ПАРАЗИТОЛОГИЯ, 2021, том 55, № 6, с. 465-475.
УДК 593.1:576.8
ЭНДОБИОНТНЫЕ ИНФУЗОРИИ ИЗ РУБЦА КОСУЛИ СИБИРСКОЙ
CAPREOLUS PYGARGUS
© 2021 г. О. А. Корниловаа, Л. В. Чистяковаb,*,
И. В. Серёдкинc, И. П. Грабарникd
aРоссийский государственный педагогический университет им. А.И. Герцена,
наб. р. Мойки, д. 48, Санкт-Петербург, 191186 Россия
bЗоологический институт РАН,
Университетская наб., 1, Санкт-Петербург, 199034 Россия
cТихоокеанский институт географии ДВО РАН,
ул. Радио, 7, Владивосток, 690041 Россия
dФГАОУ ВО Первый Московский государственный медицинский университет
им. И.М. Сеченова Минздрава России (Сеченовский Университет),
ул. Трубецкая, д. 8, Москва, 119991 Россия
* e-mail: pelomixa@mail.ru
Поступила в редакцию10.10.2021 г.
После доработки20.10.2021 г.
Принята к публикации 23.10.2021 г.
Исследована фауна эндобионтных инфузорий рубца косули сибирской Сapreolus pygargus
из Тернейского района Приморского края России. У исследованных косуль обнаружен един-
ственный вид инфузорий Entodinium dubardi, при этом эндобионты были выявлены менее
чем у 40% хозяев в обследованной популяции. Мы предполагаем, что крайне низкое видовое
разнообразие эндобионтных инфузорий и невысокая степень заражения хозяев эндобионтами
связаны в первую очередь со снижением уровня стадности, а также особенностями пищевого
рациона косуль.
Ключевые слова: эндобионтные инфузории, Entodinium, Сapreolus pygargus
DOI: 10.31857/S0031184721060028
Эндобионтные инфузории были обнаружены в пищеварительном тракте различных
позвоночных животных; их наибольшее разнообразие отмечается у парнокопытных
(преимущественно жвачных) и непарнокопытных млекопитающих (Корнилова, 2004;
Vd’acny, 2018). В настоящее время не вызывает сомнений, что представители различ-
ных систематических групп позвоночных (ранга отряда и выше) чётко различаются по
видовому составу сообществ эндобионтов (Корнилова, 2004а; Vd’acny, 2018; Cedrola
et al., 2020). При этом видовое разнообразие и количество инфузорий во многом
определяются особенностями биологии хозяина. В то же время вопрос о том, на-
сколько различные виды эндобионтов специфичны по отношению к определенному
виду хозяина, остаётся открытым. Для его решения необходимым представляется
465
сравнительное изучение фауны эндобионтных инфузорий у максимально широкого
спектра хозяев, в первую очередь в естественных местообитаниях. Подобное исследо-
вание позволит определить уровень различий в видовом составе сообществ инфузорий
у хозяев, различающихся по степени родства, и выявить виды эндобионтов, более или
менее специфичных по отношению к хозяевам. В связи с этим необходимо отметить,
что к настоящему времени детально исследованы сообщества инфузорий-эндобионтов
пищеварительного тракта одомашненных видов жвачных и лошадиных, в то время
как сведения об эндобионтах диких млекопитающих в природных популяциях край-
не скудны (Ito et al., 2014, 2020; Kittleman et al., 2015; Newbold et al., 2015; Cedrola
et al., 2019).
В настоящей работе представлены результаты исследования фауны эндобионтных
инфузорий рубца косули сибирской Capreolus pygargus (Pallas, 1771), полученные
данные обсуждаются в связи с проблемой идентификации видов рода Entodinium.
МАТЕРИАЛ И МЕТОДЫ
Исследование проведено на материале инфузорий из рубца 13 особей (четыре самки и
девять самцов) косули сибирской Сapreolus pygargus, добытых на территории Тернейского
района Приморского края (табл. 1). Пробы фиксировали в 4% формалине в соотношении 1:1.
Содержимое рубца косуль № 4, 7, 8, 9 и 11 было предварительно заморожено и сохранялось
так до фиксации от 4 дней (№ 11) до 13 месяцев (№ 4).
Светомикроскопические исследования и микрофотографирование проводили с использо-
ванием микроскопов МБИ-11, Альтами-Инверт-3 с фотонасадкой и Leica DM2500, оснащен-
ного дифференциальным интерференционным контрастом и цифровой камерой Leica DFC495
(8.0MP).
При изучении морфологии клетки для выявления макронуклеуса использовали 0.1% раствор
метилового зелёного в 1% растворе уксусной кислоты. Численность инфузорий в 1 мл содер-
жимого рубца определяли методом «калиброванной капли» (Корнилова, 2004б). Для видового
определения использовали работы Buisson (1923а, 1923b), Догеля (1929), Williams, Coleman
(1992) и Dehority (1993).
Для проведения филогенетического анализа были использованы нуклеотидные последо-
вательности гена 18S рРНК, полученные из базы данных GenBank для 23 различных видов
энтодиниоморфид (рис. 2). Множественное выравнивание образцов производили алгоритмом
ClustalW в интерфейсе программного обеспечения MEGA X. Для построения филогенетическо-
го дерева применяли метод максимального правдоподобия, а также модель эволюции General
Time Reversible (GTR) с гамма-распределением (5 категорий, (+ G, a = 0.6369) и оценкой
инвариантных сайтов (+ I, 63.45% позиций) (Nei, Kumar, 2000). Достоверность полученного
дерева определяли bootstrap-методом (1000 повторений); оптимальный выбор модели прово-
дили по Байесовскому информационному критерию. Для анализа использовалась программа
MEGA X (Kumar et al., 2018).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Эндобионтные инфузории были обнаружены в содержимом рубца пяти из три-
надцати косуль: у двух самок и трёх самцов. По нашим наблюдениям, хранение со-
держимого рубца в замороженном состоянии до отбора проб не оказывало суще-
ственного влияния на сохранность инфузорий. Так, среди проб из замороженного
материала инфузории были диагностированы в двух из пяти желудков косуль (40%),
а среди проб из свежего материала инфузории были обнаружены в трех из восьми
желудков (38%). Нами не было выявлено различий в пригодности клеток инфузорий,
466
фиксированных из замороженного или свежего материала, для микроскопирования и
дальнейшего изучения. В пробах содержимого рубца всех пяти особей косуль-хозяев
эндобионтных инфузорий был обнаружен единственный вид инфузорий Entodinium
dubardi Buisson, 1923 (Entodiniomorphida, Ophryoscolecidae) (рис. 1).
Таблица 1. Сводные данные по собранному материалу содержимого рубца косули сибирской
Capreolus pygargus
Table 1. Data on the material of the rumen contents of the Siberian roe deer Capreolus pygargus
Условия хранения
Колич.
№
Дата взятия
материала
Место сбора / пол животного
инфузорий
пробы
пробы
до фиксации
в 1 мл
в формалине
1
08.01.2016
Приморский край, Тернейский район,
Не более 3 ч,
0
р. Живописная / самка
заморозки
не было
2
08.01.2016
Приморский край, Тернейский район,
Около 1 ч,
0
р. Севастьяновка (бассейн р. Максимовка) /
заморозки
самец
не было
3
08.01.2016
Приморский край, Тернейский район,
Около 1 ч,
0
р. Севастьяновка (бассейн р. Максимовка) /
заморозки
самка
не было
4
19.05.2015
Приморский край, Тернейский район,
Был
0
(фиксация -
пос. Терней / самец
заморожен
22.06.2016)
5
22.10.2017
Приморский край, Тернейский район,
Несколько часов,
920 000
пос. Пластун / самка
заморозки
не было
6
13.01.2018
Приморский край, Тернейский район,
Несколько часов,
0
ключ Комсомольский (бассейн р. Амгу) /
заморозки
самец
не было
7
20.12.2017
Приморский край, Тернейский район,
Был
0
(фиксация -
р. Западная Кема (бассейн р. Кема) / самец
заморожен
12.01.2018)
8
21.12.2017
Приморский край, Тернейский район, р
Был
0
(фиксация -
. Западная Кема (бассейн р. Кема) / самец
заморожен
12.01.2018)
9
20.11.2018
Приморский край, Тернейский район,
Был
610 000
(фиксация -
окрестности пос. Терней / самка
заморожен
25.12.2018)
10
06.07.2019
Приморский край, Тернейский район,
11 ч
740 000
окрестности пос. Терней, оз. Утиное /
при тёплой
самец
погоде
11
16.02.2020
Приморский край, Тернейский район, ключ
Был
680
(фиксация -
Петлёвочный (бассейн р. Серебрянка) /
заморожен
20.02.2020)
самец
12
19.06.2020
Приморский край, Тернейский район, ключ
6 ч
320 000
Петлёвочный (бассейн р. Серебрянка) /
при тёплой
самец
погоде
13
28.12.2020
Приморский край, Тернейский район,
Около 3 ч,
0
р. Севастьяновка (бассейн р. Максимовка) /
заморозки
самец
не было
467
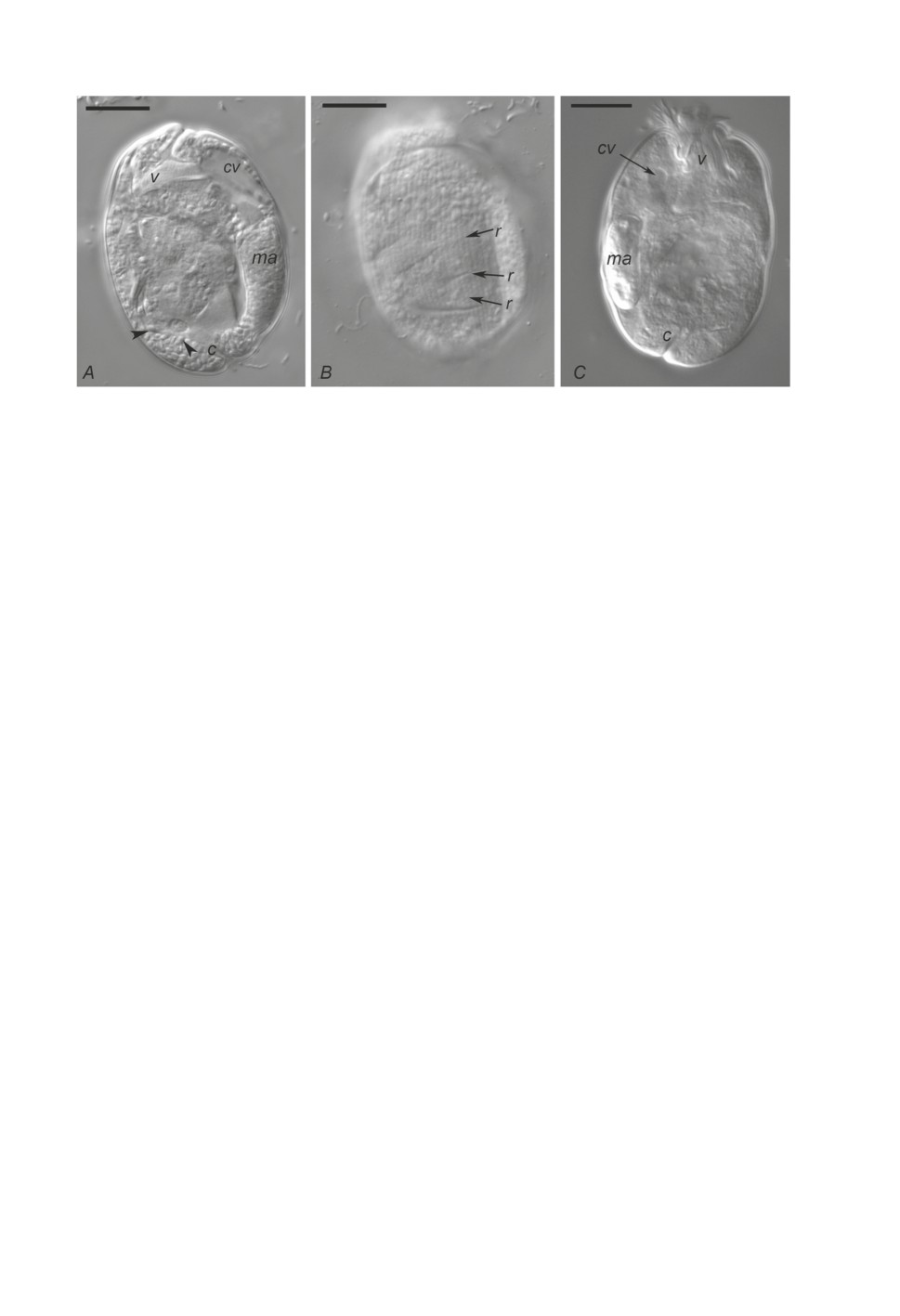
Рисунок 1. Entodinium dubardi. Световая микроскопия, DIC. А, С - детали строения клетки;
В - гребни на поверхности клетки. v - вестибулюм, ma - макронуклеус, cv - сократительная
вакуоль, с - цитопрокт, r - гребни на поверхности клетки, стрелки - включения
(пищевые частицы). Масштабная линейка 10 мкм.
Figure 1. Entodinium dubardi. Light microscopy, DIC. А, C - details of the cell morphology;
B - ridges on the cell surface. v - vestibulum, ma - macronucleus, cv - contractile vacuole,
с - cytoproct, r - ridges on the cell surface, arrowheads - inclusions (food particles).
Scale bars 10 µm.
Размеры инфузорий составляли в среднем 40.1 ± 4.8 (28.8-51.8) мкм в длину
и 26.4 ± 2.4 (20.1-34.5) мкм в ширину, отношение длины к ширине 1.4-1.6 (табл. 2,
рис. 1А, 1С). На их поверхности обычно были видны регулярно расположенные
продольные гребни толщиной около 0.6 мкм, тянущиеся от переднего конца клетки
к заднему параллельно друг другу, их число на одной стороне клетки составляло
38-40 (рис. 1В). В пробах встречались инфузории как с втянутыми, так и с расправ-
ленными околоротовыми ресничками. На заднем конце клетки был хорошо заметен
цитопрокт, его форма и расположение характерны для E. dubardi - выводное отверстие
открывается не на полюсе, а слегка смещено на дорсальную сторону клетки, сам же
выводной канал косо направлен на вентральную сторону, в среднем его длина 6 мкм
(рис. 1А, 1С). Эктоплазма на вентральной стороне клетки утолщена позади выводного
канала цитопрокта. Цитоплазма многих энтодиниумов содержала большое количество
мелких включений, вероятно, пищевых частиц (рис. 1А). Колбасовидный макрону-
клеус, иногда расширенный на переднем конце или на обоих концах, располагался в
средней части клетки (рис. 1А, 1С). Длина макронуклеуса составляла 35-50% длины
тела инфузории, расстояние от переднего конца клетки до переднего края макрону-
клеуса составляло 15-45% длины тела.
Делящиеся клетки энтодиниумов встречались редко. Количество инфузорий в 1 мл
содержимого рубца у косуль № 5, 9, 10 и 12 было сравнительно большим и состав-
ляло от 320 000 (№ 12) до 920 000 (№ 5), тогда как у косули № 11 в 1 мл рубца
содержалось всего 680 инфузорий.
468
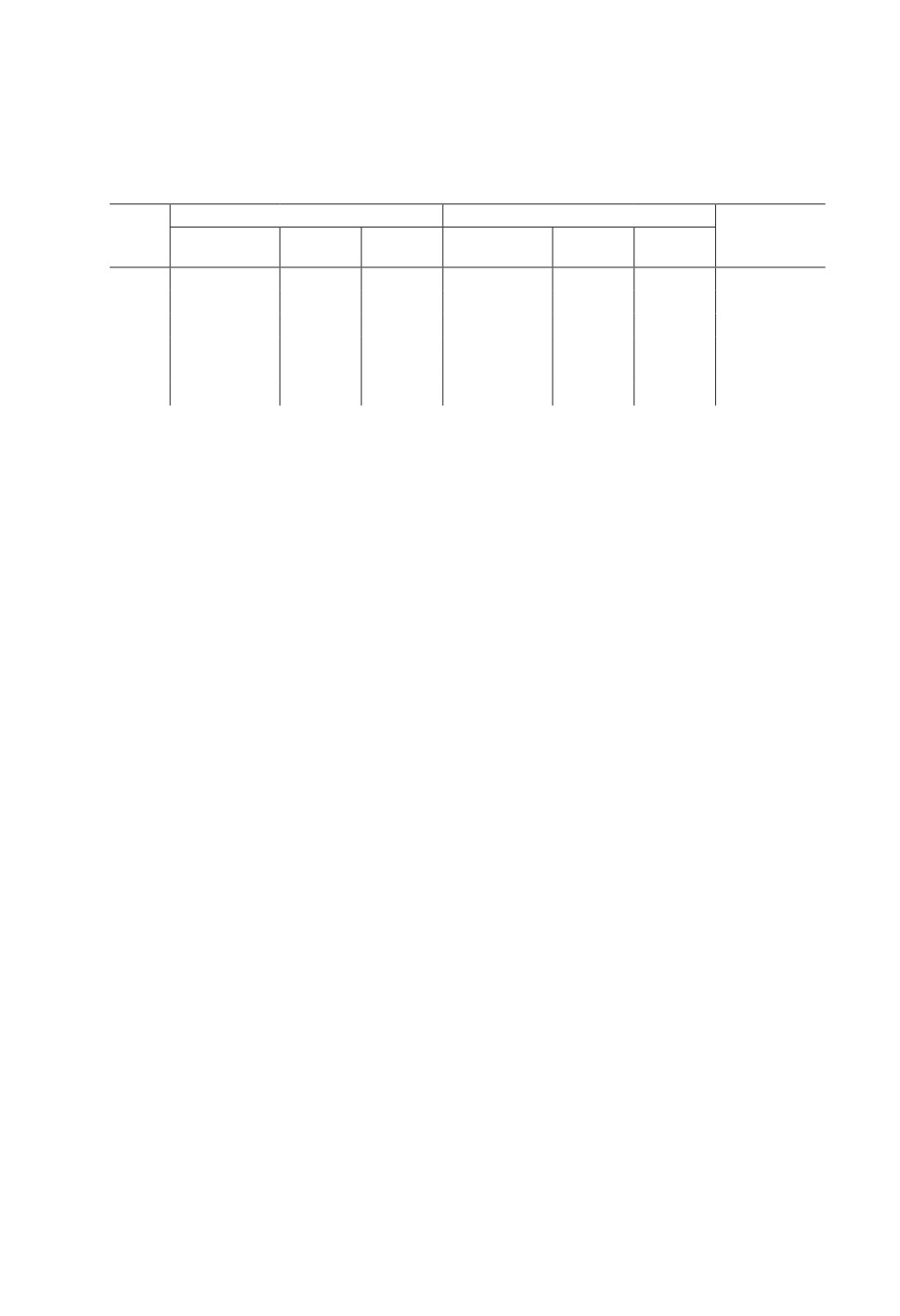
Таблица 2. Морфометрические показатели Entodinium dubardi из рубца косули
сибирской Capreolus pygargus (Тернейский р-н Приморского края)
Table 2. Morphometric parameters of Entodinium dubardi from the rumen of Siberian
roe deer Capreolus pygargus (Terneisky district of Primorsky Krai)
№
Длина, мкм
Ширина, мкм
Отношение
пробы
Среднее
max
min
Среднее
max
min
длины
к ширине
5
40.0 ± 0.48
51.8
31.6
27.6 ± 0.74
34.5
23.0
1.3-1.9
9
37.7 ± 0.47
46.0
28.8
26.6 ± 0.22
28.8
20.1
1.3-1.8
10
39.2 ± 0.94
48.9
31.6
25.2 ± 0.34
31.6
23.0
1.3-1.8
11
36.6 ± 0.65
40.3
28.8
24.8 ± 0.47
25.9
20.1
1.2-1.8
12
39.8 ±1.72
48.9
34.5
25.5 ± 0.51
28.8
23.0
1.3-1.8
Общее
39.6 ± 0.58
51.8
28.8
26.2 ± 0.55
34.5
20.1
1.2-1.9
П ри м ечан ие. Количество измеренных клеток для всех проб n=25.
Из всех трихостоматид род Entodinium наиболее богат видами - их описано более
сотни (Williams, Coleman, 1992). В то же время видоспецифичных признаков крайне
мало: длина и ширина тела, размер, форма и расположение макронуклеуса, располо-
жение сократительной вакуоли (см., например, Dehority, 1993). Соматическая цилиа-
тура у энтодиниумов отсутствует, а околоротовая цилиатура обычно полностью или
частично втянута в вестибулюм. Рисунки во многих публикациях, особенно ранних,
малоинформативны. Часто при описании энтодиниумов авторы руководствовались
формой и размерами каудальных отростков, однако в ряде экспериментов in vivo
(Стрелков, Полянский, 1937; Poljansky, Strelkow, 1938) была доказана внутривидовая
изменчивость этого признака у офриосколецид (в том числе у одного из наиболее рас-
пространённых видов - Entodinium caudatum) в зависимости от пищевого режима и
присутствия хищных инфузорий. Было установлено, что у одной и той же популяции
энтодиниумов каудальные отростки могут увеличиваться или уменьшаться вплоть до
полного исчезновения при изменении условий внешней среды.
В истории изучения офриосколецид неоднократно предпринимались попытки ре-
визии рода Entodinium и выявления дополнительных дифференциальных признаков
для отдельных видов (Kofoid, MacLennan, 1930; Lubinsky, 1957; Latteur, 1968, 1969;
Ogimoto, Imai, 1981). Тем не менее проблемы с идентификацией видовой принадлеж-
ности энтодиниумов существуют до сих пор.
Вид E. dubardi был впервые обнаружен в рубце косули европейской Сapreolus
capreolus (Linnaeus, 1758) и описан в 1923 г. (Buisson, 1923a, 1923b). В.А. Догель
(Dogiel, 1925, 1927) обозначил эту инфузорию как E. dubardi forma dubardi, тем самым
отделив её от вооружённых шипами форм. Любинский (Lubinsky, 1957) предложил
для энтодиниумов три универсальные формы - "caudatum" (с дорсальным шипом и
двумя вентральными лопастями), "loboso-spinosum" (с дорсальным шипом и одной
вентральной лопастью) и "dubardi" (без каудальных отростков). При этом описанный
Догелем E. simplex Dogiel, 1927 был признан синонимом E. dubardi Buisson, 1923.
469
Latteur (1968, 1969) провёл значительную ревизию рода Entodinium, предложив
в качестве важного определительного признака взаимное расположение ядра и со-
кратительной вакуоли. Очень многие виды были сведены в синонимику, в том чис-
ле E. dubardi, который вместе с E. nanellum Dogiel, 1923, E. simplex Dogiel, 1925,
E. elongatum Dogiel, 1927 и рядом других видов был сведён к E. furca Cunha, 1914.
Однако, согласно первоописанию Cunha (1914), E. furca значительно отличается от
E. dubardi: если первый имеет два широких заострённых хвостовых отростка (что
и дало название), для второго как раз характерно полное отсутствие каких бы то
ни было шипов; таким образом, правомочность сведения вида E. dubardi в синоним
E. furca вызывала сомнения. Кроме того, были высказаны предположения (Poljansky,
Strelkow, 1938; Hungate, 1978), что расположение сократительной вакуоли в клетке
энтодиниумов может варьировать внутри вида.
Несмотря на проведённые ревизии рода, многие исследователи рассматривали
E. dubardi как самостоятельный вид. Так, Wertheim (1933) полагал, что E. dubardi
чётко отличается от других похожих видов заметной скошенностью выводного ка-
нала цитопрокта и слегка уменьшенной задней частью эндоплазматического мешка,
следовательно, вид валиден. Ogimoto и Imai (1981) также признавали E. dubardi са-
мостоятельным видом и предлагали в качестве характерного признака использовать
особую форму макронуклеуса - он немного более толстый на концах и более тонкий
в средней части, тогда как у других близких видов макронуклеус в средней части
более утолщенный, чем на концах. В известном определителе Dehority (1993) вид
E. dubardi валиден и имеет следующие признаки: отсутствие хвостовых шиповидных
выростов, средние показатели длины тела - 41 мкм, ширины - 30 мкм, отношения
длины к ширине - 1.4, макронуклеус короче длины тела и лежит в средней части
клетки, эктоплазма утолщена на заднем конце тела. Дифференциальные признаки,
указанные Dehority, согласуются с диагнозом E. dubardi, приведенным в первоопи-
сании (Buisson, 1923a, 1923b), и были использованы нами при определении видовой
принадлежности обнаруженных инфузорий.
Мы проанализировали немногочисленные имеющиеся в Генбанке последователь-
ности гена 18S РНК видов рода Entodinium. Согласно результатам молекулярно-
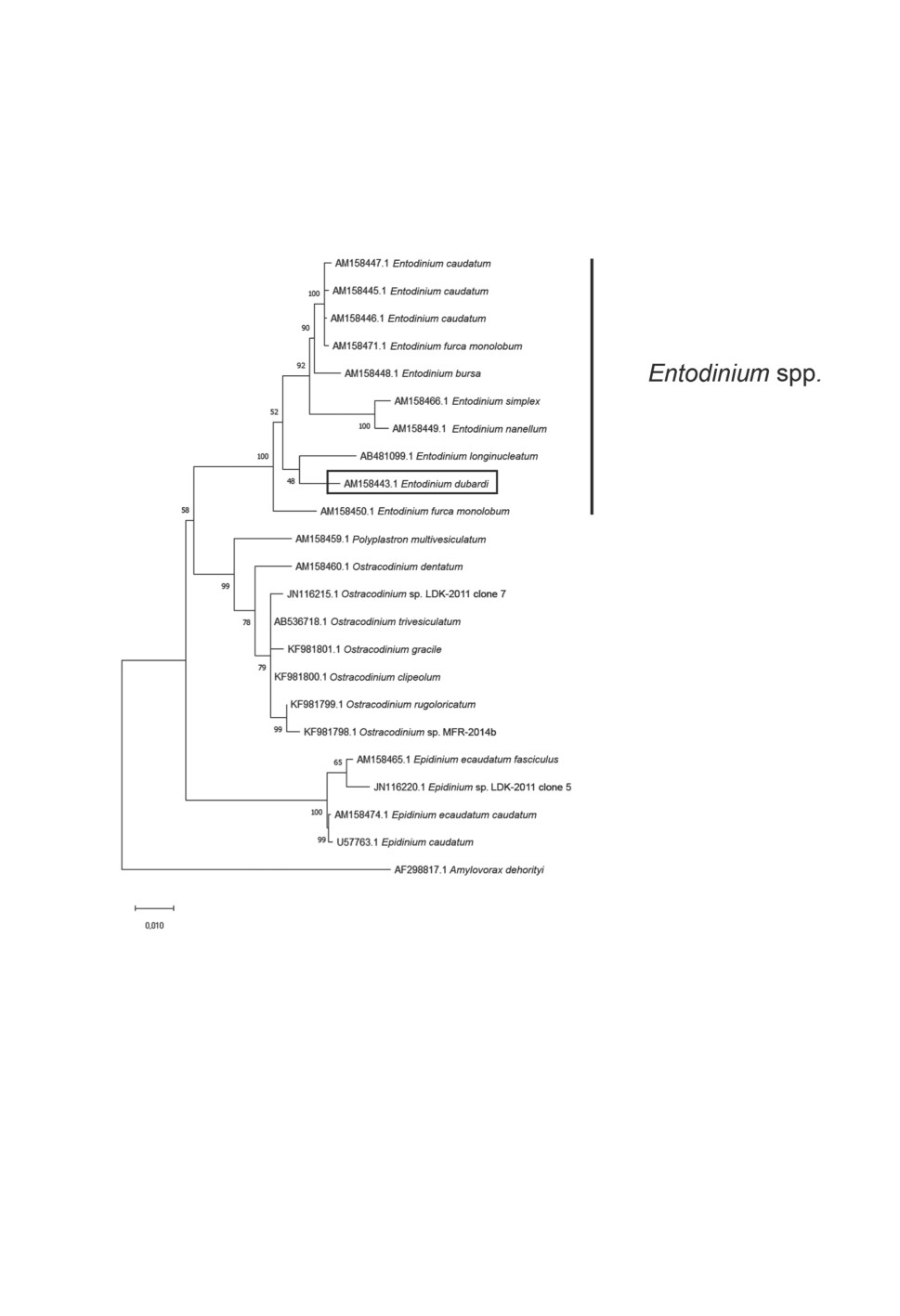
филогенетического анализа, энтодиниумы формируют единую чётко выраженную
кладу на филогенетическом дереве (рис. 2). Филогенетические связи между отдельны-
ми видами в настоящее время уверенно определить не удаётся. Однако можно заклю-
чить, что вид E. dubardi заметно отличается от остальных энтодиниумов, включённых
в анализ, и его валидность не вызывает сомнений. Таким образом, по нашему мне-
нию, вид E. dubardi валиден и является единственным представителем эндобионтных
инфузорий для исследованных нами косуль сибирских из Тернейского района При-
морского края России.
В отношении эндобионтов из рубца косули европейской были опубликованы обоб-
щающие данные по нескольким источникам (Imai et al., 1983; Williams, Coleman, 1992;
Clauss et al., 2011). В этих работах сообщается о нахождении инфузорий, преимуще-
ственно из рода Entodinium, от одного до 7-8 видов, а также приводятся данные о том,
что в рубцах более чем 50% обследованных косуль инфузории обнаружены не были.
470
При этом, согласно большинству исследований, у косули европейской находили только
один вид инфузорий E. dubardi (Buisson, 1923a, 1923b; Sladecek, 1946; Brüggemann
et al., 2009). Лишь в одной работе по эндобионтам рубца косули европейской указано
нахождение, кроме энтодиниумов, ещё нескольких видов инфузорий, как изотрихид,
так и офриосколецид, и при этом доля энтодиниумов составляла 91.9% (Enzinger,
Hartfiel, 1998).
Рисунок 2. Виды рода Entodinium на филогенетическом древе офриосколецид.
Филогенетическое дерево построено по последовательностям гена 18S рРНК методом
максимального правдоподобия, эволюционная модель GTR + G + I. Цифры рядом с узлами
дерева отражают показатели bootstrap-теста (процент деревьев из 1000 повторений).
Масштаб соответствует 1 нуклеотидной замене на каждые 100 нуклеотидов.
Figure 2. Phylogenetic tree showing the position of Entodinium species among representatives
of the family Ophryoscolecidae. Maximum likelihood phylogenetic tree inferred from 18S rRNA
gene sequences by MEGA X, based on the General Time Reversible (GTR) model with gamma
distribution and estimate of invariable sites. The numbers at the nodes are the bootstrap values
(percent out of 1000 replicates). The tree is drawn to scale, the scale bar represents
1 change per 100 positions.
471
Фауна эндобионтных инфузорий рубца косули сибирской была исследована ра-
нее в Омской и Челябинской областях России, а также в Якутии (Корнилова и др.,
2004; Аргунов, Степанова, 2015). В Омской области было исследовано 11 косуль,
у всех были обнаружены инфузории, и всего было диагностировано 10 видов инфу-
зорий - офриосколециды и изотрихиды. Из них 6 - энтодиниумы, среди которых был
и E. dubardi. В Челябинской области исследованы 10 косуль, также все с эндоби-
онтами, всего найдено 27 видов инфузорий, из них 16 - энтодиниумы, в том числе
E. dubardi. В Якутии, как и в нашем исследовании, у косули были найдены только
E. dubardi (Аргунов, Степанова, 2015).
В литературе до сих пор не существует единой точки зрения относительно видо-
вого состава рода Entodinium, и фаунистические списки эндобионтных инфузорий,
опубликованные разными авторами, различаются. Такие различия не всегда позволяют
делать однозначные выводы о характере распределения отдельных видов энтодиниу-
мов в различных популяциях хозяев. Однако мы можем заключить, что вид E. dubardi
является основным эндобионтом для большинства исследованных косуль европейских,
а также для косуль сибирских в Якутии и в Приморском крае.
В нашем исследовании эндобионтные инфузории были обнаружены менее чем
у 40% особей косули сибирской, этот показатель близок к 50% встречаемости инфу-
зорий у косули европейской, о чём было сказано выше. У подавляющего большин-
ства обследованных жвачных инфузории обычно присутствуют, и для обьяснения
относительно бедного состава эндобионтной фауны у косуль выдвигались различные
гипотезы. Так, Clauss et al. (2011) основной причиной назвали видоспецифичную дие-
ту животных-хозяев, отметив, что преобладание энтодиниумов над другими видами
инфузорий характерно для рубца копытных, в рационе которых преобладает веточ-
ный корм. Так, например, подобный рацион характерен для большинства оленевых,
и у них нередко диагностируют низкое видовое разнообразие инфузорий или даже
их отсутствие. Clauss et al. (2011) попытались выявить зависимость фаунистического
разнообразия эндобионтов рубца от массы тела хозяина, однако такая зависимость
не была установлена.
Dehority и Orpin (1997) указывали, что энтодиниумы способны жить в более кис-
лой среде, чем другие виды инфузорий, но в некоторых экспериментах связь фауни-
стического состава с кислотностью содержимого рубца не подтвердилась. В этой же
публикации показано значительное снижение численности инфузорий при длительных
промежутках времени (12 ч и более) между кормлениями животных-хозяев. Авторы
связывают этот факт с изменением кислотности в рубце во время голодания живот-
ного. В исследованиях Trabalza et al. (2005) было показано, что содержимое рубца
косули имеет среднее значение рН = 5.7. Такая относительно кислая среда может
быть приемлемой для энтодиниумов, но непригодной для обитания других инфузорий.
Существует также гипотеза о том, что видовой состав инфузорий зависит от ко-
личественного и видового состава бактерий в рубце, сформировавшегося в связи
с различными пищевыми предпочтениями разных видов эндобионтов. Согласно дан-
ным Østbye et al. (2016), микробиота косули значительно отличается от таковой других
исследованных оленевых (лось Alces alces (Linnaeus, 1758), благородный олень Cervus
472
elaphus Linnaeus, 1758) из той же среды обитания. Авторы предполагают, что при-
чиной тому могут быть различия в физиологии рубца. По нашему мнению, состав
микрофлоры может влиять на численность инфузорий, но не может способствовать
видовому обогащению их фауны. Высказывалось также мнение о том, что поедание
млекопитающими токсичных растений может вызывать угнетение фауны инфузорий-
эндобионтов (Williams, Coleman, 1992). Однако этим нельзя обьяснить тот факт, что
у разных косуль из одного и того же местообитания инфузории в рубце могут при-
сутствовать в большом количестве или же отсутствовать вовсе.
Согласно гипотезе Догеля (1946), поддержанной Imai (1998), характерный «на-
бор» эндобионтных инфузорий у каждого вида жвачных сформировался в процессе
эволюции. Инфузории передаются от одного хозяина к другому преимущественно
в пределах узкой семейной группы жвачных. В связи с этим фаунистический набор
эндобионтных инфузорий мог не пополняться новыми видами на протяжении всего
периода времени после эволюционного разделения видов хозяев.
Мы предполагаем, что изначально у общих предков жвачных могла быть доста-
точно богатая фауна эндобионтных инфузорий, однако после отделения семейства
оленевых, перехода их на преимущественно древесный корм, ряд видов инфузорий
был утрачен. Далее в связи со снижением уровня стадности и, следовательно, умень-
шением возможности обмена жвачкой между особями хозяев, список видов инфузорий
ещё сильнее сократился, а в некоторых микропопуляциях косуль полностью исчез.
Согласно данным мониторинга состояния популяции косули сибирской в Приморском
крае, её численность резко сократилась в последние десятилетия, местами косуля
полностью вытеснена другими видами копытных, в первую очередь пятнистым оле-
нем Cervus nippon Temminck, 1838 (Пикунов и др., 2009). Этим может объясняться
более низкие разнообразие и встречаемость эндобионтных инфузорий у косуль При-
морского края в сравнении с косулями Омской и Челябинской областей.
БЛАГОДАРНОСТИ
Работа выполнена при финансовой поддержке темы государственного задания
AAAA-A19-119020690109-2 (Зоологический институт РАН).
СПИСОК ЛИТЕРАТУРЫ
Аргунов А.В., Степанова В.В. 2015. Зимнее питание сибирской косули (Сapreolus pygargus Рall., 1771) в
Центральной Якутии. Вестник КрасГАУ 4: 138-143. [Argunov A.V., Stepanova V.V. 2015. The winter
diet of the Siberian roe deer (Capreolus pygargus Pall., 1771) in the Central Yakutia. Vestn. KrasGAU
4: 138-143. (In Russian)]
Догель В.А. 1929. Простейшие - Protozoa. Малоресничные инфузории - Infusoria Oligotricha. Сем.
Ophryoscolecidae. 2. Определители по фауне СССР, издаваемые Зоологическим музеем Академии
Наук. Ленинград, Изд-во Акад. наук СССР, 97 c. [Dogiel V. 1929. Protozoa infusoires oligotriches fam.
Ophryoscolecida. 2. Academie des sciences de l'union des républiques soviétiques socialistes tableaux
analytiques de la faune le l'urss, publiés par le musée zoologique de l'académie des sciences. Leningrad,
97 pp. (In Russian)]
Догель В.А. 1946. Филогения инфузорий желудка жвачных в свете палеонтологических и эколого парази-
тологических данных. Зоологический журнал 25 (5): 1162-1188. [Dogiel V.A. 1946. Philogenia infu-
soriy gheludka ghvachnych v svete paleontologicheskich I ecologoparasitologicheskich dannych. Zooogical
Journal 25 (5): 1162-1188. (In Russian)]
473
Корнилова О.А. 2004а. История изучения эндобионтных инфузорий млекопитающих. СПб., ТЕССА, 352
с. [Kornilova O.A. 2004. History of study of endobiotic ciliates of mammalia. St-Petersburg, TESSA,
352 pp. (In Russian)]
Корнилова О.А. 2004б. Метод комплексного обследования фауны эндобионтных инфузорий. В сб. Функц.
морф., экол. и жизн. циклы жив. Научн. тр. каф. зоол. РГПУ им. А.И. Герцена. Вып. 4. СПб., ТЕССА,
75-77. [Kornilova O.A. 2004b. The method of combined investigations of endobiotic ciliates. Sci. J. Dept.
Zool. Herzen SP Univ. of Russia, 4: 75-77. (In Russian)]
Корнилова О.А., Федорова П.Н., Мачахтыров Г.Н., Баймакова Л.Г. 2004. Биоразнообразие инфузорий из
кишечника лошади (Equus caballus), желудка косули (Capreolus pygargus) и северного оленя (Rangifer
tarandus) таежной зоны Сибири. В сб. Функц. морф., экол. и жизн. циклы жив. Научн. тр. каф. зоол.
РГПУ им. А.И. Герцена. Вып. 4. СПб., ТЕССА, 55-63. [Kornilova O.A., Fedorova P.N., Machactyrov
G.N., Baymakova L.G. 2004. The biodiversity of ciliates from the intestine of the horse (Equus caballus),
from the rumen of roe deer (Capreolus pygargus) and reindeer (Rangifer tarandus) in Siberian taiga. Sci.
J. Dept. Zool. Herzen SP Univ. of Russia, 4: 55-63. (In Russian)]
Пикунов Д.Г., Середкин И.В., Арамилев В.В., Николаев И.Г., Мурзин А.А. 2009. Крупные хищники и
копытные юго-запада Приморского края. Владивосток, Дальнаука, 96 с. [Pikunov D.G., Seryodkin
I.V., Aramilyov V.V., Nikolaev I.G., Murzin A.A. 2009. Krupnye khishchniki i kopytnye iugo-zapada
Primorskogo kraia. Vladivostok, Dalnauka, 96 pp. (In Russian)]
Стрелков А., Полянский Ю. 1937. О процессе естественного отбора у некоторых Infusoria Entodiniomorphida.
Зоологический журнал 16 (1): 77-87. [Strelkov A., Polyansky Yu. 1937. O processe estestvennogo otbora
u nekotorych Infusoria Entodiniomorphida. Zool. Journal. 16 (1):77-87. (In Russian)]
Brüggemann J., Giesecke D., Walser-Kärst K. 2009. Beiträge zur Wildbiologie und vergleichenden
Tierphysiologie. 1. Mikroorganismen im Pansen von Rothirsch (Cervus elaphus) und Reh (Capreolus
capreolus). Zeitschrift Für Tierphysiologie Tierernährung Und Futtermittelkunde 23 (1-5): 143-151.
https//: doi.org:10.1111/j.1439-0396. 1967. tb00972.x
Buisson J. 1923a. Les infusoires cilies du tube digestif de l'homme et des mammiferes. Paris, Legrand,
Travail du Laboratorie de Parasitologie de la Faculte de Medicine de Paris, 201 pр.
Buisson J. 1923b. Sur quelques infusoires nouveaux ou peu connus parasites des mammiferes. Ann. Parasitol.
Hum. Comp. 1: 209-246.
Cedrola F., Senra M., Rossi M.F., Fregulia P., D’Agosto M., Dias R. 2020. Trichostomatid Ciliates (Alveolata,
Ciliophora, Trichostomatia) Systematics and Diversity: Past, Present, and Future. Front. Microbiol. 10:
2967. https//:doi.org:0.3389/fmicb.2019.02967
Clauss M., Müller K., Fickel J., Streich W. J., Hatt J.-M., Südekum K.-H. 2011. Macroecology of the host
determines microecology of endobionts: protozoal faunas vary with wild ruminant feeding type and body
mass. J. Zool. 283: 169-185. https//:doi.org:10.1111/j.1469-7998.2010.00759
Cunha A.M. 1914. Ueber die Ziliaten, welche in Brasilien im Magen von Rindern und Schafen vorkommen.
Memorias do Instituto Oswaldo Cruz 6: 58-67.
Dehority B.A. 1993. Laboratory Manual for Classification and Morphology of Rumen Ciliate Protozoa. CRC
Press, Inc., Boca Raton, FL, 128 рp.
Dehority B.A. Orpin C.G. 1997. Development of, and natural fluctuations in rumen microbial populations. In: The
Rumen Microbial Ecosystem, 2nd ed., 196-235. https//:doi.org:10.1007/978-94-009-1453-7_5
Dogiel V.A. 1925. Nouveaux infusiores de la famille de Ophryoscolecidae parasites d'antilopes africaines //
Annales de Parasitologie Humaine et Comparee 3: 116-142.
Dogiel V.A. 1927. Monographie der Familie Ophryoscolecidae. Arch. Protistenk. 59: 1-288.
Enzinger W., Hartfiel W. 1998. Auswirkungen gesteigerter Energie- und Proteingehalte des Futters auf
Fermentationsprodukte, Fauna und Schleimhaut des Pansens von Wildwiederkäuern (Damhirsch/Reh) im
Vergleich zu Hauswiederkäuern (Schaf/Ziege). Z. Jagdwiss 44: 201-220.
Hungate R.E. 1978. The rumen protozoa. In: Parasitic Protozoa. London, Academic Press, vol. 2, 655-695.
Imai S. 1998. Phylogenetic Taxonomy of Rumen Ciliate Protozoa Based on Their Morphology and Distribution.
Imai S., Kinoshita M., Ogimoto K. 1983. Distribution of the rumen ciliate Protozoa in the deer. New Strateg.
Improv. Anim. Prod. Hum. Welfare. Proc. 5 World Conf. Anim. Prod., Tokyo, Aug. 14-19, 1983, 2. Free
Commun. Pap. Tokyo 1983, 337-338.
474
Ito A., Ishihara M., Imai S. 2014. Bozasella gracilis n. sp. (Ciliophora, Entodiniomorphida) from Asian elephant
and phylogenetic analysis of entodiniomorphids and vestibuliferids. Eur. J. Protistol. 50: 134-152.
Ito A., Eckardt W., Stoinsk T.S., Gillespie T.R., Tokiwa T. 2020. Three new Prototapirella species, Opisthotrichum
janus, and Troglocoryscava add to Entodiniomorphida (Ciliophora, Trichostomatia) diversity in mountain
Kittelmann S., Devente S.R., Kirk M.R., Seedorf H., Dehority B.A., Janssen P.H. 2015. Phylogeny of
intestinal ciliates, including Charonina ventriculi, and comparison of microscopy and 18S rRNA gene
pyrosequencing for rumen ciliate community structure analysis. Appl. Environ. Microbiol. 81: 2433-2444.
Kofoid C.A., MacLennan R.F. 1930. Ciliates from Bos indicus Linn. I. The genus Entodinium Stein. Univ. Cal.
Publ. Zool. 33: 471-544.
Kumar S., Stecher G., Li M., Knyaz C., Tamura K. 2018. MEGA X: Molecular Evolutionary Genetics Analysis
Latteur B. 1968. Revision systématique de la Famille des Ophryoscolecidae Stein, 1858: sous-Famille des
Entodiniinae Lubinsky, 1957, Genre Entodinium Stein, 1858. Ann. Soc. R. Zool. Belg. 98 (1): 1-4l.
Latteur B. 1969. Revision systématique de la famille des Ophryoscolecidae Stein, 1858: sous-famille des
Entodiniinae Lubinsky, 1957, Genre Entodinium Stein, 1858. Ann. Soc. R. Zool. Belg. 99 (1-2): 3-25.
Lubinsky G. 1957. Studies on the evolution of Ophryoscolecidae (Ciliata: Oligotricha). I. A new species of
Entodinium with 'caudatum', 'loboso-spinosum' and 'dubardi' forms and some evolutionary trends in the
genus Entodinium. Can. J. Zool. 35: 111-133.
Nei M., Kumar S. 2000. Molecular Evolution and Phylogenetics. New York, Oxford University Press, 333 pp.
Newbold C.J, de la Fuente G., Belanche A., Ramos-Morales E., McEwan N.R. 2015. The Role of Ciliate Protozoa
in the Rumen. Front. Microbiol. 6: 1313. https://doi.org: 10.3389/fmicb.2015.01313
Ogimoto K., Imai S. 1981. Atlas of Rumen Microbiology. Tokyo: Japan Scientific Societies Press. 231 p.
Østbye K., Wilson R., Rudi K. 2016. Rumen microbiota for wild boreal cervids living in the same habitat. FEMS
Poljansky G., Strelkow A. 1938. Étude expérimentale sur la variabilité de quelques Ophryoscolécidés. Arch.
Zool. Exp. Gen. 80: 1-123.
Sladecek F. 1946. Ophryoscolecidae z bachoru jelena (Cervus elaphus L.), dan'ka (Dama dama L.) a srnce
(Capreolus capreolus L.). Vestnik Ceskoslovenske Zoologicke Spolecnosti 10: 201-231.
Trabalza M.M., Capecci A., Riganelli N., Acuti G., Antonini C., Olivieri O. 2005. Dietary preferences and ruminal
protozoal populations in roe deer (Capreolus capreolus), fallow deer (Dama dama) and mouflon (Ovis
musimon). Ital. J. Anim. Sci. 4 (2): 401-403. https://doi.org: 10.4081/ijas.2005.2s.401
Vd’acny P. 2018. Evolutionary associations of endosymbiotic ciliates shed light on the timing of the marsupial-
Wertheim P. 1933. Ober die Beschaffenheit der Infusorien magen Fauna von Capreolus capreolus L. Zool. Anz.
106: 67-70.
Williams A.G., Coleman G.S. 1992. The Rumen Protozoa. Brock/Springer Series in Contemporary Bioscience,
ENDOBIOTIC CILIATES FROM THE RUMEN
OF THE ROE DEER CAPREOLUS PYGARGUS
O. A. Kornilova, L. V. Chistyakova, I. V. Seryodkin, I. P. Grabarnik
Keywords: endobiotic ciliates, Entodinium, Сapreolus pygargus
SUMMARY
The fauna of endobiotic ciliates of the rumen of the Siberian roe deer Capreolus py-
gargus from Terneisky region of Primorsky Territory of Russia was investigated. In the
investigated roe deer, a single ciliate species, Entodinium dubardi, was found, and it was
found in less than 40 % of hosts in the surveyed population. We assume that the extremely
low species diversity of endobiotic ciliates and the low degree of infestation of hosts are
primarily associated with a decrease in the level of herding, as well as with the peculiari-
ties of the diet of roe deer.
475