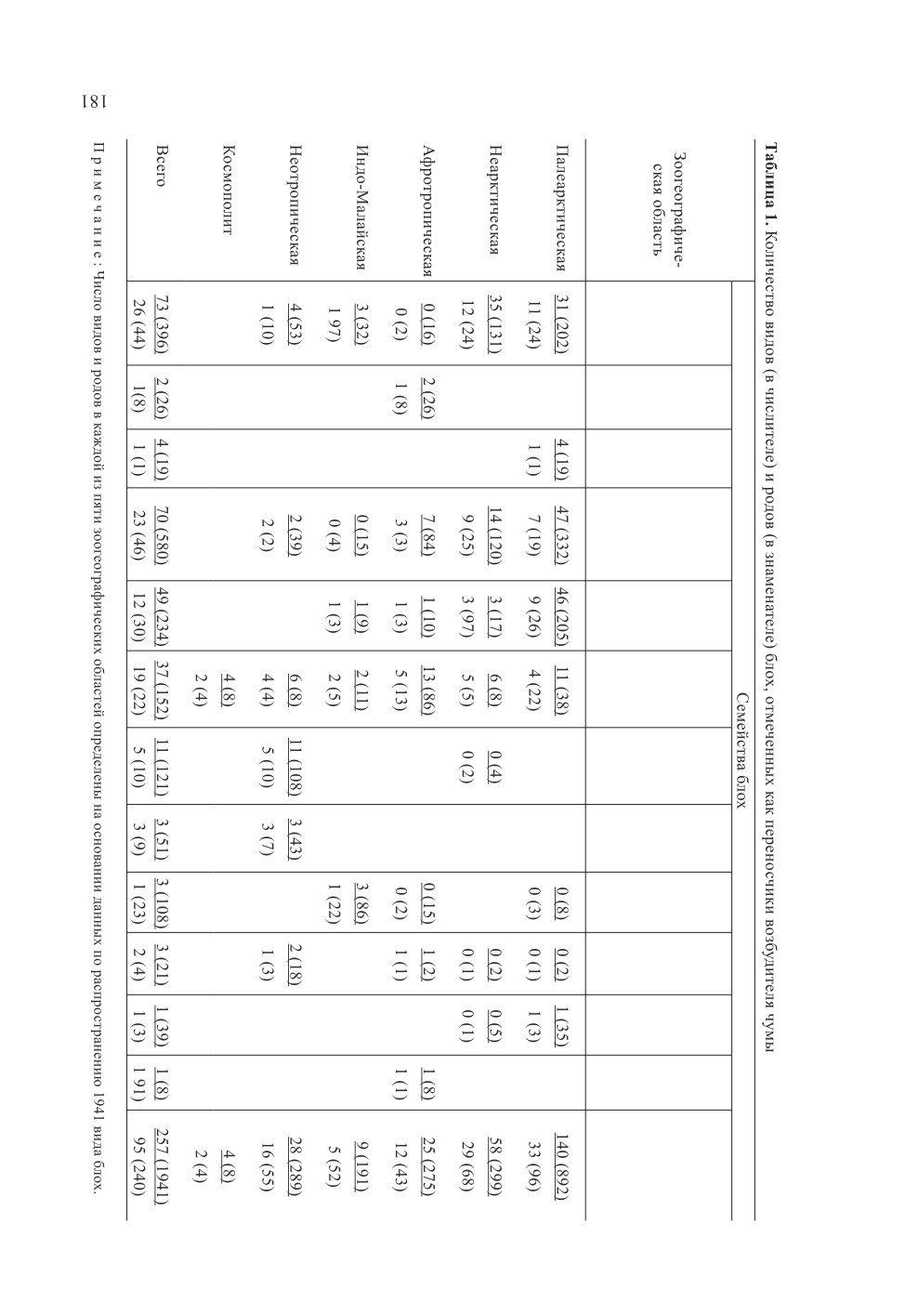
в природе, принадлежат к 95 из 240 родов и к 12 из 18 семейств блох (табл. 1). Это
составляет, соответственно, 40 % от числа родов, известных в мировой фауне. К насто-
180
Ceratophyllidae
Chimaeropsyllidae
Coptopsyllidae
Hystrichopsyllidae
Leptopsyllidae
Pulicidae
Rhopalopsyllidae
Stephanocircidae
Stivaliidae
Tungidae
Vermipsyllidae
Xiphiopsyllidae
Общий итог
ящему времени зараженность чумой отмечена у 257 видов блох, что составляет 12 %
от 2162 всех известных видов. Следует отметить, что к настоящему времени в мировой
фауне также описано 846 подвидов блох.
Наибольшее число видов, из которых был выделен возбудитель чумы в естественных
условиях в 5 зоогеографических областях мира, принадлежит к 3 крупнейшим семей-
ствам блох - Leptopsyllidae, Ceratophyllidae и Hystrichopsyllidae. В фауне этих областей
среди представителей первого семейства возбудитель был выделен у 21 % видов и
40 % родов, второго - 18 % видов и 60 % родов, третьего - 12 % и 50 %, соответствен-
но. Эти 3 семейства в целом тяготеют к Северному полушарию (Медведев, 2000), где
расположено наибольшее число очагов чумы. В фауне России они известны по 213
видам, что составляет 83 % от всех известных на этой территории (Медведев, 1998,
2013а, 2013б). Доля видов сем. Hystrichopsyllidae в фауне Палеарктики составляет око-
ло 37 %, а семейств Leptopsyllidae и Ceratophyllidae - по 23 % каждое. Высокая доля
видов (24 %), из которых выделялись возбудители чумы, а также родов, к которым они
принадлежат (86 %), наблюдается в сем. Pulicidae.
В качестве носителей возбудителя чумы к настоящему времени отмечены представи-
тели 9 отрядов млекопитающих и 1 отряда птиц (табл. 2). Носительство возбудителей
отмечено для 346 из 5937 видов (около 6 %)1, 144 из 1258 родов (около 10 %), 33 из
156 семейств (около 20 %), 9 из 28 отрядов (около 30 %) млекопитающих. Наибольшую
долю видов-носителей (34 %) составляют представители сем. хомяковых (Cricetidae).
Значительна также общая доля других представителей отряда грызунов: виды мыши-
ных (Muridae) составляют 22 %, беличьих (Sciuridae) - 15 %. Доля видов из других
отрядов и семейств млекопитающих - зайцеобразных (Leporidae), хищных (Mustelidae
и Viverridae) и насекомоядных (Soricidae) - значительно меньше (от 2 до 4 %).
Среди носителей чумы наибольшая доля видов (30 %) и родов (34 %), принадлежа-
щих к 15 семействам млекопитающих и 5 семействам птиц, отмечена на территории
Палеарктики. Отмечалось, что на территории Евразии основными носителями возбу-
дителя чумы в ее очагах являются суслики, сурки, песчанки, крысы, полевки и пищу-
хи (Каримова, Неронов, 2007). Согласно проведенному нами анализу, на территории
Палеарктики наибольшее число видов-носителей чумы отмечено не только среди
хомяковых и беличьих, но и среди тушканчиковых. На территории Неарктической
области - преимущественно среди беличьих и хомяковых, в Неотропической обла-
сти - преимущественно хомяковых, а в Афротропической и Индо-Малайской обла-
стях - мышиных. Число родов, к которым принадлежат данные виды, также отра-
жает таксономическое разнообразие носителей возбудителя чумы среди хомяковых
в Палеарктической и Неотропической областях, среди мышиных - в Индо-Малайской
и Афротропической.
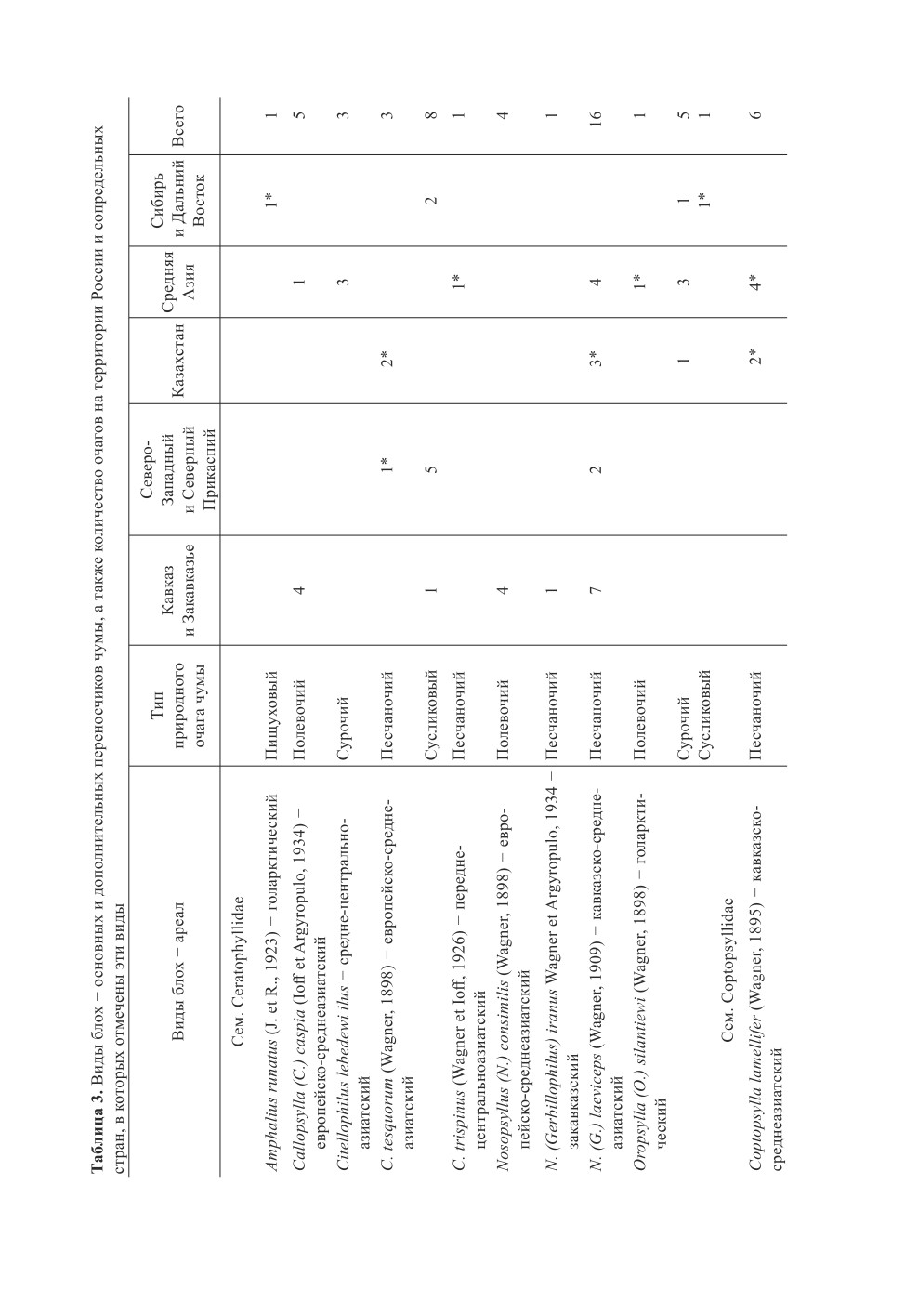
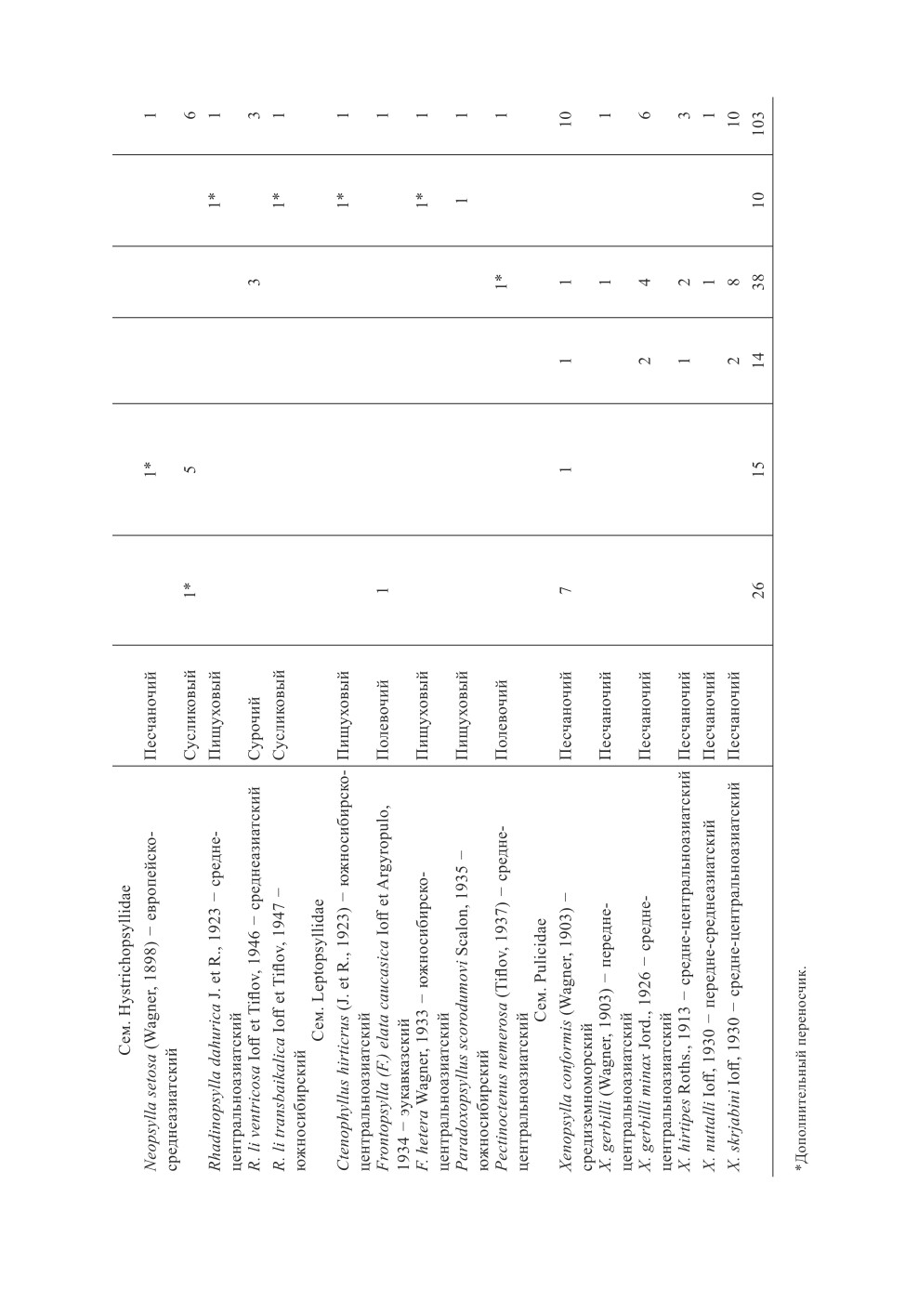
В целом на территории России и сопредельных стран отмечается следующее рас-
пределение очагов и видов блох - основных переносчиков возбудителей чумы
(табл. 3). Равнинные и горные очаги сусликового типа охватывают обширные террито-
рии Северного Кавказа, Северо-Западного и Северного Прикаспия, а также Нижнего
Поволжья. Здесь основными носителями возбудителя чумы являются малый и горный
суслики [Spermophilus pygmaeus (Pallas, 1778) и S. musicus (Menetries, 1832)]. В Сибир-
182
ской части России расположен Тувинский очаг, где носителем возбудителя является
длиннохвостый суслик [S. undulatus (Pallas, 1778)], а также Забайкальский очаг, где эту
же роль выполняет даурский суслик [S. dauricus (Brandt, 1844)]. Во всех этих очагах
основным переносчиком, а и хранителем чумной инфекции является политипический
вид - блоха Citellophilus tesquorum (Wagner, 1898), включающий 4 подвида.
Блоха C. tesquorum была отмечена в качестве основного переносчика чумы в поли-
гостальном Прикаспийском песчаночьем очаге (Гончаров и др., 2013), где основными
носителями являются не только песчанки, но и малый суслик.
Другой вид, С. trispinus - второстепенный переносчик возбудителя чумы в Му-
юнкумском пустынном песчаночьем очаге, где основным носителем является боль-
шая песчанка, а главным хозяином - С. trispinus; желтый суслик [Spermophilus fulvus
(Lichtenstein, 1823)] служит дополнительным носителем.
В 2012 г. (Балахонов и др., 2014) обнаружено проникновение возбудителя чумы ос-
новного (высоковирулентного) подвида из Монголии в Горный Алтай и его дальней-
шее распространение в популяциях серого сурка и блохи Oropsylla silantiewi (Wagner,
1898). Сейчас на этой территории фактически существуют 2 совмещенных очага,
один очаг - на монгольской пищухе и связанных с нею блохах, второй - на сурке
и O. silantiewi.
Предполагается, что блоха Neopsylla setosa (Wagner, 1898) в европейской части Рос-
сии в некоторых очагах чумы является вторым основным переносчиком. Обширный
(Европейско-сибирско-азиатский) ареал этого политипического вида (он насчитывает
2 подвида) охватывает зону степей и полупустынь от Восточной Европы до Передней,
Средней и Центральной Азии и Южной Сибири (Котти, 2018). Однако в восточной или
сибирской части ареала этот вид как переносчик не отмечался.
В Туве дополнительным переносчиком возбудителя чумы в настоящее время являет-
ся южносибирский вид - блоха Rhadinopsylla (Ralipsylla) li transbaikalica Ioff et Tiflov,
1947 (Галацевич, 2018). При этом восточнее (Забайкальский очаг) дополнительных
переносчиков выявлено не было. Однако среди случайных переносчиков отмечался
другой представитель рода Neopsylla Wagner, 1903 - южносибирский вид N. abagaitui
Ioff, 1946, а также ряд других видов.
Блохи рода Citellophilus
Видовой состав 46 родов сем. Ceratophyllidae представлен различным числом, но
только в 14 родах количество видов более 10. Так, род Ceratophyllus Curtis, 1831 на-
считывает 66 видов, а род Nosopsyllus Jordan, 1933 - 54. Согласно одним источни-
кам род Сitellophilus насчитывает 11 видов и 8 подвидов (Traub et al., 1983), согласно
другим - 13 видов и 7 подвидов (Lewis, 2003).
Расцвет наземных беличьих в Евразии приходится на позднеплиоценовое время
(2.5-3.5 млн. лет назад) (Громов, Баранова, 1981; Павлинов и др.,1995). Вероятно,
именно тогда сформировалось большинство видов рода Сitellophilus.
Блохи рода Citellophilus по признакам строения наиболее сходны с блохами другого
палеарктического рода, Callopsylla Wagner, 1934, 28 видов которого распределены по
4 подродам и паразитируют на широком круге хозяев, включая не только грызунов,
183
зайцеобразных и насекомоядных, но и птиц. Виды рода Citellophilus паразитируют на
представителях сем. Sciuridae, обитающих в норах, а среди них - преимущественно на
сусликах рода Spermophilus Cuvier, 1825 (=Citellus). Несколько видов рода Citellophilus
являются постоянными паразитами сурков рода Marmota Blumenbach, 1779 (Traub
et al., 1983). Вследствие этого, виды Citellophilus отмечаются в качестве основных пе-
реносчиков в сусликовых и сурочьих очагах чумы на территории России и сопредель-
ных стран (Онищенко, Кутырев, 2004).
Следует отметить, что, согласно данным ИАС PARHOST1 по мировой фауне блох,
виды рода Citellophilus в целом были отмечены на 66 видах из 6 семейств грызунов.
В частности, их обнаруживали на 22 видах из 4 родов сем. Sciuridae, 31 видах из 15 ро-
дов сем. Cricetidae, 8 видах из 5 родов сем. Dipodidae, 4 видах из 2 родов сем. Muridae,
1 виде из 1 рода сем. Gliridae и 1 виде из 1 рода сем. Zapodidae.
Род Citellophilus относится к 6 родам блох, имеющих транспалеарктические ареа-
лы. В частности, такой тип ареала также имеют паразиты сусликов, песчанок и туш-
канчиков из родов Ophthalmopsylla Wagner et Ioff, 1926 и Mesopsylla Dampf, 1910
(Leptopsyllidae) и Coptopsylla Jordan and Rothschild, 1908 (Coptopsyllidae), а также па-
разиты пищух, сурков, полевочьих и хомяковых из рода Callopsylla (Ceratophyllidae).
Широкое транспалеарктическое распространение имеют паразиты ежей - блохи рода
Archaeopsylla Dampf, 1908 (Pulicidae).
Виды рода Citellophilus обитают в степных и горных областях Центральной и Юж-
ной Европы, Казахстана, Передней, Средней и Центральной Азии, Южной Сибири и
Приамурья. Однако больше всего представителей этого рода обитает на территории
Казахстана, Средней и Центральной Азии.
Ниже эти данные подытожены для 11 видов и 12 подвидов рода Citellophilus.
1. Citellophilus gracilis (Mikulin,
1957). Ареал
- восточно-палеарктический,
сибирско-центральноазиатский: Джунгарский Алатау. Основной хозяин - красноще-
кий суслик [Spermophilus erythrogenys (Brandt, 1841)].
2. Citellophilus jenissejensis (Wagner, 1902). Ареал - центрально-восточнопалеаркти-
ческий, сибирский: горы Центральной Азии и юг Средней Сибири. Основной хозяин
- длиннохвостый суслик [Spermophilus undulates (Pallas, 1778)].
3. Citellophilus lebedewi (Wagner, 1933). Ареал - центрально-палеарктический, цен-
тральноазиатско-туранский: Тянь-Шань, Памиро-Алай и Гиндукуш. Основной хозя-
ин - красный [Marmota caudata (Geoffroy, 1842)] и серый [M. baibacina Kastschenko,
1899)] сурки.
Citellophilus lebedewi lebedewi. Ареал - восточно-палеарктический, центральноази-
атский: Центральный Тянь-Шань. Основной хозяин - красный и серый сурок.
Citellophilus lebedewi princeps (Ioff, 1946). Ареал - центральнопалеарктический,
туранский: Западный и Восточный Тянь-Шань, Памиро-Алай и Гиндукуш. Основной
хозяин - красный сурок.
4. Citellophilus martinoi (Wagner et Ioff, 1926). Ареал - западнопалеарктический,
европейский: Центральная и Юго-Восточная Европа. Основной хозяин - европейский
суслик [Spermophilus citellus (L., 1766)].
188
Citellophilus martinoi martinoi. Ареал - западнопалеарктический, Центральная и
Юго-Восточная Европа. Основной хозяин - европейский суслик.
Citellophilus martinoi rotundus Rosicky, 1956. Ареал - западнопалеарктический, евро-
пейский: Центральная Европа. Основной хозяин - европейский суслик.
5. Citellophilus menzbieri (Ioff, 1950). Ареал - центрально-палеарктическо-туран-
ский, туранский: Западный Тянь-Шань. Основной хозяин - сурок Мензбира (Marmota
menzbieri Kashkarov, 1925).
6. Citellophilus relicticola (Fedina, 1946). Ареал - центральнопалеарктическо-туран-
ский, туранский: Центральный и Западный Тянь-Шань, Синьцзянь. Основной хозяин -
тянь-шаньский [Spermophilus relictus (Kashkarov, 1923)] и рыжеватый [S. major (Pallas,
1779)] суслики.
7. Citellophilus simplex (Wagner, 1902). Ареал - западно-палеарктический, евро-
пейско-восточносредиземноморский: Восточная Европа и Малая Азия. Основной
хозяин - европейский и крапчатый [Spermophilus suslicus (Guldenstaedt, 1770)] сусли-
ки.
Citellophilus simplex simplex. Ареал - западно-палеарктический, европейско-восточ-
носредиземноморский: Восточная Европа и Малая Азия.
Citellophilus simplex rosickyi Cyprich, 1989. Ареал - западнопалеарктический, евро-
пейский: Центральная Европа.
8. Citellophilus sungaris (Jordan, 1929). Является самостоятельным видом по Льюису
(Lewis, 1990) и подвидом Citellophilus tesquorum согласно некоторым другим авторам.
Ареал
- центрально-восточно-палеарктический, Сибирско-Восточноазиатско-Цен-
тральноазиатский: Прибайкалье, Забайкалье, Якутия, Приамурье, Северо-Восточный
Китай, Юго-Восточный Алтай, восточная часть Котловины Больших озер и Монголь-
ского Алтая, Гобийский район (кроме восточной части), равнины Восточной Монго-
лии. Основной хозяин - даурский и длиннохвостый суслики.
Citellophilus sungaris lobatschevi (Cyprich, Kiefer et Krumpál, 1985). Ареал - восточно-
палеарктический, центральноазиатский: Монголия. Хозяева - длиннохвостый и
краснощекий [S. erythrogenys (Brandt, 1843)] суслики.
9. Citellophilus tesquorum (Wagner, 1898). Ареал - транспалеарктический, европей-
ско-сибирско-центрально-восточноазиатский: Южная Европа, Кавказ, Казахстан,
Средняя и Центральная Азия, Южная Сибирь. Паразит сусликов различных видов
рода Spermophilus.
Citellophilus tesquorum tesquorum. Ареал - западно-палеарктический, европейский:
юго-восточная часть Украины и Ростовской области до низовий Донца и Дона. Основ-
ной хозяин - малый суслик.
Citellophilus tesquorum altaicus (Ioff, 1936). Льюис (Lewis, 1990) считает C. t. altaicus
отдельным видом. Ареал - восточнопалеарктический, cибирско-восточноазиатский:
Зайсанская Котловина, Джунгария, южные склоны Джунгарского Алатау, Алтай,
Западный Саян, Хакассия, Тува, западная часть Котловины Больших озер, Северного
189
Хангая, Монгольского и Гобийского Алтая. Основной хозяин - длиннохвостый и крас-
нощекий суслики.
Citellophilus tesquorum ciscaucasicus (Ioff, 1936). Ареал - западно-палеарктиче-
ский, Европейский: между Донцом и Волгой, в Восточном Предкавказье. Основной
хозяин - малый суслик.
Citellophilus tesquorum dzetysuensis (Mikulin, 1951). Ареал - восточно-палеарктиче-
ский, центрально-азиатский: северные (в восточной части и южные) склоны Джунгар-
ского Алатау. Основной хозяин - длиннохвостый суслик.
Citellophilus tesquorum elbrusensis Goncharov, 2011. Ареал - западно-палеарктиче-
ский, европейский: центральная часть Большого Кавказа. Основной хозяин - горный
суслик [S. musicus (Menetries, 1832)].
Citellophilus tesquorum mongolicus (J. et R., 1911). Ареал - восточнопалеарктический,
центральноазиатский: Северный Китай, Манчжурия. Основной хозяин - даурский сус-
лик [Spermophilus dauriucus (Brandt, 1844)].
Citellophilus tesquorum transvolgensis (Ioff, 1936). Ареал - транспалеарктический, ев-
ропейско-сибирский: Заволжье, Казахстан и южная часть Западной Сибири. Основной
хозяин - малый суслик.
Citellophilus tesquorum transcaucasicus (Ioff et Argyropulo, 1934). Ареал - западно-
палеарктический, европейский: Малый Кавказ (Северо-Западная Армения и Севе-
ро-Восточная Турция). Основной хозяин - малоазийский суслик [Spermophilus xantho-
prymnus (Bennett, 1835)].
10. Citellophilus trispinus (Wagner et Ioff, 1926). Ареал - центрально-палеарктиче-
ский, туранско-иранский: равнины Казахстана, Средней, Центральной и Передней
Азии. Основной хозяин - желтый суслик.
Citellophilus trispinus trispinus (Wagner et Ioff, 1926). Ареал - центрально-палеаркти-
ческий, туранско-иранский: Казахстан (кроме восточной части), Средняя, Центральная
и Передняя Азия. Основной хозяин - желтый суслик
Citellophilus trispinus balkhaschensis (Mikulin, 1958). Ареал - центрально-палеаркти-
ческий, туранский: Казахстан (восточная часть), Синьцзян. Основной хозяин - желтый
суслик.
11. Citellophilus ullus (Mikulin, 1957). Ареал - центрально-палеарктический, туран-
ский: Казахстан, Средняя и Центральная Азия. Основной хозяин - краснощекий сус-
лик.
Блоха Citellophilus tesquorum
Как указывалось, основным переносчиком и хранителем возбудителя чумы в ряде
очагов инфекции на территории России является политипический вид Citellophilus
tesquorum. Он имеет ареал транспалеарктического типа, который в целом может быть
охарактеризован как европейско-сибирско-центральноазиатский. Данный вид при-
надлежит к обширной среди блох группе олигоксенных видов, паразитирующих на
близких в систематическом отношении видах хозяев. Оликогсенные паразиты могут
встречаться и на хозяевах из других таксономически групп, однако определяющей для
190
них является связь с хозяевами определенной таксономической группы. К этой груп-
пе относятся также паразиты песчанок рода Meriones Illiger, 1811 - блохи Xenopsylla
conformis (Wagner, 1903) и Nosopsyllus laeviceps (Wagner, 1909). Основными хозяевами
блохи Citellophilus tesquorum являются суслики рода Spermophilus (Медведев, 2002).
В пределах Старого Света встречается 14 видов сусликов, из которых 13 относится
к роду Spermophilus (алашанский, беренгийский, большой, горный, даурский, длин-
нохвостый, европейский, желтый, крапчатый, краснощекий, малоазийский, малый,
реликтовый) и 1 вид - к роду Spermophilopsis (тонкопалый) (Громов, Ербаева, 1995;
Wilson, Reeder, 2005). Citellophilus tesquorum либо доминирует, либо входит в число
2-3 содоминантов в сообществах блох у всех перечисленных видов сусликов, за не-
многими исключениями. Так, некоторые подвиды блох, свойственные обособленным
территориям или отдельным видам сусликов, рассматриваются в настоящее время
как самостоятельные виды, например C. jenissejensis (форма, обитающая на длинно-
хвостом суслике, на юге Красноярского края и в Хакассии) или С. relicticola (вид, оби-
тающий на Тянь-Шане и связанный с реликтовым сусликом).
В границах ареала евразийских сусликов известно 11 очагов чумы, где суслики
считаются основными носителями инфекции. Общая площадь этих очагов состав-
ляет чуть менее 500 тыс. км2 (табл. 4). С малым сусликом функционально связаны
5 очагов: Прикаспийский северо-западный степной, Терско-Сунженский низкогорный,
Дагестанский равнинно-предгорный, Волго-Уральский степной и Зауральский степ-
ной. Первые 3 находятся полностью на территории Российской Федерации, четвертый
- значительной частью заходит в Казахстан, пятый расположен в Казахстане. В двух
очагах основным носителем является даурский суслик: Забайкальский степной (на тер-
ритории РФ, Монголии и Китая) и Южно-Маньчжурский (находится на северо-востоке
Китая). В Тувинском горном (РФ) и Восточно-Тяньшаньском Северном (КНР) очагах
основным носителем является длиннохвостый суслик. В Центрально-Кавказском вы-
сокогорном очаге (РФ) в качестве основного носителя чумы выступает горный суслик,
в очаге чумы Лессового Плато (в стыке провинций Ганьсу и Нинся в КНР) - алашан-
ский суслик. В других природных очагах чумы Евразии суслики вовлекаются в эпи-
зоотии лишь спорадически - как правило, при высокой активности эпизоотического
процесса (The Atlas…, 2000; Природные…, 2004; Каримова, Неронов, 2007, Атлас…,
2012; Dubyanskiy, Yeszhanov, 2016).
Среди перечисленных природных очагов чумы, где основными носителями являются
суслики, а единственным или одним из основных переносчиков служит C. tesquorum,
2 очага - Тувинский (РФ) и Ганьсу-Нинсянский (КНР) отличаются высокой эпизооти-
ческой активностью по настоящее время.
В Зауральском степном очаге (Казахстан) в последние годы регистрируются эпизо-
отии с невысокой или средней активностью. В Забайкальском степном очаге чумной
микроб не обнаруживается с 1971 г., Прикаспийский Северо-Западный степной очаг
не проявляет себя с 1991 г., Южно-Маньчжурский - с 1996 г., Терско-Сунженский не-
активен с 2001 г., Волго-Уральский степной - с 2002 г., Дагестанский равнинно-пред-
горный - с 2004 г., Центрально-Кавказский высокогорный - с 2008 г. Следует отме-
тить, что в большинстве очагов, где перестали выделяться культуры возбудителя чумы,
периодически обнаруживают антитела к чумному микробу в крови животных или ДНК
191
возбудителя в живых объектах, что свидетельствует о сохранении патогена в местных
биоценозах.
При систематизации результатов проведенных ранее экспериментальных работ
авторы (Бибикова, Классовский, 1974) пришли к выводу о низкой способности бло-
хи C. tesquorum к формированию блока преджелудка. Ващенок (1988) отнес подвид
C. t. ciscaucasicus к активным переносчикам возбудителя чумы, а C. t. altaicus - к
малоактивным. В более поздней работе тот же автор указывает, что в очагах чумы, где
основными носителями являются малый и горный суслики, «передача инфекции осу-
ществляется высокоактивным переносчиком Neopsylla setosa и несколько уступающим
ему в этом отношении Citellophilus tesquorum» (Ващенок, 1999, с. 199). Необходимо
уточнить, что в отношении Центрально-Кавказского очага чумы - единственного, где
основным носителем является горный суслик - роль N. setosa, как переносчика чумы,
относительно велика только в восточной части очага, которой ограничено его распро-
странение. От N. setosa получено 2.4 % культур микроба, выделенных от блох, в отли-
чие от C. tesquorum, от которого изолировано 83.3 % штаммов (Природные…, 2004).
Следует учитывать, что на блокообразование и эффективность передачи возбудителя
влияет много разных факторов - температура и влажность среды, продолжительность
кормления, частота подкормок инфицированных блох, фенологический период, в кото-
рый проводится эксперимент, специфичность используемых лабораторных животных.
При экспериментальных работах в сибирских очагах чумы показано, что блохи этого
вида при подкормках на сусликах легко инфицируются и эффективно передают возбу-
дитель чумы другим зверькам (Воронова, 1984; Базанова, Маевский, 1996). Выявлено
наличие значительных сезонных изменений частоты блокирования (Воронова, 1978;
Базанова, 2009). В июле-начале августа, в период активизации эпизоотического про-
цесса в очаге, частота образования блока преджелудка у этого вида достигала макси-
мума и составляла 10.6 %. Срок от заражения блохи до появления первого блока в это
время сокращался до трех суток.
В организме C. tesquorum чумной микроб сохраняется длительное время. Ранее
(Евсеева, Фирсов, 1932) регистрировали сохранение возбудителя чумы в условиях ис-
кусственной норы на срок до 102 дней.
При проведении зимних раскопок гнезд длиннохвостого суслика в Тувинском при-
родном очаге чумы удалось дважды получить изоляты чумного микроба от блох, вы-
бранных из субстратов. В обоих случаях это были C. tesquorum (Вержуцкий и др.,
2003). В этой же работе упоминается, что среди культур возбудителя чумы, выделен-
ных в весенний период, подавляющее большинство получено тоже от блох этого вида.
В экспериментах, проведенных в Тувинском очаге в условиях, близких к естествен-
ным, показана способность блох, инфицированных возбудителем чумы, переживать
зимний период без контакта с прокормителем и передавать инфекцию на следующий
год другим зверькам. Более половины (50.0-64.3 %) насекомых сохраняют возбу-
дитель чумы 10-11 месяцев, а около 30 % блох - 13-15 месяцев. Отдельные особи,
в основном самки, могут быть носителями чумного микроба до 22 месяцев, который
затем может передаваться здоровым сусликам (Базанова, 2009). Формирование блока
в преджелудке блохи C. tesquorum регистрируется и в очень отдаленные после инфи-
цирования сроки - на 282-411-е сутки (Базанова, Маевский, 1996; Воронова, Базано-
193
ва, 2004). Отмечены значимые различия блох из разных популяций в эффективности
передачи инфекции (Базанова и др., 2000; Базанова, 2009). Анализ результатов много-
летних работ по изучению эффективности блох как переносчиков чумы показал, что
у C. tesquorum за последние десятилетия резко вырос уровень блокообразования. Дан-
ное явление отмечено во всех трех сибирских природных очагах чумы (Вержуцкий
и др., 2018).
Жизненный цикл этого вида в значительной мере определяется особенностями жиз-
недеятельности его основных прокормителей. На большей части ареала C. tesquorum
имеет 1 генерацию за сезон. Вышедшие из анабиоза имаго, а также появившиеся из
куколок молодые особи весной приступают к размножению. Летом происходит смена
поколений и нарастает волна выплода новых молодых насекомых. С середины августа
подавляющее большинство C. tesquorum прекращает питаться и остается в нежилых
гнездах летнего типа (Вержуцкий и др., 2009). Недавно выявлено, что у этого вида
в Тувинском природном очаге чумы в настоящее время развивается два поколения
за год (Галацевич, 2018). Автор связывает это с улучшением условий существования
преимагинальных стадий развития C. tesquorum в связи с изменениями климата в
Центральной Азии.
Большинство видов сусликов в неблагоприятные периоды времени впадает на бо-
лее или менее продолжительные периоды в спячку. Длиннохвостый суслик и блохи
C. tesquorum перезимовывают раздельно, в связи с этим чумной микроб около 8 меся-
цев в году находится в блохах без контакта с основным носителем (Вержуцкий и др.,
2003).
У большинства видов наземных беличьих в период беременности и выкармливания
молодняка отмечено формирование своеобразных временных группировок - агрега-
ций самок (Michener, 1983; King, 1989). Для длиннохвостого суслика показано, что
формирование агрегаций самок на небольших участках местности приводит к локаль-
ной концентрации гнезд выводкового типа и чрезвычайно высокой численности имаго
C. tesquorum на таких участках, примерно в 20 раз превышающей фоновую. Выявлено,
что участки, где в мае-июне формируются агрегации самок суслика и накапливается
аномально высокая численность блох (до нескольких тысяч особей на гектар), стано-
вятся оптимальными стациями для сохранения, накопления и распространения чумно-
го микроба (Вержуцкий, 1999).
ЗАКЛЮЧЕНИЕ
В Неарктической области очаги чумы расположены преимущественно в Западно-
американской подобласти, в Неотропической области - в Бразильской и Андийской
подобластях, в Афротропической области - в Мадагаскарской подобласти, на западе
Восточноафриканской и юге Капской. В Палеарктической области - это Туранская
провинция Турано-Иранской подобласти, Центральноазиатская и юго-восток Восточ-
ноазиатской подобластей. В Индо-Малайской области очаги ограничены западом Ин-
дийской подобласти, а также западом и востоком Индокитайской подобласти.
Проведенный анализ показывает значительное таксономическое разнообразие но-
сителей и переносчиков чумы. Виды блох, из которых в естественных условиях был
выделен возбудитель чумы, принадлежат к 40 % от общего числа родов, известных
194
в мировой фауне, и к более чем половине семейств. Данные цифры указывают на ши-
рокое распространение чумного микроба по биоценотическим цепям, что может об-
условить формирование новых очагов под воздействием, например, климатических
изменений. Однако лишь небольшое число видов блох и их хозяев в определенных
условиях обеспечивают устойчивую циркуляцию возбудителя, как в период эпизоотий,
так и в межэпизоотические периоды. При этом одним из наиболее важных и домини-
рующих переносчиков среди блох является Citellophilus tesquorum. Следует отметить,
что причины выявленных особенностей в жизнедеятельности блохи C. tesquorum еще
не вполне понятны. Возможно, что некоторые важные вопросы могут быть решены
при использовании молекулярно-генетических методов.
БЛАГОДАРНОСТИ
Работа выполнена на базе коллекции Зоологического института РАН (ЗИН РАН)
(УФК ЗИН рег. № 2-2.20) при поддержке Российского фонда фундаментальных иссле-
дований (грант № 19-04-00759).
СПИСОК ЛИТЕРАТУРЫ
Атлас распространения особо опасных инфекций в Республике Казахстан. 2012. Под ред. Л.А. Бурделова.
Алматы, 232 с.
Базанова Л.П. 2009. Взаимоотношения чумного микроба (Yersinia pestis) и блох (Siphonaptera) (на примере
сибирских природных очагов чумы). Автореф. дис
докт. биол. наук. Улан-Удэ, 46 с.
Базанова Л.П., Вержуцкий Д.Б., Хабаров А.В. 2000. Межпопуляционные различия во взаимоотношениях
с возбудителем чумы двух массовых видов блох длиннохвостого суслика из Тувы. В сб.: Карантинные
и зоонозные инфекции в Казахстане. Алматы, 2: 48-52.
Базанова Л.П., Маевский М.П. 1996. Длительность сохранения чумного микроба в организме блохи
Citellophilus tesquorum altaicus. Медицинская паразитология и паразитарные болезни 1: 45-48.
Балахонов С.В., Корзун В.М., Вержуцкий Д.Б., Чипанин Е. В., Михайлов Е.П., Денисов А.В., Глушков Э.А.,
Акимова И.С. 2014. Особенности эпизоотической активности горных природных очагов чумы Сиби-
ри в ХХI веке. Здоровье населения и среда обитания 12 (261): 48-50.
Бибикова В.А., Классовский Л.Н. 1974. Передача чумы блохами. М., Медицина, 188 с.
Ващенок В.С. 1988. Блохи - переносчики возбудителей болезней человека и животных. Л., Наука, 163 с.
Ващенок В.С. 1999. Роль блох (Siphonaptera) в эпизоотологии чумы. Паразитология 33 (3): 198-206.
Вержуцкий Д.Б. 1999. Эпизоотологическая роль популяционной организации населения блох длиннохвосто-
го суслика в Тувинском природном очаге чумы. Паразитология 33 (3): 242-249.
Вержуцкий Д.Б. 2018. Активизация природных очагов чумы в Центральной Азии: беспочвенные опасения
или реальная угроза. Природа Внутренней Азии 1 (6): 7-17.
Вержуцкий Д.Б., Ткаченко В.А., Попов В.В., Колосов В.М. 2003. О сохранении возбудителя чумы в Тувин-
ском природном очаге. Журнал инфекционной патологии 10 (4): 31-32.
Вержуцкий Д.Б., Чумакова Н.А., Галацевич Н.Ф., Ковалева Н.И. 2009. К экологии блохи Citellophilus
tesquorum Wagn., 1898 в Юго-Западной Туве. Байкальский зоологический журнал 1: 17-22.
Воронова Г.А. 1978. Ceratophyllus tesquorum altaicus Ioff, 1936, как основной переносчик и хранитель чум-
ного микроба в Тувинском природном очаге. В сб.: Эпидемиология и профилактика ООИ в МНР и
СССР. Улан-Батор, с. 152-155.
Воронова Г.А. 1984. Взаимоотношения возбудителя чумы с блохами грызунов и зайцеобразных в Тувинском
природном очаге чумы. Автореф. дис
канд. мед. наук. Саратов, 14 с.
Воронова Г.А., Базанова Л.П. 2004. Значение блох (Siphonaptera) разных видов в поддержании эпизоотий
чумы в сибирских природных очагах. Бюллетень ВСНЦ СО РАМН 2 (1): 58-65.
Галацевич Н.Ф. 2018. Динамика таксоценоза блох длиннохвостого суслика в Юго-Западной Туве и ее влия-
ние на эпизоотическую активность Каргинского мезоочага чумы. Автореф. дис. … канд. биол. наук.
Иркутск, 20 с.
Гончаров А.И., Плотникова Е П. 2010. Распределение по зоогеографическим областям родов и видов блох
семейства Pulicidae, из представителей которых в естественных условиях выделен возбудитель чумы
195
(Insecta, Siphonaptera). Международународный журнал прикладных и фундаментальных исследова-
ний 11: 25-26.
Гончаров А.И., Тохов Ю.М., Плотникова Е.П., Артюшина Ю. С. 2013. Список видов и подвидов блох, обна-
руженных зараженными возбудителем чумы в естественных условиях. Ставрополь, 34 с.
Громов И.М., Баранова Г.И. 1981. Каталог млекопитающих СССР. Л., Наука, 456 с.
Громов И.М., Ербаева М.А. 1995. Млекопитающие фауны России и сопредельных территорий. СПб., ЗИН
РАН, 472 с.
Евсеева В.Е., Фирсов И.П. 1932. Блохи как хранители чумного вируса в зимнее время. Вестник микробиоло-
гии, эпидемиологии и паразитологии 11 (4): 281-283.
Каримова Т.Ю., Неронов В.М. 2007. Природные очаги чумы Палеарктики. М., Наука, 199 с.
Каримова Т.Ю., Неронов В.М., Попов В.П. 2010. Развитие взглядов на природную очаговость чумы. Зооло-
гический журнал 89 (1): 71-78.
Котти Б.К. 2018. Каталог блох (Siphonaptera) фауны России и сопредельных стран. 2-е изд. Ставрополь, Изд-
во СКФУ, 128 с.
Медведев С.Г. 1998. Фауна и паразито-хозяинные связи блох (Siphonaptera) Палеарктики. Энтомологическое
обозрение 77 (2): 295-314.
Медведев С.Г. 2000. Фауна и паразито-хозяинные связи блох (Siphonaptera) различных зоогеографических
областей мира. II. Энтомологическое обозрение 79 (4): 812-830.
Медведев С.Г. 2002. Особенности распространения и паразито-хозяинных связей блох (Siphonaptera). I. Эн-
томологическое обозрение 81 (3): 737-753.
Медведев С.Г. 2013а. Таксономический состав и особенности фауны блох (Siphonaptera) России. Энтомоло-
гическое обозрение 92 (1): 85-101.
Медведев С.Г. 2013б. Палеарктические центры таксономического разнообразия отряда блох (Siphonaptera).
Энтомологическое обозрение 92 (3): 684-702.
Онищенко Г.Г., Кутырев В.В. 2004. Природные очаги чумы Кавказа, Прикаспия, Средней Азии и Сибири.
М., Медицина, 192 с.
Павлинов И.Я., Яхонтов Е.Л., Агаджанян А.К. 1995. Млекопитающие Евразии. I. Rodentia: систематико-гео-
графический справочник. М., Изд-во МГУ, 240 с.
Природные очаги чумы Кавказа, Прикаспия, Средней Азии и Сибири. 2004. Под ред. Г. Г. Онищенко, В. В.
Кутырева. М., Медицина, 192 с.
Ралль Ю.М. 1965. Природная очаговость и эпизоотология чумы. М., Медицина, 363 с.
Bertherat E. 2016. Plague around the world, 2010-2015. Weekly Epidemiological Record 91 (8): 89-104.
Dubyanskiy V.M., Yeszhanov A.B. 2016. Ecology of Yersinia pestis and the epidemiology of plague. In: Yang R.,
Anisimov A. (eds). Yersinia pestis: retrospective and perspective. Advances in Experimental Medicine and
Biology, 918: 101-170.
King J.W. 1989. Spacing of female kin in Columbian ground squirrels (Spermophilus columbianus). Canadian
Journal of Zoology 67: 91-95.
Lewis R.E. 1990. The Ceratophyllidae: currently accepted valid taxa (Insecta: Siphonaptera). Koenigstein, Koeltz
scientific books, 267 pp.
Lewis R.E. 2003. Siphonaptera. 15th ed. 62 pp.
Michener G.R. 1983. Kin identification, matriarchies and the evolution of sociality in ground-dwelling sciurids. In:
Eisenberg F., Kleiman D.G. (eds) Advances in the study of mammalian behavior, pp. 528-572.
The atlas of plague and its environment in the People’s Republic of China. 2000. Beijing, Science Press, 221 pp.
Traub R.E., Rothschild M., Haddow J.F. 1983. The Ceratophyllidae: key to the genera and host relationships, with
notes on their evolution, zoogeography and medical importance. Cambridge University Press, 288 pp.
Wilson D.E., Reeder D.M. 2005. Mammal species of the World: a taxonomic and geographic reference. 3rd ed.
Baltimore, Johns Hopkins University Press, 743 pp.
DIVERSITY OF FLEAS (SIPHONAPTERA), VECTORS OF PLAGUE
PATHOGENS: THE FLEA CITELLOPHILUS TESQUORUM (WAGNER, 1898),
PARASITE OF GROUND SQUIRRELS OF THE GENUS SPERMOPHILUS
S. G. Medvedev, B. K. Kotti, D. B. Verzhutsky
Key words: fleas, Siphonaptera, species vectors of plague pathogen, taxonomic diversity,
Citellophilus tesquorum.
196
SUMMARY
Plague foci cover significant territories of all the continents, excluding Australia and Antarctica. They
are distributed in eleven subregions of five zoogeographical provinces. Plague pathogen vectors include
representatives of 346 out of 5937 species (about 6 %), 144 out of 1258 genera, 33 out of 156 families
(about 20 %), and 9 out of 28 of orders (about 30 %) of mammals. Representatives of three rodent
families (Cricetidae, Muridae, and Sciuridae) manifest the largest fraction of plague vector species
(34 %).
Nowadays, 257 flea species were recorded as species infected with plague pathogen, constituting
12 % out of 2162 all the known flea species. Species infected with plague pathogen in nature belong
to 95 and 12 out of 240 genera and 18 flea families, respectively. This number constitutes 40 % of
the number of flea genera of the world fauna. These numbers point to the wide distribution of the
plague pathogen along biocenotic chains, stipulating the formation of new foci due to, for example,
and climatic changes.
At the same time, only significantly smaller number of fleas and their hosts in certain environmental
conditions provide a stable circulation of the pathogen, as well as during epizootics, and during inter-
epizootic periods. One of this species is represented by the flea Citellophilus tesquorum (Wagner,
1898), a parasite of ground squirrels of the genus Spermophilus.
This species is characterized by strongly developed ability to form the bacterial «plug» in the
proventriculus and the long survival of the plague pathogen in the organism of the imago. Fleas of this
species effectively transmit plague pathogen to warm-blooded hosts, retaining this ability even after
a long separate overwintering. C. tesquorum can reach very high population density in small areas
where ground squirrel females aggregate during pregnancy and milking of their offspring.
197