ПАРАЗИТОЛОГИЯ, 2021, том 55, № 5, с. 362-386.
УДК 576.895.775:574.3/579.842.23
МЕЖПОПУЛЯЦИОННЫЕ РАЗЛИЧИЯ БЛОХ (SIPHONAPTERA)
В ТРАНСМИССИИ ЧУМНОГО МИКРОБА
© 2021 г. Л. П. Базановаa,*, Д. Б. Вержуцкийa,**
aИркутский научно-исследовательский противочумный институт
Роспотребнадзора, Иркутск, 664047 Россия
*e-mail: bazanovalp@mail.ru
**e-mail: verzh58@rambler.ru
Поступила в редакцию 11.07.2021 г.
После доработки 27.07.2021 г.
Принята к публикации 30.07.2021 г.
Рассмотрены данные по взаимодействию возбудителя чумы и блох из различных природных
популяций. На основе широкомасштабных экспериментальных работ, проведенных преиму-
щественно в сибирских природных очагах чумы, показано значительное влияние популяцион-
ного фактора на формирование «блока» преджелудка, интенсивность образования микробных
«глыбок» в желудочном тракте насекомых, эффективность передачи возбудителя интактным
животным. Показано, что экологическая пластичность чумного микроба обусловлена не только
тесными историческими контактами с конкретными популяциями блох, хотя данный фактор
в значительной степени влияет на особенности взаимодействия патогена с этими насекомыми.
Ключевые слова: блохи, популяционная разнородность, Yersinia pestis
DOI: 10.31857/S0031184721050021
Наиболее подвижным и независимым компонентом природных очагов чумы яв-
ляются носители инфекции. Переносчики и сам возбудитель в своем использова-
нии пространства и локализации на конкретных выделах местности в значительной
степени зависят от носителей. Границы популяций переносчиков и возбудителя, как
правило, совпадают с границами популяций носителей или занимают только их часть
(Вержуцкий, 1999). Таким образом, природный очаг чумы располагается в пределах
границ популяции основного носителя инфекции в данном очаге. Территорию отдель-
ной популяции возбудителя методически правильно рассматривать как территорию
отдельного природного очага данной инфекции (Коренберг, 2010). Тем не менее су-
ществующая в настоящее время в Российской Федерации и во многих других стра-
нах мира система обозначений соподчиненных единиц природной очаговости чумы,
как правило, привязывает понятие «очаг» к территории, на которой расположены
362
от нескольких до нескольких десятков популяций основного носителя (Слудский и
др., 2019).
Так, например, в Закавказском равнинно-предгорном очаге выделяется 6 мест-
ных популяций краснохвостой песчанки: Джейранчельская, Кировабад-Казахская,
Ширванская, Кобыстанская, Апшеронская и Самур-Девичинская (Эйгелис, 1980). На
территории Приараксинского природного очага расположены 2 местные популяции
основного носителя - песчанки Виноградова (Эйгелис, 1980). В пределах Гиссарского
природного очага чумы выделяют 7 популяций основного носителя - арчовой по-
левки, каждая из которых занимает площадь от 30 до 300 км2 (Слудский и др., 2003).
Для Горно-Алтайского природного очага чумы приводят Уландрыкскую (530 км2),
Тархатинскую (1170 км2), Курайскую (390 км2) и Талдуаирскую (110 км2) популяции
монгольской пищухи (Балахонов и др., 2014).
В Тувинском природном очаге чумы показано (Балахонов и др., 2019) наличие
11 популяций длиннохвостого суслика, в границах территории каждой из кото-
рых зарегистрирована циркуляция возбудителя чумы: Моген-Буренская (645.8 км2),
Аспайтинская (143.4 км2 в пределах РФ), Кара-Бельдырская (454.4 км2 в пределах
РФ), Каргинская (1305.1 км2), Толайлыгская (493.0 км2), Барлыкская (321.6 км2),
Верхне-Барлыкская (150.1 км2 в пределах РФ), Саглинская (654.5 км2), Боро-Шайская
(642.6 км2 в пределах РФ), Чозинская (306.6 км2 в пределах РФ), Деспенская (489.8 км2).
Специальные исследования, проведенные на территории Тувинского природного очага
чумы, свидетельствуют, что выделяемые группировки являются именно образования-
ми популяционного ранга, т. е. функционально независимыми, структурированными
на неравнозначные устойчивые части и эволюционно самостоятельными единицами
(Попов, Вержуцкий, 1988; Вержуцкий и др., 1990; Вержуцкий, 2006).
Чтобы не ломать существующую схему деления энзоотичной по чуме территории
на природные очаги, мы предлагаем закрепить за эпизоотическими участками, рас-
положенными в пределах границ популяций основных носителей, термин «мезоочаг»
и в дальнейшем использовать его только в этом значении. Таким образом, фактически,
под «мезоочагом» какой-либо инфекции в природе подразумевается именно «очаг»
в классическом его понимании (Кучерук, 1972; Коренберг, 2010).
Каждая популяция основного носителя чумы (или популяции основных носителей
для полигостальных очагов), как и основного переносчика (или популяции основных
переносчиков для поливекторных очагов) специфичны по многим параметрам, вклю-
чая биохимические и физиологические показатели животных из таких группировок.
Показано, что в Туве соседние популяции длиннохвостого суслика достоверно отлича-
ются по структуре населения блох друг от друга, что имеет важное диагностическое
значение (Вержуцкий, 2012). Особенности жизненного цикла блохи Rhadinopsylla li
в соседних популяциях суслика настолько велики, что сказываются даже на числе
генераций этого вида блох (Вержуцкий, 2005). Соответственно, подобные различия
определяют и значительные вариации условий среды обитания для чумного микро-
363
ба на территории отдельных популяций носителей и переносчиков, что оказывает
влияние на течение и параметры эпизоотического процесса (Сержанов и др., 1979;
Князева и др., 1989; Вержуцкий, 1999; Базанова и др., 2006; Базанова, Вержуцкий,
2009; Сулейменов, 2009). Наиболее тесные связи возбудитель чумы имеет с блохами -
основными переносчиками в каждом из природных очагов этой инфекции (Бибикова,
Классовский, 1974; Апарин, Голубинский, 1989). Эволюционно отшлифованные взаи-
моотношения блох - основных переносчиков с чумным патогеном, циркулирующим
в каждом отдельном очаге, проявляют высокую стабильность и создают значительные
адаптационные барьеры при возможных заносах патогена в чуждую для него среду
обитания.
Проблема взаимоотношений блох и чумного микроба была и остается одной из
центральных в эпизоотологии чумы. Тем не менее ряд вопросов, касающихся попу-
ляционного уровня их взаимоотношений, до сих пор недостаточно изучен (Базанова,
2009; Базанова и др., 2010, 2013). Сибирские природные очаги чумы являются се-
верной окраиной Центрально-Азиатской зоны природной очаговости этой инфекции,
простирающейся в Монголии и Китае. В отдельных из них роль основного перенос-
чика играет один вид блох, например Citellophilus tesquorum в Забайкалье (Вершинин,
1993), Туве (Воронова, 1978; Вержуцкий и др., 2009), Монголии (Брюханова и др.,
1999; Цэренноров, 1999; Bolormaa et al., 2010; Адъясурэн и др., 2014) и Китае (The
Atlas …, 2000; Медведев и др., 2019). Территориальная близость очагов Монголии,
Забайкалья и Тувы не исключает возможность заноса микроба теми или иными пу-
тями с одной территории на другую.
МАТЕРИАЛ И МЕТОДИКА
Проведена серия экспериментальных исследований взаимоотношений возбудителя чумы и
блох, происходящих из различных мезоочагов одного природного очага, а также из географи-
чески удаленных очагов и территорий.
Особенности взаимоотношений изучали по активности формирования микробом агре-
гированных форм или биопленки (глыбок и блоков) в желудочно-кишечном тракте блох и
трансмиссии возбудителя зараженными эктопаразитами. В экспериментальных группах по-
сле каждого кормления учитывали долю пивших и погибших блох, а также число особей
со сформировавшимися конгломератами чумного микроба: глыбками, полными и частичны-
ми блоками преджелудка. Частоту блокообразования оценивали по отношению количества
блох с блоком к числу насекомых при первой подкормке, а также в среднем за подкормку.
Алиментарную активность и смертность инфицированных эктопаразитов, а также частоту
формирования бактериальных глыбок учитывали в среднем за подкормку, поскольку инфи-
цированных особей, в отличие от блокированных блох, не изымали из общей группы. Ста-
тистическую обработку данных проводили с помощью стандартных методов вариационной
статистики (Рокицкий, 1967; Никитин, Сосунова, 2003). Различия между двумя группами по
средним показателям оценивались с применением критерия Стьюдента, по изменчивости -
с применением F-критерия.
364
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Межпопуляционные различия блох в трансмиссии чумного микроба
в Тувинском природном очаге
Среди сибирских очагов чумы наиболее полно изучена пространственная структура
Тувинского очага, в пределах которого выделено 11 автономных участков очаговости
(мезоочагов): Моген-Буренский, Аспайтинский, Кара-Бельдырский, Каргинский, То-
лайлыгский, Барлыкский, Верхне-Барлыкский, Саглинский, Боро-Шайский, Чозинский
и Деспенский (рис. 1). Каждый из них функционально связан с самостоятельными
популяциями основного носителя - длиннохвостого суслика и основного перенос-
чика - блохи С. tesquorum altaicus (Вержуцкий и др., 2016; Балахонов и др., 2019).
В территориальном отношении в Тувинском природном очаге чумы границы попу-
ляций данного насекомого и его прокормителя совпадают (Вержуцкий и др., 2009).
В очаге экспериментально исследованы взаимоотношения возбудителя чумы и
блох длиннохвостого суслика, происходящих из разных мезоочагов. Поставлено две
серии опытов. Первая проведена на базе Монгун-Тайгинского эпидотряда Тувинской
противочумной станции (пос. Мугур-Аксы); вторая - в лаборатории эксперименталь-
ных животных Иркутского противочумного института. В опыты взято два вида блох
суслика (Citellophilus tesquorum altaicus и Rhadinopsylla li transbaikalica) и три штам-
ма чумного микроба (И-3226, И-3327 и И-3428). Всего использовано 1027 блох, для
подкормок инфицированных насекомых использовано 38 длиннохвостых сусликов
и 40 белых мышей. Все опыты проведены в летний период.
Рисунок 1. Тувинский природный очаг чумы и расположение отдельных мезоочагов.
Figure 1. Tuva natural plague focus and location of separate mesofoci.
365
В Тувинском природном очаге чумы C. tesquorum altaicus и Rh. li transbaikalica -
массовые виды блох длиннохвостого суслика: первый вид в большей степени при-
вязан к хозяину, второй - к его убежищу (гнезду). По численности Rh. li transbaika-
lica занимает в Юго-Западной Туве второе место после основного переносчика чумы
(Вержуцкий, 2005). Блохи этого подвида, инфицированные чумой в естественных
условиях, найдены в Тувинском и Горно-Алтайском природных очагах чумы, а также
в Западной Монголии, в Сайлюгемском природном очаге (Медведев и др., 2020). Для
обоих видов установлены блокообразование и способность к передаче возбудителя
чумы длиннохвостому суслику с генерализацией инфекции и бактериемией (Воронова,
1978; Базанова, Вержуцкий, 2001).
В первой серии для опытов взяты блохи (C. tesquorum altaicus и R. li transbaika-
lica) непосредственно из природных популяций Монгун-Тайгинского и Барлыкского
мезоочагов. В качестве прокормителя использовали длиннохвостого суслика, а для
заражения зверьков и эктопаразитов - штамм чумного микроба И-3226, выделенный
на Боро-Шайском участке очаговости (рис. 1). По своим питательным потребностям и
некоторым другим показателям возбудитель с этого участка близок варианту микроба
с Барлыкской территории и отличается от Монгун-Тайгинского (Логачев, 1999). Значи-
тельная дифференциация клонов возбудителя чумы, циркулирующих на этих участках
прослежена по 25-локусному VNTR-анализу (Тувинский …, 2019). Инфицировали
блох на сусликах, агонирующих от экспериментальной чумы, в период интенсивной
бактериемии (30-40 микробных тел в поле зрения микроскопа). Периодические под-
кормки проводили через двое суток на третьи. В данном опыте при анализе резуль-
татов пол блох не учитывали.
Во второй серии опытов в исследование взяты имаго C. tesquorum altaicus ин-
сектарной культуры, начало которой положили насекомые из Каргинской популяции.
Инфицировали блох штаммами Y. pestis И-3327 и И-3428, изолированными в Саглин-
ском и Монгун-Тайгинском мезоочагах соответственно (рис. 1).
Оба штамма типичны для очага, но различаются по питательным потребностям
в аминокислотах. На Саглинском участке очаговости циркулирует возбудитель, нуж-
дающийся в метионине, а на Каргинском - возбудитель, зависящий от метионина и
цистеина (Логачев, 1999). Блох заражали искусственно на биомембране. Заражающую
смесь готовили из равных частей суспензии, содержащей 2 млрд микробных клеток
в 1 мл физиологического раствора двухсуточной агаровой культуры, выращенной при
28 ºС, и дефибринированной крови морской свинки. Подкормки насекомых проводили
на беспородных белых мышах с той же периодичностью, что и в первой серии опытов
(через двое суток на третьи).
В опытах с блохами из природных популяций проведено по 10 подкормок каж-
дой группы C. tesquorum altaicus и по 7 и 8 подкормок в группах R. li transbaikalica.
Результаты опыта отражены в табл. 1.
366
Таблица 1. Различия во взаимоотношениях возбудителя чумы штамма И-3226
и блох из разных популяций
Table 1. Differences in the relationship between the plague pathogen strain I-3226
and fleas from different populations
Средние показатели за подкормку, %
Вид блох
Популяция
Частота
Частота
Алиментарная
Смертность
блоко-
формирования
активность
образования
глыбок
Citellophilus
Барлыкская
93.1±2.61
4.0±3.32
1.2±1.12
20.8±6.84
tesquorum altaicus
Каргинская
93.8±1.38
3.7±3.13
0
5.9±2.41
Rhadinopsylla li
Барлыкская
66.2±5.76
8.3±2.95
1.3±1.25
35.0±11.07
transbaikalica
Каргинская
72.1±4,49
5.7±1.95
0.1±1.12
19.0±6.48
Достоверных различий между популяциями у C. tesquorum altaicus и R. li trans-
baikalica в алиментарной активности, смертности инфицированных блох и частоте
образования у них блока преджелудка не выявлено.
Вместе с тем установлено влияние фактора «популяция» на частоту формирования
в организме блох C. tesquorum altaicus глыбок микроба (F=9.02, p<0,01), которая была
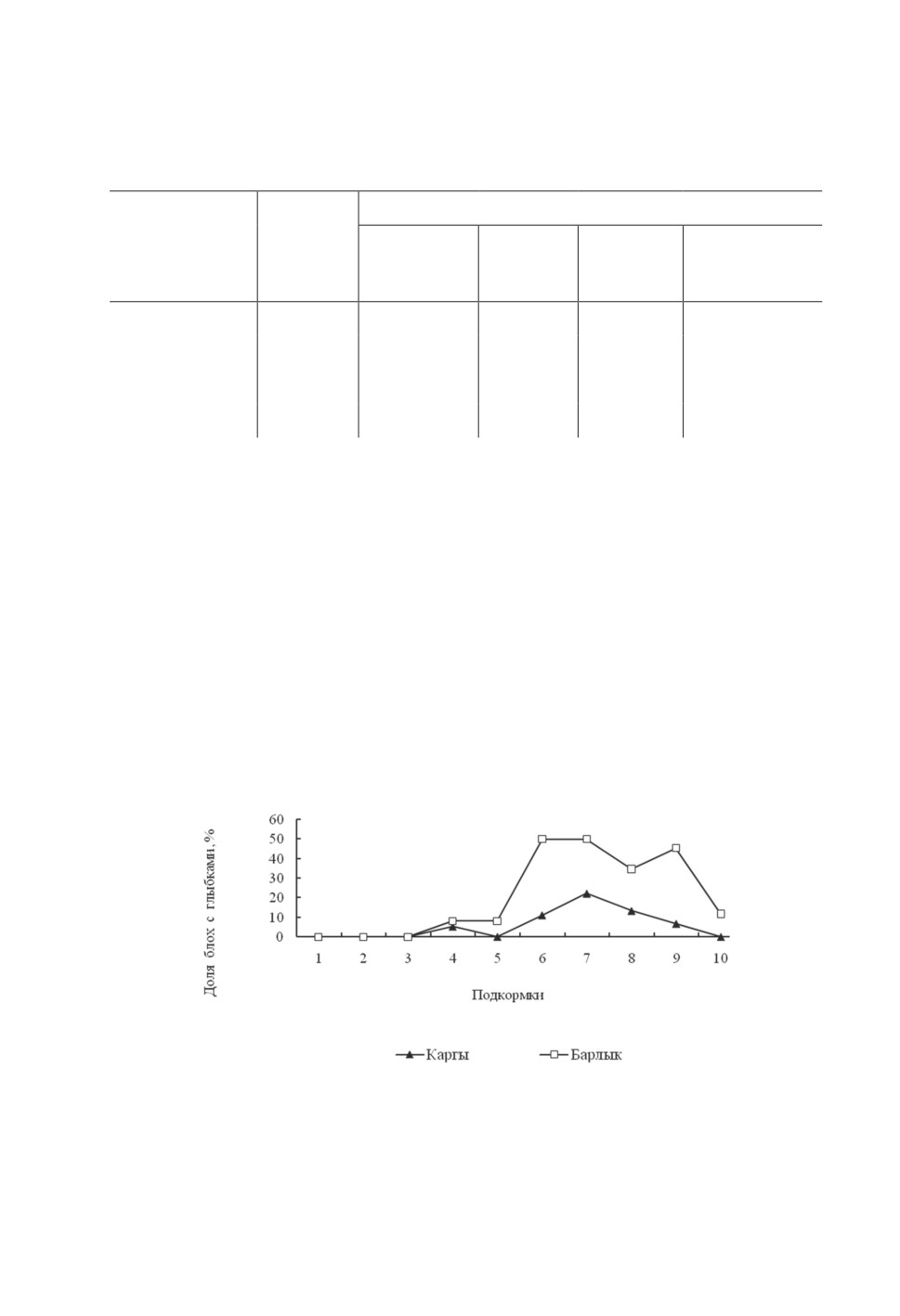
выше у насекомых Барлыкской популяции (рис. 2).
У R. li так же, как и у C. tesquorum altaicus, фактор «популяция» оказывал до-
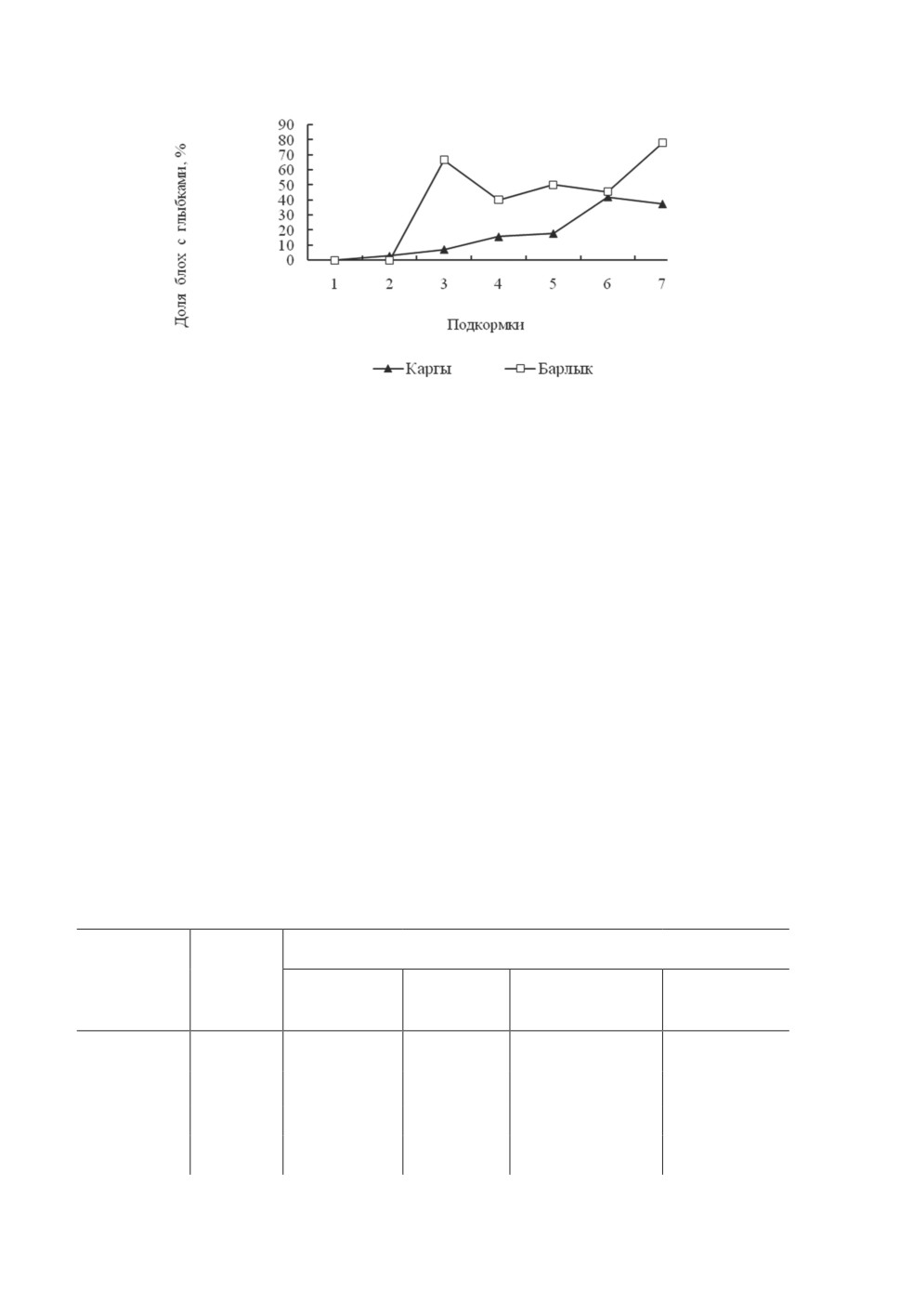
стоверное влияние на частоту формирования бактериальных глыбок (F=6.99, p<0.05).
Этот показатель был выше у блох Барлыкской популяции (рис. 3).
Рисунок 2. Частота формирования агрегированных форм чумного микроба
у блох Citellophilus tesquorum altaicus из разных популяций.
Figure 2. Frequency of formation of aggregated forms of the plague microbe
in the flea’s Citellophilus tesquorum altaicus from different populations.
367
Рисунок 3. Частота формирования агрегированных форм чумного микроба
у блох Rhadinopsylla li transbaikalica из разных популяций.
Figure 3. Frequency of formation of aggregated forms of the plague microbe
in the flea’s Rhadinopsylla li transbaikalica from different populations.
По результатам опыта более интенсивное формирование бактериальных глы-
бок исследуемым штаммом чумного микроба происходило в блохах обоих видов
с Барлыкского участка очаговости (мезоочага). Эти блохи исторически контактирова-
ли с клонами возбудителя, близкими по своим эколого-генетическим особенностям
к вариантам микроба из Боро-Шайского мезоочага (Базанова, Вержуцкий, 2019).
Во второй серии опытов проведено по 18 подкормок C. tesquorum altaicus, ин-
фицированных штаммом чумного микроба И-3428 из Каргинского мезоочага, и по
19 подкормок - штаммом И-3327 из Саглинского (табл. 2).
Таблица 2. Различия взаимоотношений инсектарной культуры блох
Citellophilus tesquorum altaicus, происходящих из Каргинского мезоочага,
и возбудителя чумы, выделенного в различных мезоочагах
Table 2. Differences in the relationship between the insect culture of the flea’s
Citellophilus tesquorum altaicus, originating from the Karginsky mesofocus,
and the plague pathogen isolated in various mesofoci
Средние показатели за подкормку, %
Исследуемый
Пол
штамм
Частота
насекомых
Алиментарная
Частота
Y. pestis
Смертность
формирования
активность
блокообразования
глыбок
И-3428
Самки
80.1±2.67
2.5±0.38
4.4±0.92
27.4±4.66
(Каргы)
Самцы
90.2±1.49
4.9±0.54
1.2±0.44
26.6±4.92
И-3327
Самки
79.8±2.16
2.0±0.58
1.9±0.52
14.2±1.69
(Саглы)
Самцы
91.6±1.50
4.7±0.60
0.4±0.15
15.5±2.93
368
В данном опыте блокированные самки, зараженные возбудителем из Каргинского
мезоочага (И-3428), встречались регулярно после каждой подкормки, гораздо чаще и
в большем количестве (до 8 особей за одну подкормку), чем инфицированные микро-
бом И-3327 (Саглы). Наибольшее количество блокированных самцов (2-5 особей)
отмечали на 20-30-е сут от заражения блох, далее обнаруживали только единичных
особей.
По результатам этой серии опытов активность кровососания и смертность блох,
инфицированных разными штаммами возбудителя чумы, не различалась у особей
обоего пола. С помощью однофакторного дисперсионного анализа установлено до-
стоверное влияние фактора «популяция» на частоту формирования бактериальных
глыбок как у самок, так и у самцов. Кроме того, отмечено влияние этого фактора на
частоту образования блока преджелудка у самок. По этим показателям штамм И-3428,
происходящий с той же территории, что и исследуемая популяция блох, значительно
превосходил штамм И-3327 (табл. 3).
Блокообразование у блох является важнейшим механизмом, обеспечивающим пере-
дачу необходимой дозы чумного микроба для эффективного заражения теплокровного
животного (Бибикова, Классовский, 1974; Ващенок, 1999). На C. tesquorum altaicus
показано, что при наличии в экспериментальных группах блокированных блох чаще
происходит передача возбудителя чумы зверькам с генерализацией инфекционного
процесса, а бактериемия обеспечивает дальнейшее инфицирование интактных на-
секомых (Базанова и др., 2003, 2004).
В анализируемых опытах у заболевших зверьков отмечена как затяжная форма
инфекции с неинтенсивной бактериемией, так и генерализованная, с агональной сеп-
тицемией, что позволяет предположить вероятность инфицирования питающихся на
таких особях насекомых.
Таблица 3. Блокообразование у Citellophilus tesquorum altaicus (инсектарная культура
из Каргинской популяции) при инфицировании двумя штаммами чумного микроба
Table 3. Block formation in Citellophilus tesquorum altaicus (insect culture
from the Karginsky population) infected with two strains of the plague microbe
Самки
Самцы
Количество
Среднее
Время
Штамм
Количество
Среднее количество
блокированных
количество
проведения
чумного
блокированных
особей с блоком
блох
особей
опыта
микроба
блох в опыте
за подкормку, %
в опыте
с блоком за
абс.
%
абс.
%
подкормку, %
Март-
И-3327
7
4.1
0.7
0
0
0
май
И-3428
11
6.1
0.8
0
0
0
Май-
И-3327
26
15.4
1.9
8
3.4
0.4
июль
И-3428
56
31.3
4.6
13
5.9
0.9
369
Проанализированы результаты бактериологического метода исследования органов
зверьков, использованных для подкормок инфицированных блох (табл. 4), которые
в наибольшей степени отражают возможность трансмиссии взятых в опыты вариантов
возбудителя.
Результаты проведенных экспериментальных исследований выявили особенности
во взаимоотношениях возбудителя чумы и блох, происходящих с различных участков
очаговости. В первой серии опытов (табл. 1) с разными популяциями C. tesquorum
altaicus и R. li transbaikalica более активное формирование агрегированных форм
чумного микроба происходило в блохах обоих видов, исторически контактировавших
с подобным, по своим питательным потребностям, вариантом возбудителя. Ранее (Сер-
жанов и др., 1979) проведены эксперименты с блохами большой песчанки Xenopsylla
skrjabini из Среднеазиатского пустынного очага чумы. В цитируемой работе показано
некоторое преимущество штаммов, адаптированных к данному виду переносчиков по
приживаемости и образованию блока преджелудка. Именно такая картина наблюдается
и в наших опытах. На этой основе можно предположить существование эволюционно
закрепленной и отшлифованной специфичности исторически сложившихся взаимо-
отношений блохи и возбудителя чумы, длительное время совместно обитающих на
одной и той же территории (Базанова и др., 2006).
Таблица 4. Трансмиссия чумного микроба блохами из разных популяций
Table 4. Transmission of the plague microbe by fleas from different populations
Количество зверьков
Количество передач
Вид
Штамм
Популяция
с
блохи
Y. pestis
сусликов
мышей
всего
генерализацией
инфекции
Rhadinopsylla
Барлыкская
И-3226
7
Не исполь-
1
0
li transbaikalica
(Боро-Шай)
зовали
Citellophilus
Барлыкская
И-3226
10
Не исполь-
3
0
tesquorum altaicus
(Боро-Шай)
зовали
R. li transbaikalica
Каргинская
И-3226
8
Не исполь-
0
0
(Боро-Шай)
зовали
C. tesquorum
Каргинская
И-3226
10
Не исполь-
1
0
altaicus
(Боро-Шай)
зовали
C. tesquorum
Каргинская
И-3428
Не исполь-
36
18
16 (88.8%)*
altaicus
(Каргы)
зовали
C. tesquorum
Каргинская
И-3327
Не исполь-
38
26
10 (38.5%)
altaicus
(Саглы)
зовали
П рим еч ани е. * - Различия между штаммами в количестве передач возбудителя блохами
с генерализацией инфекции у белых мышей достоверны (t=2.95, p<0.01).
370
Это предположение подтверждается и опытами, проведенными с двумя штаммами
возбудителя чумы из различных участков очаговости и лабораторной популяцией
C. tesquorum altaicus, происходящей с той же территории, что и один из исследуемых
штаммов. В данных опытах установлено достоверное влияние фактора «популяция»
на частоту формирования как бактериальных глыбок, так и блоков преджелудка. Не-
обходимо уточнить, что в данной серии опытов был исключен естественный прокор-
митель блохи - длиннохвостый суслик (подкормки блох проводили на белых мышах),
поэтому физиологические и биохимические особенности зверька не могли оказывать
влияние на результаты опыта.
По литературным данным при исследовании природных штаммов Y. pestis не выяв-
лено четкой зависимости способности возбудителя чумы к образованию агрегирован-
ных форм от его потребности в аминокислотах. Так, в опытах Розановой с соавторами
(1982) наблюдалась значительная вариабельность частоты блокообразования как среди
пролин-зависимых, так и среди не нуждающихся в пролине штаммов. Согласно ре-
зультатам наших исследований, триптофан-зависимые штаммы из Горно-Алтайского
очага чумы, как и типичные, в одинаковой степени формировали блок преджелудка
у блох X. cheopis (Базанова и др., 2000б). В связи с этим, можно считать, что способ-
ность исследованных штаммов чумного микроба к блокообразованию не зависит или
слабо зависит от их потребностей в аминокислотах, но, вероятно, связана с какими-то
иными, генетически закрепленными, факторами.
Таким образом, установленные в проведенных экспериментах различия во взаимо-
отношениях чумного микроба и блох по частоте формирования бактериальных глыбок
и блоков, а также активности передачи возбудителя свидетельствуют об адаптации
микроорганизма к переносчику на популяционном уровне.
Взаимоотношения чумного микроба и блох
из географически разобщенных популяций
Изучена способность C. tesquorum из сибирских регионов к инфицированию, бло-
кированию возбудителем чумы, циркулирующим в Монголии, передаче его зверькам,
и оценена возможность закрепления возбудителя в местных экосистемах при заносе
на территорию Сибири.
В опыт взяты имаго двух подвидов C. tesquorum из разных географических по-
пуляций. Блохи «Тувинской» (C. tesquorum altaicus) и «Забайкальской» (C. tesquorum
sungaris) популяций произошли от имаго, добытых в соответствующих природных
очагах чумы, блохи «Ольхонской» (C. tesquorum sungaris) - от имаго с неочаговой по
чуме территории (Ольхонский район Иркутской области). Инфицировали эктопарази-
тов чумным микробом (штамм И-3230) из Хэнтейского природного очага Монголии,
изолированным в 1988 г. от группы блох C. tesquorum sungaris. По 250 насекомых
каждой популяции заразили на биомембране и проводили подкормки на белых мышах.
Исходная зараженность блох «Тувинской» и «Забайкальской» популяций составила
371
100 %, «Ольхонской» - 70 %. Проведено по 10 периодических (через двое-трое суток)
подкормок эктопаразитов. Между подкормками насекомых содержали при температуре
18-20 ºС и относительной влажности воздуха 80-90 %. Продолжительность опыта
составила 32 дня.
Проведен сравнительный анализ частоты кровососания и смертности инфициро-
ванных насекомых разных популяций, а также активности формирования в их ор-
ганизме конгломератов чумного микроба. Дисперсионный анализ не выявил суще-
ственного влияния фактора «популяция» на алиментарную активность блох (F=2.75,
P>0.05). Средняя за подкормку доля погибших была несколько ниже среди эктопа-
разитов из Тувы (табл. 5).
Таблица 5. Алиментарная активность и смертность Citellophilus tesquorum
разных популяций при заражении чумным микробом из Монголии
Table 5. Nutritional activity and mortality of Citellophilus tesquorum
in different populations, infected with a plague microbe from Mongolia
Количество
Средняя за подкормку доля блох (%)
Популяция
блох в опыте
пивших
погибших
Самки
Самцы
Самки
Самцы
Самки
Самцы
Тувинская
156
74
91.8±2.09
93.2±2.12
3.4±0.66
8.4±1.97
Забайкальская
129
95
85.8±3.13
91.6±1.87
5.6±2.13
15.0±4.67
Ольхонская
75
36
93.6±1.33
95.6±2.66
4.2±1.24
12.7±4.05
В целом за опыт погибло (от числа имаго при первой подкормке) C. tesquorum
«Тувинской» популяции 39.0 %, «Забайкальской» - 54.6 %, «Ольхонской» - 56.8 %.
Смертность блох из Тувы была достоверно ниже, чем из Забайкалья (t=3.51, p<0.001)
и Ольхона (t=3.91, P<0.001). Доля погибших самцов всех популяций была выше, чем
самок (для «Тувинской» популяции t=4.63, p<0.001; «Забайкальской» t=6.71, p<0.001;
«Ольхонской» t=2.90, p<0.01).
С помощью дисперсионного анализа установлено влияние фактора «популяция»
на частоту формирования бактериальных глыбок у блох (F=12.02, p<0.001). Доля осо-
бей с глыбками была выше среди C. tesquorum sungaris «Забайкальской» популяции
(табл. 6). У C. tesquorum altaicus «Тувинской» и «Ольхонской» популяций чумной
микроб формировал глыбки с примерно равной частотой, однако у имаго с Ольхона
не зарегистрировано блокообразования (табл. 6).
По частоте блокообразования C. tesquorum sungaris из Забайкалья превосходи-
ли особей из Тувы более чем в пять раз. Так, блок преджелудка сформировался
у 27.2 % блох «Забайкальской» популяции и 5.2 % - «Тувинской». Различия между
популяциями по данному признаку достоверны (t=6.38, p<0.001).
372
Таблица 6. Активность формирования чумным микробом из Монголии конгломератов
в организме Citellophilus tesquorum разных популяций (без учета пола насекомых)
Table 6. Activity of the formation of conglomerates by the plague microbe from Mongolia
in the organism of Citellophilus tesquorum of different populations (excluding insect sex)
Средняя за подкормку доля блох с конгломератами (%)
Популяция
частичными
полными
глыбками
Всего
блоками
блоками
Тувинская
17.2±2.92
0.6±0.31
0.6±0.24
18.4±3.03
Забайкальская
44.8±7.33
5.0±1.64
6.2±1.90
56.0±8.36
Ольхонская
14.9±2.59
0
0
14.9±2.59
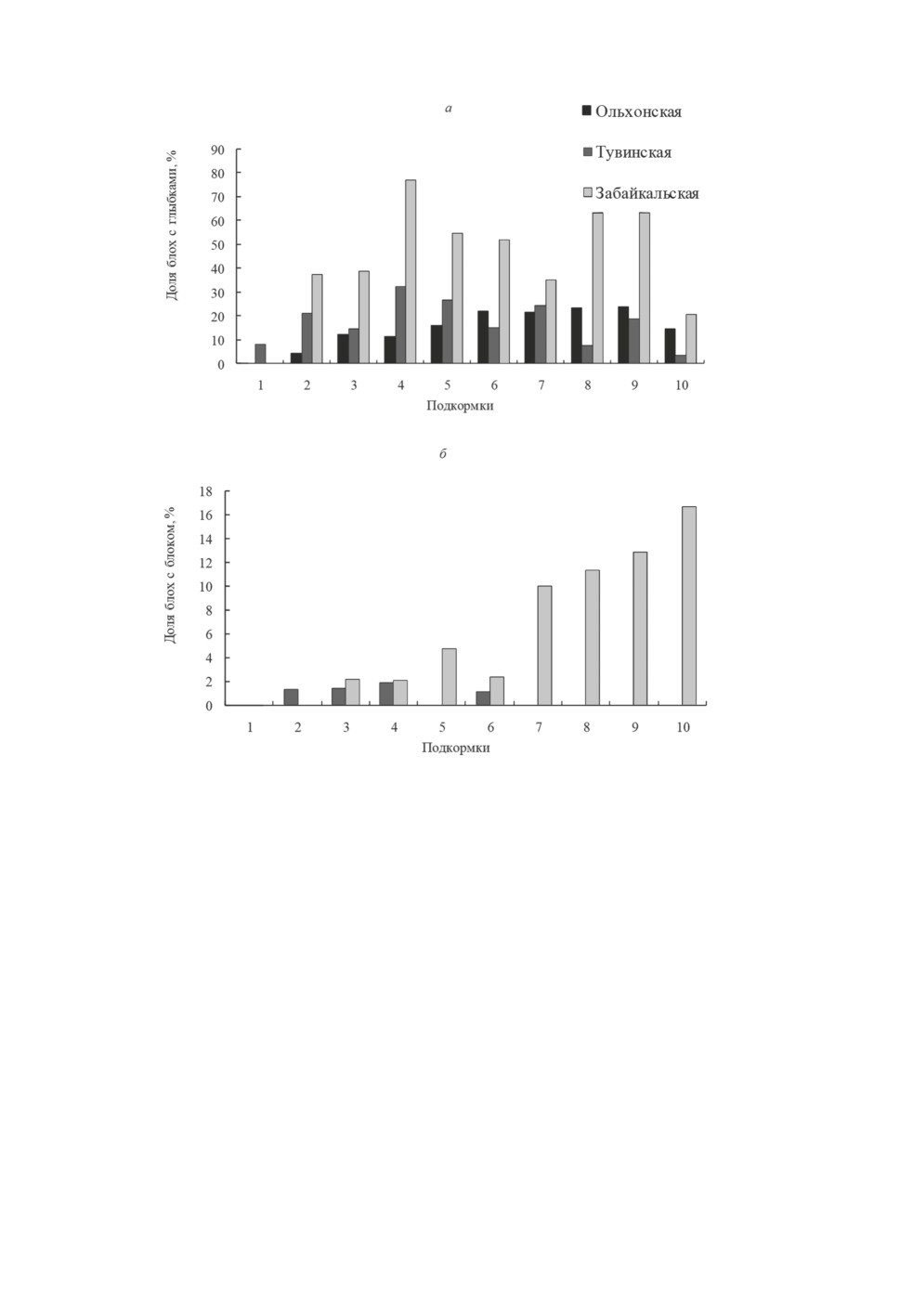
Динамика формирования бактериальных глыбок и блоков у блох представлена на
рис. 4. Начало образования глыбок отмечено у особей из Тувы после первой под-
кормки, у особей из Забайкалья и Ольхона - после второй; глыбки у насекомых вы-
являли до конца опыта. Максимум таких особей среди имаго «Забайкальской» попу-
ляции (77.0 %) зарегистрирован на 11-е, «Тувинской» (32.4 %) - на 13-е, «Ольхонской»
(23.9 %) - на 25-е сут.
Процесс блокообразования у блох «Тувинской» популяции продолжался в течение
13, «Забайкальской» - 24 сут. Формирование блоков у блох из Тувы происходило до-
статочно равномерно (1.1-1.9 % от пивших особей за одну подкормку). Доля блокиро-
ванных блох «Забайкальской» популяции после первых шести подкормок составляла
2.1-4.8 %; начиная с седьмой (20 сут) она увеличивалась с каждой подкормкой (от
10.0 до 12.9 %) и достигла максимального значения (16.7 %) в конце опыта.
Все экспериментальные группы насекомых осуществили передачу возбудителя
чумы лабораторным животным. Однако трансмиссия микроба насекомыми разных
популяций имела свои особенности. Так, блохи «Забайкальской» популяции переда-
ли возбудителя чумы семи, а блохи «Тувинской» трем зверькам из десяти. При этом
у всех павших животных, на которых кормили блох, отмечена генерализованная фор-
ма инфекции. По числу передач возбудителя (по три передачи) блохи «Тувинской» и
«Ольхонской» популяций не различались, но трансмиссия чумного микроба насекомы-
ми с Ольхона не сопровождалась гибелью мышей, а приводила к выработке антител,
титры которых составили 1:40, 1:320 и 1:160 в реакциях РПГА-РНАг.
Сравнение эколого-физиологических характеристик C. tesquorum из разных гео-
графических популяций показало, что при кровососании на белых мышах алиментар-
ная активность инфицированных насекомых не имела существенных различий. При
кормлении на биомембране менее активными были блохи «Ольхонской» популяции,
что отразилось на их исходной зараженности. Более высокая выживаемость в опыте
отмечена у C. tesquorum altaicus из Тувы, что подтверждает данные (Базанова, Маев-
ский, 1996; Токмакова и др., 2019) о способности этих блох к переживанию неблаго-
приятных условий окружающей среды, в том числе низких температур.
373
Рисунок 4. Динамика формирования чумным микробом (И-3230) из Монголии
конгломератов в организме Citellophilus tesquorum разных географических популяций:
а - бактериальных глыбок, б - блоков преджелудка.
Figure 4. Dynamics of the formation of conglomerates by the plague microbe (I-3230)
from Mongolia in the organism of Citellophilus tesquorum of different geographical populations:
a - bacterial lumps, b - blocks of the proventriculus.
Чумной микроб из Монголии формировал глыбки и блоки у C. tesquorum «За-
байкальской» популяции значительно чаще, чем у особей из других популяций.
У этих блох отмечена и самая высокая векторная способность. Необходимо отметить,
что исследуемая популяция происходит от имаго, добытых на участке Забайкальского
очага, расположенном близко к границе с Монголией. В анализируемом опыте ча-
стота блокообразования у C. tesquorum sungaris из Забайкалья достигала 27.2 %. Это
значительно превысило таковую у блох, добытых с другого участка Забайкальского
очага (3-4 %) и инфицированных типичным для данного очага штаммом (И-2621) воз-
374
будителя чумы (Феоктистов и др., 1974). У C. tesquorum sungaris, добытых на терри-
тории Монголии и зараженных штаммом Y. pestis из Прикаспийского песчаного очага,
частота формирования блока не превышала 4.5 % (Цэрэнноров, 1999). У Xenopsylla
cheopis - классического объекта экспериментальных исследований, инфицированных
чумным микробом И-3230, этот показатель равнялся 9.2 % (Воронова, Базанова, 2009),
что в три раза ниже, чем у особей C. tesquorum sungaris «Забайкальской» популя-
ции. Полученные данные позволяют предположить возможность циркуляции чумного
микроба из Монголии в случае его заноса на пограничную территорию Забайкалья
с участием C. tesquorum sungaris, которые могут являться высокоэффективными пере-
носчиками.
У C. tesquorum altaicus из Тувинского природного очага возбудитель чумы
из Монголии также формировал бактериальные глыбки и блоки преджелудка,
а инфицированные им блохи осуществляли трансмиссию микроба с генерализаци-
ей инфекционного процесса у животных. Частота блокообразования у C. tesquorum
altaicus из Тувы (5.2 %) не превышала средний показатель (5.8 %), установленный для
этих блох (Воронова, 1978). Результаты эксперимента свидетельствуют о способности
возбудителя чумы из Хэнтейского природного очага Монголии приживаться в орга-
низме основного переносчика из Тувинского природного очага, что может привести
к дополнительной активизации эпизоотий при его заносе на территорию этого очага.
Следует отметить, что изменение климатических условий, выраженное в постепенной
аридизации территории региона, привело за последние 15-20 лет к многократному
возрастанию численности C. tesquorum в Юго-Западной Туве (Вержуцкий и др., 2009).
У блох «Ольхонской» популяции глыбки формировались с не меньшей частотой,
чем у особей из Тувы, но в более поздние сроки. Однако у них не отмечено блоков
преджелудка, а у животных, на которых кормили зараженных имаго - генерализа-
ции инфекционного процесса. Так или иначе, это определяется популяционными
различиями блох из Прибайкалья, вероятно, усиленными отсутствием историческо-
го контакта насекомых этой популяции с возбудителем чумы. Имеются данные, что
у C. tesquorum с других неочаговых по чуме территорий чумной микроб формировал
блоки преджелудка, а зараженные блохи передавали его лабораторным животным
(Никитин, Базанова, 2003).
Таким образом, выявлена возможность установления функциональных контак-
тов (возбудитель-переносчик) между чумным микробом и блохами из географи-
чески разобщенных популяций. Чумной микроб, циркулирующий на территории
Хэнтейского природного очага Монголии, может формировать блоки преджелудка
у C. tesquorum из Забайкалья и Тувы, а зараженные насекомые могут осуществлять
передачу возбудителя животным с генерализацией у них инфекционного процесса, что
показывает возможность его дальнейшей трансмиссии. Внедрение чумного микроба
в экосистемы Прибайкалья достаточно проблематично, в связи с отсутствием эволю-
ционно закрепленных адаптаций при взаимодействии между патогеном и местной
популяцией блохи C. tesquorum sungaris.
375
Взаимоотношения возбудителя и переносчика чумы
из разных паразитарных систем
На северо-востоке Китая энзоотичными по чуме являются Центрально-Мань-
чжурская равнина, а также восточные и южные склоны Большого Хингана. Вся эта
огромная территория заселена даурским сусликом (Spermophilus dauricus). В земле-
дельческих районах вместе с этим сусликом в открытых стациях обитают синантроп-
ные грызуны. На диких грызунах доминирующими в сборах являются C. tesquorum
и Neopsylla bidentatiformis, на синантропных грызунах - X. cheopis (Козлов, Султанов,
2000). В Монголии и Китае почти ежегодно регистрируются вспышки заболевания
людей чумой (Enkhbaatar et al., 2003; Марамович и др., 2008). В последнее десяти-
летие в ряде очагов чумы, прилегающих к Котловине Больших озер Монголии, на-
блюдается аномально высокая активизация эпизоотий, что может привести к непред-
сказуемым эпидемиологическим последствиям (Вержуцкий, 2018). О развитии такого
неблагоприятного сценария свидетельствует резкий рост заболеваемости людей чумой
в Монголии и прилегающей части Китая в 2019-2020 гг. В этой связи представля-
ет интерес оценка взаимодействия штаммов чумного микроба из Монголии, Китая
и блох из сибирских природных очагов.
Проведены экспериментальные исследования со штаммом чумного микроба основ-
ного подвида из Китая и блохами C. tesquorum sungaris - основным переносчиком
возбудителя чумы в Забайкальском природном очаге, а также X. cheopis - класси-
ческим переносчиком в очагах чумы «крысиного» типа. Эксперименты выполнены
в весенне-летний период. В них использованы два вирулентных штамма Y. pestis:
И-3230, полученный от блох C. tesquorum из Хэнтейского природного очага чумы
Монголии, и 2155, изолированный от погибшего человека в Манчьжурии (Китай)
в 1947 г. Насекомых инфицировали с помощью биомембраны заражающей смесью
из равных частей суспензии, содержащей 2 млрд. микробных клеток в 1 мл физио-
логического раствора, выращенной на агаре при 28 ºС, и дефибринированной крови
морской свинки. Зараженных блох подкармливали через 2-3-е сут на интактных белых
мышах, между подкормками содержали при температуре 18-20 ºС и относительной
влажности воздуха 80-90%. Проведено по 10 подкормок каждой группы блох.
Результаты исследования блокообразующей способности штамма представлены
в таблице 7. У C. tesquorum sungaris, инфицированных штаммом И-3230, формирова-
ние блоков началось на 8-е сут (3 подкормка) после заражающего кормления и про-
должалось до конца опыта (по 32-е сут). Передача чумного микроба осуществлена
при групповых подкормках семи белым мышам из 10, в шести случаях среди пивших
блох выявляли от двух до 10 блокированных. В основном животные погибали на
2-4-е сут от генерализованной формы чумы.
При заражении штаммом Y. pestis 2155 у C. tesquorum sungaris процесс блокообра-
зования начался на 12-е сут (4 подкормка) и продолжался 22 дня. Зараженные блохи
осуществили передачу возбудителя при групповых подкормках трем белым мышам
376
из 10 (средняя продолжительность их жизни составила трое суток). При индивиду-
альном кормлении блокированных особей передача произошла двум животным из
10 (погибли на 3-и и 6-е сут). Сравнительный анализ блокообразующей способности
штаммов чумного микроба в организме блох этого вида показал, что формирова-
ние блоков преджелудка чаще происходило у насекомых, инфицированных штаммом
И-3230 (t=4.81, p<0.001).
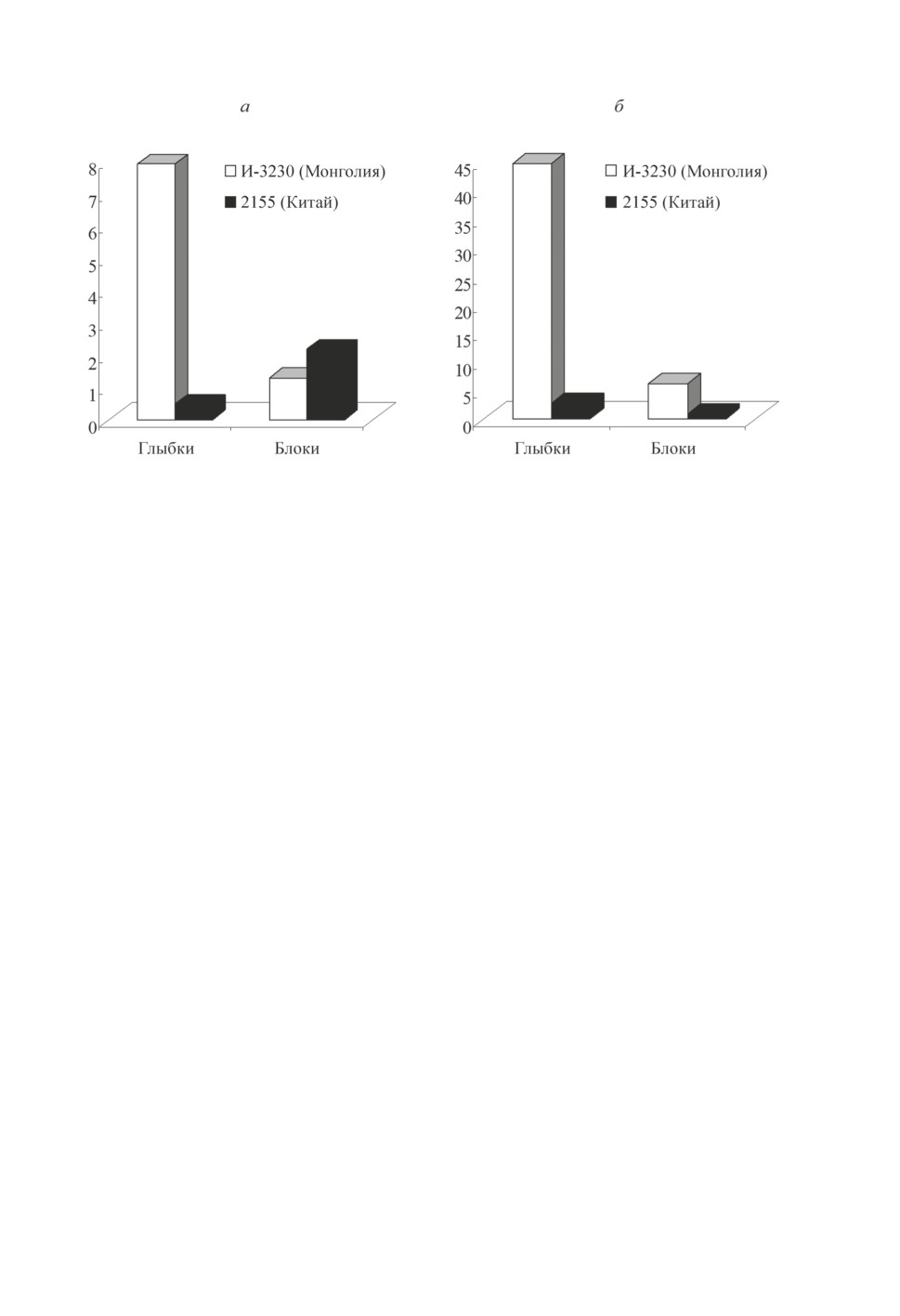
В анализируемом эксперименте штамм чумного микроба И-3230 значительно пре-
восходил штамм 2155 по способности к формированию конгломератов в виде глыбок
(рис. 5). Возможно, этот факт отражает адаптивную особенность данного штамма
чумного микроба (И-3230) к сохранению длительное время (холодный период года)
в организме блохи C. tesquorum sungaris, которая по данным Вершинина (1993) пере-
живает зимний период в основном в стадии имаго. Штамм 2155 активнее формировал
«блоки» преджелудка у X. сheopis (рис. 5), при этом сроки процесса блокообразования
у этих блох были на 3-7-е сут короче, чем у C. tesquorum sungaris, при заражении
обоими штаммами (Базанова и др., 2000а).
Представления об истории формирования очагов чумы изначально включают два
их типа: первичные природные и антропогенные, которые, в свою очередь, подраз-
деляют на синантропные крысиные и вторичные природные (Сунцов, Сунцова, 2006).
Самым распространенным и эффективным переносчиком чумного микроба в мире
является X. cheopis. У этой блохи, паразитирующей на крысах, более часто, чем у дру-
гих видов блох, чумной микроб формирует блок. Вероятно, это связано с видовыми
особенностями этого кровососущего насекомого, обеспечивающими благоприятные
условия для ускоренного размножения бактерий и образования агрегированной био-
пленки на акантах преджелудка (Jarrett et al., 2004).
Таблица 7. Образование агрегированных форм штаммами чумного микроба
из Монголии и Китая в организме блох двух видов
Table 7. Formation of aggregated forms by strains of the plague microbe
from Mongolia and China in the body of two species fleas
Средняя за подкормку доля блох
Штамм
Вид
с конгломератами микроба (%)
Y. pestis
блохи
частичными
полными
глыбками
Всего
блоками
блоками
И-3230
Citellophilus tesquorum
44.8±7.33
5.0±1.64
6.2±1.90
56.0±8.36
(Монголия)
sungaris
Xenopsylla cheopis
7.9±3.42
0.9±0.26
1.3±0.57
10.0±3.55
2155
C. tesquorum
2.7±0.77
0.1±0.11
0.9±0.35
3.7±0.91
(Китай)
sungaris
X. cheopis
0.5±0.20
1.5±0.50
2.2±0.67
4.2±1.13
377
Рисунок 5. Формирование бактериальных глыбок и блоков преджелудка
штаммами чумного микроба из Китая и Монголии в организме блох:
а - Xenopsylla cheopis, б - Citellophylus tesquorum sungaris.
Figure 5. Formation of bacterial lumps and blocks of the proventriculus by strains
of the plague microbe from China and Mongolia in the body of fleas:
a - Xenopsylla cheopis, b - Citellophylus tesquorum sungaris.
Высказывалось мнение, что наиболее вероятным районом, где чумной микроб
сурчиного варианта мог адаптироваться к организму крыс, являются склоны Гима-
лаев в пределах Юго-Западного Китая (провинция Юньнань). Там крысы рода Rattus
обитают в диких условиях и блохи X. cheopis, имея оптимальные условия существо-
вания, выступают в качестве основных переносчиков чумы и в наше время (Козлов,
Султанов, 2000). Следует отметить сомнительность данного высказывания, поскольку
горная система Гималаев не достигает провинции Юньнань, а в этих местах и сейчас,
и в обозримом прошлом отсутствуют и отсутствовали сурки. Кроме того, в есте-
ственных биотопах Юньнани массовым видом грызунов является желтобрюхая крыса
(Rattus flavipectus), не устраивающая сложных нор и не имеющая специфических
блох (Сунцов, 2020). Блоха Х. cheopis имеет африканское происхождение и появилась
в Юго-Восточной Азии с расселяющимися синантропными крысами не ранее середи-
ны XIX столетия. Более вероятным местом возможного возникновения океанической
расы чумного микроба является Северо-Западная Индия или Пакистан, где имеются
тесные контакты красных (или гималайских) сурков с индийской песчанкой (Tatera
indica) и паразитирующей на ней блохой Xenopsylla astia, являющейся эффектив-
ным переносчиком чумы. Дефицит метаболического глицерина в тканях индийской
песчанки и привел к появлению глицерин-отрицательных штаммов и их дальнейше-
му распространению среди синантропных крыс и распространившихся с ними блох
X. cheopis (Сунцов, 2020).
378
Исследуемый штамм чумного микроба из Маньчжурии изолирован от человека,
что позволяло бы предположить его возможную исходную циркуляцию в парази-
тарной системе синантропные крысы рода Rattus - X. cheopis. Но эпидемия чумы,
произошедшая в Маньчжурии в 1947 г., имела источником эпизоотию этой инфекции
среди даурских сусликов (Краминский, 1953). Т.е. использованный в опытах штамм
континентального биовара был исторически адаптирован к системе даурский суслик -
блоха C. tesquorum, что и подтверждается в нашем эксперименте с выявленным невы-
соким уровнем блокообразования у блох X. cheopis. Чумной микроб, изолированный
на территории Монголии от блохи C. tesquorum, скорее всего, циркулирует с помощью
блох этого вида в популяциях даурского суслика, демонстрируя высокую степень при-
способленности к передаче инфекции через образование блока. Тем не менее стоит
отметить, что в проведенных опытах экспериментальным путем установлена возмож-
ность достаточно высокого уровня трансмиссии данных штаммов эктопаразитами из
других паразитарных систем.
ЗАКЛЮЧЕНИЕ
Выявлено, что взаимоотношения возбудителя чумы и блох, происходящих
с различных участков очаговости, имеют свои особенности. В первом случае, при
исследовании разных популяций C. tesquorum altaicus и R. li transbaikalica, более
активное формирование агрегированных форм чумного микроба происходило в бло-
хах обоих видов, исторически контактировавших с подобным вариантом возбудите-
ля. Ранее некоторое преимущество штаммов по приживаемости в организме и об-
разованию блока преджелудка, адаптированных к данному виду переносчиков, было
показано на блохах больших песчанок X. skrjabini из Среднеазиатского пустынного
очага (Лухнова, Казакбаева, 1985). Сходная картина наблюдается и в наших опытах.
В данном случае можно предположить существование специфичности исторически
сложившихся взаимоотношений популяции блохи и возбудителя чумы, циркулирую-
щего на ее территории. Последнее подтверждается и опытами, проведенными с дву-
мя штаммами возбудителя чумы из различных участков очаговости и лабораторной
популяцией C. tesquorum altaicus, происходящей с той же территории, что и один
из исследуемых штаммов. Достоверные различия во взаимоотношениях чумного
микроба и блох по двум показателям (частоте формирования бактериальных глыбок
и блока преджелудка) свидетельствуют об адаптации микроорганизма к переносчи-
ку на популяционном уровне. У штамма И-3428, циркулирующего на Каргинском
мезоочаге, способность формировать бактериальные глыбки и блоки преджелудка
у C. tesquorum altaicus, происходящих с той же территории, была значительно выше,
чем у штамма И-3327 из Саглинского мезоочага. Необходимо отметить, что осо-
бенности взаимоотношений между переносчиком и возбудителем, адаптированным
к организму данного переносчика, не всегда можно выявить, регистрируя среди
инфицированных блох только особей с блоком преджелудка. Например, достовер-
379
ность различий между двумя природными популяциями C. tesquorum altaicus и
R. li transbaikalica во взаимоотношениях с возбудителем чумы была установлена
благодаря тому, что в экспериментальных группах насекомых учитывали не только
блокированных особей, но и всех блох со сформировавшимися агрегатами (конгло-
мератами) чумного микроба.
Выявлена возможность установления функциональных контактов (возбудитель-
переносчик) между чумным микробом и блохами из географически разобщенных
популяций. Чумной микроб, циркулирующий на территории Хэнтейского природ-
ного очага Монголии, может формировать блоки преджелудка у C. tesquorum из За-
байкалья и Тувы, а зараженные насекомые могут осуществлять передачу возбуди-
теля животным с генерализацией у них инфекционного процесса, что показывает
принципиальную возможность его укоренения и дальнейшей трансмиссии в местных
биоценозах. Укоренение и дальнейшую циркуляцию микроба в популяциях грызу-
нов и насекомых на неочаговой по чуме территории пока следует рассматривать как
маловероятные. Показано, что значительно чаще блок преджелудка формировался
у имаго «Забайкальской» популяции. Насекомые, взятые в этот опыт, выведены от
блох из сборов с участка Забайкальского очага, расположенного вблизи границы
с Монголией. Частота блокообразования в несколько раз превосходила ранее установ-
ленную величину показателя для данного вида блох. Эти насекомые характеризова-
лись и более высокой векторной способностью. У C. tesquorum altaicus «Тувинской»
популяции блокообразование происходило с такой же частотой, что и при инфициро-
вании блох типичным для Тувинского природного очага штаммом возбудителя чумы.
У C. tesquorum с неочаговой по чуме территории случаев блокообразования не заре-
гистрировано. Полученные данные предполагают возможность циркуляции чумного
микроба из Монголии в случае его заноса на территорию Забайкалья с участием
C. tesquorum sungaris.
Нельзя исключить возможность взаимодействия чумного микроба и блох из разных
паразитарных систем (из сибирских природных очагов, а также Монголии и Китая).
При этом необходимо учитывать, что адаптивная пластичность возбудителя чумы
определяется не только особенностями структурной организации сочленов парази-
тарной системы и условиями существования, но и наличием исторических контактов
местных популяций переносчика с возбудителем чумы.
СПИСОК Литературы
Апарин Г.П., Голубинский Е.П. 1989. Микробиология чумы. Иркутск, Иркутский госуниверситет, 92 с. [Apa-
rin G.P., Golubinsky E.P. 1989. Plague microbiology. Irkutsk, Irkutsk State University, 92 pp. (In Russian)]
Адъяасурэн З., Цэрэнноров Д., Мягмар Ж., Ганхуяг Ц., Отгонбаяр Д., Баяр Ц., Вержуцкий Д.Б., Ганболд Д.,
Балахонов С.В. 2014. Современная ситуация в природных очагах чумы Монголии. Дальневосточный
журнал инфекционной патологии 25: 22-25. [Adjasuren Z., Tserennorov D., Mjagmar Zh., Ganhujag
C., Otgonbayar D., Bayar C., Verzhutsky D.B., Ganbold D., Balakhonov S.V. The current situation in the
plague natural foci in Mongolia. Far Eastern Journal of Infectious Pathology 25: 22-25. (In Russian)]
380
Базанова Л.П. 2009. Взаимоотношения чумного микроба (Yersinia pestis) и блох (Siphonaptera) (на примере
сибирских природных очагов чумы): Автореф. дис. … докт. биол. наук. Улан-Удэ, 46 с. [Bazanova
L.P. 2009. The relationship between the plague microbe (Yersinia pestis) and fleas (Siphonaptera) (on
the example of Siberian natural plague foci): Abstract. dis
doct. biol. sciences. Ulan-Ude, 46 pp. (In
Russian)]
Базанова Л.П., Вержуцкий Д.Б. 2001. К оценке эпизоотологической роли блохи Rhadinopsylla li transbai-
kalica Ioff et Tifl. (1946) в Тувинском природном очаге чумы. Актуальные проблемы инфектологии
и паразитологии: Материалы международной научной конференции. Томск, 42. [Bazanova L.P., Ver-
zhutsky D.B. 2001. Evaluation of the epizootic role of the flea Rhadinopsylla li transbaikalica Ioff et Tifl.
(1946) in the Tuva natural plague focus. Actual problems of infectious diseases and parasitology: Materials
of the international scientific conference. Tomsk, 42. (In Russian)]
Базанова Л.П., Вержуцкий Д.Б. 2009. Эпизоотологическое значение блох (Siphonaptera) в Тувинском при-
родном очаге чумы (обзор). Байкальский зоологический журнал 3: 13-22. [Bazanova L.P., Verzhutsky
D.B. 2009. Epizootological significance of fleas (Siphonaptera) in the Tuva natural plague focus (review).
Baikal Zoological Journal 3: 13-22. (In Russian)]
Базанова Л.П., Вержуцкий Д.Б. 2019. Межпопуляционные различия блох в трансмиссии чумного микро-
ба в Тувинском природном очаге. Дальневосточный журнал инфекционной патологии 37: 49-50.
[Bazanova L.P., Verzhutsky D.B. 2019. Interpopulation differences of fleas in the transmission of the plague
microbe in the Tuva natural focus. Far Eastern Journal of Infectious Pathology 37: 49-50. (In Russian)]
Базанова Л.П., Вержуцкий Д.Б., Никитин А.Я., Токмакова Е.Г., Воронова Г.А., Хабаров А.В. 2006. Осо-
бенности взаимоотношений возбудителя чумы и блох с различных участков Тувинского природного
очага. Медицинская паразитология и паразитарные болезни 3: 35-38. [Bazanova L.P., Verzhutsky D.B.,
Nikitin A.Ya., Tokmakova E.G., Voronova G.A., Khabarov A.V. 2006. Peculiarities of the relationship
between the causative agent of plague and fleas from different parts of the Tuva natural focus. Medical
Parasitology and Parasitic Diseases 3: 35-38. (In Russian)]
Базанова Л.П., Вержуцкий Д.Б., Никитин А.Я., Токмакова Е.Г., Хабаров А.В. 2004. Различия между двумя
популяциями Citellophilus tesquorum altaicus из Тувинского природного очага чумы по особенно-
стям взаимоотношений с возбудителем и морфологическим признакам. Медицинская паразитология
и паразитарные болезни 1: 37-39. [Bazanova L.P., Verzhutsky D.B., Nikitin A.Ya., Tokmakova E.G.,
Khabarov A.V. 2004. Differences between two populations of Citellophilus tesquorum altaicus from the
Tuva natural plague focus in terms of the relationship with the pathogen and morphological characteristics.
Medical Parasitology and Parasitic Diseases 1: 37-39. (In Russian)]
Базанова Л.П., Вержуцкий Д.Б., Хабаров А.В. 2000а. Межпопуляционные различия во взаимоотношениях
с возбудителем чумы двух массовых видов блох длиннохвостого суслика из Тувинского природного
очага. Карантинные и зоонозные инфекции в Казахстане: Материалы Международного симпозиума.
Алматы, 2: 48-52. [Bazanova L.P., Verzhutsky D.B., Khabarov A.V. 2000. Interpopulation differences in
the relationship with the plague pathogen of two common species of long-tailed ground squirrel fleas from
the Tuva natural focus. Quarantine and Zoonotic Infections in Kazakhstan: Materials of the International
symposium. Almaty, 2: 48-52. (In Russian)]
Базанова Л.П., Воронова Г.А., Косилко С.А. 2010. Взаимоотношения возбудителя и переносчика чумы
из географически разобщенных популяций. Проблемы особо опасных инфекций 4 (106): 66-69.
[Bazanova L.P., Voronova G.A., Kosilko S.A. 2010. The relationship between the causative agent and the
carrier of plague from geographically dispersed populations. Problems of Especially Dangerous Infections
4 (106): 66-69. (In Russian)]
Базанова Л.П., Воронова Г.А., Косилко С.А. 2013. Взаимоотношения возбудителя и переносчика чумы
из разных паразитарных систем. Медицинская паразитология и паразитарные болезни 1: 12-16.
[Bazanova L.P., Voronova G.A., Kosilko S.A. 2013. The relationship between the causative agent and the
carrier of plague from different parasitic systems. Medical Parasitology and Parasitic Diseases 1: 12-16.
(In Russian)]
381
Базанова Л.П., Воронова Г.А., Токмакова Е Г. 2000б. Различия в образовании блока преджелудка у самцов
и самок Xenopsylla cheopis (Siphonaptera: Pulicidae). Паразитология 34 (1): 56-59. [Bazanova L.P.,
Voronova G.A., Tokmakova E.G. 2000. Differences in the formation of a pre-stomach block in males
and females of Xenopsylla cheopis (Siphonaptera: Pulicidae). Parasitologiya 34 (1): 56-59. (In Russian)]
Базанова Л.П., Маевский М.П. 1996. Длительность сохранения чумного микроба в организме блохи Citel-
lophilus tesquorum altaicus. Медицинская паразитология и паразитарные болезни 1: 45-48. [Bazanova
L.P., Mayevsky M.P. 1996. Duration of preservation of the plague microbe in the organism of the flea
Citellophilus tesquorum altaicus. Medical Parasitology and Parasitic Diseases 1: 45-48. (In Russian)]
Базанова Л.П., Токмакова Е.Г., Маевский М.П. 2003. Значение блокированных и неблокированных блох
Citellophilus tesquorum altaicus в передаче чумной инфекции. Проблемы особо опасных инфекций
86: 14-20. [Bazanova L.P., Tokmakova E.G., Mayevsky M.P. 2003. The role of blocked and unblocked
fleas Citellophilus tesquorum altaicus in the transmission of plague infection. Problems of Especially
Dangerous Infections 86: 14-20. (In Russian)]
Балахонов С.В., Корзун В.М., Чипанин Е.В., Афанасьев М.В., Михайлов Е.П., Денисов А.В., Фомина
Л.А., Ешелкин И.И., Машковский И.К., Мищенко А.И., Рождественский Е.Н., Ярыгина М.Б. 2014.
Горно-Алтайский природный очаг чумы. Ретроспективный анализ, Эпизоотологический мониторинг,
современное состояние Новосибирск, Наука-Центр, 272 с. [Balakhonov S.V. et al. 2014. Gorno-Altai
natural plague focus. Retrospective analysis, epizootological monitoring, current state. Novosibirsk, Science
Center, 272 pp. (In Russian)]
Балахонов С.В., Вержуцкий Д.Б., Холин А.В., Базанова Л.П., Климов В.Т., Косилко С.А., Окунев Л.П.,
Токмакова Е.Г., Шестопалов М.Ю., Галацевич Н.Ф., Акимова И.С., Салчак Л.К. 2019. Тувинский
природный очаг чумы Иркутск, Иркутский госуниверситет, 286 с. [Balakhonov S.V. et al. 2019. Tuva
natural plague focus. Irkutsk, Irkutsk State University, 286 pp. (In Russian)]
Бибикова В.А., Классовский Л.Н. 1974. Передача чумы блохами. М., Медицина, 188 с. [Bibikova V.A.,
Klassovsky L.N. 1974. Transmission of the plague by fleas. M., Medicine, 188 p. (In Russian)]
Брюханова Г.Д., Цэрэнноров Д., Щедрин В.И., Чумакова И.В., Грижебовский Г.М. 1999. Особенности
передачи чумы блохами Citellophilus tesquorum sungaris. Scient. Journal Centre Control and Research
of Nat. Infect. Diseases. Ulaanbaatar, 7: 205-206. [Bryukhanova G.D., Tserennorov D., Shchedrin V.I.,
Chumakova I.V., Grizhebovsky G.M. 1999. Peculiarities of transmission of plague by fleas Citellophilus
tesquorum sungaris. Scient. Journal Center Control and Research of Nat. Infect. Diseases. Ulaanbaatar,
7: 205-206. (In Russian)]
Ващенок В.С. 1999. Роль блох в эпизоотологии чумы. Паразитология 33 (3): 198-209. [Vaschenok V.S. 1999.
The role of fleas in plague epizootology. Parasitologiya 33 (3): 198-209. (In Russian)]
Вержуцкий Д.Б. 1999. Эпизоотологическая роль популяционной организации населения блох длиннохво-
стого суслика в Тувинском природном очаге чумы. Паразитология 33 (3): 242-249. [Verzhutsky D.B.
1999. Epizootological role of the population organization of the long-tailed ground squirrel fleas in the
Tuva natural plague focus. Parasitologiya 33 (3): 242-249. (In Russian)]
Вержуцкий Д.Б. 2005. Пространственная организация населения хозяина и его эктопаразитов: теорети-
ческие и прикладные аспекты (на примере длиннохвостого суслика и его блох). Автореф. дис. …
докт. биол. наук. Иркутск, 46 с. [Verzhutsky D.B. 2005. Spatial organization of the host population and
its ectoparasites: theoretical and applied aspects (on the example of the long-tailed ground squirrel and its
fleas). Abstract dis
doct. of biol. sciences. Irkutsk, 46 pp. (In Russian)]
Вержуцкий Д.Б. 2006. Межпопуляционные связи у длиннохвостого суслика в Юго-Западной Туве. Бюл-
летень МОИП, отд. биол. 111 (5): 52-59. [Verzhutskiy D.B. 2006. Interpopulation relationships in the
long-tailed ground squirrel in South-West Tuva. Bulletin MOIP, dep. biol. 111 (5): 52-59. (In Russian)]
Вержуцкий Д.Б. 2012. Маркерная роль эктопаразитов в популяционных исследованиях их хозяев. Байкаль-
ский зоологический журнал 10 (2): 94-102. [Verzhutsky D.B. 2012. The marker role of ectoparasites in
population studies of their hosts. Baikal Zoological Journal 10 (2): 94-102. (In Russian)]
382
Вержуцкий Д.Б. 2018. Активизация природных очагов чумы в Центральной Азии: беспочвенные опасения
или реальная угроза. Природа Внутренней Азии 6 (1): 7-17. [Verzhutsky D.B. 2018. Activation of
natural foci of plague in Central Asia: groundless fears or a real threat. The Nature of Inner Asia 6 (1):
7-17. (In Russian)]
Вержуцкий Д.Б., Зонов Г.Б., Попов В.В. 1990. Эпизоотологическое значение накопления блох в агрегациях
самок длиннохвостого суслика в Тувинском природном очаге чумы. Паразитология 24 (3): 186-192.
[Verzhutsky D.B., Zonov G.B., Popov V.V. 1990. Epizootological significance of flea accumulation in
aggregations of long-tailed ground squirrel females in the Tuvinian natural focus of plague. Parasitologiya
24 (3): 186-192. (In Russian)]
Вержуцкий Д.Б., Ткаченко С.В., Галацевич Н.Ф., Чумакова Н.А., Немкова Н.К., Акимова И.С., Холин А.В.,
Балахонов С.В. 2016. Обнаружение новых эпизоотических участков в Тувинском природном очаге
чумы. Национальные приоритеты России 22 (4): 17-21. [Verzhutsky D.B., Tkachenko S.V., Galatsevich
N.F., Chumakova N.A., Nemkova N.K., Akimova I.S., Kholin A.V., Balakhonov S.V. 2016. Discovery
of new epizootic sites in the Tuva natural plague focus. National Priorities of Russia 22 (4): 17-21. (In
Russian)]
Вержуцкий Д.Б., Чумакова Н.А., Галацевич Н.Ф., Ковалева Н.И. 2009. К экологии блохи Citellophilus tes-
quorum Wagn., 1898 в Юго-Западной Туве. Байкальский зоологический журнал 1: 17-22. [Verzhutsky
D.B., Chumakova N.A., Galatsevich N.F., Kovaleva N.I. 2009. To the ecology of the flea Citellophilus
tesquorum Wagn., 1898 in South-West Tuva. Baikal Zoological Journal 1: 17-22. (In Russian)]
Вершинин Е.А. 1993. Экологические особенности блох даурского суслика в Забайкальском природном
очаге чумы. Дис
канд. биол. наук. Иркутск, 243 с. [Vershinin E.A. 1993. Ecological features of the
fleas of the Daurian ground squirrel in the Transbaikal natural plague focus. Dis
cand. biol. sciences.
Irkutsk, 243 pp. (In Russian)]
Воронова Г.А. 1978. Ceratophyllus tesquorum altaicus Ioff 1936 как основной переносчик и хранитель чумно-
го микроба в Тувинском природном очаге. Эпидемиология и профилактика особо опасных инфекций
в МНР и СССР. Улан-Батор, Госиздат, 152-155. [Voronova G.A. 1978. Ceratophyllus tesquorum altaicus
Ioff 1936 as the main carrier and guardian of the plague microbe in the Tuva natural focus. Epidemiology
and prevention of especially dangerous infections in the Mongolian People’s Republic and the USSR.
Ulaanbaatar, State Publishing House, 152-155. (In Russian)]
Воронова Г.А., Базанова Л.П. 2009. О возможности возникновения локальных эпизоотий на территории
Сибири при заносе возбудителя чумы из Монголии. Журнал инфекционной патологии 16 (3): 88-89.
[Voronova G.A., Bazanova L.P. 2009. On the possibility of local epizootics on the territory of Siberia
when the plague pathogen is brought in from Mongolia. Journal of Infectious Pathology 16 (3): 88-89.
(In Russian)]
Князева Т.В., Топорков В.П., Бережнов А.З., Величко Л.Н., Балухин В.Н., Гордиенко Т.Г., Подсвирова В.В.
1989. Сравнение эффективности передачи чумы блохами малого суслика из разных природных попу-
ляций. Природная очаговость, микробиология и профилактика зоонозов. Саратов, 116-121. [Knyazeva
T.V., Toporkov V.P., Berezhnov A.Z., Velichko L.N., Balukhin V.N., Gordienko T.G., Podsvirova V.V. 1989.
Comparison of the efficiency of transmission of plague by small ground squirrel fleas from different natural
populations. Natural focus, microbiol. and profilact. zoonoses. Saratov, 116-121. (In Russian)]
Козлов М.П., Султанов Г.В. 2000. Чума (природная очаговость, эпизоотология). Махачкала, Мавел, 304 с.
[Kozlov M.P., Sultanov G.V. 2000. Plague (natural foci, epizootology). Makhachkala, Mavel, 304 p. (In
Russian)]
Коренберг Э.И. 2010. Природная очаговость инфекций: современные проблемы и перспективы исследо-
ваний. Зоологический журнал 89 (1): 5-17. [Korenberg E.I. 2010. Natural focus of infections: current
problems and research prospects. Zoologicheskiy Zhurnal 89 (1): 5-17. (In Russian)]
Краминский В.А. 1953. Материалы по военной эпидемиологии Китая. Дис. … канд. мед. наук. Вороши-
лов, 612 с. [Kraminsky V.A. 1953. Materials on the military epidemiology of China. Dis
cand. med.
sciences. Voroshilov, 612 p. (In Russian)]
383
Кучерук В.В. 1972. Структура, типология и районирование природных очагов болезней человека. Итоги
развития учения о природной очаговости болезней человека и дальнейшие задачи. М., Медицина,
180-212. [Kucheruk V.V. 1972. Structure, typology and regionalization of natural foci of human diseases.
The results of the development of the doctrine of natural foci of human diseases and further tasks. M.,
Medicine, 180-212. (In Russian)]
Логачев А.И. 1999. Особенности штаммов чумного микроба, изолированных на различных участках Тувин-
ского природного очага чумы. Chinese Journal of Control of Endemic Diseases 14: 210-212. [Logachev
A.I. 1999. Features of the strains of the plague microbe isolated in different parts of the Tuva natural
plague focus. Chinese Journal of Control of Endemic Diseases 14: 210-212. (In Russian)]
Лухнова Л.Ю., Казакбаева Р.А. 1985. Приживаемость и конкурентная способность штаммов возбудите-
ля чумы в организме разных переносчиков. Тезисы докл. XII Межреспубликанской науч.-практ.
конф. противочумных учреждений Средней Азии и Казахстана по профилактике чумы. Алма-Ата,
69-71. [Lukhnova L.Yu., Kazakbaeva R.A. 1985. The survival and competitive ability of strains of the
plague pathogen in the body of different vectors. Abstracts of the XII Inter-republican scientific-practical
conference of anti-plague institutions in Central Asia and Kazakhstan on the prevention of plague. Alma-
Ata, 69-71. (In Russian)]
Марамович А.С., Косилко С.А., Иннокентьева Т.И., Воронова Г.А., Базанова Л.П., Никитин А.Я., Окунев
Л.П. 2008. Чума в Китае. Опасность заноса в регионы Сибири и Дальнего Востока. Журнал микро-
биологии, эпидемиологии и иммунобиологии 1: 95-99. [Maramovich A.S., Kosilko S.A., Innokentieva
T.I., Voronova G.A., Bazanova L.P., Nikitin A.Ya., Okunev L.P. 2008. Plague in China. The danger
of drifting into the regions of Siberia and the Far East. Journal of Microbiology, Epidemiology and
Immunobiology 1: 95-99. (In Russian)]
Медведев С.Г., Вержуцкий Д.Б., Котти Б.К. 2020. Разнообразие переносчиков возбудителя чумы: по-
лигостальные паразиты - блохи рода Rhadinopsylla Jordan et Rotschild, 1911 (Siphonaptera:
Hystrichopsyllidae). Паразитология 54 (3): 205-230. [Medvedev S.G., Verzhutsky D.B., Kotti B.K. 2020.
Diversity of carriers of the plague pathogen: polygostal parasites - fleas of the genus Rhadinopsylla
Jordan et Rotschild, 1911 (Siphonaptera: Hystrichopsyllidae). Parasitologiya 54 (3): 205-230. (In Russian)].
Медведев С.Г., Котти Б.К., Вержуцкий Д.Б. 2019. Разнообразие блох (Siphonaptera) - переносчиков воз-
будителей чумы: паразит сусликов - блоха Citellophilus tesquorum (Wagner, 1898). Паразитология
53 (3): 179-197. [Medvedev S.G., Kotti B.K., Verzhutsky D.B. 2019. Diversity of fleas (Siphonaptera) -
carriers of plague pathogens: parasite of ground squirrels - flea Citellophilus tesquorum (Wagner, 1898).
Никитин А.Я., Базанова Л.П. 2003. Исследование возможности образования эффективных ксенопар между
Yersinia pestis и блохой Citellophilus tesquorum. Бюллетень восточно-сибирского научного центра
СО РАМН 3: 152-155. [Nikitin A.Ya., Bazanova L.P. 2003. Investigation of the possibility of effective
xenopair formation between Yersinia pestis and the flea Citellophilus tesquorum. Bulletin of the West
Siberian Scientific Center of the Siberian Branch of the Russian Academy of Medical Science 3: 152-155.
(In Russian)]
Никитин А.Я., Сосунова И.А. 2003. Анализ и прогноз временных рядов в экологических наблюдениях и
экспериментах (Учебное пособие). Иркутск, ИГПУ, 81 с. [Nikitin A.Ya., Sosunova I.A. 2003. Analysis
and forecast of time series in environmental observations and experiments (Textbook). Irkutsk, IGPU, 81
pp. (In Russian)]
Попов В.В., Вержуцкий Д.Б. 1988. Характеристика внутрипопуляционных группировок длиннохвостого
суслика (Citellus undulatus Pall.) в период депрессии численности. Бюллетень МОИП, отд. биол. 93
(6): 47-50. [Popov V.V., Verzhutsky D.B. 1988. Characteristics of intrapopulation groups of long-tailed
ground squirrel (Citellus undulatus Pall.) during population depression. Bulletin MOIP, dep. biol. 93 (6):
47-50. (In Russian)]
384
Розанова Г.Н., Осипова С.П., Багдасарова Т.Г. 1982. Способность штаммов чумного микроба с различным
характером питания к образованию блока в организме блох. Болезни с природ. очагов. на Кавказе:
Тез. докл. науч. конф. Ставрополь, 122-123. [Rozanova G.N., Osipova S.P., Bagdasarova T.G. 1982.
The ability of strains of the plague microbe with different feeding patterns to form a block in the body
of fleas. Disease from natural foci in the Caucasus: Abstracts report scientific. conf. Stavropol, 122-123.
(In Russian)]
Рокицкий П.Ф. 1967. Биологическая статистика. Минск, Вышэйшая школа, 328 с. [Rokitsky P.F. 1967.
Biological statistics. Minsk, Vyshyeishaja Shkola, 328 pp. (In Russian)]
Сержанов О.С., Хрусцелевская Н.М., Чумаченко В.Д., Асенов Г.А., Матаков М.И. 1979. Блокообразование
у блох Xenopsylla gerbilli caspica из различных ландшафтно-экологических участков Кызылкумов.
Проблемы особо опасных инфекций 4: 58-60. [Serzhanov O.S., Khruscelevskaya N.M., Chumachenko
V.D., Asenov G.A., Matakov M.I. 1979. Block formation in fleas Xenopsylla gerbilli caspica from various
landscape-ecological areas of the Kyzyl Kum. Problems of especially dangerous infections 4: 58-60. (In
Russian)]
Слудский А.А., Бойко А.В., Ляпин М.Н., Тарасов М.А. 2019. Популяционный подход к определению границ
и количеству природных очагов чумы. Поволжский экологический журнал 4: 493-502. [Sludsky A.A.,
Boyko A.V., Lyapin M.N., Tarasov M.A. 2019. A population-based approach to defining the boundaries
and the number of natural foci of plague. Povolzhskiy Journal of Ecology 4: 493-502. (In Russian)].
Слудский А.А., Дерлятко К.И., Головко Э.Н., Агеев В.С. 2003. Гиссарский природный очаг чумы. Саратов:
изд-во Саратовск. ун-та, 248 с. [Sludsky A.A., Derlyatko K.I., Golovko E.N., Ageev V.S. 2003. Gissar
natural plague focus. Saratov, Saratov State University, 248 pp. (In Russian)]
Сулейменов Б.М. 2009. Энзоотия и эпизоотия чумы. Алмааты, 476 с. [Suleimenov B.M. 2009. Enzooty and
epizooty of plague. Almaty, 476 pp. (In Russian)]
Сунцов В.В. 2020. Гостальный эффект территориальной экспансии микроба чумы Yersinia pestis из попу-
ляций монгольского сурка-тарбагана (Marmota sibirica). Зоологический журнал 99 (11): 1307-1320.
[Suntsov V.V. 2020. Gostal effect of the territorial expansion of the plague microbe Yersinia pestis from the
populations of the Mongolian tarbagan (Marmota sibirica). Zoologicheskiy Zhurnal 99 (11): 1307-1320.
Сунцов В.В., Сунцова Н.И. 2006. Чума. Происхождение и эволюция эпизоотической системы (экологиче-
ские, географические и социальные аспекты). М., КМК Scientific Press, 247 с. [Suntsov V.V., Suntsova
N.I. 2006. Plague. Origin and evolution of the epizootic system (ecological, geographic and social aspects).
M., KMK Scientific Press, 247 pp. (In Russian)]
Токмакова Е.Г., Базанова Л.П., Вершинин Е.А. 2019. Способность блох Citellophilus tesquorum и
Frontopsylla luculenta (Siphonaptera, Ceratophilidae) к переживанию отрицательных темпера-
тур в зимний период. Паразитология 53 (2): 120-128. [Tokmakova E.G., Bazanova L.P., Vershinin
E.A. 2019. Ability of fleas Citellophilus tesquorum and Frontopsylla luculenta (Siphonaptera,
Ceratophilidae) to survive negative temperatures in winter. Parasitologiya 53 (2): 120-128. (In Russian)].
Феоктистов А.З., Даниленко А.Ф., Юзвик Л.Н., Шведко Л.П. 1974. Эффективность массовых видов блох
Забайкалья как переносчиков чумы. Доклады Иркутского противочумного института 10: 206-208.
[Feoktistov A.Z., Danilenko A.F., Yuzvik L.N., Shvedko L.P. 1974. Efficiency of common species of fleas of
Transbaikalia as carriers of plague. Reports of the Irkutsk Anti-Plague Institute 10: 206-208. (In Russian)]
Цэрэнноров Д. 1999. Эпизоотологическое значение блох Citellophilus tesquorum sungaris (Jordan, 1929) и
Frontopsylla luculenta luculenta (Jordan et Rothschild, 1923) в природных очагах чумы Монголии:
Автореф. дис
канд. биол. наук. Ставрополь, 18 с. [Tserennorov D. 1999. Epizootological significance
of fleas Citellophilus tesquorum sungaris (Jordan, 1929) and Frontopsylla luculenta luculenta (Jordan et
Rothschild, 1923) in natural foci of plague in Mongolia: Abst. dis
cand. biol. sciences. Stavropol, 18
pp. (In Russian)]
385
Эйгелис Ю.К. 1980. Грызуны Восточного Закавказья и проблема оздоровления местных очагов чумы.
Саратов: изд-во Саратовск. ун-та, 262 с. [Eigelis Yu.K. 1980. Rodents of the Eastern Transcaucasia
and the problem of improving local plague foci. Saratov: Saratov State University, 262 pp. (In Russian)]
Bolormaa G., Undraa B., Baigalmaa M., Otgonbaatar D. 2010. Plague in Mongolia. Vector-Borne and Zoonotic
Diseases 10 (1): 69-75.
Enkhbaatar L., Jamyansbarav B., Ariuntuya O., Batsaikhan B. 2003. The Prevalence and Prognosis of Plague in
Mongolia. Scientific Journal Center for Infectious Diseases with Natural Foci (Ulaanbaatar) 11: 253-257.
Jarrett C.O., Deak E., Isherwood K.E., Oyston P.S., Fischer E.R., Whitney A.R., Kobayashi S.D., DeLeo F.R.,
Hinnebusch B.J. 2004. Transmission of Yersinia pestis from an infectious biofilm in the flea vector. Journal
The Atlas of Plague and Its Environment in the People’s Republic of China. 2000. Beijing, Science Press, 221 pp.
INTERPOPULATION DIFFERENCES OF FLEAS (SIPHONAPTERA)
IN THE TRANSMISSION OF PLAGUE PATHOGEN
L. P. Bazanova, D. B. Verzhutsky
Keywords: fleas, population diversity, Yersinia pestis
SUMMARY
The interactions of the plague pathogen with fleas from various natural populations have
been considered. A significant effect of the population factor on the proventriculus block
formation, on the intensity of the microbial clumps formation in the gastric tract of insects,
and on the efficiency of the pathogen transmission to intact animals has been demonstrated
on the basis of large-scale experimental work carried out mainly in the Siberian natural
foci of plague. The results led us to assume that the ecological plasticity of the plague
pathogen was caused not only by its close contacts with specific flea populations in the
past. However, this factor significantly affects the characteristics of the interaction between
the pathogen and these insects.
386