ПАРАЗИТОЛОГИЯ, 2021, том 55, № 1, с. 48-62.
УДК 595.771
РАЗЛИЧИЕ В РАСПРЕДЕЛЕНИИ ЗИМУЮЩИХ САМОК
КРОВОСОСУЩИХ КОМАРОВ ИЗ РОДОВ CULEX И CULISETA
(DIPTERA, CULICIDAE) В ПРИВХОДОВЫХ ЧАСТЯХ ПЕЩЕР:
СВЯЗЬ С РАЗНЫМИ ТРЕБОВАНИЯМИ К ТЕМПЕРАТУРЕ
И ВЛАЖНОСТИ ВОЗДУХА
© 2021 г. А. В. Разыграев*
Зоологический институт РАН,
Университетская наб., 1, Санкт-Петербург, 199034 Россия
Санкт-Петербургский государственный химико-фармацевтический университет,
ул. Профессора Попова, 14, лит. А, Санкт-Петербург, 197376 Россия
*е-mail: a.v.razygraev@gmail.com
Поступила в редакцию 25.05.2020 г.
После доработки 20.11.2020 г.
Принята к печати 29.11.2020 г.
В зимний период в привходовых частях Саблинских пещер (Тосненский район, Ленин-
градская обл.) измерены температура и влажность воздуха в местах максимального скопле-
ния зимующих взрослых самок Culex L., 1758 и Culiseta Felt, 1904. Показано, что самки рода
Culiseta (преимущественно, Cs. annulata) по сравнению с самками рода Culex (в основном
Cx. pipiens) занимают, как правило, более прохладные участки пещер при совместной зимовке
(средняя разница температур - 1.6 °C, p = 0.0039, парный критерий Уилкоксона; критерий Байеса
в пользу альтернативной гипотезы выше 40). Влажность воздуха, как правило, также ниже
в местах максимальных скоплений самок Culiseta по сравнению с таковыми для самок Culex
(в среднем на 9.4 %, р = 0.0051; критерий Байеса в пользу альтернативной гипотезы выше 20).
Подтверждается представление о том, что имаго рода Culiseta более адаптированы к низким
температурам и/или лучше переносят сниженную влажность воздуха. Вероятно, при понижении
температуры и влажности в привходовых частях пещер самки Culiseta более склонны оставаться
в тех участках убежищ, которые были заняты ими ранее, тогда как самки Culex перемещаются
в участки с более высокой температурой и влажностью воздуха.
Ключевые слова: кровососущие комары, зимовочные убежища, температура, влажность
воздуха, Culiseta annulata, Culex pipiens, Culicidae, Саблинские пещеры
DOI: 10.31857/S003118472101004X
Разные виды кровососущих комаров различаются по своим температурным тре-
бованиям, что отражается на их географическом распространении и на активности
имаго при различных температурах воздуха. В роде Culiseta Felt, 1904 известны виды,
адаптированные к низким температурам (WRBU). Согласно Becker et al. (2010), вид
48
Cs. annulata (Schrank, 1776), широко распространенный в Европе, более обычен
в северных широтах. Имаго сохраняют активность при низких температурах возду-
ха, например при +6 °C (Skierska, 1965; Wegner, 2009). Вид Cs. alaskaensis (Ludlow,
1906) также распространен в основном в областях с более прохладным климатом: он
типичен для тундры и бореальной зоны, а на юге тяготеет к горным регионам (Becker
et al., 2010). Многие виды рода Culex L., 1758 распространены в тропиках (Becker
et al., 2010). В Европе два близких вида подрода Culex - Cx. pipiens L., 1758 (подвид
pipiens, биотип “pipiens”) и Cx. torrentium Martini, 1925 - распространены симпа-
трично, но доля Cx. torrentium увеличивается в северных и горных регионах (Hesson
et al., 2014). Вероятно, климатические (в частности, температурные) предпочтения
этих двух видов различны.
В условиях северо-запада РФ Cx. pipiens pipiens (биотип “pipiens”), Cx. torrentium,
Cx. territans Walker, 1856, Cs. annulata, Cs. alaskaensis и Anopheles maculipennis s. l.
зимуют во взрослой фазе. Осенью оплодотворенные самки ищут зимовочные убежи-
ща, а самцы погибают до наступления зимы. При выполнении наших предыдущих
исследований осенью 2019 г. представители всех вышеупомянутых видов (и видовых
комплексов) были обнаружены в привходовых частях Саблинских пещер (Тосненский
р-н, Ленинградская обл.). С использованием подсчета самцов в убежищах в осенний
период было косвенно подтверждено, что при низких температурах (+8 °C и ниже)
имаго Cs. annulata более активны, чем имаго Cx. pipiens (Разыграев, 2020). Было также
замечено, что диапаузирующие самки рода Culex предпочитают скапливаться в пред-
положительно более теплых и менее продуваемых участках убежищ и чаще встре-
чаются на сводах пещер, чем на стенах, в противоположность самкам рода Culiseta.
Соответственно, участки максимальной плотности самок Culex и Culiseta зачастую
не совпадают друг с другом. Это значит, что гипотеза о различных температурных
требованиях имаго родов Culex и Culiseta может быть проверена путем сравнения
температур на участках с максимальной плотностью самок этих двух родов.
Цель настоящего исследования - проверка гипотезы о том, что в убежищах самки
рода Culiseta зимуют при более низких температурах воздуха, чем самки рода Culex.
Задачей было сравнить значения температуры (а также дополнительно значения влаж-
ности воздуха) на участках с максимальной плотностью зимующих самок рода Culex
и самок рода Culiseta в одних и тех же убежищах.
Материал и методика
Измерения температуры и влажности воздуха в зимних убежищах, в местах максимальных
скоплений самок родов Culex и Culiseta, проводились в Саблинских пещерах (окрестности пос.
Ульяновка, Тосненский р-н, Ленинградская обл.) в период с 10 января по 29 февраля 2020 г.
Вдоль левого берега р. Тосна обследованы пещеры Трехглазка (59°40´05.5˝ N, 30°47´41.4˝ E) и
Санта-Мария (59°39´41.1˝ N, 30°48´02.5˝ E). На правом склоне каньона р. Тосна обследованы
пещеры Жемчужная (координаты входа 59°40´06.4˝ N, 30°48´00.2˝ E) и Псевдо-Санта-Мария
(отрезанная обвалами часть пещеры Жемчужная, координаты входа 59°40´05.2˝ N, 30°48´02.9˝ E).
В пещерах Графский грот и Штаны, обследованных ранее (Разыграев, 2020), не было найдено
участков с достаточно высокой плотностью самок Culiseta (описание таких участков см. ниже),
поэтому попарные измерения в местах скопления самок Culiseta и Culex там не проводились.
Всего обследовано 11 ниш/участков в привходовых частях пещер (в частях, удаленных от вы-
хода менее чем на 50 м), в 4 из них были проведены также повторные попарные измерения
в разные дни (табл. 1).
49
Согласно предварительной оценке, температура в различных привходовых частях пещер
вариабельна при разных погодных условиях снаружи. Поэтому было решено одновремен-
но оценивать температуру и влажность воздуха на участках максимальной плотности самок
Culiseta и Culex, расположенных поблизости друг от друга (на расстоянии не более 2-3 м, кото-
рое соответствует ширине и глубине многих привходовых ниш) в одном и том же зимовочном
убежище. Ниже приведено подробное описание оригинальной методики сравнения участков,
занятых самками Culiseta и Culex, и подробно указаны причины ее использования.
Были замечены следующие особенности распределения самок Culiseta и Culex. Самки Culiseta
в основном встречаются разрозненно, поодиночке. При этом на многих участках их нет вовсе,
но иногда они образуют неплотные и, вероятно, неслучайные скопления, оцененные автором
как 3 особи на 272 см2 (272 см2 - это площадка с практически невариабельными температурой
и влажностью воздуха на ней, выделенная, например, квадратной рамкой 16.5×16.5 см). Также
встречаются 2 особи на указанную площадь, что в меньшей степени может считаться неслучай-
ным скоплением. Совсем редко число особей на указанную площадь может доходить до 6, что
явно может считаться неслучайным скоплением Culiseta при их общей немногочисленности.
Для гораздо более многочисленных самок Culex, напротив, на площадях такого размера нередки
скопления по несколько десятков особей.
Оптимальным решением для сравнения участков, занятых самками Culiseta и Culex, было
бы исследование распределений плотностей самок Culiseta и Culex по значениям температуры/
влажности и сравнение их между собой. Однако при предварительных измерениях стало ясно,
что для разных ниш и, тем более, разных пещер эти распределения будут разными для одного
и того же рода или вида комаров - вероятно, ввиду относительно быстрой динамики темпе-
ратуры/влажности в продуваемых привходовых частях и относительно низкой мобильности
диапаузирующих комаров, а также, возможно, различий ниш и пещер по другим, неизвестным
факторам, отражающимся на распределении комаров. Это, в принципе, означает несопостави-
мость распределений Culiseta с распределениями Culex из разных ниш и пещер, но оставляет
возможность попарных сравнений распределений Culiseta и Culex в одной и той же нише.
Полное изучение распределений Culiseta и Culex в каждой нише с последующим их попарным
сравнением представляет собой работу повышенной трудоемкости. При этом предсказуемо, что
результат, в случае Culiseta, будет малоинформативен: частоты встречаемости особей этого рода
для большинства значений температуры/влажности будут представлены значениями 0 и 1, лишь
иногда и далеко не для всех ниш - 3 и 4 и совсем редко - до 6 особей на выбранную площадь.
Оценить форму распределения Culiseta по значениям температуры/влажности в большинстве
ниш будет затруднительно или даже невозможно.
Технически приемлемой альтернативой явился быстрый метод: поиск такой ниши, в которой
присутствует скопление самок Culiseta с плотностью ≥ 3 особи на 272 см2, а также скопление
самок Culex (при этом визуально обнаружить наиболее плотное скопление Culex в нише не со-
ставляет труда), и измерение в этих скоплениях температуры и влажности. Если скопление самок
Culiseta обнаружено не в нише, а на стене просторного зала или коридора, то в качестве пары
можно использовать область максимальной концентрации самок Culex в пределах расстояния,
типичного для ширины и глубины ниши (2-3 м). Поиск максимальных скоплений проводится
до измерения температуры и влажности в убежище, а не после.
Очевидно, в случае нормальных распределений плотностей самок по значениям темпера-
туры и влажности для участков с максимальной плотностью характерны средние арифметиче-
ские и медианные значения температуры и влажности, а в случае умеренных отклонений от
нормальности - более близкие к медианам, чем к средним арифметическим. Медианы вполне
могут быть использованы для сравнения распределений Culiseta и Culex по температуре и
влажности в каждой отдельной нише вместо средних арифметических. Таким образом, бы-
стрый метод - это как минимум приблизительное сравнение распределений по их медианам
без детального изучения самих распределений. Даже если не рассматривать такие параметры
как средние значения, статистически значимое различие в температуре и/или влажности между
51
максимумами распределений Culiseta и Culex по площади поверхности ниши свидетельствует
о том, что распределения по температуре и/или влажности неодинаковы.
Таким образом, в соответствии с вышеописанным принципом, в обследуемой нише находили
участок максимального скопления самок рода Culiseta (Csmax, минимум 3 самки на площадь
272 см2; если в той же нише обнаруживался участок с более высокой плотностью, измере-
ние проводили именно в нем), затем в той же нише в пределах 2-3 м находили участок того
же размера с максимальной плотностью самок Culex (Cxmax). На обоих участках попеременно
в течение 5-10 мин регистрировали температуру и влажность при помощи метеометра МЭС-200,
помещая измерительный щуп в середину участка, занятого зимующими самками кровососущих
комаров, как показано на рис. 1. Полученные для каждого участка значения усредняли. Таким
образом, получали каждую отдельную пару значений одного и того же показателя для Csmax
и Cxmax. Все измерения в Csmax и Cxmax выполнялись в вечернее время с 17:30 до 22:00.
Рисунок 1. Измерение температуры и влажности воздуха на участке максимального
скопления самок Culiseta (3 особи на 272 см2) при помощи метеометра МЭС-200.
Измерительный щуп помещали в середину участка, занятого зимующими самками
кровососущих комаров.
Figure 1. Measuring of air temperature and humidity in the area of maximum density
of Culiseta females (3 individuals per 272 cm2) using meteometer MES-200. A measuring probe
was placed in the middle of the area occupied by overwintering mosquito females.
Большинство парных измерений выполнено в нишах, на стенах и сводах в одной и той
же привходовой камере пещеры Жемчужная, но в разных участках в разные дни, что означа-
ет возможность псевдорепликации значений, полученных при участии одних и тех же групп
самок. Тем не менее данный риск представляется небольшим, если исходить из общего числа
самок Culiseta в привходовой части пещеры Жемчужная (более полусотни особей, см. раздел
52
«Результаты»), числа особей в Csmax, как правило равного 3, и небольшого числа участков из-
мерений, где возможен обмен особями. Если принять во внимание парность измерений в Csmax
и Cxmax и общее количество самок Culex, исчисляемое тысячами на всю привходовую камеру
с нишами, то риск частичного повторного участия одной и той же пары групп (Culiseta и Culex)
в других участках становится еще ниже. Естественно, согласованная миграция одних и тех же
пар групп без изменения их состава в другие участки измерений совсем маловероятна, что
делает риск псевдорепликации разниц между Csmax и Cxmax совсем несущественным.
Также в период с декабря 2019 г. по март 2020 г. измеряли температуру и влажность воздуха
в глубине пещер (где комары не встречены) для сравнения с теми же показателями в привходовых
частях. При этом измерения проводили в середине полости пещеры на одинаковом расстоянии
от пола и свода. Результаты измерений температуры, выполненных в вечернее время с 17:30 до
22:00 за 5 дней в период зимовки комаров - 7 и 21 декабря 2019 г., 18 и 24 января и 6 марта
2020 г., - наносили на план-схему системы пещер «Жемчужная-Штаны».
Определение родовой принадлежности самок проводили по общим морфологическим
признакам (Гуцевич и др., 1970; Becker et al., 2010). Самок Cs. annulata отделяли от самок
Cs. alaskaensis непосредственно на месте по наличию предвершинных светлых колец на
бедрах и срединных светлых колец на первых члениках лапок (оба признака отсутствуют
у Cs. alaskaensis). Подсчет самок Cs. annulata и Cs. alaskaensis, как дополнение к опублико-
ванным ранее результатам по оценке пропорции между видами (Разыграев, 2020), проводили
путем сплошного обследования залов и коридоров, т.е. подсчитывали абсолютно всех особей
в выбранном зале или коридоре. Самок Culex для определения видов и оценки пропорции между
ними собирали равномерно по полостям пещер в начале и в конце зимы. Комаров собирали по
одному через приблизительно равные расстояния на стенах и потолках так, чтобы в целом из
отдельного зала с примыкающими к нему нишами было собрано 14-16 особей. При значительной
высоте потолка (зал в пещере Жемчужная с выходом в провал) сбор велся только на стенах.
В итоге в четырех залах было собрано 14, 16, 16 и 14 самок Culex в начале зимы (09.12.2019)
и в тех же залах 16, 15, 16 и 17 самок Culex в конце зимы (29.02.2020). В лаборатории самок
Cx. territans отделяли от Cx. pipiens/torrentium по апикальному расположению светлых пере-
вязей на тергитах брюшка (у Cx. pipiens/torrentium светлые перевязи располагаются базально)
(Гуцевич и др., 1970; Becker et al., 2010). Culex pipiens и Cx. torrentium разделяли по морфоме-
трическим характеристикам крыла, используя индекс r2/3 / r3, рассчитываемый по расстояниям
между точками ветвления и окончаниями ветвей радиальных жилок, как описано в ряде работ
(Börstler et al., 2014; Разыграев, Шулешко, 2018). Измерения проводились по правому крылу.
Статистическое сравнение значений температуры и влажности воздуха в местах макси-
мальных скоплений зимующих самок родов Culex и Culiseta проводили 1) с использованием
р-значений, полученных парным критерием Уилкоксона с поправкой Холма-Бонферрони
(поскольку на одних и те же парах площадей сравнения проводили по двум показателям)
и 2) с использованием байесовского критерия (Bayes factor) для парного t-теста. Расчеты пер-
вым способом проводились с помощью базовых функций программной среды R (версия 3.6.2)
(R Core Team, 2019). Для расчета байесовского критерия применяли байесовский калькулятор,
использующий программный пакет BayesFactor версии 0.9.8 в среде R версии 3.3.2 (Morey
et al., 2018); при этом предварительно убеждались в том, что в распределениях разниц значений
каждого из двух параметров (температуры и влажности) между Csmax и Cxmax не выявляется от-
клонение от нормального закона (по критерию Шапиро-Уилка). При наличии повторных данных
в разные дни в одной и той же части пещеры для статистических сравнений использовали те
пары значений для Csmax и Cxmax, которые в выборке из разных дней дают медианные значения
разницы между Csmax и Cxmax.
Коэффициенты корреляции Пирсона и Спирмена для оценки связи между температурой
и влажностью воздуха в Csmax и Cxmax высчитывали в программной среде R (версия 3.6.2)
(R Core Team, 2019). При вычислениях были использованы абсолютно все значения, в том числе
53
полученные повторно для одних и тех же участков, поскольку показания в них различались
в разные вечера.
Сравнения температуры и влажности воздуха в привходовых частях пещер с теми же по-
казателями в глубине пещер выполнялись с применением критерия Манна-Уитни-Уилкоксона
для независимых выборок (измерения были получены более или менее равномерно за зимний
период, но зачастую в разные дни в разных частях пещер, и во всех случаях не одномоментно,
а последовательно в разных направлениях в разные дни). При этом замеры для трех участков
в глубине пещер и для четырех привходовых частей объединялись в две выборки, n = 9 и 16
соответственно. В этих сравнениях применяли поправку Холма-Бонферрони, а также поправку
на непрерывность.
Отсутствие значимых различий в частотах среди общего числа самок Cx. pipiens и
Cx. torrentium в начале и в конце зимы проверяли байесовским критерием. Байесовский критерий
для таблицы сопряженности 2×2 (Cx. pipiens-Cx. torrentium, декабрь-февраль) рассчитывался
с использованием байесовского калькулятора (Dienes, 2008). При этом учитывались только
те особи, чей индекс r2/3 / r3 был меньше или равен 0.218 (сильное свидетельство в пользу
Cx. pipiens) и больше или равен 0.254 (сильное свидетельство в пользу Cx. torrentium) (Разыгра-
ев, Шулешко, 2018). Проверка отсутствия значимых различий в частотах среди общего числа
самок Cx. pipiens и Cx. torrentium в начале и в конце зимы служила для подтверждения того,
что один и тот же вид стабильно преобладает над другим видом за всё время зимовки. Гипо-
теза о том, что частоты двух видов одного рода неравны, проверялась точным биномиальным
тестом в среде R (нулевая гипотеза - вероятность обнаружения каждого из двух видов равна
0.5; альтернативная - вероятность больше или меньше 0.5).
Результаты
Проведенное обследование пещер и измерения температуры и влажности воздуха
в разных залах, штреках и штольнях показали, что в пещерах большой протяжен-
ности комары сем. Culicidae выбирают для зимовки не самые теплые участки, а те,
в которых регистрируются промежуточные значения температуры, т. е. участки, рас-
положенные ближе к выходам на поверхность (в системе «Жемчужная-Штаны» -
менее 30-40 м от выхода, рис. 2). На рис. 2 представлены результаты измерений
температуры внутри пещер за 5 дней, выполненных в период зимовки комаров при
наружной температуре от +0.6 до +2.0 °C. Температура в привходовых частях пещер
составляла от +3.4 до +7.4 °C и была значимо ниже, чем в глубине пещер, где значения
варьировали от +6.7 до +8.9 °C (р = 0.00029). Значения влажности воздуха, получен-
ные в те же дни (тоже в вечернее время), в привходовых частях варьировали от 67 до
99 %, а в глубине пещер - от 80 до 95 %. Диапазон влажности воздуха снаружи от
пещер в дни измерений составлял от 76 до 94 %. В сравнении значений влажности,
выполненном аналогично сравнению значений температуры, статистически значимое
различие не выявлено (р = 0.21).
В привходовых частях в период с декабря 2019 г. по март 2020 г. найдены самки
следующих видов и групп видов кровососущих комаров: Culex pipiens/torrentium,
Cx. territans, Culiseta annulata, Cs. alaskaensis и Anopheles maculipennis s. l. Самки
A. maculipennis s. l. в зимний период в Саблинских пещерах найдены впервые
(5 экземпляров в январе и феврале в пещерах Жемчужная и Санта-Мария). Замечено,
что в пещерах в указанный зимний период, по сравнению с предыдущими зимними
периодами, самки рода Culiseta были более многочисленны. В отношении гораздо более
многочисленных самок рода Culex в ряде мест внутри пещер замечена приурочен-
ность их скоплений к углублениям сводов (рис. 3), наряду с более-менее равномерным
(редким или плотным) распределением в нишах.
54
Рисунок 2. Температура воздуха в привходовых и глубинных частях системы пещер
«Жемчужная-Штаны».
Температуру измеряли в течение пяти дней в период с декабря 2019 г. по март 2020 г.
в вечернее время (с 17:30 до 22:00), указана в градусах Цельсия. Крупным шрифтом указана
медиана, мелким шрифтом (ниже) - размах min-max, в скобках - число измерений.
План пещер составлен на основе топосъемок С. Малахова и др. (2003 г.), К.Е. Васюкевича
и др. (2009-2011 г.) и карты из архива Ю.С. Ляхницкого (Малахов и др., 2010); уточнения
внесены А.В. Разыграевым и В.В. Гордеевым (2019-2020 г.).
Условные обозначения: жирные стрелки - входы в систему (в левой части - два входа
в пещеру Жемчужная, в средней - два входа в пещеру Псевдо-Санта-Мария, в правой
части - вход в пещеру Штаны), жирные черные точки - участки, на которых были найдены
зимующие кровососущие комары, * - температура снаружи от пещер (в вечернее время),
при которой регистрировались значения температуры внутри пещер.
Figure 2. Temperature values in the near-entrance and deep parts
of the “Zhemchuzhnaya-Shtany” cave system.
Measurements of temperature were performed during 5 days in an evening time (from 17:30
to 22:00) in the period from December, 2019, to March, 2020. Temperature is in Celsius’ degrees.
The medians are designated by a large font, the min—max range is designated by a font of smaller
size, the number of measurements in the same site is indicated in parentheses.
The scheme is drown up using cave maps originally performed by S. Malakhov et al. in 2003 and
K.E. Vasyukevich et al. in 2009-2011; the map from Yu.S. Lyakhnitsky’s archive was also used;
modifications were made by A.V. Razygraev and V.V. Gordeev in 2019-2020.
Designations: arrows - entries in the cave system (left area - two entries in the Zhemchuzhnaya
cave, central area - two entries in the Pseudo-Santa-Maria cave, right area - entry in the Shtany
cave), black dots - locations where overwintering mosquitoes were found, * - temperature outside
(in evening), when temperature inside caves was measured.
55
Рисунок 3. Скопление самок рода Culex в углублениях поверхности песчаника
в привходовой части пещеры Жемчужная.
Figure 3. Concentration of Culex females in cavities of the sandstone surface;
near-entrance part of the Zhemchuzhnaya Cave.
О резком преобладании Cs. annulata над Сs. alaskaensis среди самок в привходовых
частях Саблинских пещер в ноябре 2019 - начале января 2020 г. упоминалось ранее
(Разыграев, 2020). Дополнительный подсчет 18 января 2020 г. в привходовой части
пещеры Жемчужная (сплошное обследование) выявил 51 самку Cs. annulata и 3 самки
Сs. alaskaensis (соотношение 17 : 1), а в привходовой штольне пещеры Штаны, за-
крытой оползнем, - 15 самок Cs. annulata и 1 самку Сs. alaskaensis (соотношение 15
: 1). Гипотеза о том, что частоты Cs. annulata и Сs. alaskaensis равны, опровергается
для обеих пещер при высоких уровнях значимости (р = 2.919×10-12 и 5.188×10-4, соот-
ветственно; точный биномиальный тест). При равномерных сборах самок Cx. pipiens /
torrentium в четырех привходовых частях пещер в первой половине декабря 2019 г.
и в конце февраля 2020 г. были получены следующие результаты по соотношению
между численностью Cx. pipiens и численностью Cx. torrentium. По морфометри-
ческому индексу r2/3 / r3 из 123 собранных самок Cx. pipiens/torrentium 78 получили
сильное свидетельство в пользу их принадлежности к Cx. pipiens (BF01 > 10), а 29 -
к Cx. torrentium (BF01 < 0.1). Здесь BF01 - байесовский фактор в пользу гипотезы
Cx. pipiens, т. е. во сколько раз вероятнее, что особь принадлежит виду Cx. pipiens, нежели
Cx. torrentium (Разыграев, Шулешко, 2018). Остальные 16 особей получили слабое или
средней силы свидетельство в пользу одного из двух видов (0.1 < BF01 < 10)
- они
далее не рассматривались при расчете соотношения между Cx. pipiens и Cx. torrentium
(Разыграев, Шулешко, 2018). Самки, получившие сильное свидетельство в пользу
Cx. pipiens, преобладали над самками, получившими сильное свидетельство в пользу
Cx. torrentium, примерно в 2.7 раза (при этом предполагается, что среди 16 особей
с 0.1 < BF01 < 10 соотношение между двумя видами то же самое). Гипотеза о том, что
частоты двух видов равны, опроверглась с р = 2.352×10-6 (точный биномиальный тест
для 107 комаров, из которых 78 - Cx. pipiens, а 29 - Cx. torrentium). Декабрьские и
февральские результаты были объединены перед статистическим сравнением частоты
Cx. pipiens с частотой Cx. torrentium в общей численности, поскольку по соотношению
56
между Cx. pipiens и Cx. torrentium декабрьская и февральская выборки не различа-
ются (байесовский критерий в пользу нулевой гипотезы для таблицы сопряженности
«Cx. pipiens - Cx. torrentium, декабрь-февраль» равен приблизительно 3).
Из гистограммы на рис. 4 также хорошо заметно, что самки с небольшими значе-
ниями индекса (Cx. pipiens) явно преобладают над самками с высокими значениями
индекса (Cx. torrentium). Два максимума практически идентичны таковым, полученным
ранее для самок Cx. pipiens и Cx. torrentium в работе Börstler et al. (2014). Преобла-
дание Cx. pipiens над Cx. torrentium согласуется с данными, полученными на самцах
в сентябре-октябре 2019 г. для той же местности (Разыграев, 2020).
Рисунок 4. Частотная гистограмма распределения индекса r2/3 / r3 для самок Cx. pipiens /
torrentium, собранных в Саблинских пещерах 9 декабря 2019 г. и 29 февраля 2020 г.
Декабрьские и февральские результаты объединены (общее число особей составляет 123 шт.).
Частота представлена абсолютным числом особей.
Figure 4. Frequency histogram of the r2/3 / r3 index distribution for Cx. pipiens / torrentium females
collected in the Sablino caves in December 9, 2019 and February 29, 2020.
The December and February samples were combined with each other to present the data in
histogram (the total number of specimens is 123). Frequency is in absolute numbers of specimens.
Из 124 равномерно собранных самок рода Culex, среди которых проверялось со-
отношение между Cx. pipiens и Cx. torrentium, всего один экземпляр (из декабрьско-
го сбора в главной привходовой части пещеры Жемчужная) был представлен видом
Cx. territans. Осенью 2019 г. автор активно и целенаправленно собирал самок
Cx. territans в привходовой части пещеры Жемчужная для другого исследования, и,
вероятно, подавляющее большинство самок этого вида было собрано. Таким образом,
Cx. territans, третий вид из рода Culex, не вносил значимого вклада в общее число
зимующих самок рода Culex в обследованной части.
Проведенная оценка пропорций между видами показала, что среди самок Culiseta и
Culex, зимующих в Саблинских пещерах, преобладают виды Cs. annulata и Cx. pipiens.
57
Это означает, что дальнейшее сравнение Csmax и Cxmax (основное в данной работе) от-
носится к участкам максимальных скоплений самок Culiseta и Culex, представленных
преимущественно видами Cs. annulata и Cx. pipiens, соответственно.
В период с 10 января по 29 февраля 2020 г. проведены измерения температуры и
влажности воздуха в Csmax и Cxmax (Табл. 1) с последующим статистическим сравне-
нием Csmax и Cxmax по двум данным показателям. При попарном сравнении значений
температуры в Csmax и Cxmax в одних и тех же зимовочных убежищах (n = 11) выявилось
значимое различие: температура на участке с максимальной плотностью самок рода
Culiseta в среднем (и по медиане) на 1.6 °C ниже, чем на участке с максимальной плот-
ностью самок рода Culex (р = 0.0039 после применения поправки Холма-Бонферрони).
Аналогично, статистически значимо различаются значения влажности - почти во
всех случаях на участках максимального скопления самок рода Culiseta влажность
была ниже, чем на участках с максимальной плотностью самок рода Culex: средняя
разница для Csmax и Cxmax составила 9.4 %, медиана - 9.0 % (р = 0.0051). Байесовские
факторы в пользу гипотезы о различии между Csmax и Cxmax по температуре: Scaled JZS
Bayes factor - 47.20, Scaled-Information Bayes factor - 48.14. Аналогично по влажности:
Scaled JZS Bayes factor - 27.76, Scaled-Information Bayes factor - 31.25 (используемый
байесовский калькулятор на основе программного пакета BayesFactor рассчитывает
сразу 2 вида байесовского критерия, rscale = 0.707). Байесовские факторы превышают
значение 10, т. е. для обоих показателей (и температуры, и влажности) получены силь-
ные свидетельства в пользу альтернативной гипотезы (Kass, Raftery, 1995). Из Табл. 1
хорошо видно, что почти во всех случаях значения для Csmax ниже, чем для Cxmax,
и только в одном случае (помечено знаком *) разница между Csmax и Cxmax является
положительной по каждому параметру, т. е. когда значение температуры/влажности
для Csmax выше, чем для Cxmax. Результаты указывают на то, что в целом самки рода
Culiseta, по сравнению с самками рода Culex, действительно занимают более прохлад-
ные участки зимовочных убежищ, где зачастую отмечается более низкая влажность
воздуха.
Также между температурой и влажностью воздуха, измеренными в местах ско-
плений комаров, была выявлена статистически значимая связь: значения температу-
ры и влажности воздуха положительно коррелируют друг с другом, коэффициенты
корреляции Пирсона и Спирмена составили, соответственно, 0.62 и 0.65 при n = 34
(17 пар значений для Csmax и 17 пар значений для Cxmax, рис. 5) (p < 0.0001 для обоих
коэффициентов).
Обсуждение
В предыдущих работах мы упоминали, что самки кровососущих комаров относи-
тельно многочисленны в пещерах в начале зимовки, а в конце зимовки их численность
существенно сокращается. Так, данные обследования четырех тестовых площадок
показали, что в течение зимы 2018-2019 гг. количество самок Culiseta сократилось
в 6 раз, а самок Culex - в 6.77 раз (Razygraev, 2020). При этом в конце зимы и весной
самок Culiseta обнаружить было затруднительно, что связано, по-видимому, с их ис-
ходно меньшей численностью по сравнению с численностью самок Culex (Razygraev,
2020). В 2019-2020 гг. в течение всей зимы (в противоположность предыдущим зимам)
самки Culiseta оставались относительно многочисленными, что позволило найти до-
статочное количество участков их скоплений и оценить на этих участках температуру
и влажность воздуха.
58
Рисунок 5. Связь между значениями температуры и влажности воздуха, полученными
для мест скоплений комаров.
Условные обозначения: кружки с заливкой - значения для Csmax, кружки без заливки -
значения для Cxmax. Попарные связи между значениями см. в Табл. 1.
Figure 5. Correlation between air temperature and humidity measured in sites of maximum
concentration of hibernating mosquitoes.
Designations: filled circles - values for Csmax, not filled circles - values for Cxmax.
The links between Csmax and Cxmax given in Table 1.
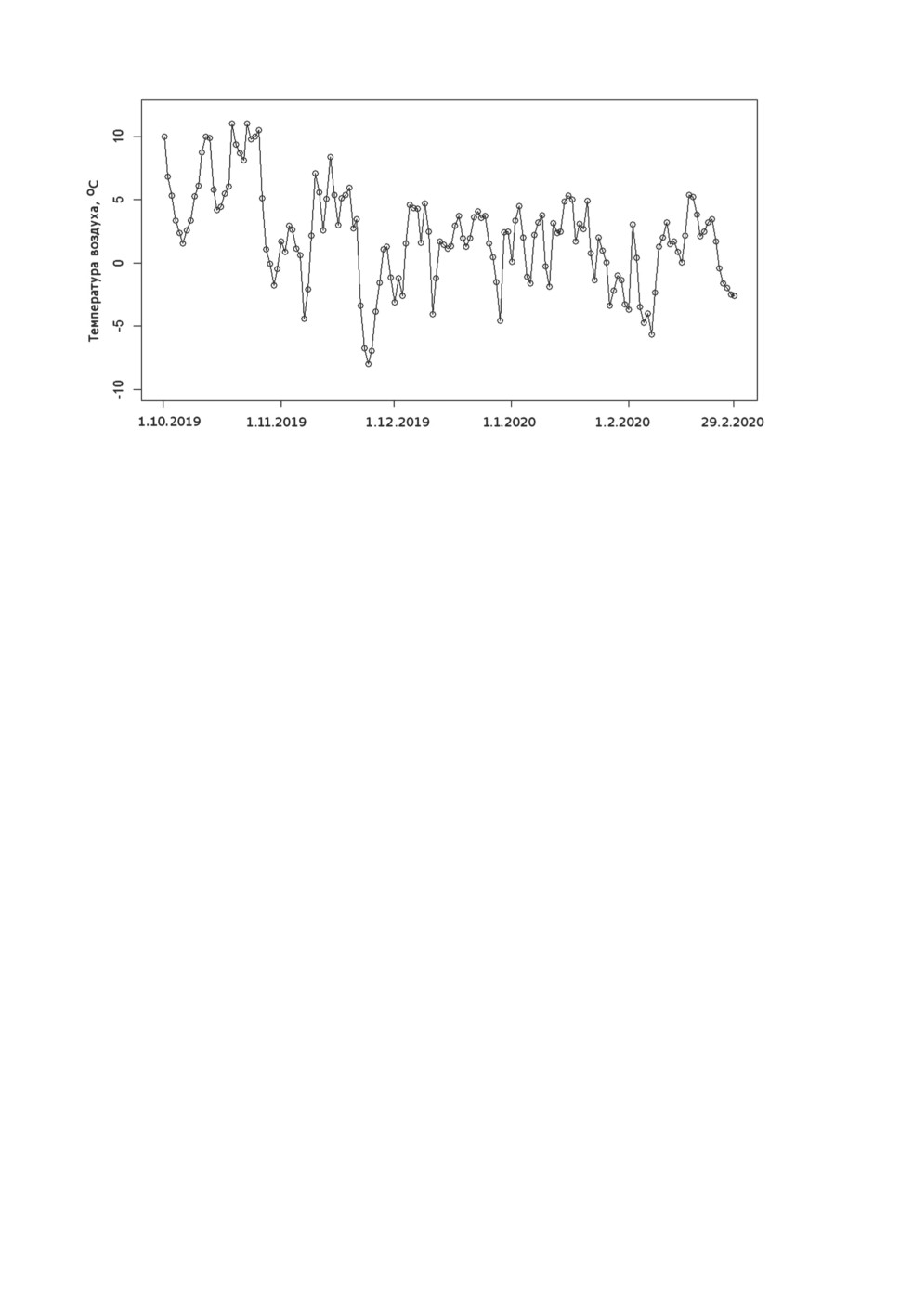
Большое количество самок Culiseta, а также первые находки в пещерах самок
Anopheles maculipennis s. l. в середине и в конце зимы, могут быть объяснены мягкой
зимой, без сильных и продолжительных морозов (рис. 6). Соответственно, привходовые
части пещер со времени ухода комаров на зимовку почти или совсем не проморажи-
вались. Это, возможно, объясняет высокую численность самок кровососущих комаров
в пещерах в целом за весь период проведения измерений в Csmax и Cxmax.
В исследовании, выполненном в сентябре-октябре 2019 г. на самцах, ищущих
убежища в пещерах при понижении температуры внешнего воздуха, было получено
статистическое обоснование различия в температурных требованиях между видами
кровососущих комаров из родов Culiseta и Culex. Самцы рода Culiseta (преимуще-
ственно Cs. annulata) были сравнительно малочисленны в привходовых частях пещер
при наружных температурах воздуха от +5 до +8 °C и сравнительно многочисленны
в тех же частях пещер при наружных температурах от +1 до +3 °C. В тех же интер-
валах наружных температур самцы Culex (преимущественно Cx. pipiens) в пещерах
одинаково многочисленны. При этом получено свидетельство в пользу сочетанного
влияния снижения влажности и температуры наружного воздуха, повышающего долю
самцов Culiseta среди общего количества самцов Culiseta и Culex в убежищах.
59
Рисунок 6. Среднесуточная температура воздуха с октября 2019 г. по февраль 2020 г.
по данным метеостанции в Пулково, южная часть Санкт-Петербурга (59.81°N, 30.27°E)
(The Weather Company, 2020).
Figure 6. Daily average air temperature from October 1, 2019 until February 29, 2020,
based on data from the Pulkovo meteo station (Southern Saint-Petersburg, 59.81°N, 30.27°E)
(The Weather Company, 2020).
Аналогично исследованию, выполненному на взрослых самцах, настоящее ис-
следование, проведенное на взрослых самках, также подтверждает различия в требо-
ваниях к температуре и/или влажности между Cs. annulata и Cx. pipiens, т. е. между
доминирующими видами из двух родов. Средняя разница температур между Cxmax
и Csmax составила всего 1.6 °C, однако результат статистически значимый. Можно
сделать вывод о небольших различиях в температурных требованиях между самками
Cs. annulata и Cx. pipiens. Различие по влажности воздуха составило 9.4 %, и оно
лишь незначительно уступает по статистический значимости различию в температуре
воздуха. Вероятно, незначительные по величине снижения температуры и влажности
в совместном убежище самок Culiseta и Culex заставляют самок Culex перемещаться
в более теплые и влажные места, тогда как самки Culiseta все еще склонны оставаться
в тех участках убежищ, которые были заняты ими ранее. Не исключено также, что
у самок Culex, уже находящихся в состоянии холодового оцепенения, несколько выше
смертность в местах с более низкой температурой и влажностью воздуха, что может
быть проверено в дальнейшем. Поскольку выявлена значимая положительная корреля-
ция между значениями температуры и влажности воздуха на участках скоплений самок
комаров, можно предполагать, что основным фактором, влияющим на распределение
самок, является только температура или только влажность воздуха, но возможно и их
сочетанное влияние.
Заключение
В проведенном исследовании показано, что самки рода Culiseta, зимуя в одних и тех
же убежищах с самками рода Culex, занимают, как правило, более прохладные участки
убежищ, имеющие зачастую более низкие значения влажности воздуха. Косвенно под-
60
тверждается более высокая приспособленность самок рода Culiseta (преимущественно,
Cs. annulata) к снижениям температуры и влажности по сравнению с самками рода
Culex (преимущественно, Cx. pipiens).
Благодарности
Работа выполнена частично при финансовой поддержке Государственной темы
«Разработка современных основ систематики и филогенетики паразитических и кро-
вососущих членистоногих» (гос. рег. № АААА-А19-119020790133-6), собранные и
определенные экземпляры комаров помещены в коллекцию Зоологического институ-
та РАН (ЗИН РАН, Санкт Петербург) (УФК ЗИН рег. № 2-2.20). Автор признателен
анонимным рецензентам за полезные замечания и вопросы: при ответах на них было
существенно расширено описание методов и результатов. Автор благодарит студен-
тов Института наук о Земле Санкт-Петербургского государственного университета
В.О. Бунина, С.Г. Каменкову и А.И. Гришнякову за помощь при измерении температуры
и влажности воздуха в привходовых и глубинных залах пещер.
Список литературы
Гуцевич А.В., Мончадский А.С., Штакельберг А.А. 1970. Комары. Семейство Culicidae. Фауна СССР. На-
секомые двукрылые, 3 (4), Ленинград, Наука, 384 c. [Gutsevich A.V., Monchadsky A.S., Stackelberg A.A.
1970. Fauna SSSR. Nasekomye dvukrylye (Fauna of the USSR. Insecta, Diptera), Vol. 3, Issue 4: Komary
sem. Culicidae (Mosquitoes, Family Culicidae), Leningrad: Nauka, 384 p. (in Russian)]
Малахов С., Шестакова К., Козлов А., Слепнева Т.Н. 2010. Саблино. Карты Саблинских пещер. Режим до-
Разыграев А.В. 2020. Численность самцов Culiseta и Culex (Diptera, Culicidae) в привходовых частях пещер
как отражение их различной устойчивости к низкой температуре наружного воздуха. Паразитология
54 (2): 137-144. [Razygraev A.V. 2020. Abundance of male mosquitoes of the genera Culiseta and Culex
(Diptera, Culicidae) in near-entrance parts of caves as a reflection of their different tolerance to low outdoor
temperatures. Parazitologiya 54 (2): 137-144 (in Russian)] DOI: 10.31857/S1234567806020042
Разыграев А.В., Шулешко Т.М. 2018. Использование фактора Байеса для определения видов Culex pipiens
и Culex torrentium (Diptera: Culicidae) по морфометрическим характеристикам крыла. Паразитология
52 (4): 304-314. [Razygraev A.V., Sulesco T.M. 2018. The use of Bayes factor for the identification of
Culex pipiens and Culex torrentium (Diptera: Culicidae) by morphometric wing characters. Parazitologiya
52 (4): 304-314 (in Russian)] DOI: 10.7868/S0031184718040054
Becker N., Petric D., Zgomba M., Boase C., Madon M., Dahl C., Kaiser A. 2010. Mosquitoes and Their Сontrol.
Second Edition. Berlin, Heidelberg, Springer-Verlag, 608 pp.
Börstler J., Lühken R., Rudolf M., Steinke S., Melaun C., Becker S., Garms R., Krüger A. 2014. The use of mor-
phometric wing characters to discriminate female Culex pipiens and Culex torrentium. Journal of Vector
Zoltan_Dienes/inference/Bayes.htm (17 апреля 2020)
Hesson J.C., Rettich F., Merdic E., Vignjevic G., Ostman O., Schäfer M., Schaffner F., Foussadier R., Besnard G.,
Medlock J., Scholte E.J., Lundström J.O. 2014. The arbovirus vector Culex torrentium is more prevalent
than Culex pipiens in northern and central Europe. Medical and Veterinary Entomology 28 (2): 179-186.
Kass R.E., Raftery A.E. 1995. Bayes factors. Journal of the American Statistical Association 90 (430): 773-795.
DOI: 10.1080/01621459.1995.10476572
Morey R.D., Rouder J.N., Jamil T., Urbanek S., Forner K., Ly A. 2018. BayesFactor: computation of Bayes fac-
(17 апреля 2020)
R Core Team 2019. R: A language and environment for statistical computing. Vienna, R Foundation for Statistical
61
Razygraev A.V. 2020. A comparative study of catalase activity in Culiseta annulata (Schrank) and Culex pipiens L.
Skierska B. 1965. Ecological studies of the occurrence and distribution of Culicinae fauna in the coastal forest
belt. Ekologia Polska Ser. A 13: 527-573.
The Weather Company. 2020. Weather Underground. Historical weather. Режим доступа: https://www.wunder-
ground.com/history (19 мая 2020)
Wegner E. 2009. The characteristics of the most troublesome mosquito species (Diptera: Culicidae) in Poland.
Fragmenta Faunistica 52 (2): 157-179. doi: 10.3161/00159301FF2009.52.2.157
DIFFERENCE IN THE DISTRIBUTION OF OVERWINTERING FEMALE
MOSQUITOES OF THE GENERA CULEX AND CULISETA (DIPTERA, CULICIDAE)
IN NEAR-ENTRANCE PARTS OF CAVES IN RELATION
TO AIR TEMPERATURE AND HUMIDITY
A. V. Razygraev
Keywords: mosquitoes, hibernacula, temperature, humidity, Culiseta annulata, Culex
pipiens, Culicidae, Sablino Caves
SUMMARY
Data on differences in ecological needs of mosquitoes of different genera and species at different
stages of the life cycle are important for the explanation of their distribution in regions with differ-
ent climate and can also be compared with differences in their physiological characteristics. In the
present paper, air temperature and humidity were measured in sites of maximum density of adult
mosquito females of the genera Culex and Culiseta, overwintering in near-entrance parts of Sablino
Caves (Northwestern Russia). Overwintering together, Culex females (predominantly Cx. pipiens)
concentrated, in general, in warmer places than Culiseta females (mainly Cs. annulata). The mean
difference in temperature between sites of maximum density of Culex and Culiseta females was
1.6 °C (p = 0.0039, Wilcoxon signed rank test; the Bayes factor in favor of alternative (BF10) was
more than 40). In general, humidity was also higher in sites of maximum density of Culex females
in comparison with those of Culiseta females - the mean difference constituted 9.4 % (p = 0.0051,
the BF10 is more than 20). These results support a hypothesis on the different temperature and/or hu-
midity requirements of Cx. pipiens and Cs. annulata: the latter species may be considered as better
adapted to low temperature and/or low humidity. Probably, when temperature and humidity fall
in near-entrance parts of caves, Culiseta females tend to stay in sites occupied earlier, while females
of Culex migrate to neighbor sites with higher temperature and humidity.
62