ПАРАЗИТОЛОГИЯ, 2021, том 55, № 4, с. 267-286.
УДК 576.895
СТРОЕНИЕ ОРГАНА ГАЛЛЕРА И СИСТЕМАТИКА
ИКСОДОВЫХ КЛЕЩЕЙ (СЕМЕЙСТВО IXODIDAE)
ПОДСЕМЕЙСТВА AMBLYOMMINAE
© 2021 г. С. А. Леонович*
Зоологический институт РАН,
Университетская наб. 1, Санкт-Петербург, 199034 Россия
*e-mail: leonssa@mail.ru
Поступила в редакцию 21.04.2021 г.
После доработки 28.04.2021 г.
Принята к публикации 15.05.2021 г.
На основании собственных и литературных данных проведен анализ строения органа
Галлера, главного рецепторного органа иксодовых клещей, изученного методами растровой
электронной микроскопии, у 40 видов клещей подсемейства Amblyomminae (Metastriata), пред-
ставителей родов Amblyomma Koch, 1844 (10 видов, включая 3 вида, ранее выделявшихся
в род Aponomma), Anomalohimalaya Filippova, 1994 (1 вид), Dermacentor Koch, 1844 (6 видов),
Haemaphysalis Koch, 1844 (6 видов), Hyalomma Koch, 1844 (6 видов), Margaropus Pomerantzev,
1947 (1 вид) и Rhipicephalus Koch, 1844 (10 видов, включая 2 вида, ранее выделявшихся
в род Boophilus). Выделены основные таксономические признаки (число и топография сенсилл
в передней ямке и форма капсулярного отверстия). Показано, что строение органа Галлера
специфично для отдельных родов, в то время как подродовые и видовые различия в пределах
каждого рода практически не выражены. На этом основании последние ревизии подсемейства,
основанные исключительно на молекулярно-генетических данных, подвергаются сомнению.
Проведено сравнение полученных данных с существующими представлениями о таксономии
подсемейства Amblyomminae.
Ключевые слова: орган Галлера, растровая электронная микроскопия, Ixodidae, Metastriata,
Amblyomminae, систематика
DOI: 10.31857/S0031184721040013
Орган Галлера, впервые описанный естествоиспытателем Галлером в 1881 г.
(Haller, 1881), - основной чувствительный орган иксодовых клещей, переносчиков
опасных трансмиссивных заболеваний. У клещей, как известно, нет антенн - при-
датков головной капсулы, несущих основную массу сенсорных образований, как это
267
наблюдается у насекомых, поэтому роль антенн насекомых у клещей выполняют
передние ноги. Клеши перемещаются на трех парах ног, а передняя пара ног слу-
жит им своеобразными аналогами антенн насекомых. Именно поэтому на передних
ногах, а точнее, на их дистальных сегментах, лапках, располагаются основные ор-
ганы чувств, позволяющие обнаруживать источник пищи или полового партнера.
У гамазовых клещей - это тарзальный рецепторный орган (Leonovich, Dimov, 2012),
а у иксодовых клещей - это орган Галлера. Кроме того, у большинства клещей от-
сутствуют глаза (Леонович, 2005)
Орган Галлера - чрезвычайно сложно устроенный орган чувств, в составе которого
обнаружены сенсиллы (элементарные чувствительные образования членистоногих),
реагирующие на запах, градиенты температуры, влажности и на некоторые другие
стимулы (Леонович, 2005).
В строении органа Галлера есть черты, определяющие его функциональные особен-
ности. И одновременно есть морфологические признаки, никак не связанные с функ-
циональными особенностями. Примеры - пористый волосок передней группы сенсилл
(обонятельный рецептор) может располагаться по-разному среди остальных сенсилл
передней группы, но при этом он остается сенсиллой, расположенной в переднем
углублении, и вместе с таким же волоском (сенсиллой), расположенным на симме-
тричной передней лапке, способен определять направление источника обонятельного
стимула. Учитывая мельчайшие размеры органа Галлера, расположение этой сенсиллы
относительно остальных сенсилл никак не влияет на ее функцию и поведенческую
роль. Далее, основное значение отверстия капсулы - обеспечить доступ обонятельных
(летучих) молекул к обонятельным сенсиллам капсулы. При этом форма отверстия,
т. е. вытянуто оно вдоль или поперек продольной оси членика, округлое оно или
овальное, или изрезанное, - никакого функционального значения не имеет. Подобные
морфологические признаки, на которые эволюционный пресс не оказывает давления,
могут быть использованы в качестве таксономических признаков.
Следует отметить, что ни в одной из таксономических работ данные по строению
органа Галлера никак не использовались (и не используются до сих пор). Вместе
с тем игнорировать данные по такому важному органу вряд ли правомочно.
Анализ современных работ, которые посвящены таксономии иксодовых клещей и
в которых в том числе использованы молекулярные данные, показывает, что на осно-
вании этих данных делаются весьма скоропалительные выводы, никак не соответству-
ющие морфологическому анализу. В связи с этим автору представляется важным рас-
смотреть строение органа Галлера, основного сенсорного органа иксодовых клещей, не
в связи с его функциональными особенностями, а в связи с таксономией этой группы.
Данная статья представляет собой продолжение серии статей автора, посвященной
строению органа Галлера в связи с систематикой Ixodidae.
В первой статье серии (Леонович, 2020) автор рассмотрел представителей одной
из основных таксономических групп семейства Ixodidae, в настоящей работе рас-
268
смотрены клещи семейства Amblyomminae в трактовке и терминологии Филипповой
(1997). В западной литературе все иксодовые клещи, не входящие в состав рода Ixodes
(подсемейство Ixodinae) обозначаются как группа семейств Metastriata. Согласно со-
временным ревизиям, род Amblyomma является полифилетическим и распадается
на ряд родов (Dobson, Barker, 1999; Burger et al., 2012), поэтому ставить такой род
в название семейства нецелесообразно. Вместе с тем группа Metastriata разделяется на
ряд подсемейств, которые Н.А. Филипповой трактуются как трибы в составе общего
подсемейства. В настоящей работе автор следует таксономическим представлениям
Филипповой (1997).
МАТЕРИАЛ И МЕТОДИКА
Всего в данной работе было проанализировано строение органа Галлера у 40 видов кле-
щей. Видовые названия клещей приведены по последней работе, в которой указаны валидные
названия видов (The Argasidae, Ixodidae and Nuttalliellidae (Acari: Ixodida) of the world: a list
of valid species names) (Guglielmone et al., 2010). Подродовые названия даны в скобках после
названия вида согласно работе Филипповой (1997). Далее в скобках дается ссылка на литера-
турный источник, из которого взяты данные. Рисунки выполнены с фотографий в растровом
электронном микроскопе. Звездочкой (*) помечены виды, изученные автором совместно с Бала-
шовым (Балашов, Леонович, 1981). Виды, изученные кроме автора и другими специалистами,
также обозначены звездочкой.
Методы подготовки препаратов для исследования в растровом электронном микроскопе
и особенности исследований изложены в предыдущей работе, посвященной органу Галлера
подсемейства Ixodinae (Леонович, 2020). Многие рисунки выполнены заново по фотографиям,
изготовленным автором с помощью растрового электронного микроскопа.
Список изученных видов, использованных в настоящей работе:
Род Haemaphysalis Koch, 1844
H. concinna Koch, 1844 (Haemaphysalis s. str.) *
H. inermis Birula, 1895 (Allocera)*
H. longicornis Neumann, 1901 (Kaizeriana) *(Leonovich, Belozerov, 2004)
H. punctata Canestrini et Fanzago, 1878 (Aboimisalis) *
H. sulcata Canestrini et Fanzago, 1878 (Herpetobia) *
H. erinacei Pavesi, 1884 (= Haemaphysalis numidiana Neumann, 1905) *
Род Dermacentor Koch, 1844
D. andersoni Stiles, 1908 (Wooley, 1972) *(Olenevia)
D. marginatus Sulzer, 1776* (Serdjukovia)
D. nuttali Olenev, 1928* (Serdjukovia)
D. niveus Neumann, 1897 *(= D. daghestanicus) (Леонович, 2009) (Serdjukovia)
D. silvarum Olenev, 1931*(Serdjukovia)
D. reticulatus Fabricius, 1794 (= D. pictus) *(Dermacentor s. str.) (Buzcek et al., 2002)
Род Rhipicephalus Koch, 1844
Rh. sanguineus Latreille, 1806*(Rhipicephalus s. str.)
Rh. evertsi Neumann, 1897 (Digeneus) (Cross et al., 1975; Arthur, 1975a, 1975b)*
Rh. pulchellus Gerstäcker, 1873*(Lamellicauda)
269
Rh. pumilio Schulze, 1935*(Rhipicephalus s. str.)
Rh. rossicus Yakimov & Kol-Yakimova, 1911*(Rhipicephalus s. str.)
Rh. shulzei Olenev, 1929*(Rhipicephalus s. str.)
Rh. simus Koch, 1844 (Arthur, 1975a) (Rhipicephalus s. str.)
Rh. (Boophilus) decoloratus (Koch, 1844)* (Arthur, Londt, 1973)
Rh. (Boophilus) microplus (Canestrini, 1887) (Waladde, 1977)
Rh. turanicus Pomerantsev 1936*(Rhipicephalus s. str.)
Род Hyalomma Koch, 1844
H. aegyptum Linnaeus, 1758*
H. anatolicum (Koch, 1844) *(Euhyalomma)
H. asiaticum Schulze & Schlottke, 1929 (Euhyalomma) (Леонович, 1978 )
H. brevipunctata Sharif, 1928*(Hyalommina)
H. marginatum Koch, 1844*(Euhyalomma) (Buzcek et al., 1998)
H. scupense Schulze, 1919 [= H. detritum Schulze (Apanaskevich et al., 2010)] *(Euhyalomma)
Род Amblyomma Koch, 1844
A. marmoreum Koch, 1844 (Axtell et al., 1973)
A. cajennense (Fabricius) (Famadas et al., 1997) (type species of the genus)
A. hebraeum Koch, 1844* (Rechav et al., 1977)
A. nuttali Dönitz, 1909*
A. testudinarium Koch, 1844 (Chow, Wang, 1975)
A. americanum Linnaeus, 1758 (Bruce, 1971; Foelix, Axtell, 1972)
A. variegatum Fabricius, 1794*
A. komodoense Oudemans, 1928* (Aponomma)
A (Aponomma) elaphense Price, 1959 (Aponomma) [= Robertsicus elaphense (Barker, Burger, 2018)]
(Keirans, Degenhardt, 1985).
A. trachysauri Neumann, 1899*(Aponomma)
Род Anomalohimalaya Filippova, 1994
An. lotozkyi Filippova et Panova, 1978 (Филиппова, 1992)
Род Margaropus Pomerantzev, 1947
M. winthemi Karsch, 1879 (Belozerov et al., 2002)
Результаты исследования проиллюстрированы снимками, полученными в растровом элек-
тронном микроскопе, а также схематическими изображениями органа Галлера, исполненными
в одинаковом ракурсе (рис. 1-10, см. ниже). В рисунках намеренно опущены изображения
дистальных сенсилл (в отличие от представителей подсемейства Ixodinae, у которых строение
данного отдела органа Галлера может варьировать (Леонович, 2020), клещи группы Metastriata
всегда характеризуются строго однотипным набором дистальных сенсилл, включающим один
пористый волосок обонятельной сенсиллы и один волосок вкусовой сенсиллы (рис. 1).
Результаты и обсуждение
У всех исследованных видов клещей Metastriata орган Галлера состоит из несколь-
ких морфологически обособленных частей: (1) капсулы, углубления в кутикуле перед-
него тарзуса, в которой размещены обонятельные сенсиллы. У всех представителей
группы капсула закрыта крышкой, сообщающейся со средой посредством сложного
270
капсулярного отверстия (рис. 1); (2) передней группы сенсилл, расположенной не
на расстоянии одна от другой в обширном углублении (anterior trough) (как это от-
мечено для представителей Ixodinae), а собранных компактно в особой ямке (anterior
pit), зачастую ограниченной общим сочленовным валиком: (3) дистальных сенсилл
(в отличие от Ixodinae, их не 4, а всегда 2), никогда не расположенных на особом
бугорке, но всегда на ровной поверхности тарзуса, и (3) посткапсулярных сенсилл
(рис. 1A, 1B). Половых различий в строении органа Галлера не было обнаружено,
поэтому в дальнейшем изложении рассматриваются взрослые клещи (самцы и самки
вместе), нимфы и личинки.
Передняя группа сенсилл всегда включает набор из 4 типов сенсилл (пористый,
тонкий, конический и двуслойный волоски сенсилл соответствующих типов) (Лео-
нович, 2005) (рис. 1А). Пористый волосок принадлежит обонятельной сенсилле, бо-
роздчатые волоски содержат хемо-терморецепторы, тонкий волосок - гигрорецептор,
функция конического волоска до конца не выяснена (рис. 1А). Количество сенсилл
разных типов может варьировать, но их набор (из 4 типов) остается неизменным, как
обычно неизменна их ориентация по отношению друг к другу (рис. 1С) (Леонович,
2005).
Капсула - отдел органа Галлера, в состав которого входят от 4 до 7 пористых во-
лосков обонятельных сенсилл. Сенсилла - элементарный орган чувств членистоногих,
включающий в себя кутикулярный отдел, представленный волоском (в таксономиче-
ской литературе волосок также обозначается как сета, щетинка, хета и т. п.). Сета
иннервируется одним или несколькими биполярными сенсорными нейронами. Ден-
дриты этих нейронов определяют модальность (функцию) сенсиллы. Так, в обонятель-
ных сенсиллах полый тонкостенный волосок (сета, хета) пронизан многочисленными
мелкими порами, а в полости волоска обнаруживаются многочисленные ветвления
рецепторных ресничек (Леонович, 2005). Более подробно о связи ультраструктуры
сенсилл клещей с их функцией можно узнать из монографии Леоновича (2005).
Одним из основных признаков, отличающих капсулу Metastriata от капсулы
Ixodinae, является присутствие плеоморфов - несенсорных выростов дна капсулы.
Предполагается, что крышка капсулы у амблиоммин образовалась за счет разрастания
верхних отделов плеоморфов (подробнее, см. Леонович, 2005). Второй главный при-
знак, отличающий орган Галлера Ixodinae от органа Галлера Amblyomminae, это на-
личие специализированных сложных желез, открывающихся либо снаружи от капсулы,
либо внутри капсулы, но всегда в ее проксимальном отделе. Подробно эти железы
исследованы у трех видов: A. americanum (Foelix, Axtell, 1972), Hyalomma asiaticum
(Леонович, 1978) и D. niveus (Леонович, 2009). Железистые клетки объединяются
в один или два общих протока, которые открываются либо проксимальнее капсулы,
так что отверстия протоков хорошо видны, либо вовнутрь капсулы.
Роль этих желез не совсем понятна, но остается очевидным, что эти железы име-
ются у всех представителей группы (в тех случаях, когда наружные отверстия про-
токов желез не видны, они обнаруживаются при исследовании на срезах (Леонович,
2005). Остается неясным, имеются ли железы у представителей рода Haemaphysalis.
271
Рисунок 1. Строение органа Галлера клещей подсемейства Amblyomminae на примере
лугового клеща Dermacentor niveus: A - внутреннее строение по данным трансмиссивной
электронной микроскопии (по: Леонович, 2005, 2009); В - общий вид в растровом
электронном микроскопе; С - схематическое изображение (локализация сенсилл в основных
отделах, форма переднего углубления и капсулярного отверстия) деталей строения,
показанных на рис. 1А и 1В.
Условные обозначения: AP - передняя ямка (anterior pit), в которой расположена передняя
группа сенсилл; DS - дистальные сенсиллы; CA - капсула (capsule); СO - капсулярное
отверстие (capsular orifice); DS - дистальные сенсиллы (distal sensilla); GC - железистые
клетки (glandular cells); Gl - отверстие капсулярной железы; GO - выводное отверстие
протока желез; PCS - посткапсулярные сенсиллы (postcapsular sensilla); PoS - сенсилла
с порами (porous sensillum) (обонятельная сенсилла); DwS - сенсилла с дополнительными
полостями (double-walled sensillum) ( комбинированная хемо-терморецепторная сенсилла);
BS - контактный хеморецептор, вкусовая сенсилла (gustatory (upper-pore) sensillum);
TS - терморецепторная сенсилла (thermo-sensitive sensillum); PM - плеоморфы;
SC - сенсорные нейроны.
Figure 1. Structure of Haller’s organ in ticks of the subfamily Amblyomminae with an example
of the meadow tick Dermacentor niveus: A - fine structure of the organ according to transmission
electron microscopy data (from Leonovich, 2009); В - general view of the organ in a scanning
electron microscope; С - scheme of the outer structure of the organ of the meadow tick (shown
in Fig. 1А and 1B).
Designations: AP - anterior pit with anterior group of sensilla; DS - distal sensilla; CA - capsule;
СO - capsular orifice; DS - distal sensilla; GC - glandular cells); GO - orifice of the common
duct of glandular cells; PCS - postcapsular sensilla; PoS - porous sensillu (olfactory sensillum);
DwS - double-walled sensillum (combined chemo-thermoreceptor); BS - contact chemoreceptor,
gustatory (upper-pore) sensillum; TS - thermo-sensitive sensillum; PM - pleomorphs;
SC - sensory neurons.
272
Для каждого вида клещей характер взаимного расположения сенсилл передней
группы остается строго постоянным. В специальных исследованиях, посвященных
регенерации органа Галлера у клеща Hyalomma asiaticum, путем суперимпозиции
(последовательного наложения изображений, полученных в одном ракурсе и при
одинаковом увеличении растрового электронного микроскопа), было показано, что
расположение сенсилл передней группы варьирует в определенных пределах, никак
не меняя характер относительного взаиморасположения сенсилл (рис. 2). При этом
вся группа сенсилл целиком располагается так, что пористый волосок занимает наи-
более дистальное положение, либо вся группа повернута так, что пористый волосок
занимает латеральное положение. Вне зависимости от ориентации всей группы сен-
силл относительное взаиморасположение пористого, двух бороздчатых, тонкого и
конического волоска остается постоянным (Леонович, 2005).
Проксимальнее капсулы располагается группа посткапсулярных волосков (сенсилл)
(рис. 1). Латеральные сенсиллы представлены вкусовыми рецепторами (так же как и
латеральные волоски по бокам сенсилл дистального бугорка), центральные сенсиллы
этой группы относятся к двухслойным сенсиллам (комбинированным хемо-, термо-,
гигрорецепторам) (рис. 1). Функция этих сенсилл остается неясной.
Посткапсулярные сенсиллы (точнее, их топография) сильно варьируют в пределах
видов, что не позволяет использовать их в качестве надежного таксономического
признака.
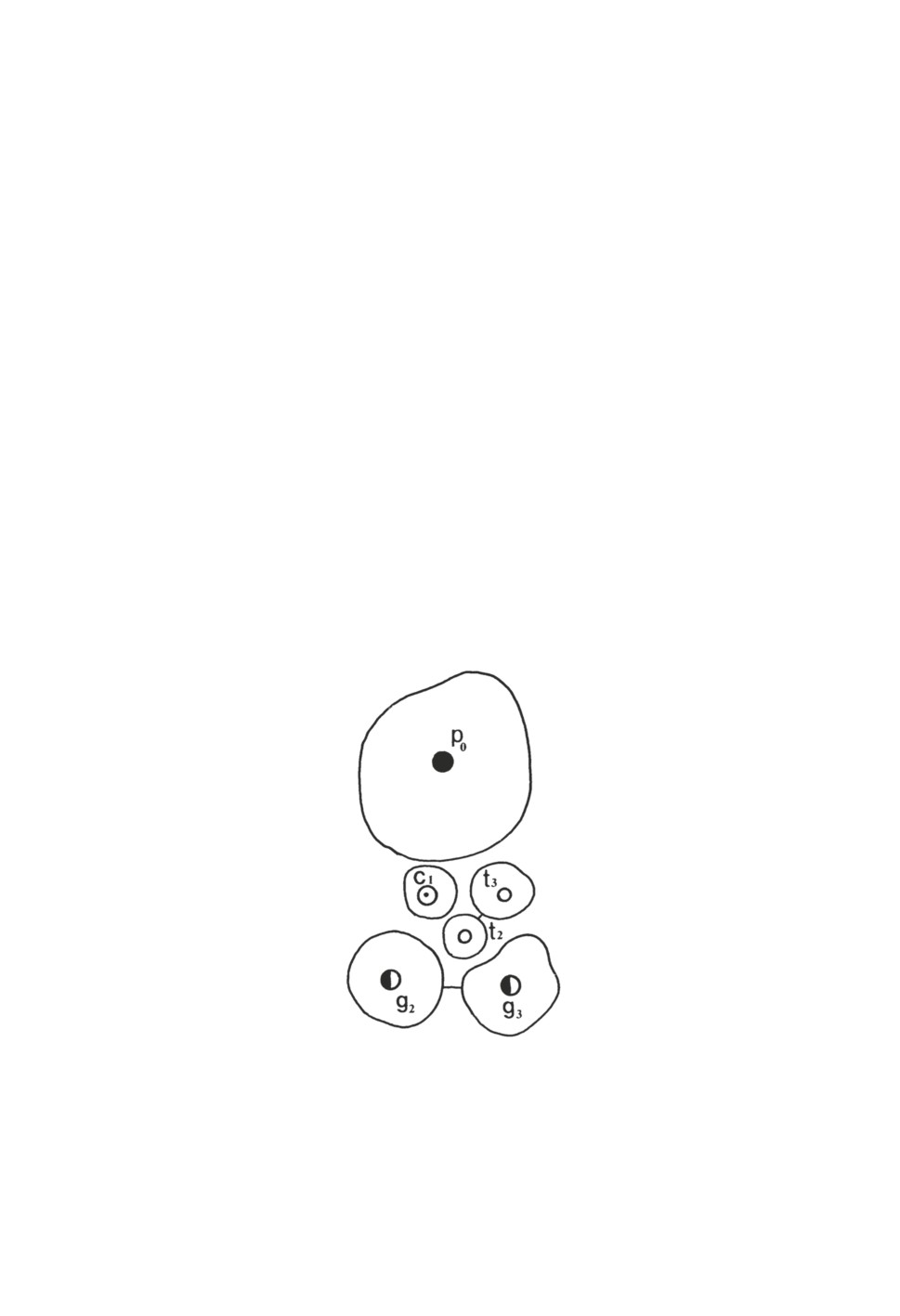
Рисунок 2. Локализация мест расположения оснований сенсилл передней группы
(anterior pit sensilla) у клеща Hyalomma asiaticum, полученная путем наложения изображений
(по: Belozerov, Leonovich, 1995): c1 - коническая сенсилла, p0 - пористая сенсилла,
t2-t3 - тонкие сенсиллы, g2-g3 - двухслойные сенсиллы
Figure 2. Topography of anterior pit sensilla in the tick Hyalomma asiaticum created by
superimposition of images obtained in scanning electron microscope (Belozerov, Leonovich, 1995).
273
Самыми надежными таксономическими признаками служат форма капсулярного
отверстия и ориентация сенсилл передней группы. Капсулярное отверстие обеспе-
чивает поступление пахучих молекул к обонятельным сенсиллам капсулы, и форма
этого отверстия (изрезанное или нет, поперечное или продольное и т. п.) не имеет
никакого функционального значения. Имеет значение общая площадь отверстия, но
она также никак не связана с формой. То же касается топографии (ориентации) сен-
силл передней группы - учитывая мелкие размеры как клещей, так и их органов
Галлера, и то, что передняя группа сенсилл сконцентрирована на очень небольшой
площади передней ямки, относительная ориентация ее сенсилл не имеет никакого
функционального значения. Таким образом, обе эти морфологические особенности -
форма капсулярного отверстия и ориентация сенсилл передней группы - могут слу-
жить хорошими таксономическими признаками.
Род Haemaphysalis Koch, 1844
Передняя группа сенсилл содержит 7 сенсилл (из них два пористых волоска)
(рис. 3A, 3B). Передняя ямка практически не выражена - сенсиллы располагаются
в относительно обширном углублении дистальнее капсулярного отверстия. Валик,
у других видов семейства совершенно отчетливый, здесь совершенно не выражен.
Передняя группа сенсилл состоит из скомпонованных на небольшом участке сенсилл
и крупного пористого волоска обонятельной сенсиллы, а также дополнительного круп-
ного пористого волоска, стоящего отдельно в латеральном положении. Капсулярное
отверстие выглядит как простая поперечная щель, иногда изогнутая (рис. 5A-5F).
У нимф и личинок размеры органа Галлера меньше, но форма капсулярного
отверстия сохраняется. В передней группе не 7, как у взрослых клещей, а только
5 сенсилл (отсутствуют один пористый и один бороздчатый волосок). Ориентация
сенсилл передней группы у личинок характеризуется смещением расположения по-
ристой сенсиллы с латерального на латеро-дистальное положение (рис. 5G, 5H).
Особенно примечательной чертой является практически полное отсутствие (за
исключением мелких несущественных деталей, или черт, находящихся в пределах
внутривидовой изменчивости) разницы в строении органа Галлера у представителей
разных подродов рода Haemaphysalis.
Род Dermacentor Koch, 1844
Передняя группа сенсилл располагается в компактной передней ямке, ограничен-
ной сочленовным валиком (рис. 1B, 3Е). В передней группе стандартный набор из
6 сенсилл, вся группа повернута так, что пористый волосок занимает латеральное
положение (рис. 3Е).
По топографии сенсилл передней группы все изученные виды сходны. У D. mar-
ginatus пористый волосок смещен более проксимально (рис. 6E), у D. andersoni общая
ячейка, в которой расположены сенсиллы, практически округлая, менее вытянутая
в сравнении с прочими изученными видами (рис. 6A-6F).
274
Форма капсулярного отверстия у всех изученных представителей рода также сход-
на: поперечная слегка изогнутая изрезанная щель (рис. 6). Наиболее характерная черта
строения капсулярного отверстия - наличие двух сочлененных лопастей в середине
дистальной части (рис. 3D, 6A-6F). У некоторых видов (D. reticulatus, D. nuttali,
D. niveus) в районе сочлененных лопастей видны наружные отверстия капсулярных
желез, напоминающие двустворчатый клапан (рис. 3F; 6A, 6С, 6D).
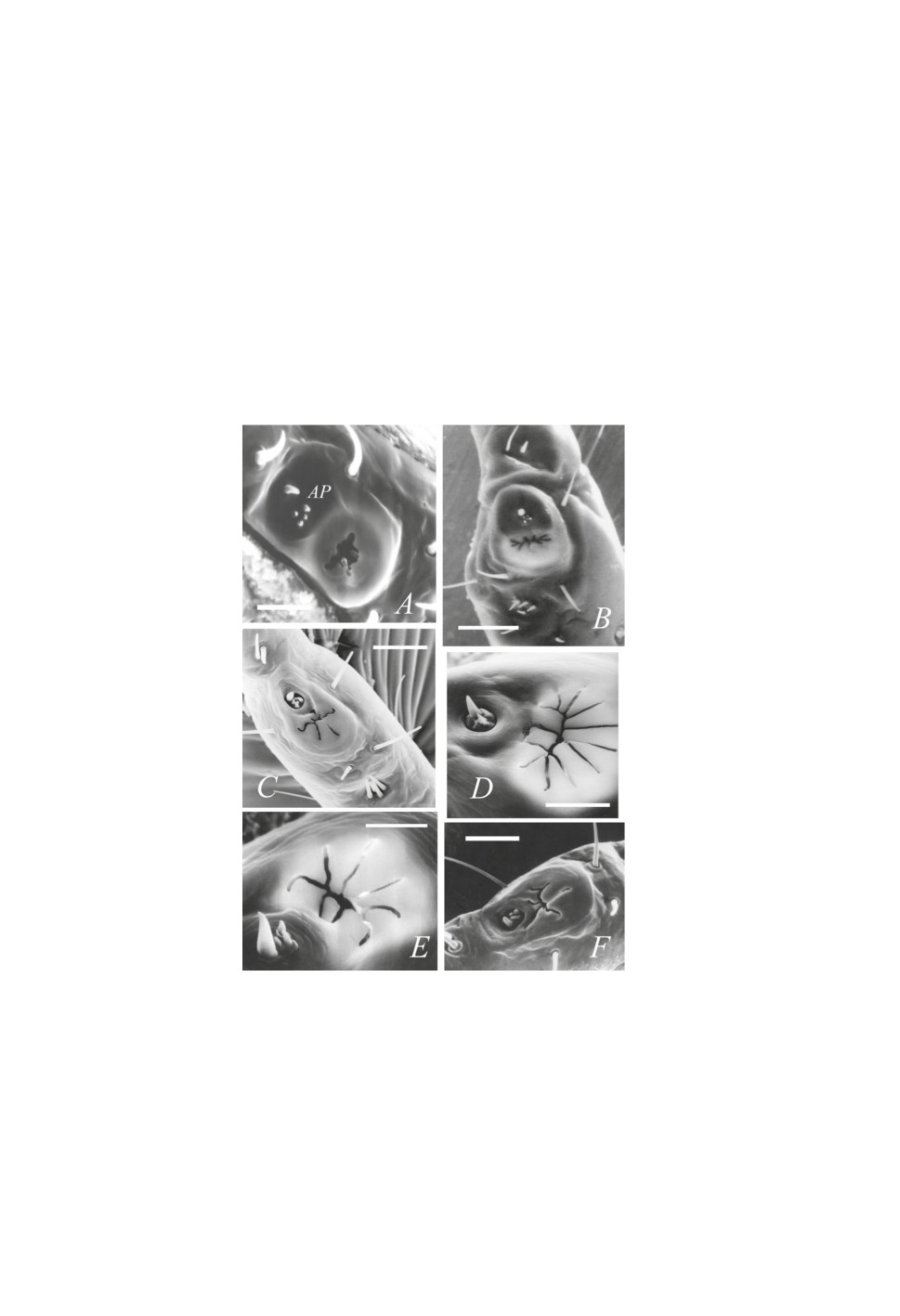
Рисунок 3. Строение органа Галлера у клещей подсемейства Amblyomminae в растровом
электронном микроскопе: A - самка Haemaphysalis longicornis (левая лапка); В - самка
H. punctata (правая лапка); С - то же, нимфа; D - то же, личинка; E - Dermacentor variabilis,
самка; F - D. niveus, капсулярное отверстие c сочлененной лопастью (JB) и отверстиями
выводных протоков капсулярных желез (GO).
Масштабная линейка, мкм: A, B, E - 30; C - 20; В - 5; D - 10.
Figure 3. Structure of Haller’s organ in ticks of the subfamily Amblyomminae in a scanning
electron microscope: A - Female of Haemaphysalis longicornis (left tarsus); В - female of H.
punctata (right tarsus); С - same species, nymph; D - same species, larva; E - Dermacentor
variabilis, female; F - D. niveus, capsular orifice with joint blade (JB) and orifices of capsular
glands.
Scale, µm: A, B, E - 30; C - 20; В - 5; D - 10.
275
Личинки представителей рода Dermacentor характеризуются наличием 5 сенсилл
передней группы (отсутствует один из бороздчатых волосков), дистальным (а не ла-
теральным) расположением пористой сенсиллы и своеобразной формой капсулярного
отверстия (рис. 4А). Это отверстие - Т-образная слегка изрезанная и довольно ши-
рокая щель (рис. 10I-10K).
Нимфы Dermacentor характеризуются достаточно широким изрезанным капсуляр-
ным отверстием, иногда крестовидным (рис. 10Е). Набор сенсилл передней группы
идентичен набору сенсилл взрослых клещей, а вот ориентация этих сенсилл иная -
пористый волосок занимает крайнее дистальное, а не латеральное положение
(рис. 10E, 10F).
Рисунок 4. Строение органа Галлера у клещей подсемейства Amblyomminae в растровом
электронном микроскопе: A - Dermacentor pictus, личинка; В - Rhipicephalus turanicus,
самка; С - тот же вид, нимфа; D - Hyalomma asiaticum, самка; E - Hyalomma anatolicum,
самка; F - H. anatolicum, нимфа.
Масштабная линейка, мкм: A, С - 5; B - 50; D, E - 30; C, F - 10.
Figure 4. Structure of Haller’s organ in ticks of the subfamily Amblyomminae in a scanning
electron microscope. A - Dermacentor pictus, larva; В - Rhipicephalus turanicus, female;
С - same species, nymph; D - Hyalomma asiaticum, female; E - Hyalomma anatolicum, female;
F - H. anatolicum, nymph.
Scale, µm: A, С - 5, B - 50; D, E - 30; C, F - 10.
276
Если проследить изменения органа Галлера в онтогенезе клещей рода Dermacentor
на примере личинки, нимфы и взрослого клеща D. silvarum (рис. 10I, 10E, и 6B) или
тех же фаз развития D. reticulatus (рис. 6A, 10E, 10J), то можно заметить следую-
щее. У личинок капсулярное отверстие вытянуто примерно одинаково в дистально-
проксимальном и латеро-латеральном направлении, у нимф отверстие становится
более вытянутым поперечно, у взрослых клещей отверстие в виде поперечной изре-
занной щели, появляются сочлененные лопасти и железы. Передняя группа сенсилл
у личинок и нимф характеризуется дистальной ориентацией пористого волоска и
отсутствием кутикулярного валика вокруг сенсилл группы.
Род Rhipicephalus Koch, 1844
Передняя группа сенсилл и капсула окружены общим кутикулярным валиком,
выдающимся над поверхностью переднего тарзуса (рис. 4В, 4С). У представителей
подродов Rhipicephalus s. str. (Rh. sanguineus, Rh. pumilio, Rh. rossicus, Rh. shulzei, Rh.
simus) и Digeneus (Rh. evertsi) (взрослые клещи) передняя группа сенсилл окружена
общим кутикулярным валиком, ориентирована пористым волоском в дистальном на-
правлении (рис. 7A-7F), Капсулярное отверстие в виде поперечной лопастной щели,
сочлененные лопасти отсутствуют. У нимф тот же набор сенсилл передней группы
и такая же как у взрослых ориентация сенсилл в этой группе, капсулярное отверстие
крестовидное (рис. 10A-10C). У личинок капсулярное отверстие крестовидное, сильно
изрезанное (рис. 7G-7L)
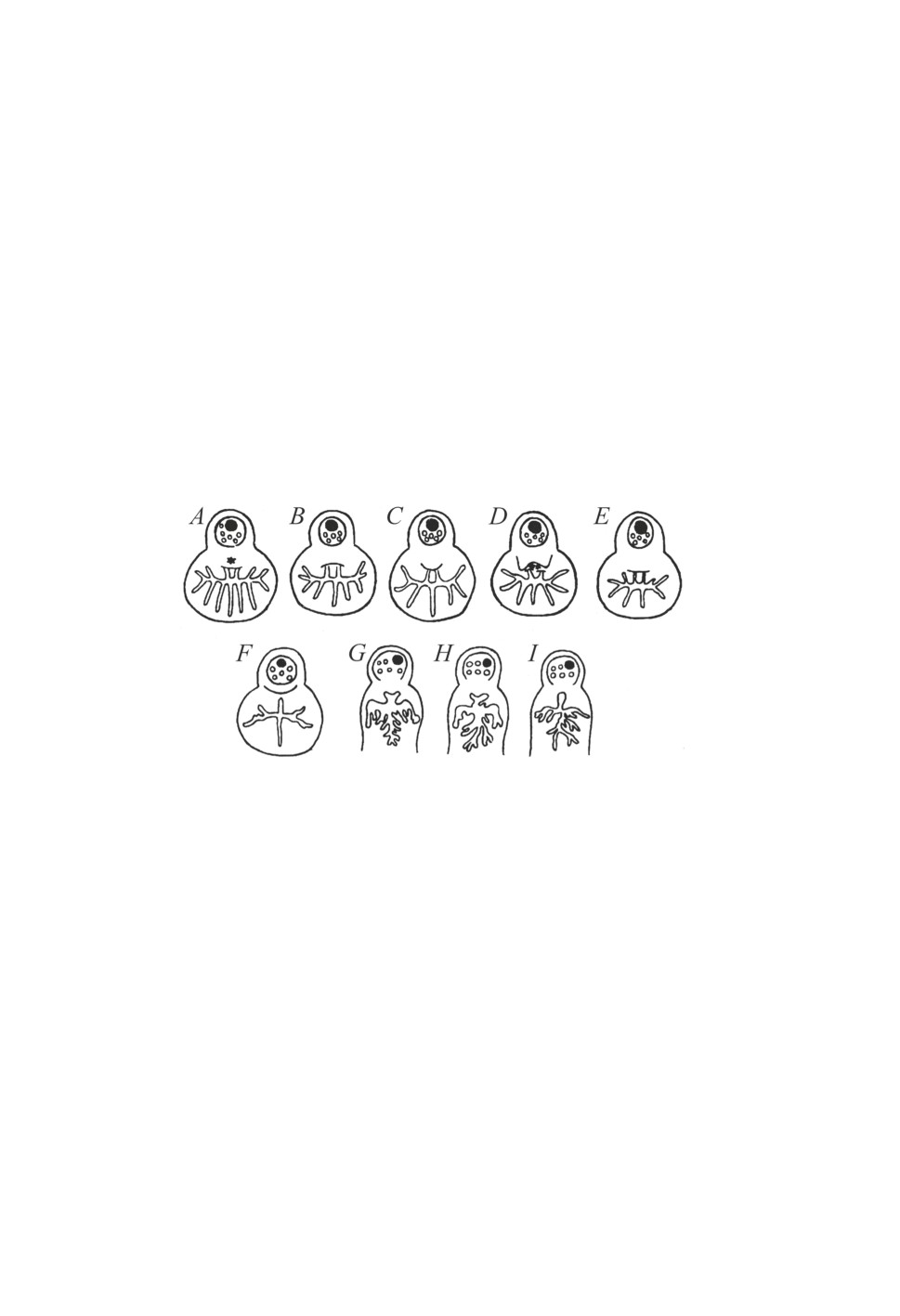
Рисунок 5. Схема строения основных отделов органа Галлера (топография сенсилл
передней группы и форма капсулярного отверстия) у представителей рода Haemaphysalis.
Взрослые клещи: A - H. erinacei, B - H. concinna, С - H. sulcata, D - H. punctata,
E - H. longicornis, F - H. inermis. Личинки: G - H. concinna, H - H. inermis.
Figure 5. Scheme of the structure of main parts of the Haller’s organ (topography of anterior pit
sensilla and shape of capsular orifice) in representatives of the genus Haemaphysalis.
Adults: A - H. erinacei, B - H. concinna, С - H. sulcata, D - H. punctata, E - H. longicornis,
F - H. inermis. Larvae: G - H. concinna, H - H. inermis.
277
Совершенно особняком стоят представители подрода Boophilus. Ранее этот род
рассматривался как самостоятельный род (Филиппова, 1997), а в последних класси-
фикациях был сведен в подрод в составе рода Rhipicephalus (Murrell, Barker, 2003;
Guglielmone et al., 2010). У двух изученных в отношении органа Галлера видов,
Rhipicephalus (Boophilus) microplus (Waladde, 1977; Балашов, Леонович, 1981) и
Rh. (B.) decoloratus (Arthur, Londt, 1973; наши данные), ориентация сенсилл перед-
ней группы резко отличается от ориентации, типичной для Rhipicephalus (рис. 7J,
7H): пористый волосок расположен не дистально, а латерально. Такое же положе-
ние передней группы характерно для нимфы B. microplus (рис. 10D). У личинки
Boophilus decoloratus (рис. 7I) ориентация передней группы дистальная (рис. 7I). Кро-
ме того, резко отличается и форма капсулярного отверстия. У представителей под-
рода Boophilus отверстие округлое, с изрезанными краями, а не лопастное (сравнить
рис. 7A-7J и рис. 7J-7H).
Род Hyalomma Koch, 1844
Сенсиллы передней группы в общей хорошо выраженной ячейке. Ориентация
передней группы дистальная, капсулярное отверстие обширное, лопастное (рис. 4D,
4E). В середине дистальной части капсулярной щели у всех видов присутствует одна
сочлененная лопасть (рис. 4D, 4E, 8A-8E). Отверстие общего выводного протока кап-
сулярных желез (Леонович, 1978) расположено в районе сочлененной лопасти, в наи-
более дистальной части отверстия видны кутикулярные выросты (шипики).
Рисунок 6. Схема строения основных отделов органа Галлера (топография сенсилл
передней группы и форма капсулярного отверстия) у представителей рода Dermacentor,
взрослые клещи: A - D. reticulatus, B - D. silvarum, С - D. nuttali, D - D. niveus,
E - D. marginatus, F - D. andersoni.
Figure 6. Scheme of the structure of main parts of the Haller’s organ (topography of anterior
pit sensilla and shape of capsular orifice) in representatives of the genus Dermacentor, adult ticks.
A - D. reticulatus, B - D. silvarum, С - D. nuttali, D - D. niveus, E - D. marginatus,
F - D. andersoni.
278
Рисунок 7. Схема строения основных отделов органа Галлера (топография сенсилл
передней группы и форма капсулярного отверстия) у представителей рода Rhipicephalus
(включая Boophilus). Взрослые клещи: A - Rh. turanicus, B - Rh. pumilio, С - Rh. shulzei,
D - Rh. rossicus, Е - Rh. sanguineus, F - Rh. evertsi, J - Rh. (Boophilus) microplus,
H - Rh. (Boophilus) decoloratus. Личинки: I - Rh. (Boophilus) decoloratus, G - Rhipicephalus
turanicus, K - Rh. shulzei, L - Rh. evertsi.
Figure 7. Scheme of the structure of main parts of the Haller’s organ (topography of anterior
pit sensilla and shape of capsular orifice) in representatives of the genus Rhipicephalus (including
Boophilus). Adult ticks: A - Rh. turanicus, B - Rh. pumilio, С - Rh. shulzei, D - Rh. rossicus,
Е - Rh. sanguineus, F - Rh. evertsi, J - Rh. (Boophilus) microplus, H - Rh. (Boophilus)
decoloratus. Larvae: I - Rh. (Boophilus) decoloratus, G - Rhipicephalus turanicus,
K - Rh. shulzei, L - Rh. evertsi.
У нимф капсулярное отверстие лопастное, но форма его близка к крестовидной
(рис. 4F, 10G, 10H). Набор и расположение сенсилл передней группы такие же, как
у взрослых клещей (рис. 8F, 10G, 10H).
У личинок капсулярное отверстие в виде широкой, сильно изрезанной Т-образной
щели (рис. 8G-8I). Сенсиллы передней группы располагаются в два ряда, причем по-
ристый волосок занимает не наиболее дистальное положение, как у нимф и взрослых
клещей, а крайнее положение в дистальном ряду (рис. 8G-8I).
Род Amblyomma Koch, 1844
Строение органа Галлера может сильно различаться у представителей рода. Капсу-
лярное отверстие в виде поперечной изогнутой щели с двумя изгибами (рис. 9A-9C).
У некоторых видов в передней группе 6 сенсилл, причем пористый волосок занимает
дистальное положение, как у Amblyomma variegatum (рис. 9A). У некоторых видов
в передней группе 7 сенсилл. В этом случае, в составе группы имеются 2 пористых
волоска: самый крупный занимает латеральное положение по отношению к основной
части группы из 6 сенсилл, включающей дистально расположенный пористый волосок
279
(A. americanum, A. nuttali, рис. 9B, 9C). У видов, ранее относимых к роду Aponomma
(автор имел возможность использовать данные по Amblyomma (Aponomma) elaphense,
A. (A.) trachysauri и A. (A.) komodoense), строение органа Галлера сильно отличается
от строения этого органа у остальных видов рода Amvblyomma. В частности, ориен-
тация передней группы сенсилл у A. (A.) komodoense и A. (A.) trachysauri уникальна
среди всех изученных Metastriata (Amblyomminae): передняя группа сенсилл ориенти-
рована таким образом, что пористый волосок занимает крайнее проксимальное (а не
дистальное) положение (рис. 9F, 9H). Капсулярное отверстие выглядит как подковоо-
бразная узкая щель (рис. 9F, 9H). У A. elaphensis ориентация передней группы сходна
с таковой A. variegatum, а капсулярное отверстие можно рассматривать как переходное
между изогнутой щелью «классических» Amblyomma и Aponomma (сравнить рис. 9Е
и рис. 9F, 9H). Весьма примечательно, что такая форма капсулярного отверстия со-
храняется и у личинок (рис. 9G), но при этом ориентация сенсилл передней группы
сходна с таковой личинок Dermacentor, Rhipicephalus и некоторых Haemaphysalis.
Рисунок 8. Схема строения основных отделов органа Галлера (топография сенсилл
передней группы и форма капсулярного отверстия) у представителей рода Hyalomma.
Взрослые клещи: A - H. asiaticum, B - H. marginatum, C - H. anatolicum, D - H. aegyptum,
E - H. brevipunctata. Нимфы: F - H. asiaticum. Личинки: G - H. asiaticum,
H - H. anatolicum, I - H. scupense (= H. detritum).
Figure 8. Scheme of the structure of main parts of the Haller’s organ (topography of anterior
pit sensilla and shape of capsular orifice) in representatives of the genus Hyalomma.
Adult ticks: A - H. asiaticum, B - H. marginatum, C - H. anatolicum, D - H. aegyptum,
E - H. brevipunctata. Nymphs: F - H. asiaticum. Larvae: G - H. asiaticum, H - H. anatolicum,
I - H. scupense (= H. detritum).
Род Margaropus Pomerantzev, 1947
У единственного исследованного в отношении органа Галлера вида M. winthemi
Karsch 1879 (Belozerov et al., 2002) передняя группа сенсилл содержит стандартный
набор из 6 сенсилл, ориентированных пористым волоском в латеральном направле-
нии (рис. 9I). Капсулярное отверстие в виде лопастной цели, в дистальной части
щели на дорзальной поверхности видны 4 кутикулярных выроста (шипика) (рис. 9I).
280
У личинки ориентация сенсилл передней группы сохраняется, при этом ориентация
передней группы сенсилл слегка смещена так, что пористый волосок занимает более
проксимальное положение. Капсулярное отверстие Т-образное (рис. 9J).
Рисунок 9. Схема строения основных отделов органа Галлера (топография сенсилл передней
группы и форма капсулярного отверстия) у представителей родов Amblyomma (включая
Aponomma), Margaropus и Anomalohimalaya. A - Amblyomma variegatum, B - A. americanum,
самка, С - A. nuttali, самка, D - A. nuttali, личинка, E - Amblyomma (Robertsicus) elaphensis,
F - Amblyomma (Aponomma) trachysauri, самка, G - тот же вид (личинка); H - Amblyomma
(Aponomma) komodoense, I - Margaropus winthemi, самка; J - тот же вид, личинка;
K, L - Anomalohimalaya lotozkyi, самка и личинка.
Figure 9. Scheme of the structure of main parts of the Haller’s organ (topography of anterior
pit sensilla and shape of capsular orifice) in representatives of the genera Amblyomma (including
Aponomma), Margaropus, and Anomalohimalaya. A - Amblyomma variegatum,
B - A. americanum, female, С - A. nuttali, female, D - A. nuttali, larva, E - Amblyomma
(Robertsicus) elaphensis, F - Amblyomma (Aponomma) trachysauri, female, same species, larva,
H - Amblyomma (Aponomma) komodoense, I - Margaropus winthemi, female, J - same species,
larva, K, L - Anomalohimalaya lotozkyi, female and larva.
Род Anomalohimalaya Filippova, 1994
У исследованного вида An. lotozkyi Filippova et Panova, 1978 (Филиппова, 1992)
ориентация передней группы сенсилл дистальная, капсулярное отверстие лопастное,
при этом лопасти расположены только с проксимальной стороны капсулярной щели.
Имеется одна сочлененная лопасть (рис. 9K). У личинки капсулярная щель в виде
сильно изрезанной изогнутой щели (рис. 9L).
Таким образом, по результатам анализа имеющихся в литературе данных по строе-
нию органа Галлера можно отметить, что в большинстве случаев особенности строе-
ния органа совпадают с разделением Metastriata (Amblyomminae) на роды. Причем
281
роды, по мнению большинства систематиков несомненно монофилетические, обла-
дают сходством в строении органа Галлера (например, род Haemaphysalis). Различия
между подродами рода Haemaphysalis в отношении таксономических признаков, вы-
деленных в данной работе, несущественны (см. рис. 5). У представителей всех под-
родов (Haemaphysalis s. str., Allocera, Kaizeriana, Aboimisalis и Herpetobia) основные
признаки строения передней группы сенсилл и капсулярного отверстия практически
идентичны.
Рисунок 10. Схема строения основных отделов органа Галлера у неполовозрелых фаз
развития: A - H: нимфы; I- L: личинки. A - Rhipicephalus turanicus, B - Rh. pumilio,
C - Rh. shulzei, D - Rh. (Boophilus) microplus, E - Dermacentor silvarum, F - D. reticulatus,
G - Hyalomma asiaticum, H - H. scupense (= H. detritum), I - D. silvarum, J - D. reticulatus,
K - D. marginatus, L - Rhipicephalus pulchellus.
Figure 10. Scheme of the structure of main parts of the Haller’s organ in immature stages.
A - H: hymphs; I- L: larvae. A - Rhipicephalus turanicus; B - Rh. pumilio; C - Rh. shulzei;
D - Rh. (Boophilus) microplus; E - Dermacentor silvarum; F - D. reticulatus; G - Hyalomma
asiaticum; H - H. scupense (= H. detritum); I - D. silvarum; J - D. reticulatus;
K - D. marginatus.
То же самое касается и представителя рода Dermacentor (6A-6F): практически
идентичная форма капсулярного отверстия и присутствие двух сочлененных лопастей;
передняя группа сенсилл, содержащая 6 рецепторных волосков; ориентация передней
группы с латеральным расположением пористого волоска (6A-6F). Такое строение
типично для изученных представителе подродов Olenevia, Serdjukovia и Dermacen-
tor s. str.
У представителей рода Hyalomma наблюдается та же картина. Принципиальной
разницы в строении органа у изученных представителей Euhyalomma и Hyalommina
не обнаружено (рис. 8)
282
Иная ситуация характерна для рода Rhipicephalus (рис. 7). У представителей под-
родов Rhipicephalus s. str. и Digeneus (7A-7F) орган Галлера характеризуется лопаст-
ной капсулярной щелью и передней группой сенсилл, ориентированной пористым
волоском дистально. В то же время, у представителей подрода Boophilus (рис. 7G,
7H) ориентация сенсилл передней группы принципиально иная (такая же, как
у Dermacentor) (рис. 6) - т.е. не с дистальным, а латеральным расположением пори-
стого волоска (напомним, что во всех случаях взаимное расположение сенсилл в со-
ставе передней группы остается постоянным, т.е. не пористый волосок перемещается
и принимает дистальное или латеральное положение, а вся группа «разворачивается»).
Кроме того, и форма капсулярного отверстия у изученных представителей Boophilus
принципиально иная (см. рис. 7J-7H). Таким образом, наши данные категорически
не согласуются с объединением родов Rhipicephalus и Boophilus в один род. Такое
мнение против объединения указанных родов в один род на основании исключительно
молекулярных данных высказывают и другие систематики-морфологи (в частности,
Каэйро (Caeiro, 2006), который приводит данные о существовании многочисленных
морфологических различий между представителя данных родов и считает, что эти
данные необходимо учитывать, а не слепо принимать «новую» молекулярную клас-
сификацию). Некоторые генетические данные также опровергают подродовой статус
Boophilus (Abdigoudarzi et al., 2011).
Многие систематики указывают на полифилию рода Amblyomma (Burger et al.,
2012), и наши данные подтверждают эту точку зрения (рис. 9). Даже в пределах «чи-
стых» представителей Amblyomma (Guglielmone et al., 2010) ориентация сенсилл пе-
редней группы может принципиально различаться (различия на родовом уровне, если
сравнить с другими родами Metastriata). Изученные представители подрода Aponomma
(A. elaphense, A. tachysauri, A. komodoense) отличаются совершенно особым, подко-
вообразным строением капсулярного отверстия (у A. elaphense эта черта строения
выражена слабее). A. trachysauri и A. komodoense дополнительно характеризуются
уникальной ориентацией сенсилл передней группы (развернутой так, что пористый
волосок занимает самое проксимальное положение). Таким образом, представители
подрода Aponomma (по крайней мере, три вида, изученных в отношении структуры
органа Галлера) имеют черты, типичные для самостоятельного рода. Видимо, реаль-
ная ревизия этого рода еще впереди. Следует надеяться, что в таких ревизиях будут
использоваться не только молекулярные данные, но и материалы по строению органа
Галлера - важнейшего рецепторного органа иксодовых клещей.
БЛАГОДАРНОСТИ
Работа выполнена по Государственному заданию «Разнообразие пара-
зитарных систем, адаптаций и путей эволюции паразитов» (номер темы:
АААА-А19-119020690109-2). В работе использованы материалы коллекции Зоологи-
ческого института РАН (ЗИН РАН) (УФК ЗИН рег. № 2-2.20).
283
СПИСОК ЛИТЕРАТУРЫ
Балашов Ю.С., Леонович С.А. 1981. Строение и пути эволюции органа Галлера иксодовых клещей под-
семейства Amblyomminae. Паразитологический сборник (ЗИН РАН, Ленинград), 5-21. [Balashov
Yu.S., Leonovich S.A. 1981. Stroenie i puti evolyutsii organa Gallera iksodovykh kleshchei podsemeistva
Amblyomminae. Parazitologicheskii Sbornik ZIN RAN, Leningrad, 5-21. (in Russian)].
Леонович C.A. 2005. Сенсорные системы паразитических клещей. Санкт-Петербург, Наука, 236 с. [Leonovich
S.A. 2005. Sensory systems of parasitic ticks and mites. St. Petersburg, Nauka, 236 pp. (in Russian)].
Леонович С.А. 1978. Тонкое строение органа Галлера иксодового клеща Hyalomma asiaticum P.Sch. et
E.Sch. (Parasitiformes, Ixodidae, Amblyomminae). Энтомологическое обозрение 57 (1): 221-226.
[Leonovich S.A. 1978. Fine structure of Haller’s organ in ixodid tick Hyalomma asiaticum P.Sch et E.Schl.
(Parasitiformes, Ixodidae, Amblyomminae). Entomologicheskoe Obozrenie 57 (1): 221-226. (in Russian)].
Леонович С.А. 2005. Сенсорные системы паразитических клещей. Спб., Наука, 232 с. [Leonovich S.A. 2005.
Sensory systems of parasitic ticks and mites, SPb., Nauka, 232 pp. (in Russian)].
Леонович С.А. 2009. Особенности строения сенсорной системы иксодового клеща Dermacentor niveus
(Parasitiformes, Ixodidae). Паразитология 43 (1): 3-11. [Leonovich S.A. 2009. Fine structural features
of the sensory system in the ixodid tick Dermacentor niveus (Parasitiformes, Ixodidae). Parazitologiya 43
(1): 3-11. (in Russian)].
Леонович С.А. 2020. Строение органа Галлера и систематика иксодовых клещей подсемейства Ixodinae
(семейство Ixodidae). Паразитология 54 (6) 470-490. [Leonovich S.A. 2020. Structure of Haller’s organ
and taxonomy of hard ticks of the subfamily Ixodinae (family Ixodidae). Parazitologiya 54 (6): 470-490).
(in Russian)]. DOI: 10.31857/S1234567806060024
Филиппова Н.А. 1992. Орган Галлера самки и личинки иксодового клеща Anomalohimalaya lotozkyi. Па-
разитология 26 (1): 78-80.[Filippova N.A. 1992.Haller’s organ in female and larva of the ixodid tick
Anomalohimalaya lotozkyi. Parazitologiya 26 (1): 78-80. (in Russian)].
Филиппова Н.А. 1997. Иксодовые клещи подсемейства Amblyomminae. Фауна России и сопредельных
стран. Паукообразные. Том 4, вып. 5. СПб., Наука, 436 с. [Filippova N.A. Ixodid ticks of the family
Amblyomminae. Fauna of Russia and neighboring countries. Arachnoidea, Vol. 4, issue 5, St. Petersburg,
Nauka, 436 pp. (in Russian)].
Abdigoudarzi M., Noureddine R., Seitzer U., Ahmed J. 2011. rDNA-ITS2 Identification of Hyalomma,
Rhipicephalus, Dermacentor and Boophilus spp. (Acari: Ixodidae) collected from different geographical
regions of Iran. Advanced Studies in Biology 3 (5): 221-238.
Apanaskevich D.A., Filippova N.A., Horak I.G. 2010. The genus Hyalomma Koch, 1844. X. Redescription of all
parasitic stages of H. (Euhyalomma) scupense Schulze, 1919 (= H. detritum Schulze) (Acari: Ixodidae)
and notes on its biology. Folia Parasitologica 57 (1): 69-78.
Arthur D.R. 1975a. The larvae of some ixodid ticks (Acarina) from the eastern Cape Province of South Africa.
Bulletin of Entomological Research 65: 405-421.
Arthur D.R. 1975b. The nymphs of some ixodid ticks (Acarina) from the eastern Cape Province of South Africa.
Bulletin of Entomological Research 65: 423-431.
Arthur D.R., Londt J.G.H. 1973. The parasitic cycle of Boophilus decoloratus (Kock 1844). Journal of
Entomological Society of South Africa 36 (1): 87-116.
Axtell R.C., Foelix R.F., Coons L.B., Roshdy M.A. 1973. Sensory receptors in ticks and mites. Proceedings of
3rd International Acarological Congress, Prague, 35-40.
Barker S.C., Burger T.D. 2018. Two new genera of hard ticks, Robertsicus n. gen. and Archaeocroton n. gen., and
the solution to the mystery of Hoogstraal’s and Kaufman’s “primitive” tick from the Carpathian Mountains.
org:pub:9B710DFE-6869-479B-A33F-79045E9A006A
Belozerov V.N., Kok D.J., Fourie L.J., Voltzit O.V. 2002. Haller’s organ and ontogenetic changes in its external
structure during life cycle of the tick Margaropus winthemi Karsch (Acari: Ixodidae). Entomological
Review 82 (7): 809- 815.
284
Belozerov V.N., Leonovich S.A. 1995. Pathways of regeneration of Haller's sensory organ in ixodid tick Hyalomma
asiaticum during life cycle. Journal of Experimental Zoology 271: 194 - 204.
Bruce W.A. 1971. Posterior capsule of Haller's organ in the lone star tick, Amblyomma americanum (Acari:
Ixodidae). Florida Entomologist 54: 65-72.
Buczek L., Buczek A., Kusmierz K., Olszewski K., Jasik K. 2002. Ultrastructural investigations of Haller's
organ in Dermacentor reticulatus (Fabr.) (Acari: Ixodida: Ixodidae), In: Acarid phylogeny and evolution.
Adaptations in mites and ticks, 227-231.
Buczek A., Jasik K., Buczek L. 1998. Sense organs in post-embryonic stages of Hyalomma marginatum
marginatum Koch, 1844 (Acari: Ixodidae). 1. Tarsal sensory system. Parassitologia (Rome) 40 (3): 279-282.
Burger T.D., Shao R., Beati L., Hilary Miller H., Barker S.C. 2012. Phylogenetic analysis of ticks (Acari: Ixodida)
using mitochondrial genomes and nuclear rRNA genes indicates that the genus Amblyomma is polyphyletic.
Caeiro V. 2006. Reflexão sobre a taxonomia actual dos Ixodidae. A sistemática morfológica versus
sistemática molecular - o género Rhipicephalus eo género Boophilus. Revista Portuguesa de Cencias
Veterinarias 101 (557-558): 37-39.
Chow Y.S., Wang C.H. 1975. External structures of Haller's organ of the Ixodes tick. Bulletin of the Institute of
Zoology, Academia Sinica (Taipei), 14: 9-17.
Cross R.H.M., Von Maltzahn H.C., Whitehead G.B. 1975. A scanning electron microscope investigation of the
structure of Haller's organ of the ixodid tick, Rhipicephalus evertsi Neumann 1897. Proceedings of Electron
Microscopy Society of South Africa 5: 77 - 78.
Dobson S.J., Barker S.C. 1999. Phylogeny of the hard ticks (Ixodidae) inferred from 18S rRNA indicates that
the genus Aponomma is paraphyletic. Molecular Phylogenetics and Evolution 11 (2): 288-295. https://doi.
org/10.1006/mpev.1998.0565
Famadas K.M., Serra-Freire N.M., Lanfredi R.M. 1997. Redescription of the larva of Amblyomma cajennense
(Fabricius) (Acari: Ixodidae) using optical and scanning electron microscopy. Acarologia 38 (2): 101-109.
Foelix R.F., Axtell R.C. 1972. Ultrastructure of Haller's organ in the tick Amblyomma americanum L. Zeitschrift
für Zellforschung und mikroskopische Anatomie 124: 275-292.
Guglielmone A.A., Robbins R.G., Apanaskevich D.A., Petney T.N., Estrada-Pena A., Horak, I.G., Shao R., Barker
S.C. 2010. The Argasidae, Ixodidae and Nuttalliellidae (Acari: Ixodida) of the world: a list of valid species
Haller G. 1881. Vorlaufige Bemerkungen Uber das Gehurorgan der Ixodiden. Zoologische Anzeiger 4: 165-167.
Keirans J.E., Degenhardt W.G. 1985. Aponomma elaphense Price, 1959 (Acari: Ixodidae) diagnosis of the adults
and nymph with first description of the larva. Proceedings of the Biological Society of Washington 98
(3): 711-717.
Leonovich S.A., Belozerov V.N. 2004. Regeneration of Haller's sensory organ in two species of hard ticks of the
genus Haemaphysalis (Acari: Ixodidae). Experimental and Applied Acarology 33: 131-144.
Leonovich S.A., Dimov I. 2012. Sense organs on palps and fore tarsi of gamasid mites (Parasitiformes,
Rhinonyssidae), parasites of the nasal cavity of the Great Tit, the Rock Dove, and the Eurasian Coot.
Entomological Review 92: 1012-1019.
Murrell A., Barker S.C. 2003. Synonymy of Boophilus Curtice, 1891 with Rhipicephalus Koch, 1844 (Acari:
Ixodidae). Systematic Parasitology 56: 169-172.
Rechav Y.H., Terry S., Knight M.M., Cross R.M.M. 1977. Chemoreceptor organs used in detection of pheromone(s)
of the tick Amblyomma hebraeum (Acarina, Ixodidae). Journal of Medical Entomology 14: 395-400.
Waladde S.M. 1977. The sensory nervous system of the adult cattle tick Boophilus microplus. II. Scanning electron
microscopy. Journal of Australian Entomological Society 16: 73-79.
Wooley T.A. 1972. Some sense organs of ticks as seen by scanning electron microscopy. Transactions of American
Microscopical Society 91 (1): 35-74.
285
STRUCTURE OF HALLER’S ORGAN AND TAXONOMY
OF HARD TICKS (FAMILY IXODIDAE)
OF THE SUBFAMILY AMBLYOMMINAE
S. A. Leonovich
Keywords: Haller’s organ, scanning electron microscopy, Ixodidae, Metastriata,
Amblyomminae, taxonomy
SUMMARY
The structure of the Haller’s organ, the main sensory organ of hard ticks, studied
by means of scanning electron microscopy, was analyzed in 40 species of the subfamily
Amblyomminae (Metastriata), representatives of the genera Amblyomma Koch, 1844
(10 species, including 3 species earlier separated as the genus Aponomma), Anomalohimalaya
Filippova, 1994 (1 species), Dermacentor Koch, 1844 (6 species), Haemaphysalis Koch,
1844 (6 species), Hyalomma Koch, 1844 (6 species), Margaropus Pomerantzev, 1947
(1 species), and Rhipicephalus Koch, 1844 (10 species, including 2 species earlier
distinguished as a separate genus Boophilus). The main taxonomic characters (number
and topography of anterior pit sensilla and shape of capsular orifice) were distinguished.
It was demonstrated that the structure of the organ is specific for separate genera, whereas
subgeneric and species differences within the genus are virtually insignificant. On the basis
of these results, some modern revisions of the subfamily based exclusively on genetic and
molecular data seem doubtful. The obtained data are compared with existing views on
taxonomy of metastriate ticks.
286