ПАРАЗИТОЛОГИЯ, 2021, том 55, № 6, с. 496-513.
УДК 576.895.42:595.421:578.4
АКТИВНОСТЬ ГЕМИПОПУЛЯЦИЙ ИМАГО ТАЁЖНОГО КЛЕЩА
(IXODES PERSULCATUS SCHULZE, 1930)
В СОЧЕТАННЫХ ПРИРОДНЫХ ОЧАГАХ
КЛЕЩЕВОГО ЭНЦЕФАЛИТА И ИКСОДОВЫХ КЛЕЩЕВЫХ
БОРРЕЛИОЗОВ ЮЖНОГО ПРИБАЙКАЛЬЯ
© 2021 г. О. В. Мельниковаа,*, Е. А. Вершинина, В. М. Корзуна,
Ю. А. Вержуцкаяа, Н. В. Яковчица, Р. В. Адельшина,б,
Ю. Н. Трушинаа, К. В. Лопатовскаяа, Е. И. Андаева
а ФКУЗ Иркутский научно-исследовательский противочумный институт Роспотребнадзора,
ул. Трилиссера, 78, Иркутск, 664047 Россия
б ФГБОУ ВО Иркутский государственный университет,
ул. К. Маркса, 1, Иркутск, 664003 Россия
*e-mail: melnikovaovit@gmail.com
Поступила в редакцию 19.03.2021 г.
После доработки 27.09.2021 г.
Принята к публикации 06.10.2021 г.
Приведены результаты многолетнего (2005-2020 гг.) мониторинга сочетанных природных
очагов клещевого энцефалита (КЭ) и иксодовых клещевых боррелиозов в южном Прибайкалье
на нескольких географически изолированных участках. Показана высокая степень неоднород-
ности обследованной территории по уровню обилия имаго Ixodes persulcatus, их зараженности
вирусом КЭ и боррелиями. Выявлены существенные различия между изученными участками
по патогенности переносчиков, содержащих маркеры вируса КЭ, для лабораторных животных.
Показано наличие прямой связи между численностью переносчика и обнаружением антигена
вируса КЭ, а также числом вирусных изолятов. Инцидентность КЭ на обследуемой террито-
рии также коррелировала с вирусофорностью по результатам иммуноферментного анализа и
с успешностью изоляции на теплокровных животных. Выявление маркеров боррелий в ПЦР
было выше в случае высокой численности переносчиков, но число заболеваний на обследуемой
территории не зависело ни от численности клещей, ни от их боррелиефорности.
Ключевые слова: таёжный клещ Ixodes persulcatus, численность, вирус клещево-
го энцефалита (ВКЭ), боррелии, вирусофорность, боррелиефорность, изоляты ВКЭ,
Прибайкалье
DOI: 10.31857/S0031184721060041
Клещевой энцефалит (КЭ) и иксодовые клещевые боррелиозы (ИКБ) - самые
распространенные трансмиссивные природноочаговые инфекции умеренного пояса
Евразии, представляющие большую проблему здравоохранения и наносящие значи-
496
тельный экономический ущерб. В азиатской части России основным резервуаром
и переносчиком патогенных агентов этих болезней - вируса клещевого энцефалита
(ВКЭ) и нескольких видов патогенных боррелий - является таёжный клещ Ixodes
persulcatus Schulze, 1930. Уровень заболеваемости КЭ и ИКБ не зависит напрямую
от численности основного переносчика ввиду сложности сочетания факторов (как
природных, так и социальных), и это сочетание обеспечивает своеобразие эпиде-
мической обстановки даже на рядом расположенных территориях (Якименко и др.,
2019). Тем не менее непременными условиями эффективной профилактики инфекций,
передаваемых клещами, являются определение динамики численности переносчи-
ка во времени и в пространстве и степени риска подвергнуться нападению клещей
в местах их обитания (Daniel et al., 2006). Структура популяций таёжного клеща отли-
чается исключительной сложностью. Одна генерация включает четыре фазы развития
(яйцо, личинка, нимфа, взрослые самки и самцы), и каждая фаза образует отдельную
гемипопуляцию, занимающую свою микросреду обитания. Обилие подстерегающих
голодных клещей в экосистеме определяется балансом между пополнением их по-
пуляции новыми особями и гибелью от различных причин (Osipova et al., 2017). При
наличии условий для существования клещей и их прокормителей размер популяции
I. persulcatus в значительной мере зависит от характера, сложности и расчлененности
рельефа, степени развития гидросети, площади и расположения безлесных участков
и некоторых других особенностей топографии местности (Коренберг, 1976). Как
результат, численность таёжного клеща в каждом регионе отличается большой пе-
стротой, и динамика его численности представляет собой сложный полициклический
процесс, складывающийся из квазипериодических колебаний с периодами разной
продолжительности (Коротков, 1998). Данчинова и соавт. (2004) выделяют в При-
байкалье три зоны с различным обилием клещей I. persulcatus: с высоким - Южное
Присаянье, средним - Верхнеленские районы, низким - степи, открытые лесостепные
участки, северные районы Иркутской области.
Цель данной работы - охарактеризовать активность гемипопуляций имаго таёж-
ного клеща, его инфицированность возбудителями КЭ и ИКБ на нескольких геогра-
фически изолированных участках южного Прибайкалья и оценить риск заражения
человека данными нозоформами на этих участках.
МАТЕРИАЛЫ И МЕТОДЫ
Работа проводилась в 2005-2020 гг. на территории Иркутского, Слюдянского, Шелеховского,
Баяндаевского и Эхирит-Булагатского районов Иркутской области, географически ограничи-
ваясь 51.39°-52.98° N и 102.53°-106.17° E с высотами 459-943 м над ур. м. Наблюдениями,
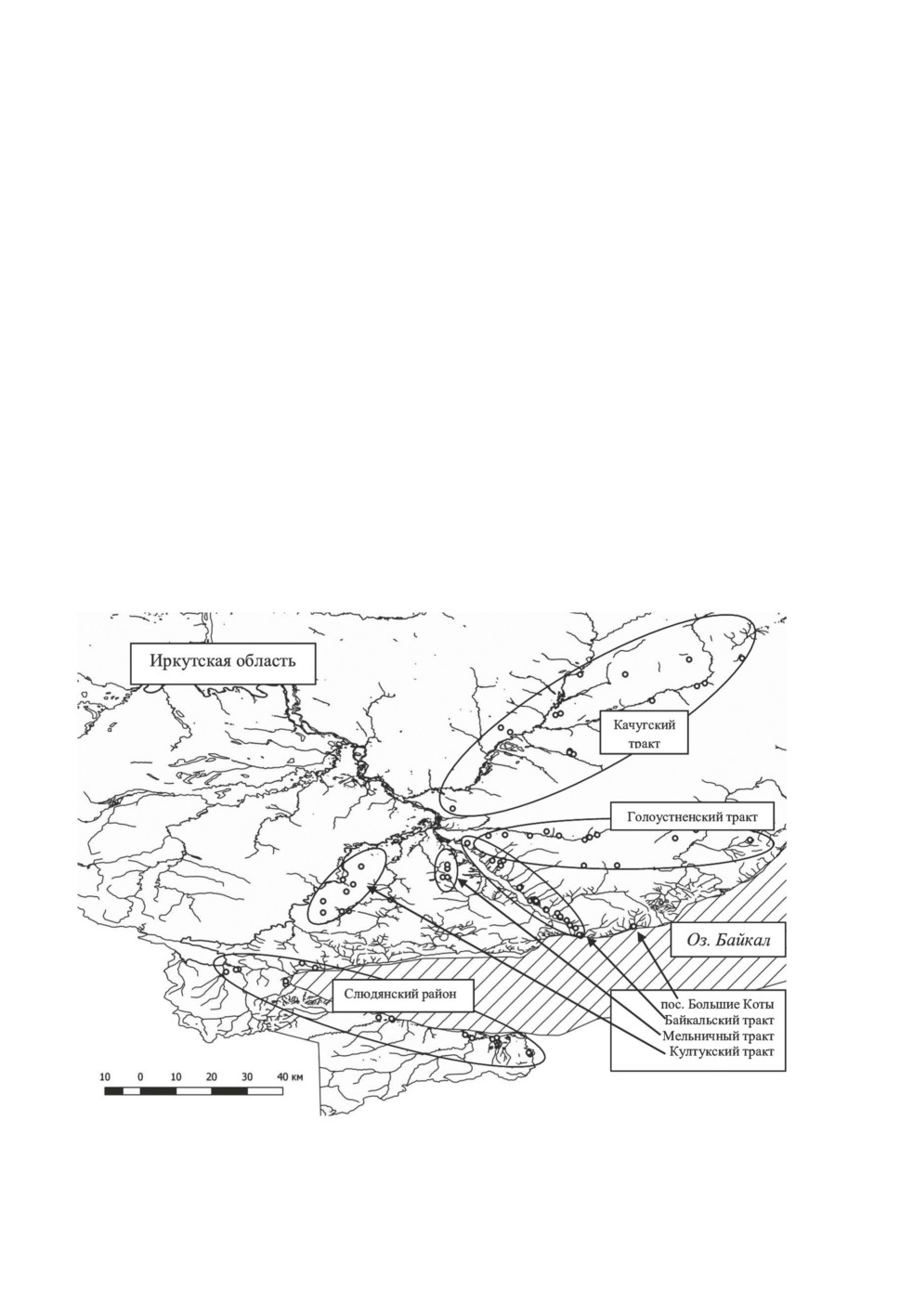
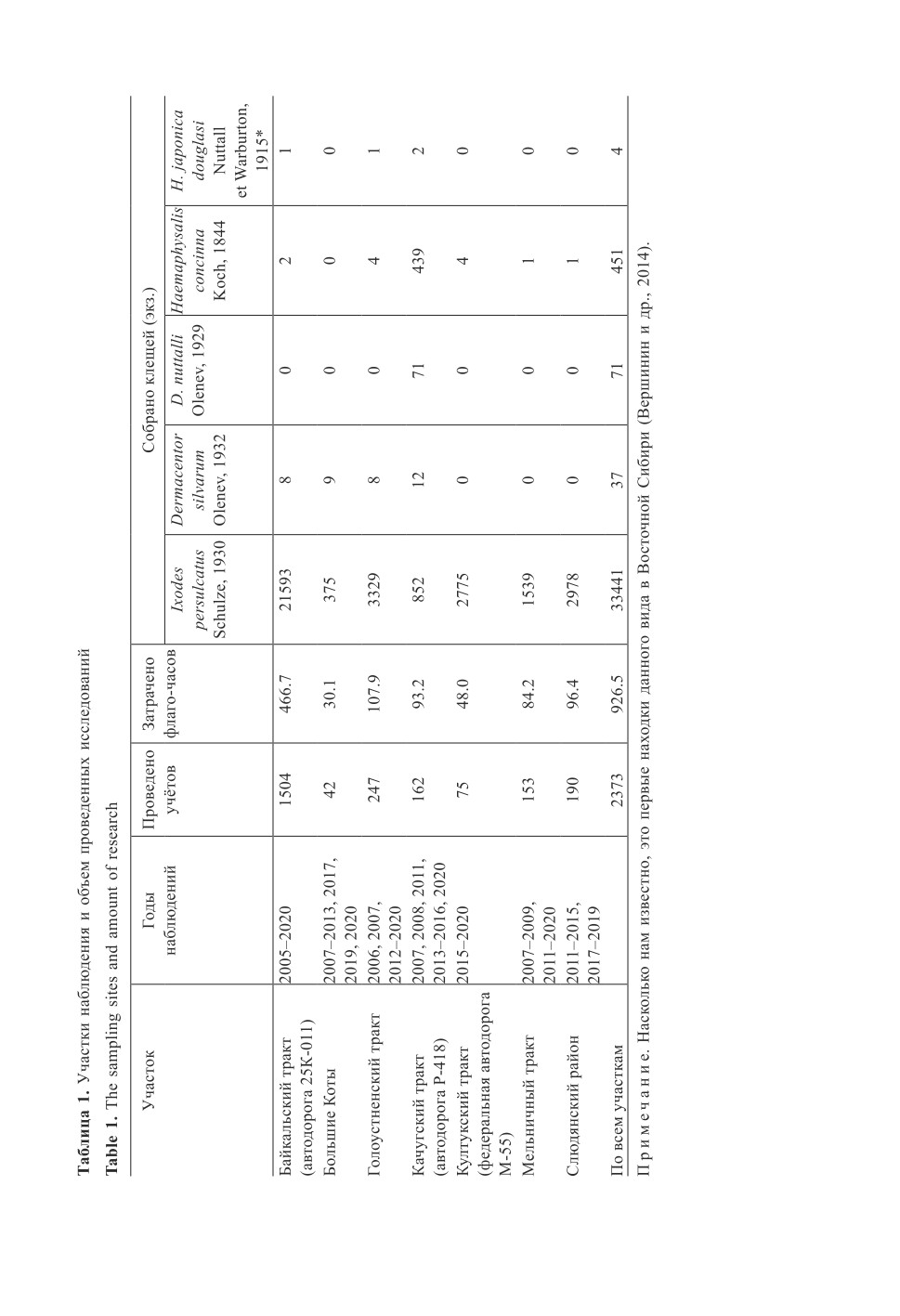
с различной степенью регулярности, было охвачено более 100 точек на семи участках (рис. 1,
табл. 1). Учетные маршруты прокладывали по лесным массивам, вдоль лесных троп и дорог
разной степени использования, просек ЛЭП.
Клещей собирали с конца апреля по начало июля, на движущегося учетчика и стандартный
флаг из фланелевой ткани 60×100 см (Таёжный клещ …, 1985); оценку уровня численности
производили в экземплярах на флаго-час (ф-ч). Среднемноголетние и среднесезонные пока-
затели обилия таёжного клеща рассчитывали по всем проведенным учётам на конкретном
участке за период исследования. Иксодид определяли до вида по морфологическим признакам
(Филиппова, 1977, 1997). Объем работы представлен в табл. 1.
497
Координаты мест сбора материала устанавливали с помощью спутникового навигатора
GPSMAP 76CSx в системе глобального позиционирования. Для построения карты использо-
материала по географическим координатам производили с использованием модульного рас-
ширения xyToPoint программы QGIS 2.18.28.
Собранных членистоногих во влажных бинтах доставляли живыми в лабораторию, где
до исследования хранили в холодильной камере при температуре -20 °. Имаго клещей ана-
лизировали индивидуально, приготавливая из них суспензию на физиологическом растворе
(по 0.5 мл на одного клеща). Наличие антигена (АГ) ВКЭ выявляли иммуноферментным ме-
тодом с помощью набора реагентов ИФА ТС АГ ВКЭ («Микроген», Томск) в соответствии
с инструкцией производителя. В 2012 г. использовали Набор реагентов для выявления анти-
генов вируса клещевого энцефалита ЗАО Биосервис. Учет результатов проводили визуально
и с помощью иммуноферментного анализатора IMARK BioRAD при длине волны 450 нм. Про-
бу считали положительной, если отношение величины её экстинкции к величине экстинкции
нормального контроля было больше или равно 2.1. Выделение вируса проводили на новорож-
дённых беспородных белых мышах (НБМ), по общепринятой методике (Вирусология, 1988).
За животными наблюдали 21 день. Зверьков с клиническими проявлениями нейроинфекции
умерщвляли, руководствуясь «Правилами лабораторной практики в Российской Федерации»
(утверждены Приказом Министерства здравоохранения и социального развития Российской
Федерации № 708н от 23.08.2010), и извлекали головной мозг для дальнейших исследований.
Рисунок 1. Места учётов и сбора иксодовых клещей.
Figure 1. Places of censuring and collecting of Ixodid ticks.
498
С 2013 г., кроме тестирования клещей на наличие АГ ВКЭ в ИФА, случайные выбор-
ки переносчиков с каждого участка исследовали в ПЦР. РНК/ДНК из клещевых и мозговых
суспензий выделяли, используя комплект реагентов «РИБО-преп»; обратную транскрипцию
проводили, используя комплект «РЕВЕРТА-L»; геномный материал ВКЭ и боррелий выявляли
в ПЦР с помощью набора реагентов «АмплиСенс® TBEV, B. burgdorferi s. l., A. phagocyto-
philum, E. chaffeensis/E. muris-FL» (ФБУН ЦНИИ эпидемиологии Роспотребнадзора, Москва)
в режиме реального времени на термоциклере C1000TM Bio-Rad CFX96TM (США).
Информация об инцидентности КЭ и ИКБ на обследуемой территории получена из попол-
няемых авторских баз данных1, созданных на основе карт эпидемиологического расследования
ФБУЗ Центра гигиены и эпидемиологии в Иркутской области.
Статистическую обработку результатов проводили общепринятыми методами вариацион-
ной статистики: критерий Стьюдента (t-критерий), регрессионный и корреляционный анализы,
коэффициент вариации, критерий χ2 (Закс, 1976); за статистически достоверный принимали
уровень значимости P < 0.05. Расчеты проводили с помощью программы Microsoft Excel 2007.
Краткая ландшафтно-климатическая характеристика
обследуемой территории
Южное Прибайкалье находится в природной зоне южной тайги и характеризуется резко
континентальным климатом, а также значительной высотой территории над уровнем моря.
Сложный горный рельеф создает разнообразные условия нагревания и охлаждения земной
поверхности, обусловливает своеобразное распределение атмосферных осадков, ветрового ре-
ландшафтов обследуемой территории. Анализ с помощью геоинформационных систем (ГИС)
показал, что размах колебаний численности иксодид больше зависит от климата и растительно-
го покрова, чем от факторов, связанных с хозяевами (Randolph, 2000). Рассмотрим ландшафтное
разнообразие обследуемых участков.
Байкальский тракт располагается по правому берегу р. Ангары, соединяя г. Иркутск с пос.
Листвянка на оз. Байкал. Голоустненский тракт проходит в юго-восточном направлении от
областного центра до пос. Большое Голоустное на Байкале (рис. 1). Обе дороги пролегают по
таёжным округам предгорий Приморского хребта, где широко представлены подгорные под-
таёжные сосновые комплексы, с подлеском из рододендрона даурского (Ландшафты …, 1977).
Пос. Большие Коты расположен в тайге у подножия Приморского хребта на побережье оз.
Байкал; сухопутных дорог, ведущих к поселку, нет. Климат умеренно- холодный с отрицатель-
ной среднегодовой температурой. В иксодофауне преобладает таёжный клещ, чья численность
невелика (табл. 1). Встречаются и клещи рода Dermacentor. Основой ландшафта являются
склоновые травяные с подлеском из рододендрона даурского остепнённые горнотаёжные со-
сновые комплексы.
Качугский тракт пролегает в северо-восточном направлении от областного центра по тер-
ритории, характеризующейся умеренно тёплым и недостаточно влажным климатом. Расти-
тельный покров представлен сложным сочетанием лесов, степей, лугов и болот. Около поло-
вины площади занимают горнотаёжные комплексы, представленные тёмнохвойными (из кедра
и ели), светлохвойными (из сосны и лиственницы) и мелколиственными (из березы и осины)
лесами. Многие участки значительно нарушены сельскохозяйственной деятельностью. На сты-
ках ландшафтов здесь можно встретить одновременно иксодовых клещей трех родов - Ixodes,
Haemaphysalis и Dermacentor (табл. 1).
1 Свидетельства о государственной регистрации базы данных № 2013620219 и 2013620220; 2013 г.
499
Култукский тракт соединяет Иркутск с оз. Байкал в южном направлении и проходит по
долине рек Иркута и Олхи на Иркутско-Черемховской равнине в зоне горнотаёжных южноси-
бирских (тёмнохвойных, кедрово-лиственичных и горнотаёжных сосновых) лесов с несколькими
группами фаций, характерных для этих геомов (Ландшафты …, 1977).
Мельничный тракт пролегает по подтаёжным биотопам вдоль левого берега р. Ангары
к юго-западу от областного центра. Преимущественные ландшафты - равнинные и днищ кот-
ловин бруснично-разнотравные с кустарниковым подлеском.
Слюдянский район расположен на южном побережье Байкала. Район отличается от дру-
гих большим увлажнением и большей теплообеспеченностью. Перепад высоты в 1800 м обу-
словливает эффект ландшафтной поясности с большим разнообразием ландшафтов, входящих
в горнотаёжные южносибирские и Байкало-Джугджурские комплексы (Ландшафты …, 1977).
В иксодофауне абсолютно доминирует I. persulcatus (табл. 1). Учеты клещей проводили
у подножья хребта Хамар-Дабан, где преобладают следующие ландшафты: равнинные и днищ
котловин бруснично-разнотравные с кустарниковым подлеском; склоновые кедрово-пихтовые
чернично-травяно-зеленомошные; долинные елово-пихтово-крупнотравные; склоновые сосново-
лиственничные бруснично-разнотравные.
По всей обследуемой территории располагается большое количество рекреационных зон
и в тёплый период года проходит множество туристических маршрутов.
РЕЗУЛЬТАТЫ
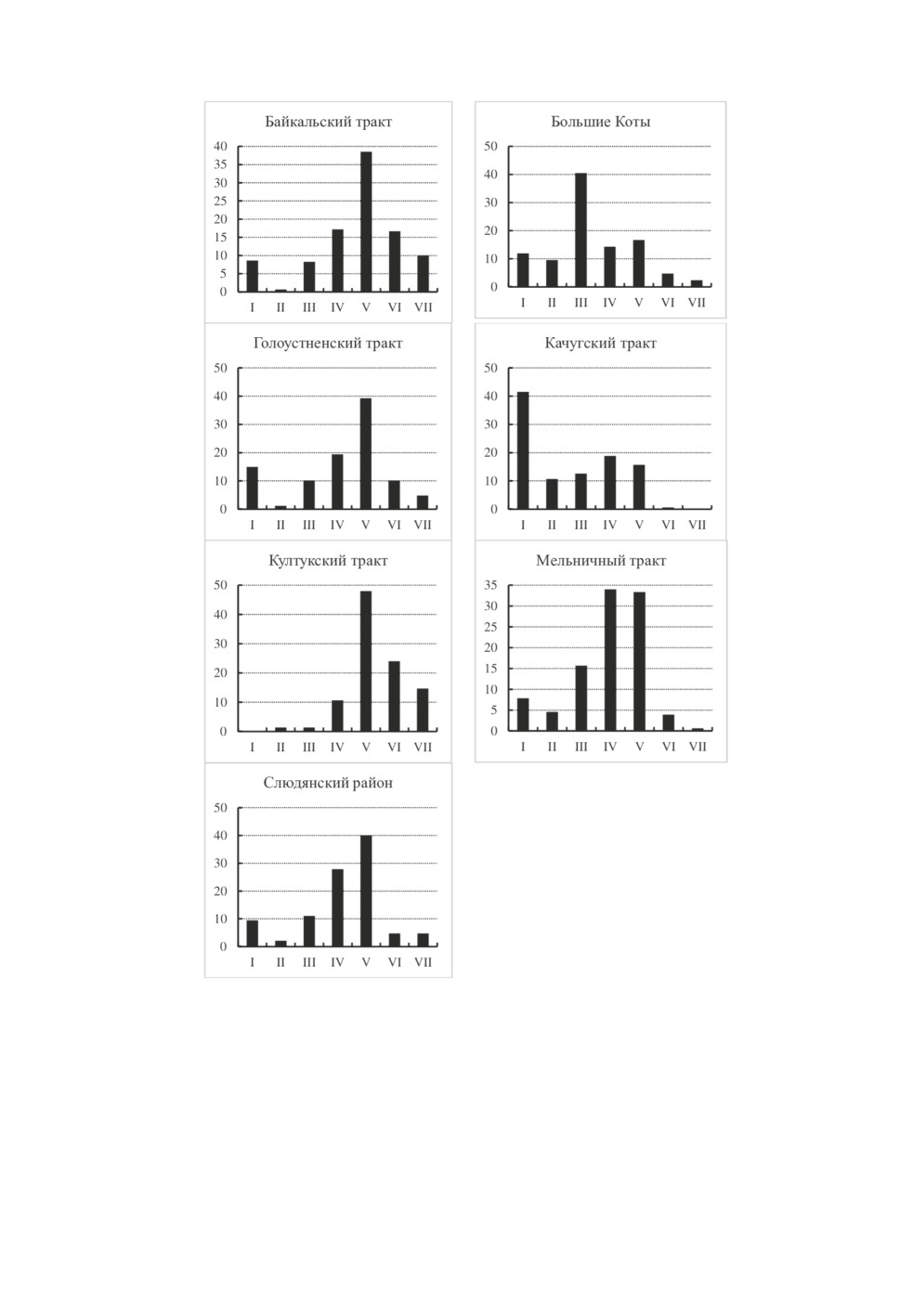
Для получения общего представления о структуре размещения клещей на обсле-
дуемых участках мы, согласно предложенной ранее классификации (Коренберг и
др., 2013, с. 114-115), построили гистограммы, отражающие относительную частоту
(в процентах) показателей различных классов обилия таёжного клеща (от полного их
отсутствия до 100 экз. на ф-ч и более) (рис. 2). Все классы численности представлены
почти на каждом участке, кроме Качугского тракта, где учёты с максимальными по-
казателями не зафиксированы, и Култукского, на котором клещи встречались на всех
пройденных маршрутах. Несмотря на кажущееся сходство некоторых гистограмм
(Байкальский и Голоустненский тракт, Мельничный тракт и Слюдянский район),
сравнение распределений частот показателей обилия на всех участках наблюдения
выявило высокую степень их неоднородности (χ² = 466.9, df = 36. P < 0.001). Обилие
имаго I. persulcatus варьировало по участкам, по годам и в течение сезона активно-
сти. Среднемноголетние показатели относительной численности таёжного клеща на
обследуемой территории, стандартное отклонение, интервалы и величина амплитуды
колебаний численности (коэффициент вариации, CV) приведены в табл. 2. Судя по
минимальной величине CV, достоверно отличающейся от таковой на всех осталь-
ных участках, наиболее стабильно состояние населения таёжного клеща на Култук-
ском тракте. Там же отмечены максимальный показатель средней численности (55.9)
и наибольшая доля учётов с численностью выше 100 экз. на ф-ч (14.7%). Наоборот,
популяции I. persulcatus в окрестностях пос. Б. Коты и по Качугскому тракту наи-
менее стабильны. Высокая достоверность этих различий подтверждается статисти-
ческими методами (табл. 3).
Более или менее продолжительные периоды непрерывных наблюдений для оцен-
ки динамики многолетней среднесезонной численности таёжного клеща имеются
только по Байкальскому (16 лет), Мельничному (14 лет) и Голоустненскому (девять
501
лет) трактам. По этим же участкам имеются сведения о среднемноголетней сезонной
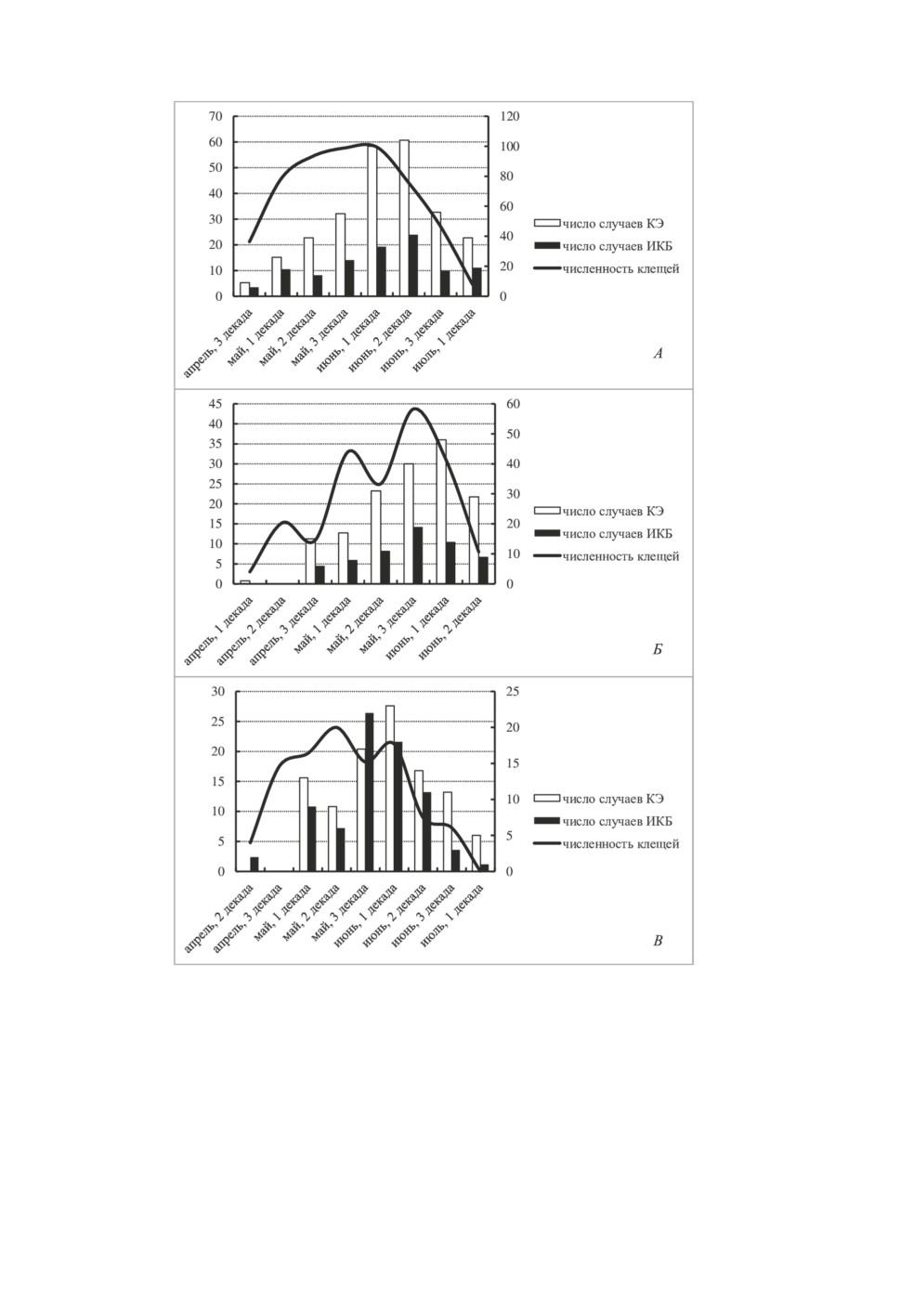
активности переносчика. На рис. 3А показано, что флуктуации численности на Голо-
устненском направлении происходят чаще, чем на двух других, с периодом колебаний
2-3 года. Периодичность циклов численности населения клещей вдоль Байкальского и
Мельничного трактов - 4-5 лет. Очевидная тенденция на повышение уровня обилия
прослеживается лишь на Мельничном тракте (коэффициент регрессии равен 0.271
± 0.1145, df = 13, P < 0.05). Что касается сезонной активности (рис. 3Б), пик её на
Мельничном тракте приходится на более ранние сроки, чем на двух других участках
и наступает уже во второй декаде мая. На Байкальском тракте максимум сезонного
обилия I. persulcatus приходится на первую декаду июня.
Таблица 2. Показатели среднемноголетней численности таёжного клеща
и амплитуда колебаний его численности (CV) на обследуемой территории
Table. 2. The multi-year averages of Ixodes persulcatus abundance and coefficient
of its variation (CV) on the territory under investigation
Средняя
Стандартное
Участок
численность
Интервал
CV
отклонение
(на ф-ч)
Байкальский тракт
43.8 ± 1.16
44.6
392
101.9 ± 1.87
Большие Коты
15.7 ± 3.74
24.2
120
154.5 ± 16.86
Голоустненский тракт
30.8 ± 2.21
34.7
286
112.8 ± 5.08
Качугский тракт
8.9 ± 1.05
13.2
66.2
142.3 ± 8.31
Култукский тракт
55.9 ± 4.04
35.0
147.1
62.6 ± 5.11
Мельничный тракт
19.7 ± 1.61
19.9
126.7
101.4 ± 5.79
Слюдянский район
28.6 ± 2.51
34.6
243.8
121.1 ± 6.21
Таблица 3. Различия амплитуды колебаний численности таёжного клеща
на обследуемой территории
Table. 3. Coefficient of Ixodes persulcatus abundance variation
on the territory under investigation
Сравниваемые участки
Значение t
df
Уровень значимости P
Култукский и Байкальский тракты
7.23
1555
< 0.001
Култукский тракт и Большие Коты
5.22
115
< 0.001
Култукский и Голоустненский тракты
6.98
320
< 0.001
Култукский и Качугский тракты
8.78
232
< 0.001
Култукский и Мельничный тракты
5.02
226
< 0.001
Култукский тракт и Слюдянский район
7.27
263
< 0.001
Б. Коты и Голоустненский тракт
2.37
287
< 0.05
Б. Коты и Качугский тракт
0.33
199
> 0.05
Б. Коты и Мельничный тракт
2.98
193
< 0.01
Б. Коты и Слюдянский район
1.86
230
> 0.05
Качугский и Мельничный тракты
4.63
310
< 0.001
Качугский тракт и Слюдянский район
2.62
347
< 0.01
502
Рисунок 2. Вариационные ряды показателей обилия (на ф-ч) имаго I. persulcatus
на обследуемых участках. По осям X - классы численности клещей: I - клещи отсутствуют,
II - 1-2, III - 3-7, IV - 8-19, V - 20-59, VI - 60-99, VII - 100 и более.
По осям Y - доля учетов (%) с данной численностью.
Figure 2. Variational series of the I. persulcatus imago abundance rates (specimen for 1 flag hour)
on the patches under examination: X-axis - the ticks’ abundance classes: I - lack of ticks,
II - 1-2, III - 3-7, IV - 8-19, V - 20-59, VI - 60-99, VII - 100 and more.
Y-axis - percentage of the censures with that abundance.
503
Рисунок 3. Многолетняя (А) и сезонная (Б) динамика численности таёжного клеща
на территориях, прилегающих к Байкальскому, Голоустненскому и Мельничному трактам.
По осям Y - численность (экз. на ф-ч), по оси X на рис. 3А - годы наблюдений,
на рис. 3Б - декады сезонной активности клещей.
Figure 3. Multi-year (A) and seasonal (B) trends of the taiga tick abundance on the territories
around the Baikalsky, Goloustnensky and Melnichny Highways. Y-axis - abundance rates (speci-
men for 1 flag hour), X-axis - Fig. 3A - years of the monitoring, Fig. 3B - decades of ticks sea-
sonal activity.
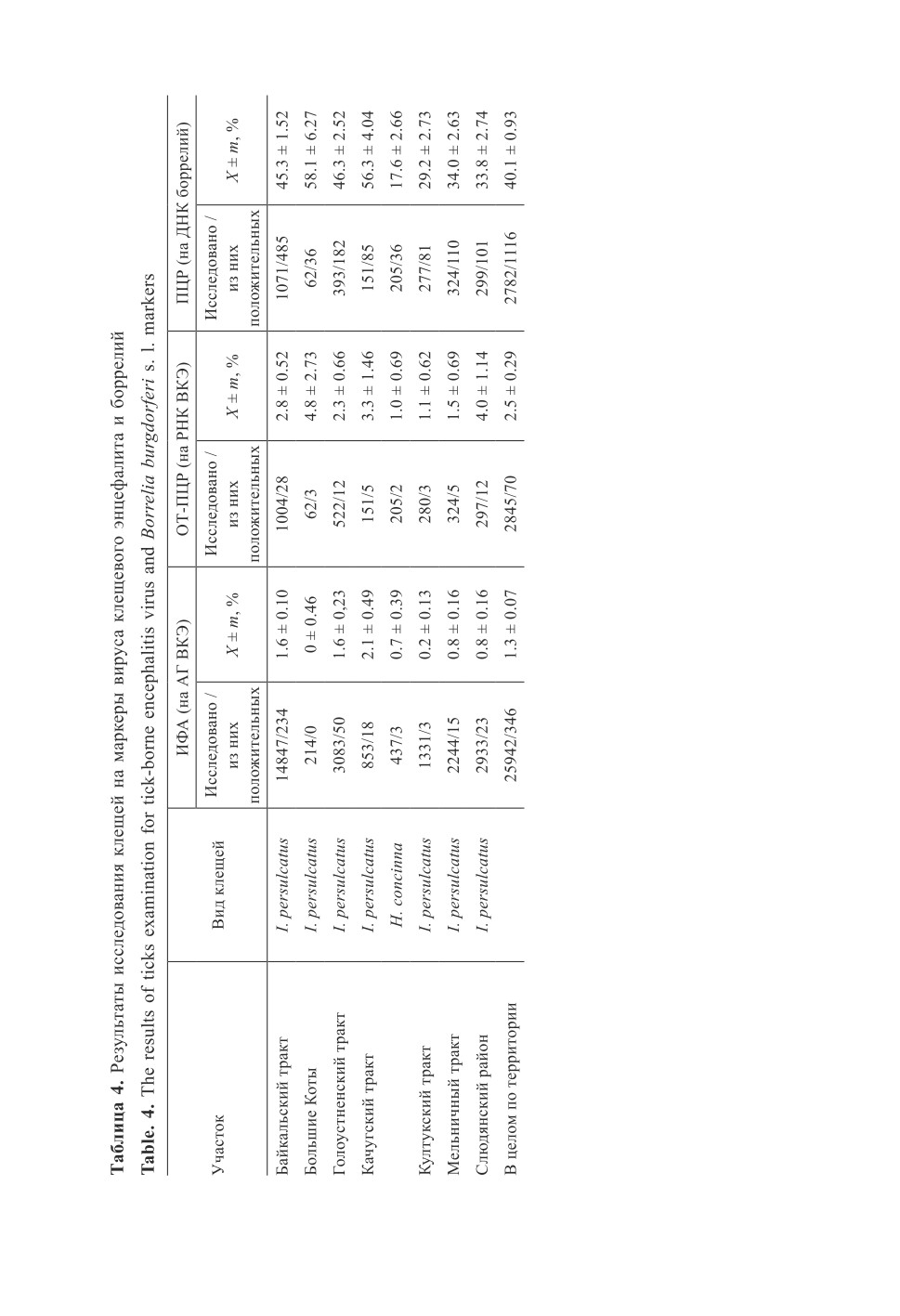
Результаты обнаружения маркеров ВКЭ и ИКБ в исследованных клещах пред-
ставлены в табл. 4. Поскольку на некоторых маршрутах, расположенных вдоль Качуг-
ского тракта, в зонах симпатрии обилие клещей H. concinna сопоставимо с таковым
I. persulcatus (табл. 1) и есть вероятность наличия дополнительного переносчика,
поиск патогенов проводили в обоих видах клещей. Средняя спонтанная заражен-
ность иксодид ВКЭ на обследуемой территории составила 1.3% по результатам ИФА
и 2.5% по результатам ПЦР, но эта разница существенна только по выборке в целом
(t = 3.8, df = 28785, P < 0.001), Байкальскому тракту (t = 2.3, df = 15849, P < 0.05)
и Слюдянскому району (t = 2.8, df = 3228, P < 0.01). Маркеры боррелий в среднем
встречались у 40.1% исследованных клещей, более чем в 30 раз превышая показатель
обнаружения АГ ВКЭ. Следует отметить, что 48.5% клещей с маркерами ВКЭ со-
держали одновременно маркеры боррелий.
504
Имелись значимые различия в обнаружении патогенов в I. persulcatus с отдельных
участков в сравнении со средним показателем по территории: АГ ВКЭ реже обнару-
живали в суспензиях клещей из Б. Котов (t = 2.9, df = 25717, P < 0.01), с Култукского
(t = 3.6, df = 27745, P < 0.001), Мельничного (t = 7.5, df = 26834, P < 0.001) трактов
и из Слюдянского района (t = 3.1, df = 28435, P < 0.01). На Мельничном тракте реже,
чем в среднем по обследованной территории, встречались также клещи с РНК ВКЭ
(t = 2.2, df = 2918, P < 0.05) и маркерами боррелий (t = 4.4, df = 2852, P < 0.001).
Значительно реже среднего показателя по Прибайкалью боррелии встречались в кле-
щах с участков вдоль Култукского тракта и из Слюдянского района (t = 2.8, df = 2899,
P < 0.01 и t = 2.8, df = 2874, P < 0.01 соответственно), а чаще - в переносчиках
из Б. Котов и с Качугского тракта (t = 2.5, df = 2637, P < 0.05 и t = 3.5, df = 2726,
P < 0.001 соответственно). Касательно видовых различий клещей по содержанию
маркеров искомых патогенов, АГ ВКЭ и ДНК боррелий обнаруживали в суспензиях
I. persulcatus гораздо чаще по сравнению с H. concinna (t = 2.3, df = 640, P < 0.05
в случае АГ ВКЭ и t = 8.0, df = 408, P < 0.001 в случае боррелий).
Из суспензий, показавших положительный результат на наличие АГ и/или РНК
ВКЭ, изолировано на биологической модели (беспородные белые мыши) 69 штам-
мов ВКЭ. Успешность изоляции варьировала как по годам, так и по территории,
и в среднем составила 19.6 ± 0.50% от числа суспензий с положительными маркерами
ВКЭ. Доля изолятов с Байкальского тракта и из Слюдянского района была наиболее
высокой (24.5 ± 2.82 и 20.7 ± 7.52%), из клещей с Мельничного тракта за все годы
исследований не удалось выделить ни одного штамма.
Доля жителей Иркутска, заразившихся КЭ и ИКБ на каждом из участков, от со-
вокупного числа пациентов, пострадавших от присасывания клещей на обследуемой
территории, показана на рис. 4. Обращает на себя внимание неравномерность рас-
пространённости случаев заболеваний: 44% заражения КЭ и 35% ИКБ приходится на
местности, прилегающие к Байкальскому тракту. Меньше всего случаев заражений
иркутян имело место в труднодоступных Б. Котах и достаточно отдалённом от област-
ного центра Слюдянском районе. На участках, по которым имелись продолжительные
периоды наблюдений, имелась тенденция к отрицательной связи многолетней динами-
ки инцидентности обеих нозоформ и численности клещей. По Байкальскому тракту
коэффициент ранговой корреляции Спирмена составил - 0.186 для КЭ и - 0.172 для
ИКБ (n = 15 в обоих случаях), по Мельничному - rs = - 0.541 для КЭ и - 0.554 для
ИКБ (n = 13 в обоих случаях). Что касается сезонной динамики, пики нападения
клещей, приведшие к инфицированию КЭ и ИКБ, по территории неодинаковы и, по
большей части, не совпадают с пиком активности клещей на участках, по которым
имеются наблюдения (рис. 5). При этом пик заражения обеими нозоформами имеет
очевидный сдвиг вправо, как минимум, на одну декаду и приходится на спад числен-
ности клещей.
Для установления связи между исследуемыми параметрами мы провели анализ
с помощью ранговой корреляции Спирмена (табл. 5). В отношении КЭ относительная
численность переносчиков коррелировала с вирусофорностью клещей по результатам
ИФА (обнаружение АГ ВКЭ), но не по результатам ПЦР (выявление РНК ВКЭ).
506
Число изолятов ВКЭ было выше при высокой численности клещей, но не зависело
от доли инфицированных клещей. Отсутствовала связь между долей переносчиков
с АГ и РНК ВКЭ. Число случаев КЭ на обследуемой территории коррелировало
с вирусофорностью по результатам ИФА и с успешностью изоляции на теплокровных
животных, но не имело связи с вирусофорностью по результатам ПЦР. Что касает-
ся боррелий, то выявление их маркеров было выше в случае высокой численности
переносчиков, но число заболеваний на обследуемой территории не зависело ни от
численности клещей, ни от их боррелиефорности.
Рисунок 4. Доля больных клещевым энцефалитом и иксодовыми клещевыми боррелиозами
людей, заразившихся на обследуемых участках южного Прибайкалья.
Figure 4. Proportion of patients, infected with tick-borne encephalitis and tick-borne borreliosis
on the territories under examination.
Таблица 5. Установление связи между признаками по коэффициенту
ранговой корреляции Спирмена
Table. 5. Establishing a connection between features by Spearman's rank correlation coefficient
Сравниваемые признаки
rs
n
Уровень
значимости P
Численность клещей / доля клещей с АГ ВКЭ
0.312
67
< 0.05
Численность клещей / доля клещей с РНК ВКЭ
0.144
44
> 0.05
Численность клещей / число случаев КЭ
0.105
57
> 0.05
Численность клещей / число изолятов на мышах
0.534
68
< 0.001
Доля клещей с АГ ВКЭ / доля клещей с РНК ВКЭ
0.026
44
> 0.05
Доля клещей с АГ ВКЭ / число изолятов на мышах
- 0.055
39
> 0.05
Доля клещей с АГ ВКЭ / число случаев КЭ
0.506
53
< 0.001
Доля клещей с РНК ВКЭ / число случаев КЭ
0.259
37
> 0.05
Число случаев КЭ / число изолятов на мышах
0.531
57
< 0.001
Численность клещей / доля клещей с ДНК боррелий
0.345
44
< 0.05
Численность клещей / число случаев ИКБ
0.090
39
> 0.05
Доля клещей с ДНК боррелий / число случаев ИКБ
- 0.137
38
> 0.05
507
Рисунок 5. Сезонная динамика инцидентности заболеваний клещевым энцефалитом
и иксодовыми клещевыми боррелиозами и численности I. persulcatus на территориях,
прилегающих к Байкальскому (А), Голоустненскому (Б) и Мельничному (В) трактам.
По осям Y слева - численность клещей (экз. на ф-ч), справа - число случаев заболеваний;
по оси X - декады сезонной активности клещей.
Figure 5. Seasonal incidence of tick-borne encephalitis and tick-borne borreliosis cases,
and I. persulcatus abundance on the territories around the Baikalsky (А), Goloustnensky (В)
and Melnichny (C) Highways. The main Y-axis- abundance rates (specimen for 1 flag hour);
subsidiary Y-axis - number of cases. X-axis - decades of ticks seasonal activity.
508
ОБСУЖДЕНИЕ
Замечено, что ландшафтное разнообразие даже на небольших по площади участ-
ках территории может значительно влиять на обилие клещей (Václavík et al., 2020).
Daniel с соавт. (2006) на территории Богемии (западная часть Чешской Республики)
выделили девять типов растительности, соотнеся их с различными уровнями встре-
чаемости европейского клеща I. ricinus, разными уровнями контакта с ними людей
и риска нападения. Авторы не смогли связать обилие клещей с каким-то определен-
ным классом лесов и заключили, что ни один из них не может считаться эксклю-
зивно «симптоматичным» в отношении риска заболевания КЭ. Впрочем, отметили,
что ельники практически исключают наличие очага КЭ. Ястребов (2013) утвержда-
ет, что высокий риск заражения характерен для природных очагов, приуроченных
к смешанным лесам разных типов: осиново-пихтовым, сосново-осиново-берёзовым,
хвойно-широколиственным и другим с богатым подлеском и травостоем. На обсле-
дуемой нами территории имеются как разные типы смешанных лесов, так и локации,
занятые почти исключительно берёзой, сосной или елью. Клещи, содержащие маркеры
ВКЭ и боррелий, встречались на участках со всеми типами растительности, однако
численность переносчиков была заметно ниже на тех из них, где имеются остепнён-
ные биотопы (прибрежная полоса Байкала в окрестностях пос. Б. Коты, зоны вдоль
Качугского тракта).
Поскольку трансмиссия «клещевых» патогенов зависит от вектора, для получения
информации об их динамике и распространении важно понимание популяционной
структуры переносчика (Carpi et al., 2016). Václavík с соавт. (2020) обнаружили, что
все патогены, по которым они проводили поиск, чаще встречались в местах с более
высокой плотностью клещей. При изучении природных очагов клещевого боррелиоза
в Прибайкалье (Сунцова, 2004) не выявлено корреляционной связи между показате-
лями многолетней численности клещей в природе и их заражённостью боррелиями,
однако в сезонной динамике максимум заражённости клещей совпадал с пиком числен-
ности переносчиков в природе. В нашем исследовании доля переносчиков, содержа-
щих маркеры ВКЭ и боррелий, в целом коррелировала с их численностью. Однако
риск заболевания клещевой инфекцией обычно связан не просто с численностью
переносчика, а с её динамикой, в том числе сезонной (Randolph, 2000). На участках
с продолжительным периодом непрерывных наблюдений (Байкальский, Голоустнен-
ский и Мельничный тракты) колебания численности происходили с различной часто-
той: от 2-3 до 4-5 лет. Вероятно, это можно объяснить влиянием микроклиматических
факторов и увеличением в этой связи продолжительности жизни одной генерации
клещей (Коротков, 1998; Osipova et al., 2017). В целом по территории число случаев
КЭ коррелировало с обилием переносчика, чего нельзя сказать о числе случаев ИКБ.
Что касается соответствия сезонной динамики численности клещей и динамики приса-
сывания к людям инфицированных переносчиков, в нашем исследовании пик зараже-
ния людей происходил как минимум на декаду позже пиковой активности клещей на
конкретном участке. Алексеев (2007) объяснял такой сдвиг вероятным возрастанием
к концу сезона числа особей с высокими титрами вируса.
509
Спонтанная заражённость иксодовых клещей патогенами вирусной и бактери-
альной природы неодинакова в разных частях их ареала (Коренберг и др., 2013),
в том числе, как показали более ранние наши исследования (Melnikova et al., 2019),
в различных районах Прибайкалья. В настоящей работе подтверждены значимые рас-
хождения в зараженности клещей как ВКЭ, так и боррелиями по обследованным
направлениям. Опираясь на эти результаты, можно говорить о риске заражения дан-
ными патогенами в той или иной местности. Кроме того, в зонах симпатрии патогены
могут обнаруживаться не только у основного переносчика, но и у других видов кле-
щей (Katargina et al., 2013). На некоторых лесных участках территории, прилегающей
к Качугскому тракту, ареалы I. persulcatus и H. concinna перекрываются, а показатели
численности этих видов сопоставимы. Результаты нашего анализа показывают, что
при прочих равных условиях (благоприятная среда обитания, схожие кормовая база
и показатели численности популяций двух видов клещей) таёжный клещ, несомненно,
является основным переносчиком искомых патогенов, поскольку соответствующие
маркеры в его суспензиях обнаруживали значительно чаще.
Наиболее достоверным подтверждением устойчивой циркуляции патогена на тер-
ритории является получение изолята в биопробе с последующей его идентификацией
(Ястребов, 2013). Гипотетически, риск заражения человека КЭ на конкретной тер-
ритории находится в прямой зависимости от количества изолятов вируса, которые
удается получить на теплокровных животных. В доступной литературе мы не нашли
исследований, посвященных этому вопросу. В нашей работе показан высокий уровень
корреляции между числом заболеваний КЭ и числом изолятов в целом по южному
Прибайкалью, хотя на отдельных участках (Слюдянский район) доля патогенных для
НБМ суспензий довольно высока, а доля заразившихся КЭ минимальна. Вероятно,
в данном случае вступают в действие другие факторы. Уровень заражаемости при-
родноочаговыми инфекциями - это результат взаимодействия двух величин: лоймопо-
тенциала очага и интенсивности контакта людей с природными очагами (Коренберг
и др., 2013). Одна из особенностей современной эпидемиологии КЭ состоит в том,
что большинство встреч с клещами происходит не в лесу и не на местности, грани-
чащей с лесом, а в рекреационных зонах (Zeman, Benes, 2013), и количество случаев
заражения напрямую зависит от плотности населения (коэффициент корреляции 0.97)
(Дружинина и др., 2002). Ключевым фактором, контролирующим контакт с пере-
носчиком и инфицирование, является поведение человека (Estrada-Peña, de la Fuente,
2014; Lambin et al., 2010). Как результат комплексного влияния всех исследованных
в настоящей работе факторов (высокие показатели численности, вирусофорности
и боррелиефорности переносчика, большое количество изолятов ВКЭ на лаборатор-
ных животных), наибольший риск заражения КЭ и ИКБ на обследованной территории
связан с посещением Байкальского тракта. Немаловажную роль играет также значи-
тельная плотность жилых поселений и рекреационных зон, расположенных вдоль
этой трассы. Именно с этого направления имеет место около 30% всех обращений
по поводу присасывания клещей (Козлова, 2008), происходит не меньше трети за-
ражений КЭ и четверти ИКБ (Мельникова, Андаев, 2017). Уменьшение воздействия
любого из вышеперечисленных факторов снижает вероятность заражения на кон-
кретной территории.
510
ВЫВОДЫ
1. Сравнение распределений частот показателей обилия I. persulcatus по критерию
χ² на обследуемых участках выявило высокую степень их неоднородности. Показаны
значимые различия среднемноголетних показателей относительной численности та-
ёжного клеща на этих участках, а также многолетней и сезонной динамики.
2. Средняя спонтанная зараженность иксодид ВКЭ на обследуемой территории
составила 1.3% по результатам ИФА и 2.5% по результатам ПЦР, маркеры боррелий
выявлены у 40.1% исследованных клещей. При этом имелись значимые различия
в обнаружении патогенов в I. persulcatus с отдельных участков в сравнении со сред-
ним показателем по территории. В суспензиях I. persulcatus АГ ВКЭ и ДНК боррелий
обнаруживали гораздо чаще, чем в суспензиях H. concinna.
3. Выявлены существенные различия между обследованными участками южного
Прибайкалья по патогенности переносчиков, содержащих маркеры ВКЭ, для лабо-
раторных животных.
4. Корреляционный анализ показал наличие прямой связи между численностью
переносчиков и обнаружением АГ ВКЭ, но не РНК ВКЭ. Число изолятов ВКЭ было
выше при высокой численности клещей, но не зависело от доли инфицированных
клещей. Отсутствовала связь между долей переносчиков с АГ и РНК ВКЭ. Число
случаев КЭ на обследуемой территории коррелировало с вирусофорностью по ре-
зультатам ИФА и с успешностью изоляции на теплокровных животных, но не имело
связи с вирусофорностью по результатам ПЦР. Что касается боррелий, то выявление
их маркеров было выше в случае высокой численности переносчиков, но число за-
болеваний на обследуемой территории не зависело ни от численности клещей, ни от
их боррелиефорности.
5. На основании совокупности исследованных параметров определён участок наи-
большего риска заражения КЭ и ИКБ.
БЛАГОДАРНОСТИ
Авторы выражают признательность бывшим и настоящим сотрудникам ФКУЗ
Иркутский научно-исследовательский противочумный институт Роспотребнадзора
Р.А. Бондарюк, А.Н. Бондарюку, М.А. Борзенко, Т.И. Борисовой, С.А. Борисову,
Д.Б. Вержуцкому, П.Е. Вершинину, М.О. Гориной, И.М. Морозову, А.Я. Никитину,
Н.Н. Пивень, А.В. Севостьяновой, Е.А. Сидоровой, А.В. Холину и др., в разные годы
принимавшим участие в сборе клещей.
СПИСОК ЛИТЕРАТУРЫ
Алексеев А.Н. 2007. Современное состояние знаний о переносчиках клещевого энцефалита. Вопросы
вирусологии 5: 21-26. [Alekseev A.N. 2007. Modern knowledge about tick-borne encephalitis vectors.
Problems of virology 5: 21-26. (In Russian)].
Вершинин Е.А., Мельникова О.В., Морозов И.М. 2014. Клещи рода Haemaphysalis в южной части При-
байкалья. Известия Иркутского государственного университета. Серия «Биология. Экология» 8:
92-95. [Vershinin E.A., Меl’nikova O.V., Morozov I.M. 2014. Haemaphysalis ticks in the southern part
of Pribaikalie. The Bulletin of Irkutsk State University. Series “Biology. Ecology” 8: 92-95. (In Russian)]
Вирусология. Методы. 1988. Пер. с англ. Под ред. Б. Мейхи. М., Мир, 344 с. (Virology. A practical approach.
1985. Ed. by B.W.J. Mahy. Oxford, Washington DC, IRL Press).
511
Данчинова Г.А., Хаснатинов М.А., Сунцова О.В. и др. 2004. Переносчики возбудителей трансмиссивных
клещевых инфекций на юге Восточной Сибири и севере Монголии. Бюллетень ВСНЦ СО РАМН
1(3): 107-112. [Danchinova G.A., Khasnatinov M.A., Suntsova O.V. et al. 2004. Vectors of transmissible
tick-borne infections in the South of East Siberia and North of Mongolia. The Bulletin of East Siberia
branch of Russian Academy of Sciences 1(3): 107-112. (In Russian)]
Дружинина Т.А., Ющенко Г.В., Мелюк С.А. и др. 2002. Клещевой боррелиоз в Ярославской области.
Медицинская паразитология и паразитарные болезни 2: 9-12. [Druzhinina T.A., Yushchenko G.V.,
Melyuk S.A. et al. 2002. Tick-borne borreliosis in Yaroslavl region. Medical parasitology and parasitic
diseases 2: 9-12. (In Russian)]
Закс Л. 1976. Статистическое оценивание. М., Статистика, 598 с. [Zaks L. 1976. The statistical estimation.
Moscow, 598 pp. (In Russian)]
Козлова И.В. 2008. Научное обоснование и пути совершенствования экстренной диагностики и профи-
лактики трансмиссивных клещевых инфекций в условиях сочетанности природных очагов. Дис. …
докт. мед. наук. Иркутск, 307 с. [Kozlova I.V. 2008. Nauchnoe obosnovanie i puti sovershenstvovaniya
ekstrennoj diagnostiki i profilaktiki transmissivnyh kleshchevyh infekcij v usloviyah sochetannosti prirodnyh
ochagov. Thesis for Dr of Science Degree. Irkutsk, 307 pp. (In Russian)]
Коренберг Э.И. 1976. Популяция и группа популяций у Ixodes persulcatus Р. Sch. Медицинская паразитоло-
гия и паразитарные болезни 45 (3): 297-303. [Korenberg E.I. 1976. Ixodes persulcatus Р. Sch. popula-
tion and group of populations. Medical parasitology and parasitic diseases 45 (3): 297-303. (In Russian)]
Коренберг Э.И., Помелова В.Г., Осин Н.С. 2013. Природноочаговые инфекции, передающиеся иксодовыми
клещами. М., 463 с. [Korenberg E.I., Pomelova V.G., Osin N.S. 2013. Infections with Natural Focality
transmitted by Ixodid ticks. Moscow, 463 p. (In Russian)]
Коротков Ю.С. 1998. Циклические процессы в динамике численности таёжного клеща и их связь с по-
годными и климатическими условиями. Паразитология 32 (1): 21-31. [Korotkov Yu.S. 1998. Cikli-
cheskie processy v dinamike chislennosti tayozhnogo kleshcha i ih svyaz' s pogodnymi i klimaticheskimi
usloviyami. Parasitology 32 (1): 21-31. (In Russian)]
Ландшафты юга Восточной Сибири (карта масштаба 1:1 500 000). 1977. Ред. Михеев В.С., Ряшин В.А. М.,
ГУГК. [South of East Siberia landscape (map on a scale of 1:1,500000). 1977. Ed.: Miheev V.S., Ryashin
V.A. Moscow, GUGK. (In Russian)]
Мельникова О.В., Андаев Е.И. 2017. Эколого-эпидемиологические особенности клещевого энцефалита и
иксодовых клещевых боррелиозов в г. Иркутске. Эпидемиология и вакцинопрофилактика 6 (97):
20-26. [Mel'nikova O.V., Andaev E.I. 2017. Ecological and Epidemiological Peculiarities of the Tick-
Borne Encephalitis and Ixodid Tick-Borne Borreliosis in Irkutsk. Epidemiology and Vaccine Prevention
6 (97): 20-26. (In Russian)]
Сунцова О.В. 2004. Эколого-паразитологическая характеристика природных очагов клещевого боррелиоза
в Прибайкалье. Автореф. дис. … канд. биол. наук. Иркутск, 10 с. [Suntsova O.V. 2004. Ekologo-
parazitologicheskaya harakteristika prirodnyh ochagov kleshchevogo borrelioza v Pribajkal'e. Thesis for
PhD Degree. Irkutsk, 10 pp. (In Russian)]
Таежный клещ Ixodes persulcatus Schulze (Acarina, Ixodidae): морфология, систематика, экология, меди-
цинское значение. 1985. Л., Наука, 416 с. [Taiga tick Ixodes persulcatus Schulze (Acarina, Ixodidae):
Morphology, Systematics, Ecology, Medical importance. 1985. Leningrad, 416 pp. (In Russian)]
Филиппова Н.А. 1977. Иксодовые клещи подсем. Ixodinae (Фауна СССР. Паукообразные; IV (4)). Л., Наука,
396 с. [Filippova N.A. 1977. Ixodid ticks of subfamily Ixodinae (Fauna of USSR. Arachnoidea IV (4).
Leningrad “Nauka”, 396 pp. (In Russian)]
Филиппова Н.А. 1997. Иксодовые клещи подсем. Ambliominae. (Фауна России и сопредельных стран. Пау-
кообразные; IV (5)). СПб., Наука, 436 с. [Filippova N.A. 1997. Ixodid ticks of subfamily Ambliominae.
(Fauna of Russia and neighboring countries. Arachnoidea IV (5). St. Petersburg, Nauka Publishing house,
436 pp. (In Russian)]
Якименко В.В., Малькова М.Г., Тюлько Ж.С. и др. 2019. Трансмиссивные вирусные инфекции Западной
Сибири (региональные аспекты эпидемиологии, экологии возбудителей и вопросы микроэволюции).
Омск: Издательский центр КАН, 312 с. [Yakimenko V.V., Mal'kova M.G., Tyul'ko Zh.S. et al. 2019.
Transmissivnye virusnye infekcii Zapadnoj Sibiri (regional'nye aspekty epidemiologii, ekologii vozbuditelej
i voprosy mikroevolyucii). Omsk, 312 pp. (In Russian)]
Ястребов В.К. 2013. Эпидемиологическая оценка очагов клещевого энцефалита для обоснования рацио-
нальных мер профилактики. Диагностика и профилактика инфекционных болезней: Материалы
научно-практической конференции. Новосибирск: Издательство «АРЕАЛ»: 30-31. [Yastrebov V.K.
512
2013. Epidemiologicheskaya ocenka ochagov kleshchevogo encefalita dlya obosnovaniya racional'nyh mer
profilaktiki. Diagnostika i profilaktika infekcionnyh boleznej: Materialy nauchno-prakticheskoj konferencii.
Novosibirsk: 30-31. (In Russian)]
Carpi G., Kitchen A., Kim H.L. et al. 2016. Mitogenomes reveal diversity of the European Lyme borreliosis
vector Ixodes ricinus in Italy. Mol. Phylogenet. Evol. 101:194-202. doi: 10.1016/j.ympev.2016.05.009
Daniel M., Zitek K., Danielova V. et al. 2006. Risk assessment and prediction of Ixodes ricinus tick questing
activity and human tick-borne encephalitis infection in space and time in the Czech Republic. International
Journal of Medical Microbiology 296 (1): 41-47.
Estrada-Peña A., de la Fuente J. 2014. The ecology of ticks and epidemiology of tick-borne viral diseases. An-
tiviral Res. 108C: 104-128. doi: 10.1016/j.antiviral.2014.05.016
Katargina O., Russakova S., Geller J. et al. 2013. Detection and characterization of tick-borne encephalitis virus
in Baltic countries and eastern Poland. PLoS One 8 (5): e61374. doi: 10.1371/journal.pone.0061374
Lambin E.F., Tran A., Vanwambeke S.O. et al. 2010. Pathogenic landscapes: interactions be-
tween land, people, disease vectors and their animal hosts. Int. J. Health. Geogr. 9 (54).
Melnikova O.V., Adelshin R.V., Trushina Yu.N. et al. 2019. Identification of the Spectrum of Pathogens in Ixodid
Ticks from Natural Co-Infection Foci of the Baikal Region. Entomological Review 99 (2): 262-273. DOI:
10.1134/S001387381902012X
Osipova T.N., Grigoryeva L.A., Samoylova E.P. et al. 2017. The influence of Meteorological Factors on the
Activity of Adult Taiga Ticks (Ixodes persulcatus Sch., Ixodinae) in St. Petersburg and Its Environs En-
tomological Review 97 (4): 554-563. DOI: 10.1134/S0013873817040169
Randolph S.E. 2000. Ticks and tick-borne disease systems in space and from space. Adv Parasitol. 47: 217-243.
Václavík T., Balážová A., Baláž V. et al. 2020. Landscape epidemiology of neglected tick-borne pathogens in
central Europe Transbound Emerg Dis. doi: 10.1111/tbed.13845. Online ahead of print.
Zeman P., Benes C. 2013. Spatial distribution of a population at risk: an important factor for understanding the
recent rise in tick-borne diseases (Lyme borreliosis and tick-borne encephalitis in the Czech Republic).
Ticks Tick Borne Dis. 4 (6): 522-30. doi: 10.1016/j.ttbdis.2013.07.003
THE ACTIVITY OF ADULT TAIGA TICKS
(IXODES PERSULCATUS SCHULZE, 1930)
HEMIPOPULATIONS IN COMBINED TICK-BORNE ENCEPHALITIS
AND TICK-BORNE BORRELIOSES NATURAL FOCI
OF SOUTHERN BAIKAL REGION
O. V. Mel'nikova, E. A. Vershinin, V. M. Korzun, Yu. A. Verzhutskaya,
N. V. Yakovchits, R. V. Adel'shin, Yu. N. Trushina, K. V. Lopatovskaya, E. I. Andaev
Keywords: taiga tick Ixodes persulcatus, abundance, tick-borne encephalitis virus
(TBEV), Borrelia, virus prevalence, Borrelia prevalence, TBEV isolates, Baikal Region
SUMMARY
The results of long-term (2005-2020) monitoring of combined tick-borne encephalitis
(TBE) and tick-borne borrelioses (TBB) natural foci in several sites of southern part of
Baikal region are presented. The studied territory showed high degree of heterogeneity of
adult Ixodes persulcatus population density and their infection with TBEV and borrelians.
Significant difference between the studied sites regarding pathogenicity of vectors with
positive TBEV - ELISA results for laboratory mice has been revealed. Positive correlation
between vector abundance and TBEV antigen prevalence and the number of murine viral
isolates has been shown. The incidence of TBE on the studied territory also correlated with
ELISA results and success of isolation on worm-blooded animals. The number of Borrelia
PCR positive results was higher if the ticks abundance was high, but TBB incidence cor-
related neither with ticks abundance, no with their Borrelia infection rates.
513