ПАРАЗИТОЛОГИЯ, 2019, том 53, № 6, с. 463-473.
УДК 595.421.(470.22)
ЧИСЛЕННОСТЬ ИКСОДОВЫХ КЛЕЩЕЙ (ACARI: IXODIDAE)
НА МЕЛКИХ МЛЕКОПИТАЮЩИХ В ЛЕСНЫХ БИОТОПАХ
СРЕДНЕТАЕЖНОЙ ПОДЗОНЫ КАРЕЛИИ
©2019 г. Л. А. Беспятоваa, *, С. В. Бугмыринa,
С. А. Кутенковa, И. А. Никонороваa
aИБ КарНЦ РАН, Пушкинская ул., д. 11, Петрозаводск 185910, Россия
*e-mail: gamasina@mail.ru
Поступила в редакцию 05.08.2019 г.
После доработки 14.08.2019 г.
Принята к публикации 28.08.2019 г.
Изучены видовой состав и численность иксодовых клещей на мелких млекопитающих в лес-
ных биотопах среднетаежной подзоны Карелии (д. Гомсельга, Кондопожский р-н, 62.07° с. ш.,
33.96° в. д.). Всего с 2011 по 2017 гг. было отработано 8 316 давилко-суток, отловлено 455 особей
зверьков, с которых собрано 889 особей клещей. Видовой состав иксодовых клещей с мелких
млекопитающих (3 вида мышевидных грызунов и 4 вида бурозубок) представлен двумя видами
клещей Ixodes persulcatus Schulze, 1930 и I. (Exopalpiger) trianguliceps Birula, 1895, с домини-
рованием на 59% I. persulcatus.Численность и размещение отдельных популяционных групп
(гемипопуляций) иксодовых клещей, паразитирующих на мелких млекопитающих, варьировали
в зависимости от типа биотопа. В более влажном биотопе - лиственном лесу у ручья - находит
более благоприятные условия для обитания клещ I. (E.) trianguliceps, а I. persulcatus предпочитает
дренированные и прогреваемые почвы, характерные для смешанного леса.
Ключевые слова: иксодовые клещи, Ixodes persulcatus, I. (Exopalpiger) trianguliceps, мелкие
млекопитающие, лесные биотопы, видовой состав, численность.
DOI: 10.1134/S0031184719060036
Иксодовые клещи (сем. Ixodidae) - временные эктопаразиты, специфические пере-
носчики многих инфекционных заболеваний вирусной, бактериальной и паразитарной
природы. В таежных биоценозах Карелии встречаются 5 видов иксодовых клещей, из
которых только 4 вида - Ixodes (Exopalpiger) trianguliceps Birula, 1895, I. ricinus (Lin-
naeus, 1758), I. apronophorus Schulze, 1924 и I. persulcatus Schulze, 1930 - паразитируют
на мелких млекопитающих (Бобровских, 1989). Республика Карелия относится к числу
эндемичных территорий по ряду клещевых инфекций. Уже на начальных этапах изуче-
ния иксодовых клещей они вошли в ряд объектов первоочередного исследования как
переносчиков наиболее острой природноочаговой трансмиссивной вирусной нейроин-
фекции - клещевого энцефалита, повышение напряженности и появление новых очагов
463
которой отмечено в последние десятилетия (Государственный доклад …., 2006-2017;
Беспятова, Бугмырин, 2017). Важнейшим фактором, определяющим потенциальную
эпидемиологическую опасность, границы и структуру ареала клещевого энцефалита,
является таёжный клещ I. persulcatus, рост численности и расширение границ обитания
которого были установлены нами по всей среднетаежной подзоне Карелии (Беспятова
и др., 2006; 2009; Bugmyrin et al., 2013; Bugmyrin et al., 2019).
Изучение функционирования природных очагов клещевых инфекций, начатое в 50-е
годы прошлого столетия, приобретает особую актуальность на современном этапе -
в условиях усиливающегося антропогенного пресса на природные экосистемы.
В основе решения этой задачи стоит определение видового состава и границ распростра-
нения, а также экологических особенностей развития иксодовых клещей, обитающих
на различных территориях естественных и трансформированных таежных экосистем.
В формировании, развитии и динамике природных очагов клещевых инфекций особая
роль принадлежит и активным предимагинальным фазам развития клещей - личинкам
и нимфам.
Целью данного исследования является изучение видового состава, численности
и биотопического размещения отдельных фаз развития (гемипопуляций) иксодовых
клещей, паразитирующих на мелких млекопитающих в лесных биотопах среднетаеж-
ной подзоны Карелии.
МАТЕРИАЛЫ И МЕТОДЫ
Исследования проведены на протяжении семи лет с 2011 по 2017 г. на базе научного стацио-
нара Института биологии КарНЦ РАН в окрестностях д. М. Гомсельга (62.07° с.ш., 33.96° в. д.)
в Кондопожском р-не. Выбранная точка является наиболее типичной и характерной для обитания
иксодовых клещей в пределах среднетаежной подзоны Карелии. Преобладающими коренными
суходольными лесами окрестностей стационара являются сосняки и, в меньшей степени, ель-
ники. Основными типами лесорастительных условий являются черничные (Крышень, 2010).
В настоящий момент значительную часть территории занимают вырубки различной давности,
а также развивающиеся на их месте молодняки и производные леса (Гусева и др., 2014).
Материал собран в июне-сентябре в трех лесных биотопах: линия № 1 - по границе сосново-
березового травяно-черничного леса и разнотравно-злакового луга (экотон); линия № 2 - осиново-
березовый травяной приручейный лес в распадке (лиственный лес); линия № 3 - сосново-
березовый травяной лес на пологом склоне (смешанный лес).
В ходе полевых и лабораторных исследований использованы стандартные зоологические,
паразитологические и геоботанические методы исследования.
Геоботанические. Геоботанические описания проведены с использованием стандартных,
принятых в отечественной геоботанике методов (Дылис, 1974). Всего заложено 8 площадок:
2 по линии № 1, в лесу и на лугу; 5 вдоль линии № 2, на различных по составу древостоя и по-
ложению в долине ручья участках; 1 на линии № 3 в достаточно однородных условиях склона.
Значение экологических факторов среды рассчитано на основе экошкал Эленберга (Ellenberg
et al., 1991) методом средневзвешенной середины интервала в программе Ecoscale for Windows
(Грохлина, Ханина, 2006). Для расчета значения фактора освещенности использованы данные
по проективному покрытию наземных ярусов, остальных факторов - также яруса подроста
и подлеска. Все использованные шкалы девятибалльные, за исключением влажности, включаю-
щей, помимо 9 градаций для наземных, 3 градации для водных местообитаний.
464
Зоологические. Отлов мелких млекопитающих проведен с помощью давилок трапиковых
и Геро, выставленных в линии по 25-50 штук на 3-5 дней в трех биотопах. Относительную
численность выражали числом особей на 100 давилко-суток. Всего за 7 лет отработано 8 316
давилко-суток, отловлено 455 особей зверьков, с которых собрано 889 экз. клещей. Единствен-
ный раз в давилку была поймана белка обыкновенная (линия № 2), с которой собраны 2 нимфы
клеща I. persulcatus. В связи с тем, что добыча была случайной, собранные клещи не учитывали
при сравнительном анализе материала.
Паразитологические. Сбор иксодовых клещей с мелких млекопитающих проведен по стан-
дартной методике (Беспятова, Бугмырин, 2012). Для оценки относительной численности клещей
использовали общепринятые в экологической паразитологии индексы: индекс встречаемости
(доля особей хозяев, у которых обнаружен клещ, %), индекс обилия (среднее число особей одного
вида клеща на особь хозяина, экз.), показатель прокормления (произведение индекса обилия на
относительную численность хозяина), индекса доминирования (доля особей вида по отношению
к суммарному количеству особей всех сравниваемых видов, %) (Беклемишев, 1961). Для оценки
значимости межбиотопических различий численности иксодовых клещей были использованы
критерии Краскела-Уолиса и Манна-Уитни, анализ данных выполнен в программ Past (Hammer
et al., 2001). Согласование наблюдаемого распределения численности клещей с законом негатив-
ного бинома рассчитано в программе Quantitative Parasitology 3.0 (Rózsa et al., 2000).
Видовая идентификация иксодовых клещей проводилась на микроскопе Olympus CX 41
c цифровым модулем визуализации и документирования VIDI-GAM - оборудовании Центра
коллективного пользования Федерального исследовательского центра «Карельский научный
центр Российской академии наук».
РЕЗУЛЬТАТЫ
Анализ широтной структуры ценофлор лесных биотопов показывает, что изученные
участки имеют более «южные» черты, не характерные для основной территории средне-
таежной Карелии. Суходольные вторичные леса развились на месте экстразональных
сосняков вейниковых (травяно-зеленомошных), в которых травы играют более значи-
тельную роль, чем бореальные лесные кустарнички (Кучеров, 2016).
Экологические факторы среды, рассчитанные на основе экошкал Эленберга (Ellenberg
et al., 1991), показали, что по значению величины богатства почвы минеральным азотом,
отражающей общий запас питательных веществ в почве, наиболее бедным биотопом
является лесной участок у линии № 1, при этом луговое местообитание - наиболее
богатое в отношении насыщенности почвы доступными минеральными элемента-
ми. Линии № 2 и № 3 умеренно обеспеченные, занимают промежуточное положение
(табл. 1). В отношении реакции (кислотности) почвы местообитания распределились
сходным с богатством почвы азотом образом. Почвы наиболее бедных местооби-
таний (лес по линии № 1) оказались и наиболее кислыми, реакция на лугу близка
к нейтральной, линии № 2 и № 3 с умеренно кислыми почвами занимают промежуточ-
ное положение. В отношении влажности почв линии № 1 и № 3 относятся к свежим
(средне-влажным), линия № 2 - от свежих до влажных местообитаний. Фактор осве-
щенности также варьирует. Наиболее тенистыми условиями характеризуются участки
с обилием ольхи серой у ручья по линии № 2, достаточно освещенные - луг по линии
№ 1, остальные местообитания занимают промежуточное положение.
465
Таблица 1. Значения экологических факторов для трех исследованных биотопов
Линия № 1
Фактор
Линия № 2
Линия № 3
Леc
Луг
Богатство почвы минеральным азотом
4.7
3
6.3
5.1
[4.3-5.1]
Кислотность почвы
5.1
3.3
5.8
4.8
[4.5-5.6]
Влажность почвы
6.7
5.3
5.1
5.7
[6.3-7.2]
Освещенность
5.2
5.3
6.8
5.5
[4.2-5.9]
П ри меч ани я. Для Линии №2 приведены: среднее (по 5 описаниям), минимальное и макси-
мальное значения фактора.
В обследованных биотопах было отмечено семь видов мелких млекопитающих, из
которых 3 вида - мышевидные грызуны и 4 вида - бурозубки (табл. 2). В суммарных
уловах почти в равных соотношениях доминировали два вида - европейская рыжая по-
левка (Myodes glareolus Schreb.) - 48.6% и обыкновенная бурозубка (Sorex araneus L.) -
42.9%. Эти виды были массовыми во всех биотопах с небольшими колебаниями
по степени доминирования на линиях. Так, M. glareolus преобладала в смешанном
лесу (ИД 58.7%), а S. araneus - в экотонной зоне на границе леса и луга (ИД 51.6%),
в лиственном лесу незначительно доминировала M. glareolus. (табл. 2). По мере воз-
растания численности мелких млекопитающих, биотопы расположились в следующем
порядке - экотонная зона на границе леса и луга, смешанный и лиственный леса.
В разных биотопах численность массовых видов варьировала: полевка M. glareolus
преобладала в смешанном и лиственном лесах, составляя 3.6 и 3.5 экз. на 100 лов-сут.
соответственно, а S. araneus - в экотонной зоне - 2.5 на 100 лов-сут.
По результатам исследований установлено, что на мелких млекопитающих, оби-
тающих в трех биотопах, прокармливались личинки и нимфы I. persulcatus, а также
личинки, нимфы и взрослые (самки и самцы) I. trianguliceps (табл. 3, 4). Клещ I. per-
sulcatus был отмечен только на личиночной фазе у малой и средней бурозубок, на
личиночной и нимфальной фазах - на всех остальных видах. Клещ I. trianguliceps на
всех активных фазах развития был собран с рыжей и пашенной полевках, на личи-
ночной и нимфальной фазах - с обыкновенной и равнозубой, только на личиночной
фазе - с малой и средней бурозубок.
В общих сборах с мелких млекопитающих в видовом составе клещей преобладал
таежный клещ с ИД 59% (табл. 4). Степень доминирования (суммарная доля особей
всех активных фаз развития) варьировала по биотопам у I. persulcatus в интервале
50-69%, с максимальным значением в смешанном лесу, у клеща I. trianguliceps - 32-
50% с максимумом в экотонной зоне у луга (табл. 4). На рыжей полевке доминировал
I. persulcatus с колебанием по биотопам от 61 до 72%, с максимумом в смешанном
лесу. На обыкновенной бурозубке соотношение двух видов клещей было почти равным
в лиственном и смешанном лесах, исключение составила экотонная зона, где числен-
ность I. trianguliceps превысила почти в два раза I. persulcatus (табл. 4).
466
Таблица 2. Видовой состав и численность мелких млекопитающих в лесных биотопах
Биотопы
ИД,
Вид
№ 1
№ 2
№ 3
%
1
2
3
1
2
3
1
2
3
Sorex araneus L.
42.9
63
51.6
2.5
76
41.5
2.9
56
37.3
2.8
S. isodon L.
2.4
0
0
0
10
5.5
0.4
1
0.7
0.05
S. minutus L.
1.3
5
4.1
0.2
1
0.5
0.03
0
0
0
S. caecutiens Laxm.
0.9
3
2.5
0.1
1
0.5
0.03
0
0
0
Sicista betulina Pall.
0.4
1
0.8
0.04
1
0.5
0.03
0
0
0
Myodes glareolus Sch.
48.6
45
36.9
1.7
88
48.1
3.5
88
58.7
3.6
Microtus agrestis L.
3.5
5
4.1
0.2
6
3.3
0.22
5
3.3
0.2
∑455
100
122
100
4.7
183
100
7.3
150
100
6.8
П ри меч ан ия. № 1 линия по границе сосново-березового травяно-черничного леса и
разнотравно-злакового луга, № 2 линия осиново-березовый травяной приручейный лес в рас-
падке, № 3 линия сосново-березовый травяной лес на пологом склоне.
1 - число отловленных зверьков (экз.); 2 - доля вида в суммарных отловах мелких млекопитаю-
щих, %; 3 - относительная численность мелких млекопитающих (экз. на 100 ловушко-суток).
Таблица 3. Роль доминирующих видов мелких млекопитающих в прокармливании клещей
в лесных биотопах
№ 1
№ 2
№ 3
Суммарные
Фаза развития
Sa
Mg
Sa
Mg
Sa
Mg
Sa
Mg
Ixodes trianguliceps
Личинка
55.6
31.1
30.2
42.6
28.9
64.5
34.4
47.2
Нимфа
35.7
57.1
17.2
63.8
13.5
83.8
18.3
69.7
Имаго
0
1 из 1
0
4 из 5
0
2 из 2
0
7 из 8
∑ (личинка, нимфа, имаго)
50.0
38.3
25.5
50.0
23.5
72.0
28.9
54.2
Ixodes persulcatus
Личинка
26.9
57.7
23.5
74.3
14.3
84.2
19.7
76.8
Нимфа
2 из 8
6 из 8
2.9
80.0
5.6
92.6
5.2
86.6
∑ (личинка, нимфа)
26.7
60.0
20.1
75.2
12.4
86.0
17.2
78.6
П рим еч ани я. Доля особей каждой фазы развития клещей от общего количества особей на
Sorex araneus (Sa) и Myodes glareolus (Mg); № 1, 2, 3 номера биотопов, которые соответствуют
номерам биотопов в табл. 1 и 2.
Таблица 4. Видовой состав иксодовых клещей на мелких млекопитающих в лесных биотопах
№ 1
№ 2
№ 3
Суммарные
Вид
1
2
3
1
2
3
1
2
3
1
2
3
Ixodes
65.2
39
50
53.3
37.4
47.3
46.6
27.6
31.5
54.1
32.5
41.2
trianguliceps
Ixodes
34.8
61
50
46.7
62.6
52.7
53.4
72.4
68.5
45.9
67.5
58.8
persulcatus
Пр им еч ания. Доля личинок и нимф I. persulcatus, а также личинок, нимф и имаго
I. trianguliceps, %. 1 - на Sorex araneus, 2 - на Myodes glareolus, 3 - на всех мелких млеко-
питающих.
467
Анализ роли основных хозяев в прокармливании клещей и характера их распреде-
ления по биотопам проведен для каждой активной фазы жизненного цикла. Массовые
виды рыжая полевка и обыкновенная бурозубка прокармливали суммарно по всем
линиям около 96% личинок и нимф I. persulcatus и около 85% личинок, нимф и има-
го I. trianguliceps. Большую часть особей активных фаз развития как таежного клеща
около 79%, так и I. trianguliceps около 55%, прокармливала полевка.
Основную роль в прокармливании и личинок (около 77%) и нимф (87%) клеща
I. persulcatus по всем биотопам играла рыжая полевка (табл. 3). В разных биотопах
степень доминирования в прокармливании как личинок, так и нимф таежного клеща
полевкой была выше 50%, достигнув максимальных значений в смешанном лесу - 84%
личинок и 93% нимф. Что касается I. trianguliceps, рыжая полевка в целом также имела
лидирующие позиции, в большей мере, по взрослым (почти 90%) и нимфам (почти
70%), в меньшей - по личинкам (около 50%), прокармливая в сумме более 50% осо-
бей трех активных фаз развития клеща. Максимальные значения по прокармливанию
нимф (84%) и личинок (65%) отмечены у рыжей полевки в смешанном лесу, а взрос-
лых (4 из 5) - в лиственном лесу (табл. 3). Прокармливание взрослой фазы развития
I. trianguliceps отмечено только у рыжей полевки (7 экз. самки) во всех биотопах и на
пашенной полевкой (1экз. самец) в лиственном лесу. Обыкновенная бурозубка лиди-
ровала в прокармливании только личинок клеща (около 56%) в экотонной зоне луга.
Показатели прокормления и встречаемости двух видов клещей на разных фазах раз-
вития варьировали по биотопам и проанализированы как в целом на всех мелких мле-
копитающих, так и на доминирующих видах в каждом конкретном биотопе (табл. 5).
Наиболее высокими эти показатели были у I. persulcatus для личинок и нимф на всех
мелких млекопитающих и на рыжей полевке в смешанном лесу, минимальными - для
личинок на обыкновенной бурозубке в экотонной зоне на границе луга и леса, для
нимф - в лиственном лесу. Показатели численности и встречаемости всех активных
предимаго клеща I. trianguliceps, уступали таковым I. persulcatus. Наиболее высо-
кими эти показатели для всех активных фаз развития клеща были в лиственном лесу
(табл. 5). Личинки и нимфы клеща доминировали на рыжей полевке в смешанном
лесу, c минимумом в экотонной зоне. Встречаемость (суммарная) всех фаз развития
была максимальной у рыжей полевки в лиственном лесу, а минимальной - личинок
у рыжей полевки в экотонной зоне, нимф - у обыкновенной бурозубки в смешанном
лесу.
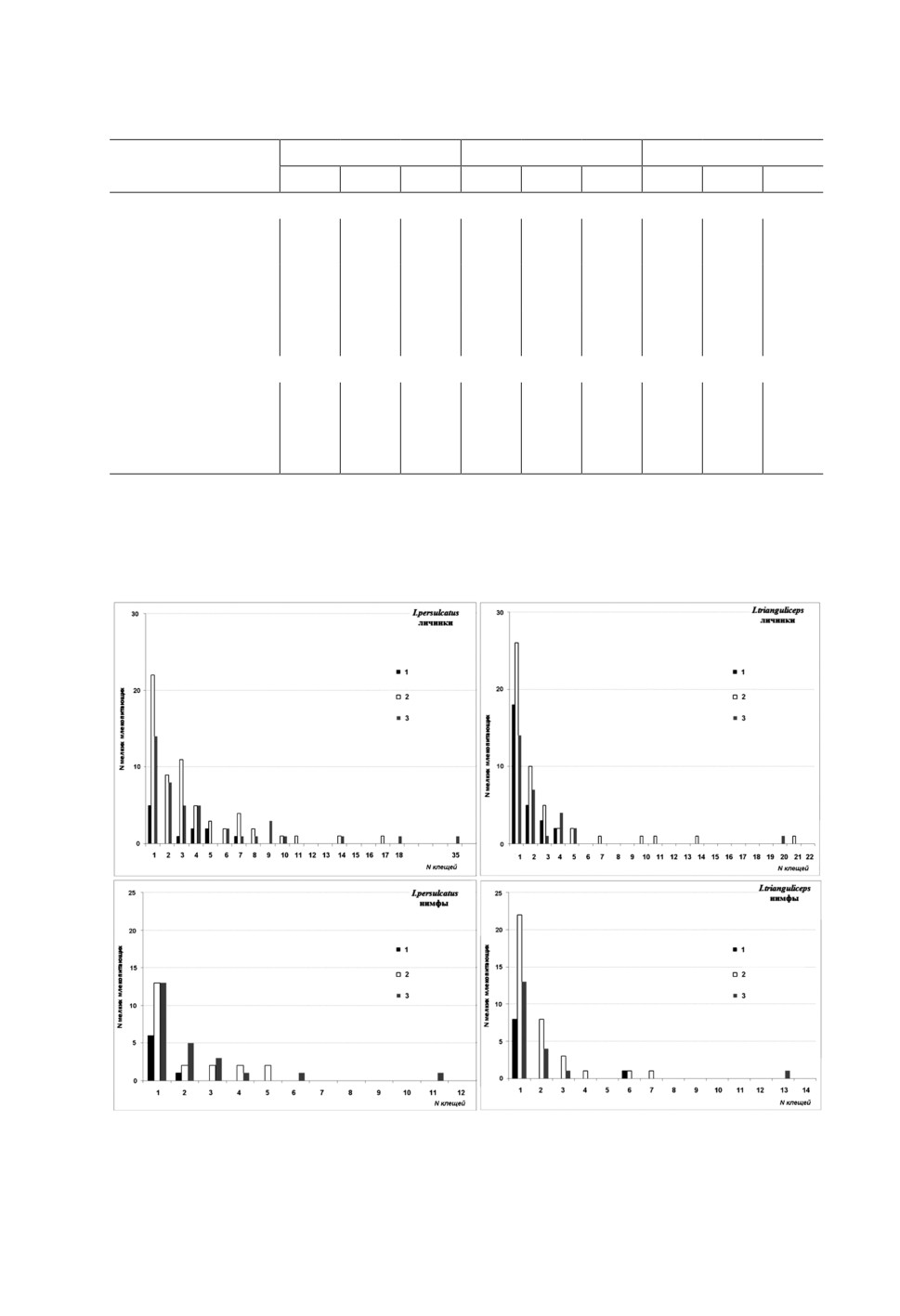
Распределение гемипопуляций иксодовых клещей в рассмотренных биотопах было
агрегированным и согласовывалось с моделью негативно биномиального распреде-
ления (рис. 1). Особенности биотопа вносят существенный вклад в наблюдаемую
биотопическую изменчивость показателей численности I. persulcatus, как личинок
(χ2 11.3, p<0.001), так и нимф (χ2 2.6, p≤0.01). Основные наблюдаемые различия, глав-
ным образом, определяются низкой численностью I. persulcatus в экотонной зоне по
сравнению со смешанным и лиственным лесом. У I. trianguliceps межбиотопические
различия мене выражены и получены только для нимф (χ2 2.6, p<0.05). В свою очередь,
значимые межбиотопические отличия показателя прокормления (критерий Манна -
Уитни) иксодовых клещей на мелких млекопитающих получены только при сравнении
численности нимф в экотонной зоне c биотопами смешанный лес для I. persulcatus
и лиственный лес для I. trianguliceps.
468
Таблица 5. Относительная численность (индекс прокормления и встречаемость) иксодовых
клещей на мелких млекопитающих в разных биотопах
№ 1
№ 2
№ 3
Фаза развития
Mg
Sa
ММ
Mg
Sa
ММ
Mg
Sa
ММ
Ixodes trianguliceps
Личинка
0.8
1.4
2.1
2.5
2.3
6.3
3.3
1.1
4.8
11.1
23.8
19.7
28.4
15.8
21.7
17.0
23.2
19.3
Нимфа
0.2
0.3
0.5
1.4
0.6
4.4
3.1
0.3
1.9
8.9
7.9
7.3
22.7
9.2
17.4
15.9
7.1
12.7
Имаго
0.02
0.01
0.08
0.12
0.05
0.02
0
0
0
2.2
0.8
4.5
2.7
2.3
1.3
Ixodes persulcatus
Личинка
1.3
0.4
2.0
6.4
1.3
8.1
6.9
1.2
8.3
11.1
7.9
9.8
31.8
17.1
23.9
38.6
19.6
30.0
Нимфа
0.32
0.09
0.43
1.62
0.06
1.86
1.85
0.29
2.19
13.3
1.6
5.7
13.6
1.3
8.7
25.0
1.8
16.0
П ри мечан ия. Над чертой - индекс прокормления, под чертой - встречаемость, %; Mg -
Myodes glareolus, Sa - Sorex araneus, ММ - мелкие млекопитающие.
Рис. 1. Распределение численности личинок и нимф клещей Ixodes persulcatus и I. trianguliceps
на мелких млекопитающих в лесных биотопах (1, 2, 3)
469
ОБСУЖДЕНИЕ
Основным направлением современных исследований является изучение фактиче-
ских изменений, произошедших в последнее время в распространении и численности
наиболее важного компонента таежных биоценозов - иксодовых клещей. Получение
целостной картины пространственной структуры популяций иксодовых клещей как
основных, так и второстепенных переносчиков в очагах клещевого энцефалита в Ка-
релии складывается в изучении размещения каждого вида переносчика в пределах
отдельных небольших по размерам сообществ. В этой связи существенным аспектом
является исследование факторов, влияющих на это размещение. Лесовосстановление
после вырубок происходит через смену древесных пород и изменение мощности лесных
подстилок. Поэтому именно вырубки определяют тренды трансформации местообита-
ний таежных животных, как обитателей лесной подстилки иксодовых клещей, так и
хозяев мелких млекопитающих. Ранее нами был проанализирован материал по обилию
и встречаемости активных фаз развития клещей I. persulcatus и I. trianguliceps с мелких
млекопитающих на разных этапах сукцессии леса после рубки (Бугмырин и др., 2009;
Bespyatova, Bugmyrin, 2015).
Стационарные исследования показали, что в районе исследования микроклимати-
ческие условия в целом близки к оптимальным для развития клещей. Богатые (менее
кислые более «теплые») почвы определяют и высокую численность клещей здесь.
В типичных лесных стациях модельного полигона на мелких млекопитающих про-
кармливались личинки и нимфы таежного клеща I. persulcatus с пастбищным типом
паразитизма, а так же личинки, нимфы и имаго клеща I. trianguliceps c примитивным
пастбищным типом паразитизма. В видовом составе клещей преобладал I. persulcatus
с доминированием до 60%. Отсутствие у личинок и нимф этих видов строгой спец-
ифичности к виду хозяина ведет к их перераспределению на виды, преобладающие
в биотопе.
Сравнительный анализ разных биотопов по встречаемости и обилию личинок и
нимф иксодовых клещей на мелких млекопитающих в разных биотопов показал, что
уровень численности каждой из активных фаз развития, а также роль отдельных видов
прокормителей различаются и обусловлены в том числе и особенностями биотопов.
В расположенном на склоне смешанном относительно сухом лесу, наиболее богатом
по общему запасу питательных веществ в почве и по составу растительности, со зна-
чительным участием бореонеморальной (южной) фракции, количественно преобладал
I. persulcatus. Oсновным прокормителем всех активных фаз развития двух видов клещей
была рыжая полевка
В лиственном лесу у ручья, чуть менее богатом по запасу питательных веществ
в почве, с широким спектром влажности и кислотности почв при уклоне к влажным
местообитаниям оба вида иксодид были примерно в равном соотношении с незна-
чительным преобладанием таежного клеща. Рыжая полевка была основным про-
кормителем личинок и нимф I. persulcatus, а также нимф и взрослых I. trianguliceps,
в прокормлении личинок последнего важную роль играла и обыкновенная бурозубка.
В экотонном биотопе на границе луга и леса, относительно сухого и бедного как по
запасу питательных веществ в почве, так и с более бореальными (северными) чертами
растительности, виды клещей на мелких млекопитающих представлены в равном со-
470
отношении. Численность каждой из фаз развития двух видов клещей была минималь-
ной относительно двух других биотопов. Основную роль в прокармливании личинок
и нимф I. persulcatus, а также взрослых особей клеща I. trianguliceps выполняла ры-
жая полевка, обыкновенная бурозубка лидировала в прокармливании только личинок
клеща I. trianguliceps.
Таким образом, наиболее благоприятные условия для обитания личинок и нимф
таежного клеща складывались в смешанном и лиственном лесах, где доминировала
рыжая полевка, прокармливая их большую часть. В экотонной зоне на границе леса
и луга, где численность рыжей полевки была невысокой, численность и личинок,
и нимф была выше у более массового вида - обыкновенной бурозубки. При этом
наиболее благоприятные условия для I. persulcatus складывались все-таки в смешанном
лесу, где доминирующие позиции по отношению к общей численности мелких млеко-
питающих имела рыжая полевка, которая прокармливала значительную долю личинок
и нимф клеща. Основываясь на численности активных предимаго I. persulcatus, можно
утверждать, что рыжая полевка в лесных биотопах является основным прокормителем
личинок и нимф, а ее численность определяет численность их гемипопуляций.
Наиболее благоприятные условия для клеща I. trianguliceps складывались в листвен-
ном лесу, где численность каждой активной фазы развития на всех хозяевах была
максимальной. В лиственном и смешанном лесах, где доминировала рыжая полевка,
она и прокармливала большую часть всех активных фаз развития клеща. В экотонной
зоне численность клеща на личиночной и нимфальной фазах была выше на массовой
здесь обыкновенной бурозубке. Роль обыкновенной бурозубки сводится к прокармли-
ванию в основном личинок I. trianguliceps, а соотношение доминирующих видов хозяев
в биотопе определяет роль каждого из них в поддержание численности личиночной
фазы развития клеща.
Мозаичное распределение иксодовых клещей связано с тем, что они могут раз-
виваться лишь в лесах определенного типа, при условиях умеренной температуры
и влажности, при наличии хорошо развитого травяного покрова, лесной подстилки
и хорошего дренажа почвы (Хейсин, 1953; 1954 а, б; Хейсин, Лебешева, 1955; Хейсин,
Кузнецова, 1956; Лутта и др., 1958). Анализ характера распределения клещей, про-
веденный для разных активных фаз жизненного цикла клещей I. persulcatus (личинок
и нимф) и I. trianguliceps (личинок, нимф и имаго), показал несовпадение мест их
высокой численности. Мозаичность распространения I. trianguliceps усиливается его
выраженной гигрофильностью, который для своего развития нуждается в высокой влаж-
ности, что доказано экспериментально (Кузнецова, 1964; Бобровских, 1965). Поэтому
в более влажном биотопе - лиственном лесу у ручья - этот клещ находит оптимальные
условия для своего обитания, а I. persulcatus предпочитает дренированные и прогре-
ваемые почвы, характерные для смешанного леса.
Таким образом, численность, видовой состав и размещение отдельных популяци-
онных групп двух видов иксодовых клещей I. persulcatus и I. trianguliceps, паразити-
рующих на мелких млекопитающих, варьируют в лесных биотопах. Эти колебания
обусловлены в основном особенностями местообитаний, определяющими как состав и
численность иксодовых клещей на разных фазах их развития, так и их прокормителей.
471
БЛАГОДАРНОСТИ
Финансовое обеспечение исследований осуществлялось из средств федерального
бюджета на выполнение государственного задания КарНЦ РАН (№ 0218-2019-0075)
и гранта РФФИ (№ 16-44-100109 р_а).
СПИСОК ЛИТЕРАТУРЫ
Беклемишев В.Н. 1961. Термины и понятия, необходимые при количественном изучении популяций экто-
паразитов и нидиколов. Зоологический журнал 40 (2): 149-158.
Беспятова Л.А., Бугмырин С.В. 2012. Иксодовые клещи Карелии (распространение, экология, клещевые
инфекции). Петрозаводск, Карельский научный центр РАН, 100 с.
Беспятова Л.А., Бугмырин С.В. 2017. Видовой состав, распространение основных переносчиков и эпи-
демиологическая ситуация по клещевому энцефалиту в Республике Карелия. Актуальные вопросы
ветеринарной биологии (1): 13-20.
Беспятова Л.А Бугмырин С.В., Коротков Ю.С., Иешко Е.П. 2009. Природные очаги клещевого энцефалита
на северо-западной периферии обитания таежного клеща (Ixodes persulcatus Schulze,1930). Труды
Карельского Научного центра РАН (4): 96-101.
Беспятова Л.А., Иешко Е.П., Ивантер Э.В., Бугмырин С.В. 2006. Межгодовая динамика численности ик-
содовых клещей и формирование очага клещевого энцефалита в условиях средней тайги. Экология
37 (5): 360-364.
Бобровских Т. К. 1965. Биология и экология Ixodes trianguliceps в Карелии. Автореф. дис. … канд. биол.
наук. Петрозаводск, 23 с.
Бобровских Т.К. 1989. Иксодовые клещи (подсемейство Ixodinae) Карелии. Петрозаводск, Карельский фи-
лиал АН СССР, 85 с.
Бугмырин С.В., Беспятова Л.А, Аниканова В.С., Иешко Е.П. 2009. Численность личинок и нимф Ixodes
persulcatus (Acari: Ixodidae) у мелких млекопитающих на вырубках среднетаежной подзоны Карелии.
Паразитология 43 (4): 338-346.
Государственный доклад о состоянии окружающей природной среды Республики Карелия в 2006-2017 г.г.
Петрозаводск, «Карелия» 1997-2018
Грохлина Т.И., Ханина Л.Г. 2006. Автоматизация обработки геоботанических описаний по экологическим
шкалам. В кн.: Принципы и способы сохранения биоразнообразия: сборник материалов II Всероссий-
ской научной конференции. Йошкар-Ола, Марийский государственный университет, 87-89.
Гусева Т.Л., Коросов А.В., Беспятова Л.А., Аниканова В.С. 2014. Многолетняя динамика биотопического
размещения обыкновенной бурозубки (Sorex araneus Linnaeus, 1758) в мозаичных ландшафтах Ка-
релии. Ученые записки Петрозаводского государственного университета. Серия: Естественные и
технические науки (8/2): 13-20.
Дылис Н.В. (ред.). 1974. Программа и методика биогеоценологических исследований. М., Наука, 403 с.
Крышень А.М. 2010. Типы лесорастительных условий на автоморфных почвах в Карелии. Ботанический
журнал 95 (3): 281-297.
Кузнецова Т.К. 1964. Материалы к развитию Ixodes trianguliceps Bir. в лабораторных условиях. В кн.: Лутта
А.С. (ред.). К природной очаговости паразитарных и трансмиссивных заболеваний в Карелии. М.- Л.,
Наука, 79-83.
Кучеров И.Б. 2016. О подразделении типов ареалов полизональных и плюрирегиональных видов для целей
сопряженного анализа флор сосудистых растений, мохообразных и лишайников. Комаровские чтения,
64: 138-197.
Лутта А.С., Хейсин Е.М. Шульман Р.Е. 1958. Влияние микроклиматических условий луга и лесана выжи-
ваемость и активность клещей Ixodes ricinus L. Зоологический журнал 37 (12): 1813-1822
Хейсин Е.М. 1953. Поведение взрослых Ixodes persulcatus в зависимости от температуры и влажности
окружающей среды. Зоологический журнал 32 (1): 77-87.
472
Хейсин Е.М. 1954а. Наблюдения над развитием скотского и таежного клещей в лабораторных условиях.
Труды Карело-Финского университета 5 (3): 88-106.
Хейсин Е.М. 1954б. Продолжительность развития личинок и нимф Ixodes ricinus L. и Ixodes persulcatus P.
Sch. в разные сезоны года (К вопросу о диапаузе). Труды Карело-Финского университета 6: 28-44.
Хейсин Е.М., Кузнецова Т.К. 1956. Холодостойкость яиц личинок и взрослых клещей Ixodes ricinus L. и
Ixodes persulcatus P. Sch. Труды Карело-Финского филиала АН СССР 4: 116-130.
Хейсин Е.М, Лебешева М.А. 1955. Яйцекладка и развитие Ixodes ricinus L. и Ixodes persulcatus P. Sch. при
разной температуре и влажности окружающей среды. Труды Карело-Финского университета 6: 5-27.
Bespyatova L.А., Bugmyrin S.V. 2015. Ixodid ticks (Parasitiformes: Ixodidae) from small mammals in reforesting
boreal habitats of the Northern European Russia. Entomological Review 95 (9): 1308-1316. DOI: 10.1134/
S0013873815090183
Bugmyrin S.V., Bespyatova L.A., Korotkov Yu.S. 2019. Long-term dynamics of Ixodes persulcatus (Acari: Ixodidae)
abundance in the north-west of its range (Karelia, Russia). Experimental and Applied Acarology 77 (2):
Bugmyrin S.V., Bespyatova L.A., Korotkov Y.S., Burenkova L.A., Belova O.A., Romanova L.Iu., Kozlovskaya L.I.,
Karganova G.G., Ieshko E.P. 2013. Distribution of Ixodes ricinus and I. persulcatus ticks in southern Karelia
Ellenberg H., Weber H.E., Dull R., Wirth V., Werner W., Paulisen D. 1991. Zeigerwerte von Pflanzen in Mit-
teleuropa [Indicator values of plants in Central Europe]. Scripta Geobotanics 18. Gottingen, Verlag Erich
Goltze KG, 248 pp.
Hammer O., Harper D., Ryan P. 2001. PAST: Paleontological statistics software package for education and data
analysis. Palaeontologia Electronica 4 (1): 9.
Rózsa L., Reiczigel J., Majoros G. 2000. Quantifying parasites in samples of hosts. The Journal of Parasitology
86 (2): 228-232.
THE ABUNDANCE IXODID TICKS (ACARI: IXODIDAE) ON SMALL MAMMALS
IN FOREST BIOTOPES OF THE MIDDLE TAIGA SUBZONE OF KARELIA
L. A. Bespyatova, S. V. Bugmyrin, S. A. Kutenkov, I. A. Nikonorova
Keywords: ixodid ticks, Ixodes persulcatus, I. (Exopalpiger) trianguliceps, small mam-
mals, forest biotopes, species composition, abundance.
SUMMARY
Species composition and abundance of ixodid ticks on small mammals in forest biotopes of the
middle taiga subzone of Karelia (Vlg. Gomselga, Kondopozhsky District, 62.07° N, 33.96° E, were
examined. In 2011-2017, 455 specimens of small mammals were collected with the use of span traps
(8316 traps/days). 889 tick specimens were collected from these mammals. Species composition of
ixodid ticks collected from seven species of small mammals (3 rodent species and 4 species of shrews)
was represented by two tick species Ixodes persulcatus Schulze, 1930 and I. (Exopalpiger) trianguliceps
Birula, 1895, with the dominance of 59% I. persulcatus. The number and location of individual popula-
tion groups (hemipopulations) of ixodid ticks feeding on small mammals varied depending on the type
of the biotope. More humid biotope, the deciduous forest near a stream, was more optimal for I. (E.)
trianguliceps, while I. persulcatus preferred dryer and heated soil in a mixed forest on a gentle slope.
473