ПАРАЗИТОЛОГИЯ, 2020, том 54, № 1, с. 3-12.
УДК 576.891:599.363:591.557
СВЯЗЬ ВИДОВОГО БОГАТСТВА
СООБЩЕСТВ ГЕЛЬМИНТОВ И ЧИСЛЕННОСТИ ХОЗЯИНА
(НА ПРИМЕРЕ ОБЫКНОВЕННОЙ БУРОЗУБКИ SOREX ARANEUS)
© 2020 г. Е. П. Иешкоa,*, А. В. Коросовb,**,
И. А. Никонороваa, С. В. Бугмыринa
aИБ КарНЦ РАН,
Пушкинская ул., 11, Петрозаводск, 185910, Россия
bПетрГУ,
Ленина пр., 33, Петрозаводск, 185910, Россия
* e-mail: ieshko@krc.karelia.ru
** e-mail: korosov@psu.karelia.ru
Поступила в редакцию 13.08.2019 г.
После доработки 20.01.2020 г.
Принята к печати 20.01.2020 г.
Изучено видовое богатство компонентных сообществ паразитов обыкновенной бурозубки
Sorex araneus в периоды её высокой и низкой численности. Анализ строился на сравнительной
оценке паразитарных инфрасообществ и кривых накопления видового богатства гельминтов.
Достоверность наблюдаемых различий определялась при анализе коэффициентов уравнения
степенной регрессии, моделирующей динамику накопления видового богатства паразитарных
сообществ. Показано, что при возрастании численности обыкновенной бурозубки наблюдается
снижение средних значений видового богатства паразитарных инфрасообществ, при этом растет
их β-разнообразие. Различия кривых накопления видового богатства паразитов в условиях низкой
и высокой численности хозяина определяются особенностями распределения видового богатства
инфрасообществ гельминтов. В период относительной депрессии численности S. araneus в струк-
туре инфрасообществ паразитов преобладали особи со средними значениями видового богатства.
В годы высокой численности бурозубок их популяция характеризуется преобладанием особей
с бедным видовым богатством паразитов.
Ключевые слова: паразитарные сообщества, компонентное, инфрасообщество, динамика
численности, кривые накопления видового богатства, степенное уравнение, регрессия
DOI: 10.31857/S1234567806010010
Состав и структура сообществ паразитов во многом определяются объемом ресур-
сов, предоставляемых хозяином - средой их обитания. Пространственным аспектом,
или «территорией», для паразитов является популяция хозяина, следовательно, состав
и структуру паразитарного сообщества будет определяться разнообразием и уровнем
численности их хозяев (Иешко, Голицына, 1984; Иешко и др., 2009). Иными словами, при
анализе пространственной структуры паразитарных сообществ более важным является
3
не размер исследуемой выборки, а экологическое соответствие изучаемых особей хозяев
требованиям жизненных циклов населяющих их паразитов.
Как уже отмечалось, экологическая паразитология, в отличие от экологии свободно-
живущих организмов, имеет определенные преимущества, связанные с представлениями
минимальной единицы паразитарного сообщества - инфрасообщество (Poulin, 2007).
Инфрасообщества - паразитарные населения отдельной особи хозяина, при соблюдении
определенных принципов однородности, являются статистически равноценными повтор-
ностями, количество и состав которых обусловливает структурированность видового
богатства компонентного сообщества паразитов (Иешко и др., 2019). Инфрасообщество
паразитов следует рассматривать как островную экосистему (MacArthur, Wilson, 1967;
Goater et al., 1987), где «островом» является особь хозяина, заселенная различным чис-
лом видов паразитов. Видовое богатство инфрасообществ паразитов может меняться
в широком диапазоне, но всегда уступает видовому богатству компонентного сообщества
(Poulin, 2007; Иешко и др., 2019). Наряду с числом видов для инфрасообществ важную
роль играет видовое разнообразие паразитов, что определяет степень сходства или раз-
личия состава паразитов у разных особей хозяина (β-разнообразие). Чем выше значения
β-разнообразия паразитарных инфрасообществ, тем больше вероятность выявления
новых видов паразитов с увеличением числа исследованных хозяев, что отражается на
скорости накопления видового богатства компонентного сообщества паразитов.
Для оценки богатства фауны паразитов может быть применен широко используемый
в классической экологии метод, который учитывает зависимость видового богатства
от площади обследованной территории, числа проб или, в случае паразитологических
исследований, количества особей хозяина (Джиллер, 1988). Для описания этой зависи-
мости для свободноживущих животных часто используется степенная регрессия. Было
показано, что форма кривых накопления зависит от относительного обилия и видового
разнообразия отдельной выборки, а также от порядка или последовательности анали-
зируемых проб (Thompson, Withers, 2003; Thompson, Thompson, 2007).
При достаточно проработанном алгоритме анализа и общей интерпретации результа-
тов найдется не много исследований, посвященных изучению формирования видового
богатства паразитов (Иешко, Коросов, 2012; Salgado-Maldonado et al., 2016; Pelegrini
et al., 2018; Иешко и др., 2019). Первыми в отечественной паразитологии данный подход
реализовали Петрушевский и Петрушевская (1960), показавшие, что богатство парази-
тофауны имеет закономерную связь с числом исследованных рыб.
В большинстве опубликованных работ построение кривых накопления служит ин-
струментом оценки репрезентативности выборочного усилия для характеристики той
или иной локальной паразитофауны и устраняет различия по количеству исследованных
особей хозяев сравниваемых выборок (Colwell et al., 2004; Dove, Cribb, 2006). Модели-
рование кривых видового богатства может иметь более широкое применение, например,
для оценки закономерностей накопления видового богатства паразитов в естественном
ареале хозяина и в условиях его интродукции (Иешко и др., 2019).
В настоящей работе рассмотрены особенности кривых накопления видового богат-
ства гельминтов обыкновенной бурозубки Sorex araneus Linnaeus, 1758 на стационарных
площадках в годы, различающиеся по уровню численности хозяина. Предпринята по-
4
пытка выявить влияние численности обыкновенной бурозубки на параметры накопле-
ния видового богатства компонентных сообществ паразитов. Будет проверена гипотеза
о влиянии численности хозяев на распределение и видовое богатство инфрасообществ
паразитов обыкновенной бурозубки.
МАТЕРИАЛ И МЕТОДЫ
Анализ видового богатства гельминтов обыкновенной бурозубки выполнен по материалам
паразитологических исследований, проведенных в районе Гомсельского научного стационара
(62.0697 с.ш., 33.9614 в.д.) в течение 8 лет с 2000 по 2007 годы. Для корректного сопоставления
результатов данные взяты только за август каждого года. Исключение составляет 2005 г., анализ
данных за этот год выполнен по материалам июльских сборов (отловы в августе 2005 г. не про-
водились).
За указанный период методом полного гельминтологического вскрытия (Аниканова и др.,
2007) обследовано 623 особи Sorex araneus (табл. 1). Показатели относительной численности
обыкновенной бурозубки - число особей на 100 ловушко-суток - представляют собой средние
значения по разнотипным биотопам в августе каждого года (Гусева и др., 2014).
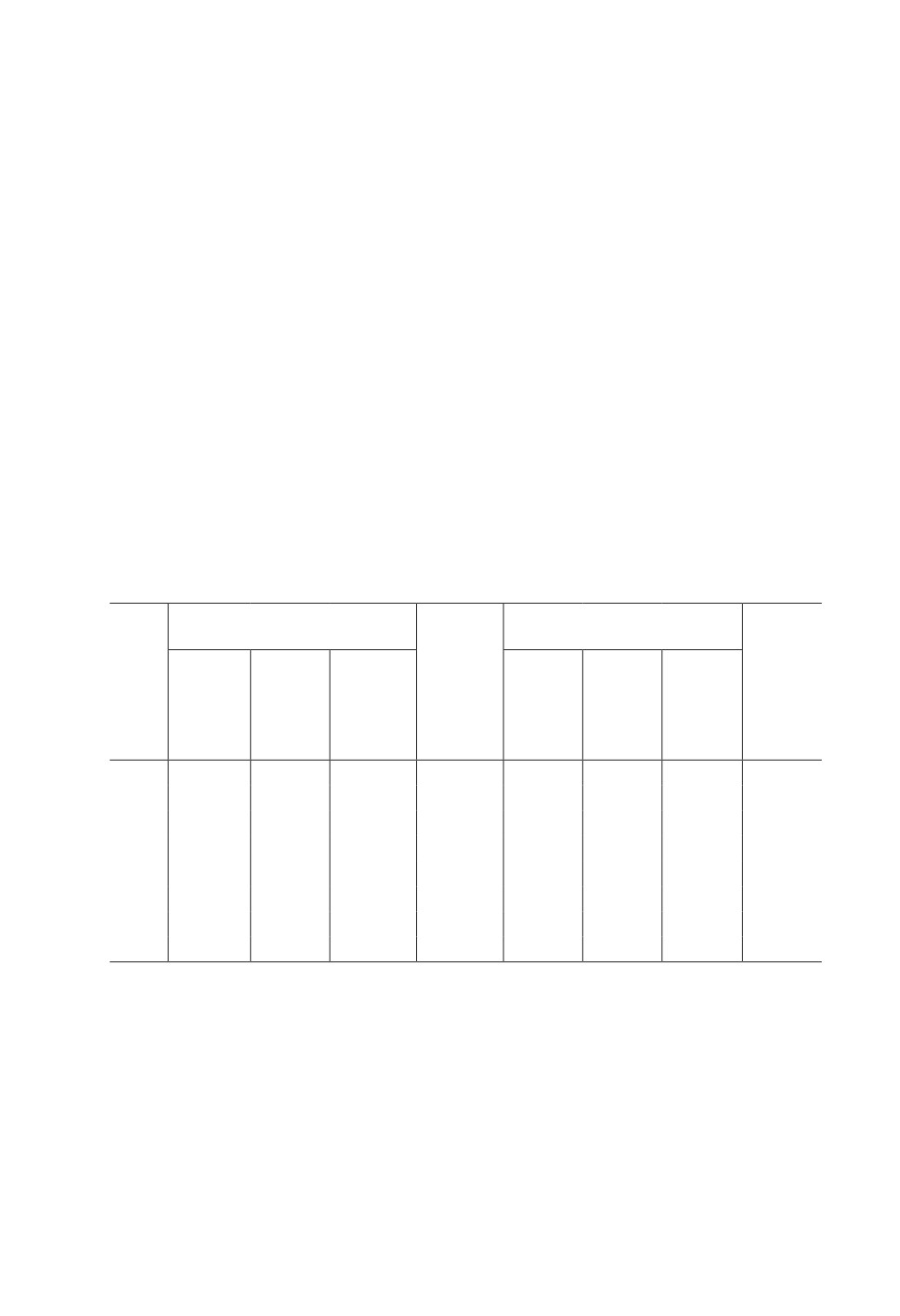
Таблица 1. Параметры видового разнообразия, численности и возрастной структуры популяции
обыкновенной бурозубки Sorex araneus в разные годы
Table 1. Parameters of species diversity, abundance and age structure of the Sorex araneus population
in different years
Видовое богатство
Бурозубки
инфрасообществ паразитов
Доля
Видов
Число
Числен-
зимо-
гельмин-
βWhit
исследо-
ность
Год
вавших
тов
Среднее
min-max
Медиана
ванных
на 100
особей,
особей
лов/сут
%
2000
18
6
2
18
5.2
2-9
5
2.5
2001
192
7
8
23
4.3
1-11
4
4.4
2002
96
7
6.5
26
3.8
1-11
3
5.8
2003
119
5
5.9
22
4.3
1-10
3
4.1
2004
56
11
2.7
25
5.8
1-13
6
3.3
2005
44
30
н.д.
19
4.5
1-10
4
3.2
2006
41
15
5
22
5
1-11
5
3.4
2007
57
16
3.8
22
5
1-9
5
3.4
П ри меч ан ия. βWhit - показатель β-разнообразия Уиттекера: компонентное паразитарное со-
общество состоит из инфрасообществ (сообществ гельминтов отдельных особей хозяина), ве-
личина β-разнообразия показывает степень различий (непохожести) состава гельминтов между
отдельными особями S. araneus (паразитарными инфрасообществами).
Для описания связи количества обнаруженных видов паразитов с увеличением объема выбор-
ки хозяина использовали уравнение степенной регрессии s=anb. Было показано, что параметры
уравнений регрессии могут иметь биологическую интерпретацию: коэффициент b характеризует
скорость нарастания видового богатства компонентных сообществ паразитов (s) при росте числа
5
проанализированных особей хозяина (n), коэффициент a равен числу видов паразитов первой
особи, взятой из данной выборки (Иешко и др., 2019).
С целью стандартизации выборки исследуемых популяций зверьков в разные годы в настоя-
щем исследовании уравнение степенной регрессии накопления видового богатства паразитов
строились по 50 особям хозяина. Возможные потери информации при сокращении выборки устра-
нялись путем рандомизации - расчета нескольких уравнений регрессии по серии ограниченных
выборок, взятых случайным образом из исходной выборки. В результате получаем множество
коэффициентов регрессии и рассчитываем их распределение. На этом основании можно выпол-
нять статистические сравнения пары коэффициентов регрессии, относящихся в нашем случае
к разным фазам динамики численности. В годы депрессии объемы выборок были невелики
(в 2000 г. всего 18 особей), поэтому для размножения малочисленной выборки и увеличении до
50 экз. использовали рандомизацию со случайным отбором и возвратом.
Обработка и анализ данных выполнен с использованием программ Past (Hammer et al., 2001)
рандомизации (Мастецкий, Шитиков, 2014) строит 300 серий кривых накопления видового богат-
ства паразитов и для каждого ряда рассчитывает коэффициенты а и b регрессионного уравнения.
Были рассчитаны группы коэффициентов по всем данным, для каждого года отдельно (2000-
2007 гг.) и для двух групп лет с высоким (годы 2001, 2002, 2003) и низким (годы 2000, 2004, 2007)
уровнем численности хозяина. Ряды этих коэффициентов использовали для расчета средних, их
распределений с оценкой значимости различий по W-критерию Вилкоксона.
В работе используются два ключевых понятия: «инфрасообщество гельминтов» - все виды
гельминтов, обнаруженные у отдельной особи хозяина, и«компонентное сообщество» - виды
гельминтов данной популяции хозяина (Holmes, Price, 1986).
РЕЗУЛЬТАТЫ
Характеристика видового богатства компонентных сообществ гельминтов
обыкновенной бурозубки в разные годы
За исследуемый период у обыкновенной бурозубки было выявлено 29 видов гель-
минтов трех систематических групп: трематоды (2), цестоды (16) и нематоды (11 видов).
Среди обнаруженных гельминтов у 17 видов (59 % фауны) встречаемость варьировала
от 1 до 10 % (Brachylaima fulvum, Rubenstrema exasperatum, Lineolepis scutigera, Vigisol-
epis spinulosa, Gulyaevolepis tripartite, Dilepis undula, Porrocaecum depressum,Calodium
soricicola, Stefanskostrongylus soricis, Soboliphyme soricis, Porrocaecum sp., Cladotaenia
globifera, Versteria mustelae, Polycercus lumbrici, Urocystis prolifer, Spirometra erinacei-
europaei, Staphylocystoides stefanskii). Менее многочисленную группу составили 8 видов
(27 %) встречаемость которых изменялась от 10 до 50 % (Ditestolepis diaphana, Staphylo-
cystis furcata, Neoskrjabinolepis schaldybini, N. singularis, Liniscus incrassatus, Aonchotheca
kutori, Eucoleus oesophagicola, Longistriata depressa). К массовым можно отнести только
4 вида (14 % от общей гельминтофауны), экстенсивность инвазии которых была выше
50% (Monocercus arionis, Longistriata didas, L. codrus, Parastrongyloides winchesi).
Богатство компонентных сообществ (число обнаруженных видов паразитов) в раз-
ные годы исследований варьировало от 18 до 26 видов. Средние многолетние значения
видового богатства инфрасообществ паразитов S. araneus (число видов гельминтов
у одной особи хозяина) составил 4.5 (табл. 1). В сборах не было бурозубок, не заражен-
6
ных гельминтами, минимальное богатство инфрасообществ бурозубки представлено
1-2, а максимальное 13 видами.
Распределение видового богатства инфрасообществ паразитов в исследованные
годы преимущественно носило агрегированный характер, при котором в популяции
доминировали животные с малым числом видов паразитов, особи с большим числом
видов паразитов были единичны. Распределения видового богатства инфрасообществ
паразитов в периоды, которые различались по численности бурозубок, имели досто-
верные различия (тест Колмогорова-Смирнова DN=0.43, p<0.05) (рис. 1). В период
относительной депрессии численности хозяина распределение инфрасообществ па-
разитов моделировалось нормальным законом, с преобладанием инфрасообществ со
средними значениями видового богатства. В годы с высокой численностью бурозубок
распределение инфрасообществ паразитов имело выраженный агрегированный характер,
и доминировали классы с малыми значениями видового богатства инфрасообществ
(рис. 1). Отмеченные различия структуры инфрасообществ паразитов подтвержда-
ются отрицательной корреляцией (r = -0.83, R2 = 0.69, р<0.05) между средними значе-
ниями богатства инфрасообществ и численностью бурозубок в исследованный период
(рис. 2). При этом наблюдается положительная корреляция (r=0.89, R2=79.5, p<0.05)
β-разнообразия паразитарных инфрасообществ и численности хозяина (рис. 2).
Рисунок 1. Частотное распределение видового богатства инфрасообществ паразитов в годы
низкой (1) и высокой численности (2) обыкновенной бурозубки.
Figure 1. Frequency distribution of species richness of parasite infra-communities in years of low (1)
and high (2) abundance.
7
Рисунок 2. Связь среднего видового богатства S inf (1) и β-разнообразия Уиттекера βWhit (2) инфра-
сообществ паразитов с относительной численностью обыкновенной бурозубки (Sorex araneus)
для исследованных лет.
Figure 2. Relationship between average species richness S inf (1) and β-diversity (2) of parasite infra-
communities with the relative abundance of Sorex araneus for the years studied.
Характеристика кривых накопления видового богатства
По данным каждого года были построены рандомизированные кривые накопления
видового богатства и рассчитаны значение коэффициентов a и b уравнения степенной
регрессии (рис. 3).
Различия кривых накопления видового богатства компонентных сообществ парази-
тов, построенные для обобщенных данных по периоду низкой и высокой численности
обыкновенной бурозубки (рис. 4), оценивали по распределению коэффициентов a и b
(рис. 5). Достоверные различия (p< 0.01) получены только для коэффициентов a, которые
отражают видовое богатство инфрасообществ. Для коэффициентов b, характеризующих
скорость накопления видового богатства, различия были не значимы (рис. 5).
Рисунок 3. Коэффициенты а и b cтепенных уравнений (даны среднее значение, ошибка и стан-
дартное отклонение), характеризующих кривые накопления видового богатства гельминтов Sorex
araneus в разные годы.
Figure 3. Indices a and b for degree equations (mean, standard error and standard deviation are given)
characterizing the curves of helminthic species richness accumulation of Sorex araneus in different years.
8
Рисунок 4. Кривые накопления видового богатства гельминтов (S) в годы низкой (1) и высокой
(2) численности обыкновенной бурозубки. По осям абсцисс - число исследованных хозяев, по
осям ординат - число обнаруженных видов паразитов.
Figure 4. Curves of the helminthic species richness accumulation (S) in the years of low (1) and high
(2) abundance of Sorex araneus. The abscissa axis is the number of studied hosts; the ordinate axis is
the number of detected parasite species.
Рисунок 5. Распределение коэффициентов a и b уравнений степенной регрессии; s=anb, рассчи-
танных для групп хозяев в периоды низкой (1) и высокой (2) численности. По осям абсцисс -
значения коэффициентов, по осям ординат - частота.
Figure 5. Distribution of indices a and b of the degree regression equations; s=anb, calculated for host
groups in periods of low (1) and high (2) abundance. The abscissa axis are the index values, the ordinate
axis presents the frequency values.
Обсуждение
Многолетний мониторинг видового богатства паразитов обыкновенной бурозубки
показал, что ядро сообщества формируют 18 видов, которые встречались в каждый год
исследования, как в период высокой, так и низкой численности хозяина. Варьирование
видового богатства компонентных сообществ паразитов в различные годы в большей
степени обусловлено редкими видами, которые были обнаружены всего 1 или 2 раза.
9
Многолетний стационарный мониторинг паразитофауны обыкновенной бурозубки
позволяет решать целый ряд задач экологической паразитологии, дает ответ на раз-
личные вопросы динамики паразитарных сообществ, устойчивости видового состава
и разнообразия паразитов. Полученные результаты показали, что видовое богатство
инфрасообществ обыкновенной бурозубки заметно уступает видовому богатству ком-
понентных сообществ. В популяции преобладали особи хозяев с малым числом видов
паразитов. Данный факт свидетельствует о том, что исследованное нами паразитарное
сообщество является устойчивым, не имеющим выраженной динамики в значениях
видового богатства паразитов, а различия богатства компонентных сообществ в разные
годы, которые мы выявляем в ходе паразитологических исследований, связаны с раз-
личиями частотного распределения видового богатства инфрасообществ паразитов.
Средняя продолжительность жизни S. araneus в условиях Карелии составляет око-
ло одного года. Бурозубки, родившиеся в текущем году, как правило, становятся по-
ловозрелыми и приступают к размножению только после зимовки (Ивантер, 1975).
Наши исследования мы проводили в августе, когда популяция S. araneus представлена
преимущественно молодыми незрелыми животными возрастом 1-2 мес., зараженными
небольшим числом паразитов. В годы высокой численности резко увеличивается доля
в популяции именно этой возрастной группы животных, что в свою очередь может
приводить к возрастанию в выборке животных с небольшим числом паразитов. Число
видов паразитов у одной особи S. araneus варьировало от 1 до 13. В период относительно
низкой численности бурозубок популяция характеризуется преобладанием относительно
«богатых» паразитарных инфрасообществ, и, наоборот, в период высокой численности
зверьков в структуре распределения богатства инфрасообществ преобладают низкие
значения видового богатства.
Выявляемое видовое богатство компонентного сообщества паразитов бурозубки
зависит от числа обследованных особей хозяина. В литературе взаимосвязь данных
параметров неоднократно анализировалась с помощью степенных или логарифмиче-
ских кривых (например, Walther et al., 1995; Dove, Cribb, 2006; Sarabeev, 2015; Sellers
et al., 2015). Особый интерес в использовании данного подхода состоит в биологиче-
ской интерпретации коэффициентов полученных степенных уравнений. Рассчитанные
параметры уравнений свидетельствуют о различиях инфрасообществ паразитов при
разном уровне численности хозяев. Периоды высокой и низкой численности бурозубок
достоверно различались по значениям коэффициента a и не различались по коэффици-
енту b. Иными словами в периоды высокой и низкой численности бурозубок скорость
накопления видового богатства (появление новых видов паразитов с увеличением числа
исследованных хозяев) практически не различается.
Моделирование кривых накопления видового богатства имеет важное значение
в характеристике видового разнообразия паразитов, выступая как инструмент опреде-
ления достаточности размера выборки и прогнозирования видового богатства. Кроме
того коэффициенты степенных уравнений являются информативными параметрами
структуры и разнообразия сообществ паразитов. Сравнительный анализ паразитарных
сообществ в разные годы показал информативность степенных уравнений регрессии
10
и перспективность биологической интерпретации коэффициентов, где один характери-
зует видовое богатство инфрасообществ (a), а второй определяет скорость накопления
компонентных сообществ паразитов (b). Исследование связи численности S. araneus
и видового богатства паразитов выявило различия частотного распределения богатства
инфрасообществ в годы высокой и низкой численности хозяина, что в свою очередь
может определять различия в числе обнаруженных видов гельминтов.
БЛАГОДАРНОСТИ
Появление статьи стало возможным благодаря многолетним исследованиям
с.н.с. лаборатории Паразитологии животных и растений ИБ КарНЦ РАН к.б.н. Ва-
лентины Семеновны Аникановой. Финансовое обеспечение осуществлялось из
средств федерального бюджета на выполнение государственного задания КарНЦ РАН
(№ 0218-2019-0075).
Список литературы
Аниканова В.С., Бугмырин С.В., Иешко Е.П. 2007. Методы сбора и изучения гельминтов мелких млекопи-
тающих. Учебное пособие. Петрозаводск, Карельский научный центр РАН, 115 с.
Гусева Т.Л., Коросов А.В., Беспятова Л.А., Аниканова В.С. 2014. Многолетняя динамика биотопического раз-
мещения обыкновенной бурозубки (Sorex araneus Linnaeus 1758) в мозаичных ландшафтах Карелии.
Ученые записки Петрозаводского государственного университета (8): 13-20.
Джиллер П. 1988. Структура и экологическая ниша. М., Мир, 184 с.
Ивантер Э.В. 1975. Популяционная экология мелких млекопитающих таежного Северо-Запада СССР. Л.,
Наука, 246 с.
Иешко Е.П., Бугмырин С.В., Аниканова В.С., Павлов Ю.Л. 2009. Особенности динамики и распределения
численности паразитов мелких млекопитающих. Труды Зоологического института РАН 313 (3): 319-328.
Иешко Е.П., Голицына Н.Б. 1984. Анализ пространственной структуры популяции трематоды
Bunodera luciopercae из окуня Perca fluviatilis. Паразитология 18 (5): 374-382.
Иешко Е.П., Коросов А.В., Соколов С.Г. 2019. Видовое богатство сообществ паразитов ротана Perсcottus glenii
(Actinopterygii: Odontobutidae) в нативной и приобретенной частях ареала хозяина. Паразитология 53
(2): 145-158. doi: 10.1134/S0031184719020066
Иешко Е.П., Коросов А.В. 2012. Оценка видового богатства паразитофауны рыб: экологический подход.
Принципы экологии (4): 28-40.
Мастицкий С.Э., Шитиков В.К. 2014. Статистический анализ и визуализация данных с помощью R. Электрон-
Петрушевский Г.К., Петрушевская М.Г. 1960. Достоверность количественных показателей при изучении
паразитофауны рыб. Паразитологический сборник ЗИН РАН 19: 333-343.
Colwell R.K., Mao C.X., Chang J. 2004. Interpolating, extrapolating, and comparing incidence-based species
Dove A. D.M., Cribb T.H. 2006. Species accumulation curves and their applications in parasite ecology. Parasitology
Goater T.M., Esch G.W., Bush A.O. 1987. Helminth parasites of sympatric salamanders: ecological concepts at
infracommunity, component and compound community levels. American Midland Naturalist 118: 289-300.
Hammer O., Harper D., Ryan P. 2001. PAST: Paleontological statistics software package for education and data
analysis. Palaeontologia Electronica 4 (1): 9.
Holmes J.C., Price P.W. 1986. Communities of parasites. In: Kikkawa J. and Anderson D.J. (eds). Community ecology:
patterns and processes. Melbourne, Blackwell Scientific Publications, 187-213.
MacArthur R.H., Wilson E.O. 1967. The theory of Island Biogeography. Princeton, Princeton University Press, 203 pp.
11
Pelegrini L.S., Januário F.F., Kozlowisky de Azevedo R., Abdallah V.D. 2018. Biodiversity and ecology of the parasitic
infracommunities of Loricaria prolixa (Siluriformes: Loricariidae) from the Tietê-Batalha Basin, SP, Brazil.
Acta Scientiarum. Biological Sciences 40: e36294. DOI: 10.4025/actascibiolsci.v40i1.36294
Poulin R. 2007. Evolutionary Ecology of Parasite (second edition). New Jersey08540, Princeton University Press,
332 pp.
Salgado-Maldonado G., Novelo-Turcotte M.T., Caspeta-Mandujano J.M., Vazquez-Hurtado G., Quiroz-Martínez
B., Mercado-Silva N., Favila M. 2016. Host specificity and the structure of helminth parasite communities of
fishes in a Neotropical river in Mexico. Parasite 23: 1-11. DOI: 10.1051/parasite/2016073
Sarabeev V. 2015. Helminth species richness of introduced andnative grey mullets (Teleostei: Mugilidae). Parasitology
International 64 (4): 6-17. DOI:10.1016/j.parint.2015.01.001
Sellers A.J, Ruiz G.M, Leung B., Torchin M.E. 2015. Regional variation in parasite species richness and abundance
in the introduced range of the invasive Lionfish, Pteroisvolitans. PLOS ONE 10 (6): e0131075. https://doi.
org/10.1371/journal.pone.0131075
Thompson G.G., Thompson S.A. 2007. Using species accumulation curves to estimate trapping effort in fauna surveys
and species richness. Austral Ecology 32 (5): 564-569. DOI:10.1111/j.1442-9993.2007.01728.x
Thompson G.G., Withers P.C. 2003. Effect of species richness and relative abundance on the shape of the species
Walther B.A., Cotgreave P., Price R.D., Gregory R.D., Clayton D.H. 1995. Sampling effort and parasite species rich-
ness. Parasitology Today 11 (8): 306-310.
SPECIES RICHNESS OF HELMINTH COMMUNITIES
IN RELATION TO HOST ABUNDANCE VARIATIONS
(THE CASE OF THE COMMON SHREW SOREX ARANEUS)
E. P. Ieshko, A. V. Korosov, I. A. Nikonorova, S. V. Bugmyrin
Keywords: parasite communities, component-, infracommunity, abundance variation,
species accumulation curves, power law equation, regression
SUMMARY
The aim of this study was to compare the parameters of the parasite species richness in common
shrew Sorex araneus in periods its population highest and lowest population abundance. The analysis
was mostly based on the estimation of differences in the species richness of helminth infracommunities
and coefficients of the power law equation of species accumulation curves for parasite component
communities. It was demonstrated that where common shrew abundances increased, the mean species
richness of infracommunities decreased, but their β-diversity rose. Differences between the species
accumulation curves for periods of shrew population highs and lows were dependent on the species
richness of parasite infracommunities. When the host abundance was relatively low, individuals
with average levels of parasite species richness prevailed in the population. In years with high shrew
abundances, the species richness of their parasitic infracommunities was low.
12