ПАРАЗИТОЛОГИЯ, 2020, том 54, № 1, с. 32-41.
УДК 576.895.132
CALODIUM SORICICOLA (YOKOGAWA ET NISCHIGORI, 1924)
(NEMATODA: CAPILLARIIDAE): ЭКОЛОГИЧЕСКИЕ ОСОБЕННОСТИ
ЗАРАЖЕННОСТИ ЗЕМЛЕРОЕК ЮГА ЗАПАДНОЙ СИБИРИ
© 2020 г. С. В. Карпенко a,*, В. В. Панов a,**
a Институт систематики и экологии животных СО РАН,
ул. Фрунзе 11, Новосибирск, 630091
* e-mail: swkarpenko@mail.ru;
** e-mail: panovv53@mail.ru
Поступила в редакцию 08.09.2019 г.
После доработки 20.01.2020 г.
Принята к печати 20.01.2020 г.
Представлены результаты шестилетних исследований (1983-1988 гг.) зараженности земле-
роек нематодой Calodium soricicola (Yokogawa et Nischigori, 1924) Moravec, 1982 (Nematoda,
Capillariidae) - паразитом паренхимы печени насекомоядных млекопитающих. В зоне подта-
ежных мелколиственных лесов (Северный район Новосибирской области) зарегистрировано
6 видов окончательных хозяев нематоды (Sorex araneus L., S. caecutiens Laxmann, S. daphaenodon
Thomas, S. minutus L., S. tundrensis Merriam и Neomys fodiens Pennant). Экстенсивность инвазии
всех насекомоядных C. soricicola в 1983-1988 гг. составляла 6.6 ± 0.8 % и по годам изменялась
от 3.2 ± 1.6 % в сухие годы до 9.1 ±1.7 % - во влажные. Наиболее многочисленной и наиболее
зараженной была обыкновенная бурозубка 10.3 ±1.1 %, которая является основным хозяином
C. soricicola в сообществе землероек. Выявлены значительные изменения зараженности её по
годам - от 5.2 ±1.7 % в сухие годы до 17.4 ±2.2 % - во влажные. Остальные 5 видов землероек
отнесены к второстепенным хозяевам из-за их малой численности и низкой зараженности. Про-
ведено исследование стадий и сроков развития личинок C. soricicola и C. hepaticum до инвазион-
ного состояния в лабораторных условиях. Результаты экспериментального заражения золотистых
хомячков инвазионными личинками обоих видов нематод показало, что хомячки заражались
C. hepaticum, но не C. soricicola.
Ключевые слова: Nematoda, Capillariidae, Calodium, зараженность, Soricidae, землеройки,
Западная Сибирь
DOI: 10.31857/S1234567806010046
В паренхиме печени у грызунов и насекомоядных млекопитающих паразитируют
разные виды нематод рода Calodium Dujardin, 1845: у грызунов - Calodium hepaticum
(Bancroft, 1893) Moravec, 1982 , а у насекомоядных - Calodium soricicola (Yokogawa
et Nischigori, 1924) Moravec, 1982. По поводу самостоятельности вида C. soricicola
в отечественной и зарубежной литературе нет единой точки зрения (Скрябин и др.,
32
1957). Основные морфологические отличия C. hepaticum от C. soricicola заключаются
в том, что C. hepaticum крупнее и размеры её органов больше, чем у C. soricicola. Анато-
мия половых органов у них во многом сходна. Недостаточно изучены жизненный цикл
C. soricicola, характер заражённости разных видов насекомоядных в естественных био-
ценозах и эпизоотология нематодоза. Отсутствуют сведения о животных, поедающих
зараженных нематодами землероек и рассеивающих яйца калодиумов. Суммарная за
все годы исследований экстенсивность инвазии калодиумами основных видов хозяев,
отловленных в одних и тех же биоценозах Северной Барабы, у водяной полёвки Arvicola
amphibius (Linnaeus, 1758) (syn. Arvicola terrestris L.) нематодой C. hepaticum - 10.4 ±
1.02 % (Чечулин и др., 2011), а у землероек S. araneus - нематодой C. soricicola - 10.3 ±
1.1 % - т. е. одинакова. Наши исследования были предприняты с целью уточнения систе-
матического статуса вида, а также изучения экологических особенностей зараженности
нематодами C. soricicola насекомоядных млекопитающих.
Нематода C. soricicola зарегистрирована в России у землероек в Алтайском крае
(Павлов, Юнь Лянь, 1962; Карпенко, 1998а), Бурятии (Карпенко, 1985; Карпенко и др.,
1989), Калининской (Викторов и др., 1964), Ленинградской (Васильев, 1949) и Ново-
сибирской областях (Карпенко, 1998б), а также в Бельгии (Bernard, 1969), Белоруссии
(Морозов, 1956; 1958; Меркушева, 1966; Арзамасов и др., 1969), Болгарии (Prokopic,
Genov, 1974; Генов, 1984), Венгрии (Matskási et al., 1990), Германии (Stammer, 1955),
Испании (Mas Coma, 1977), Польше (Soltys, 1952), Чехии (Prokopic, 1959), Франции
(Dollfus, 1961), Швейцарии (Jourdane, 1971) и Японии (Nischigori, 1924).
Район исследований расположен в среднем течении р. Тартас. Он относится к подзоне
мелколиственных лесов Тартас-Ичинского ландшафтно-зоогеографического региона и
представляет собой макросклон от Обь-Иртышского междуречья к низинам централь-
ной Барабы. В целом для этого ландшафта характерны сильная мозаичность и мелко-
контурность почвенно-геоботанических выделов (Гантимуров, Супряга., 1967; Глотов
и др., 1978). Равнинный рельеф и близкое залегание глинистых пород определяют фор-
мирование обширных массивов болот различного типа. Дренированные возвышенные
участки занимают березовые, реже березово-осиновые леса. В понижениях рельефа на
открытых пространствах образуются кочкарные болота с зарослями тростника и осоки,
в лесных массивах - болота лесного типа. Залесенность территории составляет около
60 %, заболоченность - более 30 %. (Глотов и др., 1978). Pайон исследований можно
охарактеризовать как единый лугово-лесо-болотный ландшафт (Чечулин и др., 2011).
Для Северной Барабы характерно чередование сухих и влажных периодов в 11-летних
циклах увлажненности. Это явление, первоначально описанное при анализе вспышек
массового размножения водяных полевок (Максимов, 1982), оказывает существен-
ное влияние на изменения гидрологического режима, сукцессии животного населения
и состояние растительного мира (Максимов и др., 1979; Максимов, 1989). Влажные годы
характеризуются высокими и продолжительными разливами рек, заполнением мелких
пойменных водоемов, значительным увеличением площади и длительности обводнения
болот. На приболотные леса и луга проникают гидрофильные виды растений, возрас-
тает численность водных и гидрофильных животных, которые широко расселяются по
территории. В сухие годы болота обводнены лишь весной, к середине лета практически
33
вся их площадь обсыхает. На болота проникают луговые виды растений, численность
гидрофильных видов животных снижается.
Сообщество землероек района исследований насчитывает 8 видов двух родов: обык-
новенная бурозубка Sorex araneus L., средняя бурозубка Sorex caecutiens Laxmann,
крупнозубая бурозубка Sorex daphaenodon Thomas, малая бурозубка Sorex minutus L.,
тундряная бурозубка Sorex tundrensis Merr, крошечная бурозубка Sorex minutissimus
Zimmermann, равнозубая бурозубка Sorex isodon Turov и обыкновенная кутора Neomys
fodiens Pennant.
Обыкновенная бурозубка S. araneus является доминантным видом в сообществе
землероек Северной Барабы. Она населяет почти все биотопы в районе исследований,
причем почти во всех доминирует по численности. Это обусловлено экологической
пластичностью S. araneus и, не в последнюю очередь, крупными размерами зверьков,
что увеличивает спектр кормов, доступных виду и повышает его конкурентоспособность
(Долгов, 1985). Другие виды бурозубок для ослабления конкурентных отношений либо
ограничивают набор местообитаний (средняя и тундряная бурозубки), либо «расходят-
ся» в динамике численности (малая бурозубка). У последнего вида пики численности
не совпадают с пиками вида-доминанта. Кутора N. fodiens - это околоводный зверек
и в других биотопах встречается в основном при расселении. Околоводный образ жиз-
ни и крупные размеры обусловливают специфику питания - набор кормов этого вида
значительно отличается от такового бурозубок.
Цель исследований - изучение динамики зараженности землероек нематодой
C. soricicola на юге Западной Сибири.
МАТЕРИАЛ И МЕТОДИКА
Гельминтологические исследования насекомоядных млекопитающих проведены в течение
летне-осенних сезонов 1983-1988 гг. в Северной Барабе (окрестности с. Усть-Ургулька Северного
района Новосибирской области). Наши исследования проведены как в сухие (1983-1984), так
и во влажные (1985-1988) годы.
Землеройки были отловлены с помощью стандартных ловчих канавок (50 м длиной с 5 ловчими
цилиндрами). Результаты учета животных пересчитаны в единицы относительной численно-
сти - количество экземпляров на 100 цилиндро-суток (цс). Учеты мелких млекопитающих про-
водились в 1978-1990 гг. и среднемноголетние данные по численности и демографии землероек
в тексте даны за весь период учетов, а в таблице и на графике - приводятся за период 1983-1988,
когда проводились гельминтологические исследования. Материал по динамике численности
мелких млекопитающих частично опубликован (Макимов, Ердаков, 1985; Panov, 2000; Панов,
Карпенко, 2004).
Методом полных гельминтологических вскрытий исследовано 1407 экз. восьми видов землеро-
ек. В качестве основной характеристики зараженности животных были использованы показатели
доли зараженных особей в исследованной выборке (Р %) - экстенсивность инвазии (Э.И.) и ее
ошибки - (± m). Вычисление индекса обилия C. soricicola (количество нематод на одно иссле-
дованное животное) вызывает затруднения, а при значительном поражении печени практически
невозможно. Это связано с особенностями локализации паразитов и их тонким нитевидным строе-
нием. Часто мы видим либо клубки зрелых самцов и самок, выделить и подсчитать которых не
34
представляется возможным, либо яйца, окружённые соединительными тканями хозяина. Иногда
в паренхиме печени обнаруживаются только переплетенные яйцевые тяжи, а сами нематоды уже
погибли. Достоверность различий в частоте встречаемости C. soricicola у разных видов сорицид
(P) оценивали по параметрическому критерию Стъюдента - t.
Для экспериментального изучения развития нематод червей изымали из печени грызунов
и землероек. Зрелые яйца извлекались из взрослых самок нематод методом разрыва стенок мат-
ки. Они были помещены в 5 чашек Петри на влажную фильтровальную бумагу по 5-6 групп из
25-35 яиц в каждой чашке. Яйца содержали при температуре 20-24 ºC и постоянной влажности
в условиях естественного освещения, но без прямого солнечного воздействия. Яйца периодиче-
ски, 1-2 раза в неделю, исследовали под микроскопом. Отслеживали стадии и сроки развития
личинок, проверяли их жизнеспособность. Основной нашей задачей было выяснить, смогут ли
инвазионные личинки C. soricicola прижиться в печени грызунов и развиться до половозрелого
состояния. В качестве экспериментальных дефинитивных хозяев были использованы молодые зо-
лотистые хомяки, свободные от нематодной инвазии. Яйца, содержащие подвижных инвазионных
личинок, помещались на кусочки хлеба. Хлеб скармливали золотистым хомякам, которые были
предварительно выдержаны на голодной диете. Для предотвращения гиперинвазии на каждый
кусочек наносили не более 8-12 яиц, а каждому хомяку скармливался только один кусочек. Через
4-14 дней хомячков исследовали на предмет обнаружения нематод. Эксперименты по изучению
развития нематод до стадии инвазионных личинок и заражение млекопитающих инвазионными
личинками, выращенными в лабораторных условиях, проводили в 1983 и 1985 гг. Исследования
были проведены при соблюдении правил проведения научных исследований с использованием
экспериментальных животных, утвержденных распоряжением Президиума АН СССР от 2 апреля
1980 N 12000-496 и приказом Минвуза СССР от 13 сентября 1984 N 22.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Динамика численности хозяев
Средняя многолетняя численность сообщества бурозубок для всех восьми изучен-
ных видов составляет 30.46±5.19 экз./100 цс. Численность отдельных видов сообще-
ства землероек в период гельминтологических исследований представлена в табл. 1.
Численность большинства видов землероек по годам изменяется не синхронно, лишь
изменение численности обыкновенной и средней бурозубок происходит более или менее
синхронно. Большинство видов имеют свою собственную динамику численности. По
нашим данным среди исследованных землероек преобладают неполовозрелые сеголет-
ки, а среди взрослых особей самцов немного больше, чем самок, эта картина типична
для материалов, полученных при отлове в канавки (Щипанов и др., 2003). Среднемно-
голетняя численность вида-доминанта (S. araneus) составила 16.60±3.01 экз./100 цс,
а ее доля в сообществе землероек - 54.5±1.4 %. Демографическая структура обычна
для мелких млекопитающих (53.9±0.7 % самцов и 80.3±0.6 % сеголеток). Второй по
обилию вид в этом сообществе землероек - кутора. Ее среднемноголетняя численность
5.33±2.26 экз./100 цс, а доля в сообществе достигает 17.6±1.1 %. Средняя бурозубка
в данном сообществе занимает третье место по численности - 3.75±0.81 экз./100 цс, что
составило 12.3±0.9 % от всего сообщества. Малая бурозубка лишь немного уступает
средней по численности (3.25±0.5 экз./100 цс), ее доля в сообществе составляет 10.7±
35
0.9 %. Тундряная бурозубка малочисленна (1.06±0.21 экз./100 цс), а ее доля в сообществе
составляет лишь 3.5±0.5 %. Крупнозубая и равнозубая бурозубки самые малочисленные
представители сообщества землероек Усть-Ургульки. Численность крупнозубой бурозуб-
ки - 0.41±0.2 экз./100 цс, что составляет 1.3±0.3 % от общей численности сообщества.
Численность равнозубой - 0.03±0.01 экз./100 цс, а доля ее в сообществе - 0.1±0.1 %.
Крошечная бурозубка встречается в единичных экземплярах и не ежегодно. Численность
отдельных видов сообщества землероек в период гельминтологических исследований
представлена в табл. 1.
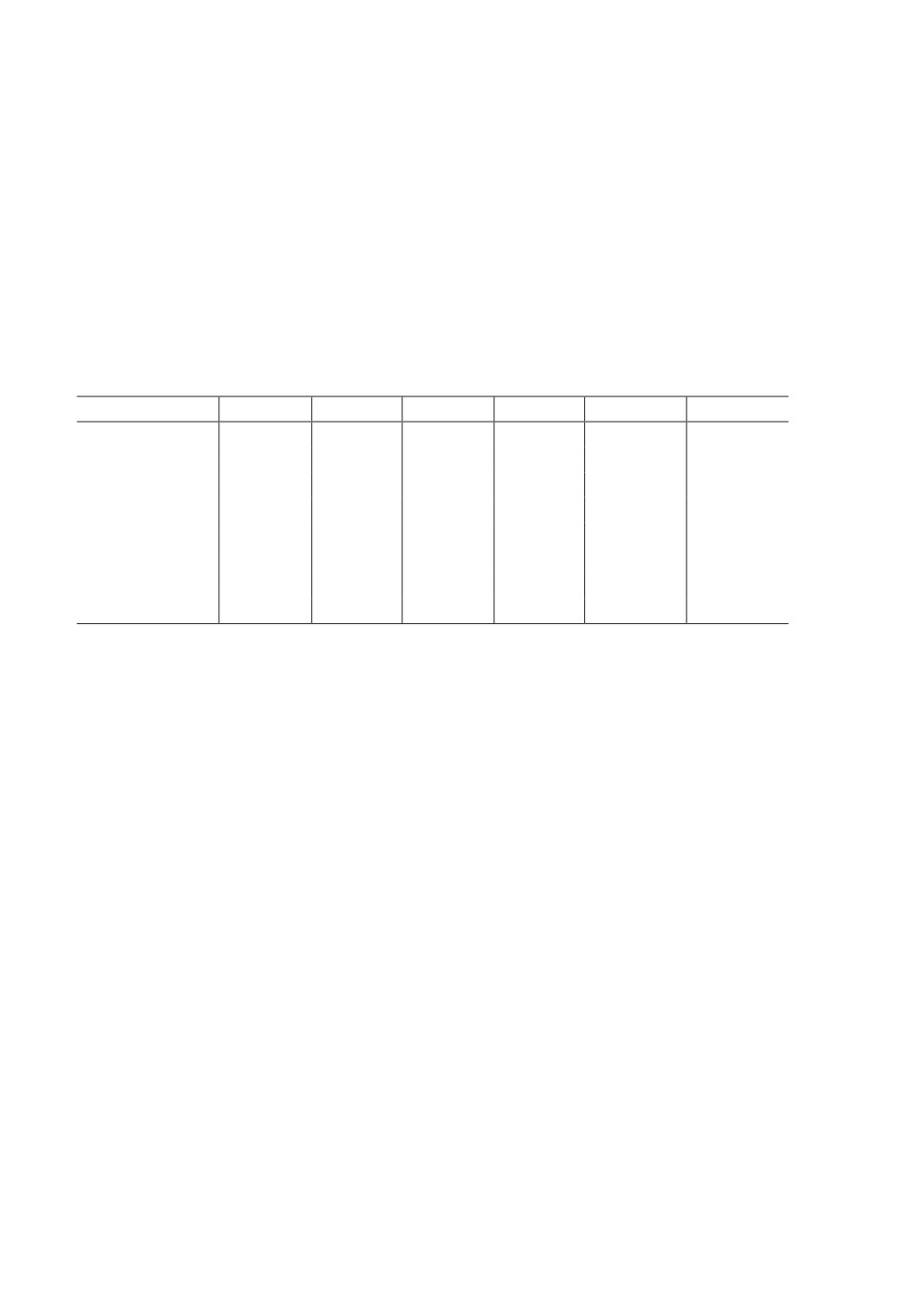
Таблица 1. Изменения относительной численности (на 100 цилиндро-суток)
популяций землероек по годам
Table 1. Dynamics in relative number (per 100 cylindro-days)
of shrews populations, by years (mean ± m)
Вид
1983
1984
1985
1986
1987
1988
S. araneus
13.6 ± 0.6
35.4 ± 1.1
32.3 ± 1.2
26.6 ± 1.1
23.9 ± 0.8
18.3 ± 0.2
S. caecutiens
1.8 ± 0.2
6.8 ± 0.6
6.1 ± 0.6
3.9 ± 0.5
5.9 ± 0.4
4.2 ± 0.1
S. daphaenodon
0
0
0
0
2.6 ± 0.3
0.7 ± 0.04
S. minutus
3.4 ± 0.3
5.5 ± 0.5
2.4 ± 0.4
4.6 ± 0.5
4.9 ± 0.4
6.3 ± 0.1
S. tundrensis
1.7 ± 0.2
2.7 ± 0.4
0.9 ± 0.2
1.4 ± 0.3
1.1 ± 0.1
1.03 ± 0.05
S. minutissimus *
0.03 ± 0.03
0
0
0.1 ± 0.1
0.03 ± 0.03
0.04 ± 0.01
S. isodon *
0.1 ± 0.1
0
0.07 ± 0.07
0.1 ± 0.1
0
0
N. fodiens
0.1 ± 0.1
0.8 ± 0.2
7.0 ± 0.7
28.5 ± 1.1
7.2 ± 0.5
13.6 ± 0.2
П ри м еча ние. * Виды землероек, у которых нематоды не обнаружены.
Характер зараженности отдельных видов землероек C. soricicola
Методом полных гельминтологических вскрытий исследовано 1407 экз. землеро-
ек 8 видов: 736 экз. S. araneus, 176 экз. S. caecutiens, 10 экз. S. daphaenodon, 161 экз.
S. minutus, 52 экз. S. tundrensis, 1 экз. S. minutissimus, 4 экз. S. isodon и 267 экз. N. fodi-
ens. В Северной Барабе нематода C. soricicola зарегистрирована у 6 видов землероек:
обыкновенной, средней, крупнозубой, малой, тундряной бурозубок и куторы.
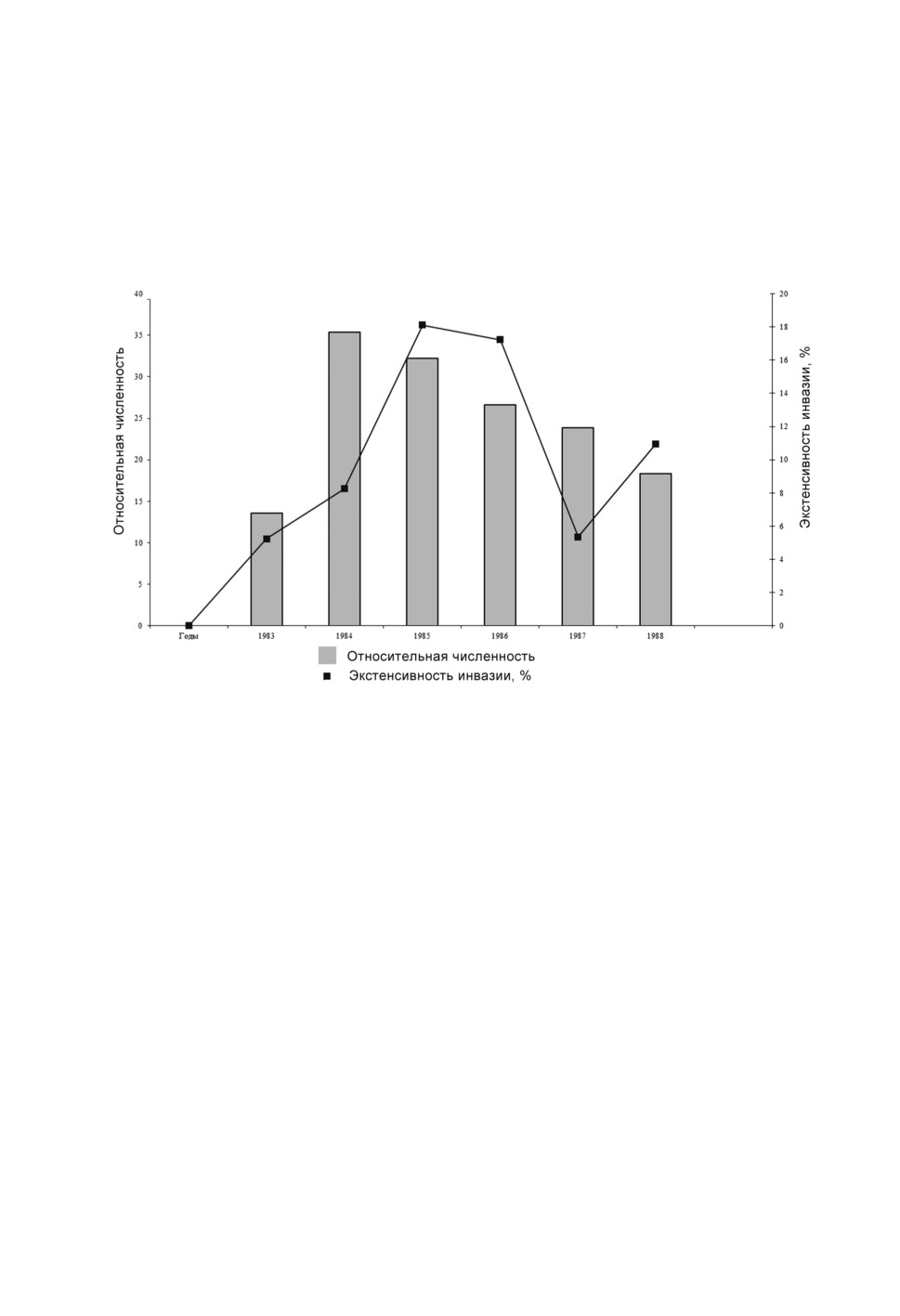
Суммарная за все годы исследований экстенсивность инвазии популяции обык-
новенной бурозубки C. soricicola составляла 10.3 ± 1.1 %. Наименьшие показатели
зараженности отмечены в 1983 году в начале подъема численности S. araneus - 5.23 ±
1.7 %. Пик численности бурозубки зарегистрирован в 1984 году, затем наблюдалось по-
степенное её снижение (табл. 1, рис. 1). Экстенсивность инвазии S. araneus нематодой
в 1984 г. увеличилась до 8.25 ±1.92 %, достигнув в 1985 году максимальных значений
18.11 ±3.27 %, на следующий год после пика численности бурозубки. Несколько ниже
была зараженность S. araneus в следующем году - 17.24 ±4.1 %, а затем этот показатель
снижался вместе с численностью хозяев: в 1987 г. - 5.36 ± 3.0 % и несколько выше
в 1988 г. - 10.97 ± 3.66 %.
Остальные виды землероек заражены в гораздо меньшей степени, а у некоторых
видов нематод не обнаружено, возможно, это обусловлено недостаточно большой выбор-
кой (табл. 1). Суммарная экстенсивность инвазии нематодой средней бурозубки в 1983-
1988 гг. составляла 1.14 ± 0.8 %, малой бурозубки - 3.1 ± 1.37 %, тундряной бурозубки -
36
5.77 ± 3.23 %. Зараженных особей в выборках этих видов было очень мало, а в некоторые
годы их не было совсем. Крупнозубая бурозубка в малых количествах встречалась в отло-
вах лишь в 1987-1988 гг., и всего 1 экз. был заражен нематодой. Кутора была доминантом
в сообществе землероек в 1986 г. Зараженные нематодой особи куторы зарегистрированы
лишь при повышении и на пике ее численности: в 1985 г. (Э.И. 1.35 ± 0.16 %) и 1986 г.
(Э.И. 2.65 ± 0.14 %), т.е. в годы её наиболее активного расселения.
Рисунок 1. Динамика численности S. araneus и зараженности её C. soricicola в Северной Барабе.
Figure 1. Population dynamics of S. araneus and its infection in the C. soricicola in North Baraba.
Сообщество землероек по степени зараженности нематодой можно условно разде-
лить на две группы - высоко зараженных и мало зараженных. В первую группу входит
только S. araneus, а во вторую - все остальные виды землероек. Экстенсивность инвазии
всех видов второй группы суммарно и за все годы исследований - 2.25 ± 0.6 %, а экс-
тенсивность инвазии S. araneus за все годы исследований - 10.3 ± 1.1 %, т.е. достоверно
выше, чем у видов второй группы.
В биоценозах Северной Барабы жизненный цикл нематоды С. soricicola осуществля-
ется с участием шести видов землероек в качестве окончательных хозяев. Но именно
обыкновенная бурозубка является основным окончательным хозяином C. soricicola.
Экстенсивность инвазии остальных видов сообщества землероек суммарно за все годы
исследований достоверно ниже, что означает их второстепенную роль в качестве хозяев
нематоды.
Изменения экстенсивности инвазии обыкновенной бурозубки по годам коррелируют
с колебаниями численности хозяина с задержкой в 1 год. При возрастании численности
S. araneus в следующем году увеличивается экстенсивность инвазии, а при снижении
численности в следующем году уменьшается и зараженность.
37
Экспериментальные исследования
С целью получения дополнительных доказательств самостоятельности вида
C. soricicola было принято решение экспериментально вырастить личинок C. hepaticum
и C. soricicola до инвазионного состояния и выяснить возможность перекрестного зара-
жения грызунов и насекомоядных млекопитающих. Содержание землероек в вольерных
или клеточных условиях весьма затруднительно, поэтому мы ограничились неполной
моделью перекрестного заражения (нематодами от грызунов к грызунам и нематодами
от землероек также к грызунам). Материалы по экспериментальным исследованиям
частично опубликованы (Карпенко, Чечулин, 2004).
Развитие C. hepaticum. Через 20 дней после начала эксперимента содержимое
яиц находится на стадии 8 бластомеров. Через 31 день личинки находятся на стадии
16 бластомеров. Через 42 дня личинки находятся на стадии морулы. Через 57 дней
в яйцах наблюдаются полностью сформированные и активно движущиеся личинки не-
матоды. Такие яйца с помощью пипетки были перенесены на кусочки хлеба, которые
скормлены 4 особям золотистого хомячка, свободным в отношении C. hepaticum. Через
4 дня после заражения вскрыто 2 хомяка. У одного найдено 2 экз. C. hepaticum с фор-
мирующимися в матке яйцами. У 2-го хомяка найдено 4 экз. молодых нематод. Через
10 дней вскрыто 2 хомяка, у одного обнаружено 3 экз. C. hepaticum с формирующимися
в матке яйцами. У второго нематод не найдено.
Развитие C. soricicola. Через 12 дней после начала эксперимента содержимое яиц
находится на стадии 8 бластомеров. Через 20 дней личинки находятся на стадии 16 бла-
стомеров. Через 30 дней личинки находятся на стадии морулы. Через 49 дней после на-
чала эксперимента в яйцах найдены полностью сформированные и активно движущиеся
личинки нематоды. Такие яйца с помощью пипетки были перенесены на кусочки хлеба
и впоследствии скормленны 6 особям золотистого хомячка, свободным в отношении
C. hepaticum. Через 7 дней после заражения вскрыты все 6 особей экспериментально
зараженных C. soricicola хомяков. Инвазированных нематодами рода Calodium Dujardin,
1845 не найдено.
Так как результаты экспериментального заражения хомяков нематодой C. soricicola
были отрицательными, было решено повторить лабораторное развитие нематоды до по-
явления инвазионных личинок и эксперименты по заражению ими золотистых хомяков.
Яйца C. soricicola от землероек содержались в чашках Петри на влажной фильтро-
вальной бумаге при 20-24 ºС в течение 49 дней. Через 45 дней развития в яйцах найдены
полностью сформированные и активно движущиеся личинки. Такие яйца с помощью
пипетки были перенесены на кусочки хлеба, впоследствии скормленные свободным
в отношении C. soricicola 11 экз. золотистых хомяков. Через 7 дней после зараже-
ния вскрыто 5 хомяков. Зараженных нематодами рода Calodium не обнаружено. Через
14 дней после заражения вскрыто ещё 6 хомяков. Зараженных нематодами рода Calo-
dium не обнаружено.
Яйца, отложенные самками нематод в паренхиму печени хозяина, при жизни его во
внешнюю среду не выделяются. При экспериментальном изучении развития личинок
C. soricicola выяснено, что при температуре 18 ºC зародыш формируется в течение
38
68 дней, а при 32 ºC - в течение 28 дней (Скрябин и др., 1957). При скармливании вполне
созревших яиц белым мышам личинки вылупляются в кишечнике, затем проникают
в брюшную полость и мигрируют на поверхность печени. Однако в организме белых
мышей, согласно экспериментам Nischigori (1924), развитие нематод до половозрелой
стадии не происходит.
В условиях наших экспериментов, проведенных в одно и то же время, при одинако-
вых температурах и влажности, сроки развития личинок C. hepaticum и C. soricicola до
инвазионного состояния различаются на 8 -9 дней. Различаются и сроки формирования
отдельных стадий развития личинок. Различия в сроках развития и полученные дважды
отрицательные результаты по экспериментальному заражению хомяков инвазионными
личинками C. soricicola подтверждают видовую самостоятельность C. soricicola.
БЛАГОДАРНОСТИ
Исследования выполнены по программе фундаментальных научных исследований
государственных академий наук на 2013-2020 гг.,
проект № VI. 51.1.5. (0311-2019-0016) - (АААА-А16-116121410121-7)
и проект № VI.51.1.9. (0311-2019-0018) - (АААА-А16-116121410119-4).
Список литературы
Арзамасов И.Т., Меркушева И.В., Михолап О.Н., Чикилевская И.В., 1969. Насекомоядные и их паразиты на
территории Белоруссии. Минск, Наука и техника, 175 с.
Васильев В.В. 1949. Паразитофауна грызунов и насекомоядных из окрестностей Ленинграда. Ученые записки
ЛГУ. Серия биологическая 101 (19): 73-81.
Викторов Л.В., Головин О.В., Савинов В.А. 1964. Материалы по гельминтофауне насекомоядных млеко-
питающих Калининской области. Ученые записки Калининского государственного педагогического
института 31: 289-292.
Гантимуров И.И., Супряга И.К. 1967. Агропочвенное районирование Новосибирской области. Новосибирск,
Наука, 127 с.
Генов Т., 1984. Хелминти на насекомоядните бозайници и гризачите в България. София, Издательство БАН,
348 с.
Глотов И.Н., Ердаков Л.Н., Кузякин В.А., Максимов А.А., Мерзлякова Е.П., Николаев А.С., Сергеев В.Е. 1978.
Сообщества мелких млекопитающих Барабы. Новосибирск, Наука, 237 с.
Долгов В.А. 1985. Бурозубки Старого Света. М., Изд-во МГУ, 220 с.
Карпенко С.В. 1985. Гельминты насекомоядных млекопитающих семейства Soricidae зоны влияния Байкало-
Амурской магистрали. Автореф. дис. … канд. биол. наук. Алма-Ата. 22 с.
Карпенко С.В., 1998а. Гельминты землероек предгорий Западного Алтая. В кн. Паразиты в природных ком-
плексах и рисковые ситуации. Новосибирск, Наука, 69-71.
Карпенко С.В., 1998б. Гельминты землероек (Mammalia, Insectivora, Soricidae) Северной Барабы. В кн. Бес-
позвоночные животные Южного Зауралья и сопредельных территорий. Курган, Изд-во Курганского
ун-та, 182-187.
Карпенко С.В., Федоров К.П., Чечулин А.И. 1989. Нематоды бурозубок зоны влияния Байкало-Амурской
магистрали. В кн. Экология гельминтов позвоночных Сибири. Новосибирск, Наука, 86-105.
Карпенко С.В., Чечулин А.И. 2004. Мониторинг зараженности микромаммалий Северной Барабы нематодами
рода Hepatica Hall, 1916 и экспериментальное доказательство валидности вида Hepaticola soricicola
Yokogawa in Nischigori, 1924. Сибирская зоологическая конференция, Тезисы докладов всероссийской
конференции, посвященной 60-летию ИСиЭЖ СО РАН, 15-22 сентября 2004. Новосибирск, 380.
39
Максимов А.А. 1982. Исследование смен фаз увлажненности территорий лесостепной зоны Западной Сиби-
ри в 11-летних циклах. В кн. Природные циклы Барабы и их хозяйственное значение. Новосибирск,
Наука, 6-24.
Максимов А.А. 1989. Природные циклы. Причины повторяемости экологических процессов. Л. Наука. 236 с.
Максимов А.А., Ердаков Л.Н. 1985. Циклические процессы в сообществах животных. Новосибирск, Наука,
236 с.
Максимов А.А., Понько В.А., Сытин А.Г. 1979. Смена фаз увлажненности Барабы (характеристика и прогноз).
Новосибирск, Наука, 64 с.
Меркушева И.В., 1966. Гельминтофауна обыкновенной бурозубки (Sorex araneus L.) в Белоруссии. Материалы
конф. Всесоюзного об-ва гельминтологов. М., т. 3, 178-183.
Морозов Ю.Ф. 1956. К познанию гельминтофауны грызунов и насекомоядных СССР и опыт ее эколого-
фаунистического анализа. Автореф. дис. … канд. биол. наук. М., 16 с.
Морозов Ю.Ф. 1958. Материалы по гельминтофауне грызунов и насекомоядных Беловежской Пущи. Труды
заповедно-охотничьего хозяйства "Беловежская Пуща" 1: 151-175.
Павлов А.Ф., Юнь Лянь 1962. К обнаружению нематоды Hepaticola soricicola у землероек Алтая. Труды
ГЕЛАН СССР 12: 90-92.
Панов В.В., Карпенко С.В. 2004. Динамика популяции куторы обыкновенной Neomys fodiens Pennnant (Mam-
malia, Soricidae) и ее гельминтофауны в Северной Барабе. Паразитология 38 (5): 426-437.
Скрябин К.И., Шихобалова Н.П., Орлов И.В. 1957. Трихоцефалиды и капиллярииды животных и человека и
вызываемые ими заболевания. Основы нематодологии. М., Наука, т. 6., 585 с.
Чечулин А.И., Карпенко С.В., Панов В.В. 2011. Экологические особенности заражения нематодой Hepaticola
hepatica (Bancroft, 1893), Hall, 1916 (Nematoda, Capillariidae) грызунов юга Западной Сибири. Сибирский
экологический журнал 4: 563-569.
Щипанов Н.А., Купцов А.В., Калинин А.А., Олейниченко В.Ю. 2003. Конуса и живоловки ловят разных
землероек-бурозубок (Insectivora, Soricidae). Зоологический журнал 82 (10): 1258-1265.
Bernard J., 1969. Observations sur les helminthes parasites de Mammifères et d'oiseaux de la faune de Belgique.
Archives de l’Institute Pasteur de Tunis 46: 137-193.
Dollfus, R.Ph. 1961. Contribution a la faune parasitaire regionale. Station experimentale de parasitologie de Richelieu
(Indre-et-Loire). Annales de Parasitologie Humaine et compare 36 (5-6): 171-354.
Jourdane J. 1971. Helminthes parasites des Micromammiferes des Pyrenees Orientales. 2. Les Plathelminthes de
Sorisinae. Annales de Parasitologie Humaine et compare 46 (5): 553-573.
Mas Coma S. 1977. Contribution al conocimiento de la helmintofauna de micromamiferos ibericos. II. Parasitos de
Crocidura russula (Hermann, 1780) (Insectivora: Soricidae). Säugetierk. Mitteil. 25. (1). 67-78.
Matskási I. , Mészáros F., Murai E. 1990. Helminthological investigations of vertebrates in the Bátorliget Nature
Reserves (Trematoda, Cestoda, Nematoda, Acanthocephala). In: Mahunka, S. (ed.): The Bátorliget Nature
Reserves-after forty years. Budapest, Hungarian Natural History Museum, 121-128.
Nischigori M. 1924. On a new speсies of the genus Hepaticola. Taiwan Ig. Kw. Z. № 236. (Abstr.) Japanese Journal
of Zoology (1926), Vol. 1, 124-125.
Panov V.V. 2000. Diversity of the shrew Communities in the Southern part of West Siberia. Biodiversity and Dy-
namics of ecosystems in North Eurasia. Diversity of the fauna in North Eurasia. Novosibirsk, IC&G, Vol. 3,
Part 1, 191-193.
Prokopic J. 1959. Cisopasni cervi nasich hmuzozravcu. Folia parasitologica 6 (2): 87-134.
Prokopic J., Genov T. 1974. Distribution of helminthes in micromammals (Insectivora and Rodentia) under different
ecological and geographical conditions. Praha, Academic Press, 159 pp.
Soltys A. 1952. Pasozyty wewnetrezne rujowki aksamitnei (Sorex araneus L.) Bialowieskiego Parcu Narodowego.
Annales Universitatis Mariae Curie-Skłodowska: S. T. C. 6 (5): 165-209.
Stammer H.J. 1955. Die Parasiten deutscher Kleinsauger. Verhandlungen der Deutschen Zoologischen Gesellschaft,
Erlangen 16 (6): 362-390.
40
CALODIUM SORICICOLA (YOKOGAWA ET NISCHIGORI, 1924)
(NEMATODA, CAPILLARIIDAE): ECOLOGICAL CHARACTERIZATION
OF NEMATODE INFECTION OF SHREWS IN SOUTH-WEST SIBERIA.
S. V. Karpenko, V. V. Panov
Keywords: Nematoda, Capillariidae, Calodium, infection, Soricidae shrew, Western Siberia
SUMMARY
The results of six-year studies (1983-1988) of shrew infestation with the nematode Calodium
soricicola (Yokogawa et Nischigori, 1924) (Nematoda, Capillariidae) - a parasite of liver parenchyma
of insectivorous mammals are presented. There are 6 species of the final nematode hosts (Sorex araneus
L., S. caecutiens Laxmann, S. daphaenodon Thomas, S. minutus L., S. tundrensis Merriam and Neomys
fodiens Pennant) detected in the subtaiga deciduous forest zone (northern regions of the Novosibirsk
Region). The intensity of invasion of all insectivores by C. soricicola in 1983-1988 was 6.6 ± 0.8 %
and changed from 3.2 ± 1.6 % in dry years to 9.1 ± 1.7 % in wet years. The most numerous and the most
contaminated was the Sorex araneus, 10.3 ± 1.1 %, which is the main host of C. soricicola in the shrews
community. Significant changes in its infestation by years were found - from 5.2 ± 1.7 % in dry years, to
17.4 ± 2.2 % in wet years. Other 5 species of shrews are classified as the second value hosts because of
their small number and low contamination. The stages and timing of development of C. soricicola and
C. hepaticum larvae were studied in laboratory conditions up to their invasive state. Infection of golden
hamsters with invasive larvae of both nematode species showed that they easily infected C. hepaticum and
did not infect by C. soricicola. The results obtained confirm the species independence of C. soricicola.
41