ПАРАЗИТОЛОГИЯ, 2021, том 55, № 2, с. 91-100.
УДК 576.311.344:595.121.3:597.593
LYSOSOMAL ENZYMES IN ADAPTIVE RESPONSES
OF CESTODES OF THE GENUS TRIAENOPHORUS
© 2021 R. U. Vysotskayaа, *, E. P. Ieshkoа, M. Yu. Krupnovaа,
L. V. Anikievaа, D. I. Lebedevaа
а Institute of Biology, Karelian Research Centre, Russian Academy of Sciences,
11 Pushkinskaya St., Petrozavodsk, 185910 Russia
*e-mail: vysotskayaru@gmail.com
Received 28.01.2021
Received in revised form 04.03.2021
Accepted 06.03.2021
The activity of seven acid hydrolases (acid phosphatase, DNase, RNase, β-glucosidase,
β-galactosidase, cathepsin В, cathepsin D) and tissue protein content were comparatively studied in
two cestode species of the genus Triaenophorus (T. crassus and T. nodulosus) sampled from pike
(Esox lucius) from Lake Kamennoye (northern Karelia). Differences between the lysosomal enzyme
profiles of these species were identified. Triaenophorus crassus demonstrated higher activities of
acid phosphatase and β-galactosidase. The activities of β-glucosidase, cathepsin В and DNase were
reliably lower than in T. nodulosus. The lower ecological plasticity of T. crassus and the differences
detected in the biochemical reactions in the two helminth species are indicative of a more strenuous
relationships of T. crassus with its definitive host, pike, in comparing with T. nodulosus.
Keywords: Triaenophorus, Esox lucius, host specificity, lysosomal enzymes
DOI: 10.31857/S0031184721020010
The cestode genus Triaenophorus Rudolphi, 1793 is chiefly represented in freshwater
bodies in Northern Europe, Siberia and North America by two species: Triaenophorus
nodulosus Pallas, 1781 and Triaenophorus crassus Forel, 1868 (Kuperman, 1973). Both
cestodes change three hosts over their complex life cycle. The first intermediate hosts for
larval stages of both parasites are Copepoda. The range of second intermediate hosts of
T. nodulosus is extensive, encompassing 57 fish species of 17 families. Plerocercoids of
this helminth are most often found in the liver of perch and ruffe. The diversity of second
intermediate hosts for T. crassusis much narrower, represented by 16 species of Salmonidae
and the related Osmeridae and Thymallidae (Kuperman, 1973). The definitive host for the
cestodes is the Northern pike Esox lucius L., in whose intestines the parasites mature and
complete their development (Kuperman, 1973).
91
Tapeworms possess some unique features they have acquired while adapting to the para-
sitic lifestyle (Dubinina, 1974). Morphofunctionally and biochemically, they are very well
adapted to their host (Shishova-Kasatochkina, Leutskaya, 1979; Sidorov et al., 1989). The
digestive system being reduced in cestodes, their tegument performs the essential secretory,
excretory, digestion and absorption functions. The helminth tegument is involved in many
physiological and biochemical processes that balance the host-parasite relationship (Davy-
dov, Mikryakov, 1988; Kuperman, 1988; Kuz’mina, 2005). The host organism responds to
helminth invasion by launching versatile protective mechanisms to minimize the damage
inflicted by the parasite (Izvekova, 2001; Sajid, McKerrow, 2002; Vysotskaya et al., 2003;
Dzik, 2006; Dezfuli et al., 2014; Nikishin, 2016).
Adaptations in intestinal cestodes are rendered more complex by the dual environment
of endoparasites, which have to adapt to the host (1st order environment) as well as respond
to changes in the host’s external environment (2nd order environment). Changes in the envi-
ronment disrupt the host’s food chains, wherefore facultative hosts get involved, resulting
in the formation of ”nonspecific parasitism” (Kuklin, Kuklina, 2005; Ieshko et al., 2012).
Over time, adaptation to new hosts results in speciation and emergence of new parasitic
systems (Kuperman, 1973). The mechanisms of the organism’s adaptation to the environ-
ment at the cellular level are built upon biochemical changes, including the reactions for
supplying the organism with matter and energy, metabolic regulation, and protection against
adverse impacts. An important role in the adaptive and protective responses of aquatic or-
ganisms belongs to lysosomal enzymes - special intracellular organelles containing several
dozens of acid hydrolases (Vysotskaya, Nemova, 2008). Information about the activity of
these enzymes in closely related species of the genus Triaenophorus and their participation
in the process of adapting to the host is in deficit.
The aim was to study in a comparative manner the activity of lysosomal enzymes in tissues
of adult cestodes Triaenophorus nodulosus and Triaenophorus crassus from pike intestines.
MATERIAL AND METHODS
Material for the study was sampled from northern Karelia, from Lake Kamennoye in the Kos-
tomukshsky Strict Nature Reserve (Kem River catchment, White Sea) in June, 2011. The 18 pike
specimens with body length (AC) ranged from 33 to 86 cm (62 ± 3) were investigated. The fish aged
2 - 14 years (7.6 ± 0.7) with body mass 328-5000 g (2184 ± 285).
The captured pikes were examined by partial helminthological dissection, with the prevalence (E)
and intensity (M) of infection with the cestodes T. nodulosus and T. crassus determined as suggested
by Bush et al. (1997). The fish were examined in June; all the retrieved cestodes were mature; data
on Triaenophorus infection rates are given in Table 1.
Pike liver was used for comparisons of biochemical indices in the parasitic systems, since this
lysosome-rich organ is actively involved in the host’s adaptive responses. Whole cestodes were taken
for the analyses. Tissue aliquots were rendered to 10% homogenates in 0.25 М sucrose solution with
EDTA and 0.1% Triton Х-100 non-ionic detergent, which destroys intracellular organelle membranes
releasing the enzymes contained therein. The samples were centrifuged at 10 000 g in centrifuges with
cooling. The supernatant fluid was analysed for the activity of seven lysosomal enzymes (acid phospha-
tase, DNase, RNase, β-glucosidase, β-galactosidase, cathepsin В, cathepsin D) and for protein content.
92
Analytical studies were done using equipment of the Core Facility of the Karelian Research Centre
of the Russian Academy of Sciences (Tissue Lyser LT homogenizer, Qiagen, Germany; Allegra 64R
centrifuge, Beckman Coulter, USA; spectrophotometer SF-2000, OKB-Spektr, Russia).
The substrate in determinations of the activity of acid phosphatase (EC 3.1.3.2) was sodium
β-glycerophosphate (Barrett, Heath, 1980). The enzyme activity was expressed in micrograms of
hydrolytically generated inorganic phosphorus, whose quantity was calculated based on its reac-
tion with chromogenic reagent (Kahovkova, Odavic, 1969). The activity of acid nucleases - DNase
(EC3.1.22.1) and RNase (EC3.1.4.23) - was determined as suggested by Pokrovskii and Archakov
(1968) and Levitskii et al. (1973), respectively. The substrates were deoxyribonucleic acid (рН 5)
and ribonucleic acid (рН 5.2) solutions in acetate buffer. Hydrolytic reaction products were quanti-
fied by spectrophotometry at 260 nm. The activity of the enzymes was expressed in relative units
D260. Determination of the activity of acid β-glucosidase (EC 3.2.1.21) was based on photometric
Δ
determination of the para-nitrophenol amount released by the reaction (Pokrovskii et al., 1971). The
substrate was a p-nitrophenyl-β-D-glucopyranoside solution in a citrate buffer (рН 5). The activity of
β-galactosidase (EC 3.2.1.23) was measured as suggested by Barrett and Heath (1980). The substrate
was sodium p-nitrophenyl-β-D-galactopyranoside (рН 4). The activity of both glycosidases was ex-
pressed in micromoles of p-nitrophenol per unit time per mg protein. The activity of acid proteases
was determined by modified spectrophotometric techniques: for cathepsin В (EC 3.4.22.1) - based on
break-up of 0.065 М Na-benzoyl-L-arginine ethyl ester solution in acetate buffer (рН 5), for cathepsin
D (EC 3.4.23.5) - based on the hydrolysis of 1% bovine haemoglobin in acetate buffer at рН 3.6
(Alekseenko, 1968). Protease activity was expressed in relative units of change in optical density (∆D)
per mg protein: cathepsin В - at 525 nm, cathepsin D - at 280 nm. Protein content in the samples
was determined according to the techniques suggested by Bradford (1976).
Data on cestode infection rates in pike were processed and analysed using Past software (Ham-
mer et al., 2001), differences between biochemical parameters were tested for reliability using the
Mann-Whitney U-test (Gubler, Genkin, 1969). Differences were considered significant with p ≤ 0.05.
RESULTS
All the examined pikes from Lake Kamennoye were quite intensively infected with the
cestodes Triaenophorus crassus and T. nodulosus (Table 1). The intensity of the T. nodulosus
(1-73) infection was much lower than the T. crassus (25-175) infection rates.
Table 1. Summary statistics of infection with cestodes of the genus Triaenophorus
in pike from Lake Kamennoye
Index
T. nodulosus
T. crassus
Number of examined pikes
18
18
Infection prevalence, %
100
100
Min intensity
1
25
Max intensity
73
175
Mean intensity
21.57
64
SE
4.89
13.23
Variance
334.88
2451.53
SD
18.30
49.51
Median
15.5
42
Variance/mean s2/M
15.52
38.31
93
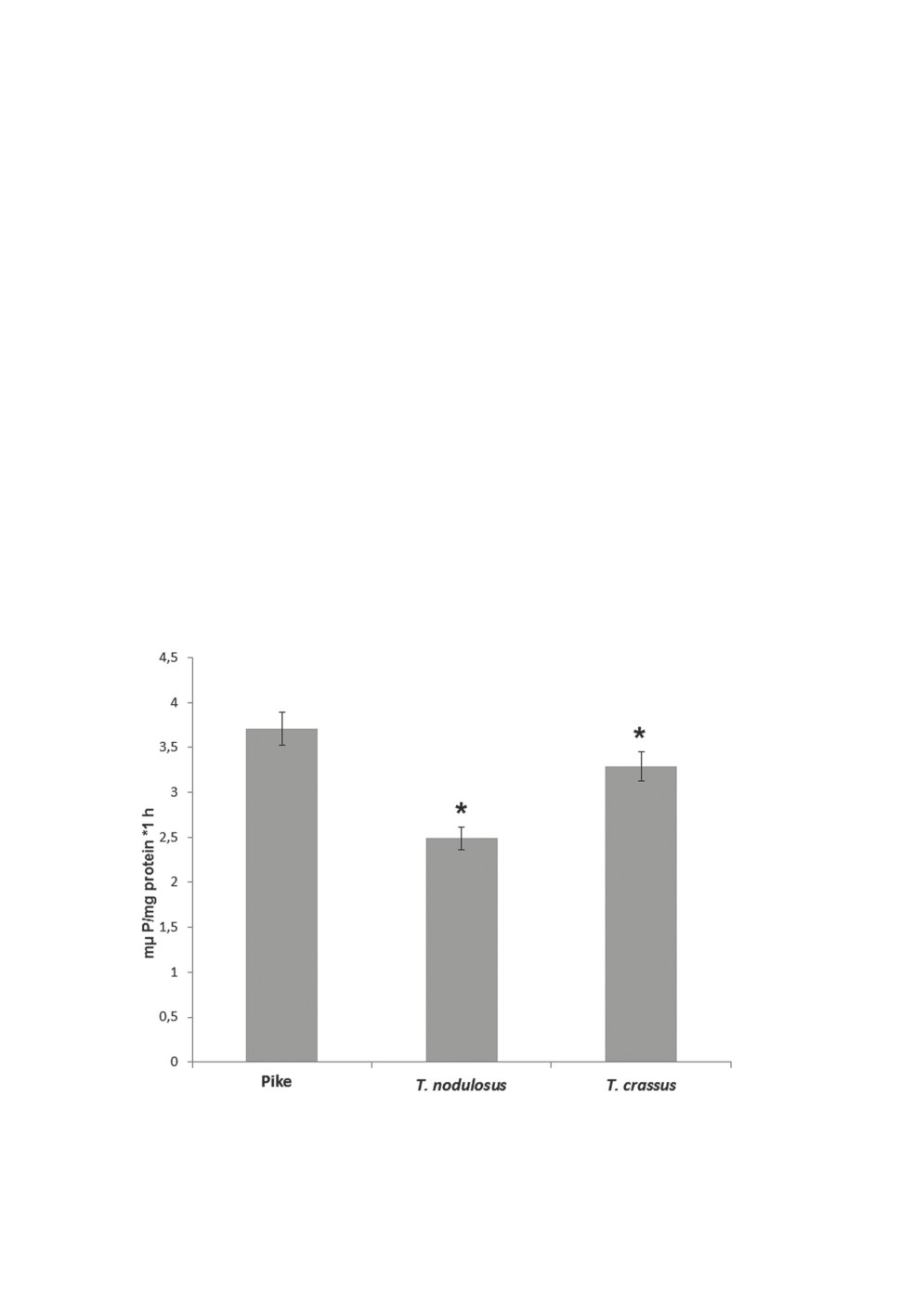
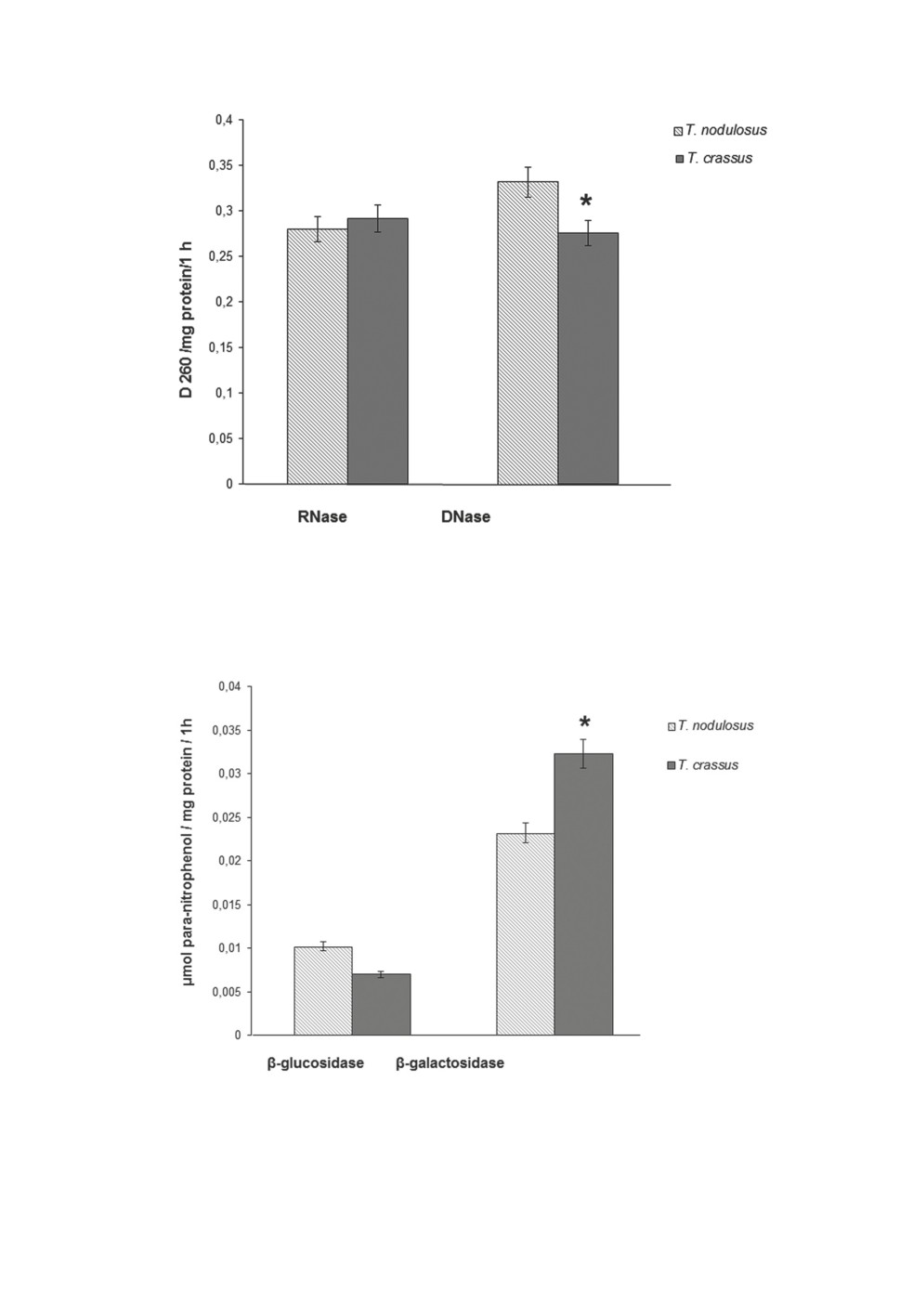
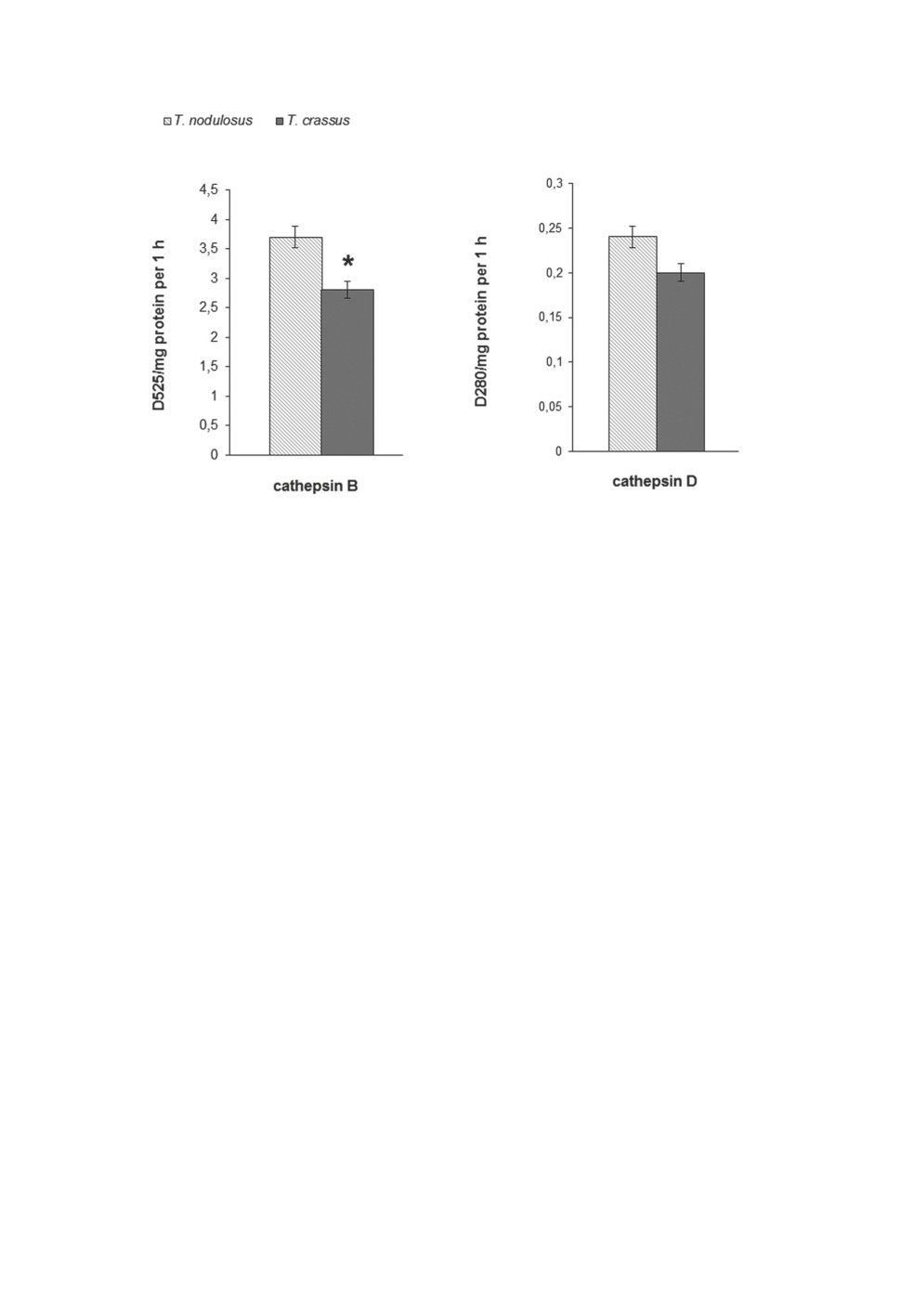
The results of the biochemical study of the cestodes are shown in Figures 1-4. It follows
from the reported data that the activity of lysosomal enzymes in parasite tissues was com-
mensurate with the respective indices in the host’s liver. This is indicated by the activity
of the lysosomal marker enzyme - acid phosphatase (Fig. 1). One should remark that the
activity of this enzyme in T. crassus was notably higher than in the other cestode.
The difference between T. nodulosus and T. crassus in tissue acid RNase activity was not
significant (Fig. 2), whereas the other nuclease - DNase, was significantly lower in T. crassus.
The most significant differences between the cestodes concerned the activity of glyco-
sidases (Fig. 3). Firstly, the absolute values of β-galactosidase activity in both worms were
significantly higher than those of β-glucosidase. Secondly, galactosidase activity in T. crassus
was higher and glucosidase activity was lower than in T. nodulosus.
The same was true for the activity of both proteases (Fig. 4). Their levels in T. crassus
were lower than in T. nodulosus. Note also that cathepsin B activity in cestodes was 2-3
times higher than in the host’s liver (1.02±0.04 relative units - ∆D525/mg protein per hour).
On the contrary, the cathepsin D activity of cestodes was 4 times lower than in the pike’s
liver (0.91±0.05 relative units - ∆D280 /mg protein per hour). The variation of soluble protein
content in the samples was not so significant: this index in T. nodulosus tissues was 74.5 ±
3.7, in T. crassus - 77.5 ± 3.2, and in pike liver - 62.7 ± 4.4 mg/g dry mass.
Figure 1. Acid phosphatase activity in tissues of adult cestodes of the genus Triaenophorus
and the liver of their host - pike. n = 5.
*Differences between variants are significant at p≤0.05.
94
Figure 2. Nuclease activity in tissues of adult cestodes of the genus Triaenophorus. n = 5.
*Differences between variants are significant at p≤0.05.
Figure 3. Glycosidase activity in tissues of adult cestodes of the genus Triaenophorus. n = 5.
*Differences between variants are significant at p≤0.05.
95
Figure 4. Protease activity in tissues of adult cestodes of the genus Triaenophorus. n = 5.
*Differences between variants are significant at p≤0.05.
DISCUSSION
It seems a highly aggregated distribution, indicated by the s2/M ratio, was demonstrated
by T. crassus (Table 1), suggesting pike was more susceptible to infestation by this species
as compared to T. nodulosus.
Cestodes of the genus Triaenophorus are systematically close species. The ranges of
distribution of both T. nodulosus and its host of the genus Esox are almost the same. The
distribution area of T. crassus is somewhat narrower. It occupies the northern part of pike’s
range and covers circumpolar Holarctic regions. The definitive and the first intermediate
hosts are the same for both helminth species. The most significant difference between them
is the localisation of plerocercoids in the second intermediate host: muscles of salmoniform
fishes for T. crassus, and usually liver of Percidae for T. nodulosus. Adaption to the environ-
ment inside their respective second intermediate hosts has been the key factor for the diver-
gence of these cestode species (Kuperman, 1973). The differences detected in the adaptive
responses of the helminths in our study also suggest that T. crassus is a stricter definitive
host specialist than T. nodulosus. The leading role in the species’ adaptations belongs to
biochemical changes, including changes in the lysosomal enzyme complex activity. These
enzymes are involved in membrane and cellular digestion processes (Vysotskaya, Nemova,
2008). Cestode tegument is rich in structures participating in lysosome formation (Kuperman,
1988). A higher activity of acid phosphatase in T. crassus tissues compared to T. nodulosus
indicates that the former generates more lysosomes and that the costs of its adaptation to
96
the host are higher. Acid phosphatase is known as a broad-spectrum phosphoric monoester
hydrolase, which has an important role in the metabolism of carbohydrates, lipids, nucleic
acids and phosphorus compounds and, hence, in supplying the organism with energy.
Another enzyme whose activity was significantly higher in T. crassus than in the other
cestode was β-galactosidase. The level of this glycosidase can be elevated when the para-
site’s adaptive reactions involve galactose-containing lipids and proteoglycans, which act
as metabolic regulators (Vdovichenko, Vysotskaya, 2013). Also, considering that carbohy-
drate metabolism is the principal source of energy for helminths, when the stores of energy
substrates have mostly been exhausted, alternative mechanisms can be launched to support
tissue bioenergy, and then the role of lysosomal glycosidases, including β-galactosidase,
will grow (Vysotskaya, Nemova, 2008).
We have previously demonstrated that the activity of the lysosomal protease cathepsin B
in tissues of T. nodulosus cestodes is several times higher than in pike organs (Vysotskaya
et al., 2015). The lysosomal proteolytic system is the main player in protein metabolism.
On top of cleaving proteins to peptides and amino acids, lysosomal proteases perform
a number of specific functions for renewal of proteins, activation of precursors of biologi-
cally active proteins and peptides, including hormones (Turk et al., 2001; Buhling et al.,
2004; Nemova, Bondareva, 2005). Cathepsin В participates in the degradation of many
intra- and extracellular proteins (Brix et al., 2008; Arampatzidou et al., 2011; Yadati et al.,
2020). Cathepsin B was shown to take part in apoptosis and immunoregulation processes
(Turk, Turk, 2009). The high activity of cathepsin B we observed in the tissues of both
cestodes suggests that this enzyme is actively involved in the parasites’ protective response
to impact from the host. Probably that activity of cathepsin B in T. crassus was significantly
lower than in T. nodulosus due to the effect of the peptide antibiotics the host produces to
protect itself against various infection agents, including invasion by helminths (Dezfuli et al.,
2014). Besides, proteases, as well as other acid hydrolases, can be inhibited by own proteins
produced by the parasite to protect against the host’s proteolytic enzymes and excreted at
host-helminth contact sites (Holt et al., 2006; Chen et al., 2017; Izvekova, Frolova, 2019;
Vidak et al., 2019). The assumption that the cestodes in question can respond differently to
the same impact from the host is supported by the recently obtained data on qualitative and
quantitative differences in the protein composition of these species (Kochneva et al., 2018).
Thus, according to the biochemical aspects of host-parasite relationships, as well as
data on the distribution, life cycles, and host affiliations, T. crassus appears to be a stricter
specialist than T. nodulosus. It is adapted to living in cold oligotrophic waters. Its first in-
termediate hosts are northern copepodite species. The parasite’s distribution in water bodies
is mainly associated with vendace - a very common species in northern lakes (Potapova,
1978; Valtonen et al., 1989). The other species - T. nodulosus has a broader distribution
and a much wider range of second intermediate hosts, including salmoniform fishes. The
key role in maintaining T. nodulosus abundance belongs to perch and ruffe - the main
items in pike’s diet (Kuperman, 1973). Differences in ecological valence and specialisation
between Triaenophorus species are the reasons for the different resilience of their parasitic
systems through the natural succession in water bodies and under human impact. To wit,
97
Lake Kostomukshskoye, contaminated by wastes from iron-ore mine and mill, exhibits
a poorer species composition of the biota, including a drastic reduction of the fish fauna.
Vanishing of vendace, the main intermediate host for T. crassus, from the lake entails the
extinction of the parasite, in spite of the presence of other aquatic organisms involved in
its life cycle. In this situation, the decline of aquatic animal diversity similarly leads to
changes in the structure of the T. nodulosus parasitic system in line with changes in the
host’s trophic links, since the main intermediate host (perch) is also absent from the lake.
A typical intermediate host is recruited into the parasite’s life cycle (Ieshko et al., 2012).
Research into the biochemical aspects of relationships in the T. nodulosus-pike parasitic
system in a water body altered by human activities has demonstrated that the main contribu-
tor to the adaptive response to the adverse environmental impact is the host. The parasite,
on the other hand, also contributes to overall homeostasis in the system by adjusting its
metabolism to the host’s condition (Vysotskaya et al., 2015). It is there for ease to say that
the host-parasite relationship in the T. nodulosus-pike system is highly balanced at the fine
biochemistry level. The fact that of the two cestode species of the genus Triaenophorus
only T. nodulosus occurred in the technogenic suppressed lake suggests that this parasitic
system has a higher adaptive potential than that of T. crassus.
Our studies revealed differences between the biochemical and population parameters
of the two cestode species, which evidence a more strenuous process of adaptation to the
host in T. crassus. This, together with a narrower range of second intermediate hosts and
a lower ecological plasticity, corroborates the assumption about a later speciation of
T. crassus compared to T. nodulosus. Of the two ancient and steady parasitic systems, the
original one is the T. nodulosus - pike Esox lucius L. system.
ACKNOWLEDGEMENTS
The studies were financed from the federal budget under state assignments to KarRC
RAS ( No. 0218-2019-0075, 0218-2019-0076 and АААА-А17-117031710039-3).
REFERENCES
Alekseenko L.P. 1968. Methods for determination of activity of proteolytic enzymes. In: Orekhovich V.N. (ed.).
Modern Methods in Biochemistry. Moscow, Meditsina, 115-130. [in Russian]
Arampatzidou M., Rehders M., Dauth S., Yu D.M.T., Tedelind S., Brix K. 2011. Imaging of protease functions-
current guide to spotting cysteine cathepsins in classical and novel scenes of action in mammalian epithelian
cells and tissues. Italian Journal of Anatomy and Embryology 116 (1): 1-19.
Barrett A.J., Heath, M.F. 1980. Lysosomal enzymes. In: Dingle J.T. (ed.). Lysosomes - A Laboratory Handbook.
Amsterdam, North Holland Publishing Co, 25-156.
Bradford M.M. 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing
the principle of protein-dye binding. Analytical Biochemistry 72: 248-254.
Brix K., Dunkhorst A., Mayer K., Jourdans S. 2008. Cysteine cathepsins: cellular roadmap to different functions.
Biochimie 90: 194-207.
Buhling F., Waldburg N., Reisenauer A., Heimburg A., Golpon H., Welte T. 2004. Lysosomal cysteine proteases
in the lung: role in protein processing and immunoregulation. European Respiratory Journal 23: 620-628.
Bush A.O., Lafferty K.D., Lotz J.M., Shostak A.W. 1997. Parasitology meets ecology on its own terms: Margolis
et al. revisited. Journal of Parasitology 83 (4): 575-583.
Chen L., He B., Hou W., He L. 2017. Cysteine protease inhibitor of Schistosoma japonicum - A parasite-derived
negative immunoregulatory factor. Parasitology Research 116 (3): 901-908.
98
Gubler E.V., Genkin, A.A. 1969. Application of Nonparametric Statistics Criteria to Assess the Differences between
Two Groups of Observations in Biomedical Research. Moscow, Meditsina, 29 pp. [in Russian]
Davydov V.G., Mikryakov V.R. 1988Adaptive structures of the integument of some cestodes related to the protection
of parasites from the effects of the host organism. In: Sonin M.D. (ed.). Immunological and Biochemical
Aspects of the Relationships between Helminth and Host. Moscow, Nauka, 88-100. [in Russian]
Dezfuli B.S., Giari L., Lorenzoni M., Manera M., Noga E.J. 2014. Perch liver reaction to Triaenophorus nodulosus
plerocercoids with an emphasis on piscidins 3, 4 and proliferative cell nuclear antigen (PCNA) expression.
Veterinary Parasitology 200: 104-110.
Dubinina M.N. 1974. State and the next tasks of systematics of tapeworms (Cestoidea Rud., 1808). Parazitologiya
8 (4): 281-292. [in Russian]
Dzik J.M. 2006. Molecules released by helminth parasites involved in host colonization. ActaBiochemicaPolonica
53 (1): 3-64.
Hammer Ø., Harper D.A.T., Ryan P.D. 2001. PAST: Paleontological statistics software package for Education
Holt O.J., Gallo F., Griffiths G.M. 2006. Regulating secretory lysosomes. Journal of Biochemistry 140 (1): 7-12.
Ieshko E.P., Lebedeva D.I., Anikieva L.V., Ilmast N.V. 2012. Population biology of cestodes of genus Triaenophorus
in natural and man-made water bodies. Parazitologiya 46 (6): 434-443. [In Russian]
Izvekova G.I. 2001. Physiological specificity of the interaction between Traenophorus nodulosus (Cestoda) and
its host - fishes. Parazitologiya 35 (1): 60-68. [in Russian]
Izvekova G.I., Frolova T.V. 2019. Certain characteristics of trypsin activity inhibition by cestodes Triaenophorus
nodulosus and Eubothrium rugosum. Parazitologiya 53 (1): 73-81. [in Russian]
Kahovkova J., Odavic R. 1969. A simple method of the quantitative analysis of phospholipids separated by thin
layer chromatography. Journal of Chromatography 40 (1): 90-96.
Kochneva A., Borvinskaya E., Bedulina D. 2018. The study of protein composition of Triaenophorus sp. at dif-
ferent stages of the life cycle and in different body segments. Bioinformatics of Genome Regulation and
Structure Systems Biology (BGRS\SB-2018): The Eleventh International Conference (20-25 Aug. 2018,
Novosibirsk, Russia); Abstracts/ Novosibirsk State University. Novosibirsk: ICGSB RAS, 2018. DOI:
10.18699/BGRSSB-2018-081
Kuzmina V.V. 2005. Physiological and biochemical basis of exotrophy in fish. Moscow, Nauka, 302 pp. [in Russian]
Kuklin V.V., Kuklina M.M. 2005. Helminths of Birds of the Barents Sea: Fauna, Ecology, and Impact on Hosts.
Apatity, Kol’sky Nauchny Tsentr RAS, 289 pp. [in Russian]
Kuperman B.I. 1973. Tapeworms of the Genus Triaenophorus - Fish Parasites. Leningrad, Nauka, 208 pp. [in
Russian]
Kuperman B.I. 1988. Functional morphology of lower cestodes: ontogenetic and evolutionary aspects. Leningrad,
Nauka, 168 pp. [in Russian]
Levitskii A.P., Barabash R.D., Konovets V.M. 1973. Seasonal characteristics of ribonuclease and α amylase activ-
ity of saliva and salivary glands in Wistar rats. In: Kreps E.M. (ed.). Biochemical Evolution, Leningrad:
Nauka, 192-195. [in Russian]
Nemova N.N., Bondareva L.A. 2005. Proteolytic Enzymes. Petrozavodsk, KRC RAS, 92 pp. [in Russian]
Nikishin V.P. 2016. Morphofunctional diversity of glycocalyx in tapeworms. Advances in Modern Biology 136
(5): 506-526. [in Russian]
Pokrovskii A.A., Archakov A.I. 1968. Methods of separation and enzymatic identification of subcellular fractions.
In: Orekhovich V.N. (Ed.). Modern Methods in Biochemistry. Moscow, Meditsina, 5-59. [in Russian]
Pokrovskii A.A., Kravchenko L.V., Tutel’yan V.A. 1971. The study of activity of lysosomal enzymes in the pres-
ence of aflatoxin and mitomycin C. Biochemistry 36 (4): 690-696. [in Russian]
Potapova O.I. 1978. Large vendace Coregonus albula L. Leningrad, Nauka, 133 pp. [in Russian]
Sajid M., McKerrow J.H. 2002. Cysteine proteases of parasitic organisms. Molecular and Biochemical Parasitol-
ogy 120: 1-21.
Shishova-Kasatochkina O.A., Leutskaya Z.K. 1979. Biochemical aspects of the relationship between helminth
and host (exchange of proteins, vitamins and steroids in the process of parasitism). Moscow, Nauka, 152
pp. [in Russian]
Sidorov V.S., Vysotskaya R.W., Smirnov L.P., Guryanova S.D. 1989. Comparative biochemistry of fish helminths.
Amino acids, proteins, lipids. Leningrad, Nauka, 152 pp. [in Russian]
99
Turk B., Turk V. 2009. Lysosomes as “suicide bags” in cell death: myth or reality? Journal of Biological Chemistry
284: 21783-21787.
Turk V., Turk B., Turk D. 2001. Lysosomal cysteine proteases: facts and opportunities. The EMBO Journal 20:
4629-4633.
Valtonen E.T., Rintamӓki P., Lappalainen M. 1989. Triaenophorus nodulosus and T. crassus in fish from Northern
Finland. Folia Parasitologica 26: 351-370.
Vdovichenko E.A., Vysotskaya R.U. 2013. The comparative characteristics of the activity of lysosomal glycosi-
dases of pikes living in water objects with different rates of anthropogenic load. Fundamental research 4
(5): 1134-1138. [in Russian]
Vidak E., Javoršek U., Virovišek M., Turk B. 2019. Cysteine cathepsins and their extracellular roles: shaping the
microenvironment. Cells 8 (3): 264-273.
Vysotskaya R.U., Ieshko E.P., Evseeva N.V. 2003. A comparative biochemical research in the Schistocephalus
solidus (Cestoda) — three-spined stickleback Gasterosteus aculeatus L. system. Parazitologiya 37: 503-511.
[in Russian]
Vysotskaya R.U., Krupnova M.Y., Ieshko E.P., Anikieva L.V., Lebedeva D.I. 2015. Ecological and biochemical
aspects of parasite-host interactions in transformed aquatic bodies: a case study of the cestode Triaenophorus
nodulosus and its host, the northern pike Esox lucius. Biology Bulletin 42 (3): 246-253.
Vysotskaya R.U., Nemova N.N. 2008. Lysosomes and lysosomal enzymes of fish. Moscow, Nauka, 284 pp. [in
Russian]
Yadati T., Houben T., Bitorina A., Shiri-Sverdlov R. 2020. The ins and outs of cathepsins: physiological function
and role in disease management. Cells 9 (7): 1679-1705.
ЛИЗОСОМАЛЬНЫЕ ФЕРМЕНТЫ
В АДАПТИВНЫХ РЕАКЦИЯХ ЦЕСТОД РОДА TRIAENOPHORUS
Р. У. Высоцкая, Е. П. Иешко, М. Ю. Крупнова,
Л. В. Аникиева, Д. И. Лебедева
Ключевые слова: Triaenophorus, Esox lucius, гостальная специфичность, лизосо-
мальные ферменты
РЕЗЮМЕ
Проведено сравнительное изучение активности семи кислых гидролаз (кислой фосфатазы,
ДНКазы, РНКазы, β-глюкозидазы, β-галактозидазы, катепсина В, катепсина D) и содержания
белка в тканях двух видов цестод рода Triaenophorus (T. crassus и T. nodulosus) из щуки озе-
ра Каменного (Северная Карелия). Установлены различия в ферментных профилях лизосом
у изученных паразитов. Для Triaenophorus crassus характерны более высокие значения активности
кислой фосфатазы и β-галактозидазы, активность же β-глюкозидазы, катепсина В и ДНКазы была
более низкой, чем у T. nodulosus. Выявленные различия в показателях заражения и параметрах
распределения численности цестод в популяции хозяина (Esox lucius Linnaeus, 1758) свидетель-
ствуют о более напряженном приспособительном процессе к хозяину у T. crassus, что наряду
с более узким кругом вторых промежуточных хозяев и меньшей экологической пластичностью
подтверждает более позднее происхождение этого вида по сравнению с T. nodulosus.
100