ПАРАЗИТОЛОГИЯ, 2022, том 56, № 1, с. 35-47.
УДК 595.77
ПЛОТНОСТЬ ОКРЫЛЕННЫХ ФОРМ ОЛЕНЬЕЙ КРОВОСОСКИ
LIPOPTENA CERVI (LINNAEUS, 1758) В РАЗЛИЧНЫХ БИОТОПАХ
И ИНТЕНСИВНОСТЬ ЗАРАЖЕНИЯ ОСНОВНОГО ХОЗЯИНА
НА ТЕРРИТОРИИ ЛЕНИНГРАДСКОЙ ОБЛАСТИ
© 2022 г. Н. В. Седихинa,b,*, А. В. Дмитрюковc
aЗоологический институт РАН,
Университетская наб., д. 1, Санкт-Петербург, 199034 Россия
bСанкт-Петербургский государственный лесотехнический университет им. С.М. Кирова,
Институтский пер., д. 5, Санкт-Петербург, 194021 Россия
cСанкт-Петербургский государственный университет,
Университетская наб., д. 7-9, Санкт-Петербург, 199034 Россия
*e-mail: sedoi1995@mail.ru; Nickolai.Sedikhin@zin.ru
Поступила в редакцию 29.11.2021 г.
После доработки 12.01.2022 г.
Принята к публикации 13.01.2022 г.
Приведены результаты исследований по определению плотности и распространению окры-
ленных форм оленьей кровососки Lipoptena cervi (Linnaeus, 1758) в различных биотопических
условиях на территории Ленинградской области, а также информация о зараженности основных
прокормителей - европейских лосей Alces alces (Linnaeus, 1758). Определение численности
окрыленных форм было проведено методом маршрутного учета с привязкой к биотопам. Про-
должительность периода лёта составляет более 91 дня (с 18 июля по 17 октября). Массовый
лёт начинается с 8-10 августа и продолжается до 18-20 сентября. Плотность кровососок >
50 экз./10 мин регистрировалась в период с 31 августа по 16 сентября. Основными биото-
пами, в которых наблюдалось значительное число окрыленных форм, являлись: смешанные
ельники черничники с наличием сфагнума в ЖНП (max = 35 экз./10 мин); зарастающие мо-
лодняками естественные поляны, перелески (max = 81.4 экз./10 мин) и линейные объекты
(ЛЭП, Газо/Нефтепроводы) (max = 31.1 экз./10 мин); сфагновые и увлажненные разнотравные
сосняки (max = 83.3 экз./10 мин); окраины верховых и переходных болот (max = 120.9 экз./
10 мин); богатые разнотравные и увлажненные березняки (max = 155 экз./10 мин); молодняки,
возобновляемые после сплошных и постепенных рубок (max = 213.3 экз./10 мин); осинники
кисличники с высоким естественным отпадом осины (max = 264.2 экз./10 мин). Впервые по-
лучены количественные данные о численности паразитирующих особей оленьей кровососки
на лосях в осенне-зимний период для Ленинградской области, которые свидетельствуют об
интенсивном заражении различных половозрастных групп: минимальное значение 17 экз./дм2
(взрослая самка, 26.12), максимальное значение 455 экз./дм2 (полуторагодовалый самец, 21.11).
Ключевые слова: оленья кровососка, Lipoptena cervi, плотность, зараженность, европей-
ский лось, Alces alces
DOI: 10.31857/S0031184722010021
35
Оленья кровососка Lipoptena cervi (Linnaeus, 1758) является кровососущим обли-
гатным моноксенным паразитом оленевых (Cervidae). Как представитель семейства
Hippoboscidae оленья кровососка характеризуется живорождением - и яйцо, и личинка
до последней стадии перед превращением в куколку развиваются в теле имаго. По-
сле отрождения в шерсти хозяина предкуколка опадает на землю, а ее покровы за-
твердевают (Балашов, 1996, 2009). Куколка, находящаяся внутри пупария, полностью
неподвижна, приспособлена к перенесению неблагоприятных климатических условий
зимы (Kaunisto et al., 2015; Бахтушкина, 2018). В условиях Северо-Запада России
окрыленные формы имаго активно нападают на людей и основных прокормителей
- европейского лося Alces alces (Linnaeus, 1758) и европейскую косулю Capreolus
capreolus (Linnaeus, 1758). По литературным данным массовое появление и, соот-
ветственно, нападение мух начинаются со второй половины августа и продолжаются
до середины октября (Попов, 1965). Насекомое подстерегает потенциальную жертву
в месте своего вылупления, однако может почувствовать движущийся объект на рас-
стоянии и подлететь к нему. В зависимости от условий окружающей среды мухи мо-
гут подлетать к потенциальной жертве с расстояния до 50 м. При низкой температуре
данное расстояние уменьшается до 15 м (Иванов, 1974, 1975; Буракова, 2002). Попав
на тело прокормителя, представители обоих полов сбрасывают крылья и начинают
питаться. В течение первых трех недель, согласно данным Бураковой (2002), проис-
ходит «созревание» мух - «темнеет окраска покровов, голова втягивается в плечевые
выросты груди, атрофируются крыловые мышцы, увеличивается размер брюшка, про-
является половой диморфизм и наблюдается половая активность». Самка воспроизво-
дит предкуколок на протяжении всего периода паразитирования. Жизнь паразитов на
хозяине продолжается, по разным оценкам, до марта следующего года (Попов, 1965).
Паразитируя, кровососки ежедневно потребляют до 0.006 г крови, доставляют бес-
покойство, дискомфорт, а при большой зараженности и наносят весомый ущерб орга-
низму хозяина (Иванов, 1974; Madslien et al., 2011). В течение последних десятилетий
в Финляндии и Норвегии наблюдается увеличение численности популяции оленьей
кровососки (Kaunisto et al., 2011; Meier et al., 2014), связанное, по всей вероятности,
с высокой численностью прокормителей. Поэтому возникла опасность широкого рас-
пространения и массового «освоения» кровосоской более северных регионов и других
хозяев, в частности Rangifer tarandus (Paakkonen, 2012).
Исследования численности окрыленных форм оленьей кровососки на Северо-
Западе России ранее практически не проводились. Чаще всего в публикациях ука-
зывались лишь усредненные или единичные значения количественных параметров
лётной активности. Из отечественных материалов по L. cervi наиболее известны ра-
боты Попова (1965) и Балашова (1996) в Ленинградской области, Иванова (1974,
1975) в Республике Беларусь и Варнакова (1977) в Вологодской области. Объективных
сведений о зараженности основных хозяев в отечественной литературе для данного
региона авторами выявлено не было.
В цели настоящего исследования входило: установление сроков, динамики и ко-
личественных характеристик лётной активности L. cervi; выявление максимальных и
минимальных значений плотности для распространенных лесорастительных форма-
ций южной и средней тайги; выявление основных биотопов Ленинградской области,
в которых наблюдается высокая плотность окрыленных форм паразита; определение
36
количественных параметров зараженности лосей оленьей кровосоской на территории
исследуемого региона. Для достижения целей были поставлены следующие задачи:
получение количественных данных о лётной активности и плотности оленьей кро-
вососки методом маршрутного учета с привязкой к различным биотопам; получение
образцов шкур основного прокормителя - европейского лося - и последующая их об-
работка; сравнение полученных расчетных результатов с предыдущими изысканиями
по данной тематике.
Актуальность данных исследований обусловлена малочисленностью информации
о распространении и состоянии популяции оленьей кровососки в исследуемом ре-
гионе. Эта муха часто нападает на человека (Иванов, 1975; Kaunisto et al., 2011)
и, как переносчик возбудителей болезни Лайма и различных дерматитов (Буракова,
2002), может быть опасна для людей, отдыхающих и работающих в лесу. Изучение ее
биотопического распределения и динамики лётной активности способствует получе-
нию информации для разработки рекомендаций по предупреждению взаимодействия
с ней и по защите популяций ценных охотхозяйственных видов копытных, поскольку
сильная зараженность, вероятно, может сказываться на физиологическом состоянии
единичных особей, а в совокупности - на продуктивности и распространении по-
пуляции.
МАТЕРИАЛ И МЕТОДИКА
Пешие маршруты
Ориентируясь на схожие проведенные Балашовым (1996) исследования, авторами были
намечены и пройдены пешие маршруты в различных районах Ленинградской области с це-
лью сбора всех нападающих на человека лётных форм L. cervi и в дальнейшем определе-
ния численности и плотности оленьей кровососки в пересеченных учетчиками биотопах.
Под «биотопом» в данной работе подразумевается участок геопространства, однородный по
условиям обитания для зооценоза и занятый однородной растительностью (фитоценозом).
Для закладки трансект осуществлялся обзор спутниковых карт в программах SASPlanet и
Google Earth Pro. Главный принцип рекогносцировки территории, пригодной для организа-
ции и закладки маршрута, - при наименьшей протяженности маршрута обеспечить прохожде-
ние максимального количества биотопов. Сохранённый вычерченный маршрут в расширении
kmz экспортировался в навигатор Garmin через приложение BaseCamp. Задача учетчика со-
стояла в прохождении маршрута вдоль трансекты, при этом необходимо было дать характе-
ристики всем выявленным биотопам, собирая активно нападающих на человека кровососок
в отдельную для каждого биотопа пробирку с 75 % спиртовым фиксирующим раствором. Марш-
руты были заложены в местах потенциального присутствия прокормителеи.
Выделялись следующие категории биотопов: поля, с уточнением их текущего состояния
(сенокос, пашня, луг, пастбище); нелесохозяиственные молодняки (зарастающие поля с/х назна-
чения, естественные поляны); вырубки (лесохозяйственные молодняки), с уточнением возраста
(свежая (текущего года), менее 5 лет, 5-10 лет, 10-20 лет); погибшие насаждения (бурелом,
ветровал, гарь, затопление, очаг вредителей); болота (верховые, переходные, низинные); линии
электропередач, газо/нефтепроводы и лесные территории (лес) - участки, занятые средневоз-
растными, спелыми и перестойными древостоями.
По преобладающей породе (порода, составляющая наибольшую часть верхнего яруса дре-
востоя по запасу) визуально выделялись следующие типы лесов: сосняки, ельники, березняки,
37
осинники, черноольшаники, сероольшаники, ивняки. Древостой каждого типа леса по преоб-
ладающей породе описывался согласно следующей последовательности:
1. Смешанный (в верхнем ярусе разные породы деревьев) или чистый (в верхнем ярусе
все деревья одной породы);
2. Сложный (многоярусный - кроме первого яруса, есть второй (нижний) ярус, который по
высоте на 20% ниже первого) или простои (одноярусный);
3. Разновозрастный (двухъярусный - в каждом ярусе присутствует преобладаю-
щая порода верхнего яруса или одноярусный с множеством разных по диаметру ство-
ла на высоте груди деревьев преобладающей породы) или одновозрастный (одноярусный
с множеством одинаковых по диаметру на высоте груди деревьев преобладающей породы или
двухъярусный, - только в верхнем ярусе присутствовали деревья преобладающей породы, ко-
торые визуально имели схожие диаметры стола на высоте груди);
4. Тип леса по живому напочвенному покрову (ЖНП): кисличник, брусничник, чернич-
ник, беломошник, сфагновый, травяно-сфагновый, сфагново-кустарничковый, приручеиный и
разнотравный. Типы леса по видам-индикаторам травянистой растительности в значительной
степени варьируют, поэтому тип леса «разнотравный» обозначает любой тип леса, где в ЖНП
преобладают различные виды травянистой растительности;
5. Дополнительные породы по ярусам для смешанных древостоев.
Таким образом для каждого биотопа категории «лес» название выглядело по примеру:
«смешанный сложный разновозрастный ельник-кисличник с примесью осины и березы» или
«чистый простой одновозрастный сосняк-беломошник».
Каждому биотопу были даны дополнительные характеристики: описание подроста/подле-
ска (визуальная оценка проективного покрытия в % (группы по 25 %) с указанием породного
состава), тип увлажнения (недостаточное увлажнение - беломошники, брусничники, выруб-
ки исходных типов, сухие луга, поля со злаковой растительностью; умеренное увлажнение
- кисличники, черничники, богатые типы леса с дубравной и разнотравной растительностью;
избыточное увлажнение - болота, сфагновые типы леса, заболоченные долины ручьев/рек;
фрагментированное увлажнение - при наличии в биотопе различных типов увлажнения, чаще
всего на вырубках), уровень захламлённости (отсутствует - не отмечена; низкая - единично
зафиксированный естественный древесный отпад в виде упавших деревьев и их частей; уме-
ренная - присутствие естественного древесного отпада, не затрудняющего передвижение по
биотопу; высокая - при наличии значительного количества естественного древесного отпада
и поврежденных деревьев, которое затрудняет передвижение по биотопу; критическая - почти
«непроходимые» ветровалы и буреломы); проводимые в биотопах лесохозяйственные, гидро-
мелиоративные и биотехнические мероприятия.
Во время прохождения маршрута учетчик фиксировал на GPS-навигаторе точки смен био-
топов. Во избежание массового переноса кровососок с предыдущего биотопа и минимизации
ошибочного расчета плотности в последующих биотопах, было принято решение пересекать
видимые границы, предварительно сняв всех напавших кровососок, достаточно быстро (бегом),
фиксируя последнюю остановку в предыдущем биотопе и новую остановку в новом биотопе
не менее чем в 50 м от визуально выявленных границ. Также отмечались следы жизнедеятель-
ности видов прокормителей.
После прохождения маршрута подсчитывалось число собранных с учетчика крово-
сосок для каждого биотопа. Видовая принадлежность кровососок была подтверждена
на основании фундаментальной коллекции Зоологического института Российской ака-
38
демии наук. В программе BaseCamp для каждого биотопа рассчитывались пройден-
ное расстояние и время, затраченное на его прохождение. Данные с описаниями биото-
пов и характеристиками пройденных маршрутов вносились в таблицы Microsoft Excel.
В дальнейшем для каждого биотопа рассчитывалась плотность кровососок на 100 м,
на 10 мин, а также на 1 га (по формуле A/(B*30/10000) (где A - количество собран-
ных экземпляров в биотопе; B - расстояние (м), пройденное сквозь биотоп; 30 (15 м +
15 м) - ширина (м) трансекты, в зоне которой, по данным Иванова (1974), кровосо-
ски потенциально могут обнаружить учетчика даже в условиях низких температур.
Для всех обобщенных биотопов были рассчитаны максимальные и минимальные зна-
чения плотности кровососок. На основании опытных данных определяли период по-
тенциального лёта кровососок - период времени, в течение которого возможно встре-
тить окрылившихся особей, и период массового лёта - период, в течение которого
плотность окрыленных форм, напавших на человека, может достигать более 10 экз./
10 мин.
Оценка зараженности основных прокормителей
Основным прокормителем L. cervi в Ленинградской области является лось. Так как это един-
ственный многочисленный вид-хозяин оленьей кровососки, не запрещенный к охоте, основной
анализ зараженности проводился на образцах шкур особей, законно добытых в разрешенные
сроки.
Результаты различных схожих исследований в Фенноскандии свидетельствуют о том, что
зараженность лосей по «анатомическим областям» шкуры различна. При исследовании разных
участков шкуры одного лося (голова, передняя часть спины, задняя часть спины, передние
конечности, задние конечности и живот) отмечается, что на передней части спины обнару-
живается более половины всех кровососок (Paakkonen, 2012). Хранение и обработка всей
шкуры добытого лося - трудоёмкий и длительный процесс. Такой подход не представлялся
авторами настоящей статьи возможным и целесообразным. За основу методики была взята
методика из другой норвежской публикации, согласно которой авторы оценивали зараженность
на одном анатомическом участке шкуры (на шее) с определенными параметрами хранения
и обработки (Samuel et al., 2012).
Основными задачами данного этапа исследования являлись обработка участков шкур лосей,
механический подсчет количества кровососок на образцах и расчет плотности имаго и пупариев
на 1 дм2. Для оценки зараженности основных хозяев-лосей оленьей кровосоской использовали
участки шкур с шеи, площадью каждого не менее 2 дм2. Предоставленные охотниками фраг-
менты шкур законно добытых животных помещали в зип-пакет 30×40 и сопровождали данными
о месте сбора и половозрастными характеристиками добытого животного. Пакеты с образцами
хранились в морозильной камере при температуре - 18-22°С до окончательной обработки. Воз-
раст добытых лосей определяли по общепринятой методике по степени стертости коренных
зубов нижней челюсти (Кнорре, Шубин, 1959), у самцов с еще не сброшенными рогами для
определения возраста использовали дополнительную характеристику - окружность ствола рога
(Колесников, Макарова, 2014).
При обработке фрагмент размораживали в течение нескольких часов, далее измеряли его
площадь (дм2) с помощью измерительной ленты. Шерсть не полностью выстригали ножницами,
оставляя расстояние около 1 см от эпидермиса, оставшийся слои проверяли на наличие крово-
сосок. Подстриженный и измеренный кусок прочёсывали гребнем до полного освобождения от
39
кровососок. Количество кровососок подсчитывали для каждого образца. В случае разрезания
насекомого на 2 части учитывалась только передняя часть (голова). Кроме имаго, отмечали
количество отрожденных пупариев. Данные по зараженности были соотнесены с датами и
половозрастной характеристикой хозяев. Была измерена плотность заражения кровососками
для каждой особи.
РЕЗУЛЬТАТЫ
Биотопическое распределение окрыленных форм кровососок,
динамика лёта
Работа была выполнена в течение двух сезонов - 2020 и 2021. С 3 по 30 августа
2020 г. было пройдено 100 км. С 18 июля по 30 сентября 2021 г. было пройдено 92.1 км.
Всего учетами (79 маршрутов в 623 различных биотопах) были охвачены следую-
щие раионы Ленинградскои области: Выборгскии, Всеволожскии, Волосовскии,
Гатчинскии, Кингисеппскии, Ломоносовскии, Лужскии, Приозерскии. Всего было
собрано 3 902 крылатые особи. За 2 полевых сезона отмечено, что первые крылатые
особи появляются еще в июле. Независимо от проведенных исследований, автора-
ми при посещении лесов отмечены единичные нападения кровососок на человека
в октябре. Продолжительность периода лёта составляет более 91 дня (с 18.07 по
16.10 - даты первой и последней регистрации нападения кровососки на человека).
Период массового лёта по мнению авторов начинается в первой половине августа
(первая регистрация плотности >10 экз./10 мин 8 августа) и продолжается до второй
половины сентября (последняя регистрация плотности >10 экз./10 мин - 18 сентя-
бря). Наибольшие показатели плотности >50 экз./10 мин фиксировались в период
с 31 августа по 16 сентября. В табл. 1 приведены основные характеристики плотности
по выявленным биотопам.
Высокие показатели плотности напавших на человека кровососок фиксировались
в смешанных ельниках черничниках с наличием сфагнума в ЖНП (max = 35 экз./
10 мин); на зарастающих молодняками естественных полянах, перелесках (max =
81.4 экз./10 мин) и линейных объектах (ЛЭП, Газо/Нефтепроводы) (max = 31.1 экз./
10 мин); в сфагновых и увлажненных разнотравных сосняках (max = 83.3 экз./
10 мин); по окраинам верховых и переходных болот (max = 120.9 экз./10 мин);
в богатых разнотравных и увлажненных березняках (max = 155 экз./10 мин); в мо-
лодняках, возобновляемых после сплошных и постепенных рубок (max = 213.3 экз./
10 мин); в осинниках-кисличниках с высоким естественным отпадом осины (max =
264.2 экз./10 мин). Наиболее высокие значения плотности чаще всего фиксировались
в биотопах с избыточным увлажнением. При этом в сухих биотопах показатели мак-
симальной плотности в 4-5 раз ниже, чем в биотопах с избыточным увлажнением.
Разница между максимальными показателями плотности в биотопах с умеренным
увлажнением и в биотопах с избыточным тоже достаточно велика - 30-50%. Высокая
плотность >50 экз./10 мин наблюдалась в биотопах с критической захламленностью
и в биотопах с проективным покрытием подроста и подлеска 50-75%. Наблюда-
лось также, что плотность в схожих биотопах может быть разной в пределах одного
маршрута.
40
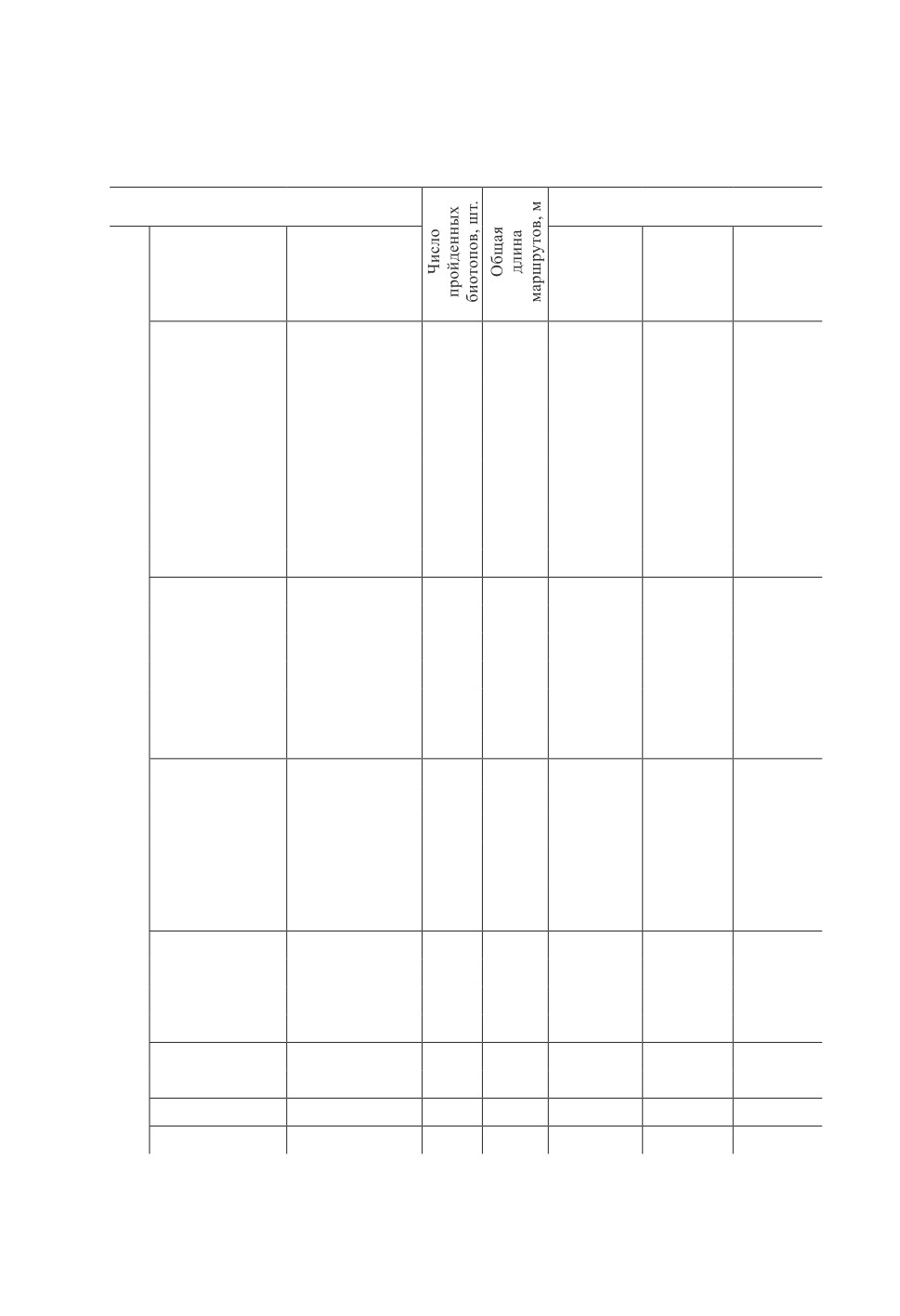
Таблица 1. Плотность оленьей кровососки в исследованных категориях биотопов
и основные количественные показатели измерений
Table 1. Density of deer ked in the studied biotope categories
and the main quantitative indicators of measurements
Биотоп
Плотность кровососки (мin-max)
Тип леса по
Тип леса по
преобладающей
ЖНП (биотопы
экз./100 м
экз./га
экз./10 мин
породе (биотопы
«Лес»)
«Лес»)
беломошник
1
598
0.5
1.7
1.5
брусничник
12
4032
0.1-1.3
0.4-4.5
0.5-5.7
кисличник
11
2263
0.4-1.3
1.5-4.4
0.2-3.8
черничник
41
11097
0.4-13.7
1.3-45.7
0.6-36.7
сосняк
разнотравный
3
436
0-22.1
0-73.5
0-21.4
сфагново-
30
7657
0.1-20.9
0.5-69.5
0.6-37.5
кустарничковый
сфагново-
13
3449
3.9-33.3
12.9-111.1
9.0-83.3
травяный
сфагновый
18
4297
0.7-38.0
2.4-126.6
2.5-49.0
кисличник
31
8170
0.3-38.0
0.9-126.8
1.1-155.0
приручейный
3
887
0.3-1.8
0.9-6.0
0.9-6.3
черничник
14
2610
1.9-5.0
6.5-16.8
2.0-13.3
березняк
разнотравный
44
9796
0.1-33.3
0.3-111.1
0.2-54.4
Лес
сфагново-
18
3935
0.6-26.2
1.9-87.2
1.3-85.0
травяный
сфагновый
9
2116
0.6-12.4
2.1-41.4
1.2-23.3
кисличник
55
19506
0.2-9.3
0.7-30.9
0.2-29.3
черничник
43
15095
0.3-12.7
1.1-42.2
0.8-35.0
разнотравный
5
2090
0-8.0
0-26.6
0-13.8
ельник
сфагново-
9
2995
0
0
0
кустарничковый
сфагново-
4
899
0-0.6
0-1.9
0-1.5
травяный
кисличник
30
7990
0.6-77.7
2.0-259.1
0.8-264.3
черничник
7
2572
0.3-9.3
1.0-31.0
0.3-9.5
осинник
разнотравный
6
1188
0.4-40.3
1.2-134.2
1.3-75.0
сфагновый
1
60
23.3
77.8
28.0
кисличник
4
762
0-0.3
0-1.2
0-0.5
сероольшанник
разнотравный
13
2689
0.9-4.3
2.8-14.5
2.5-13.3
черноольшанник
разнотравный
4
803
1.3-9.9
4.4-32.9
2.5-23.3
ивняк
разнотравный
1
131
0
0
0
41
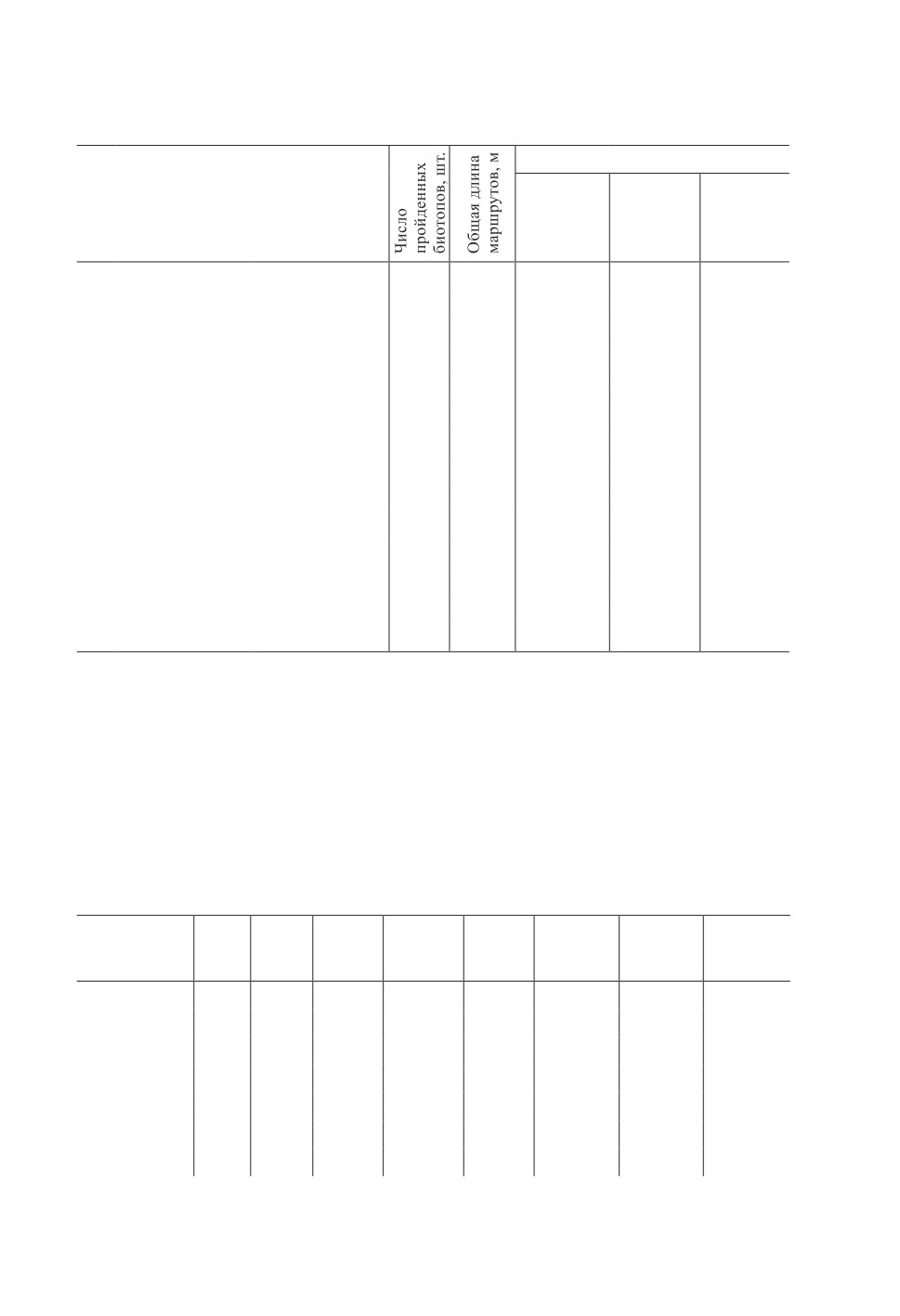
Таблица 1. Продолжение
Table 1. Continuation
Плотность кровососки (мin-max)
Биотоп
экз./100 м
экз./га
экз./10 мин
Болото верховое
15
5197
0.5-53.0
1.7-176.6
0.3-120.9
переходное
10
1690
0.7-11.8
2.3-39.5
0.9-10.7
Вырубка <1 года (свежая)
3
1053
0
0
0
<5 лет
21
7542
0.1-9.0
0.4-29.9
0.8-27.0
5-10 лет
25
7639
0.1-5.9
0.5-19.6
0.3-21.4
10-20 лет
28
11430
0.2-59.8
0.8-199.4
0.6-213.3
Погибшие насаждения
4
545
0-2.6
0-8.8
0-6.0
Гарь
1
289
1.7
5.8
5.6
ЛЭП/Газопровод
21
7778
0.1-7.2
0.4-23.9
0.5-31.1
Поля
18
10939
0.1-0.4
0.4-1.2
0.6-1.4
Пойменные луга
1
121
0
0
0
Пойменный лес
6
3659
0.2-1.5
0.6-4.9
0.5-4.3
Нелесохозяйственные молодняки
40
16160
0.2-35.0
0.6-116.6
0.7-81.4
Общий итог
623
192165
0.1-77.7
0.3-259.1
0.2-264.3
П рим еч ани я. Плотность кровососок рассчитана в разных единицах для возможного кор-
ректного сравнения результатов с результатами других аналогичных исследований.
Зараженность добытых животных
Всего за охотничий сезон 2020 г. было получено 8 образцов шкур животных, за-
конно добытых разными охотничьими коллективами, из Всеволожского (4 шт.), Луж-
ского (3 шт.) и Тосненского (1 шт.) районов Ленинградской области. Животные были
добыты в период с 17 октября по 26 декабря (табл. 2).
Таблица 2. Сведения о зараженности лосей, добытых в 2020
Table 2. Data on infestation of moose harvested in 2020
Площадь
Общее
Число
Плотность
Плотность
Район
Месяц
Пол
Возраст
образца,
число
пупариев,
мух экз./
пупариев,
дм2
мух, экз.
экз.
дм2
экз./дм2
Самец
Всеволожский
X
3.5
5.10
1362
10
267.1
1.96
Лужский
X
Самец
6.5-7.5
5.58
555
3
99.4
0.54
Самец
Всеволожский
XI
1.5
4.22
1937
25
459.0
5.92
Всеволожский
XI
Самка
3.5
3.72
231
1
62.1
0.27
Самец
Лужский
XII
2.5
2.98
466
7
156.4
2.35
Самец
Всеволожский
XII
4.5-5.5
2.54
272
1
107.2
0.39
Лужский
XII
Самка
3.5
8.72
156
0
17.9
0.00
42
После обработки и расчета были получены следующие данные: плотность крово-
сосок варьировала по образцам от 6 до 480 шт./дм2. Наиболее зараженный образец
шкуры принадлежал полуторагодовалому самцу, добытому во Всеволожском районе
21 ноября, наименее зараженный - самке в возрасте 3.5 года, добытой в Лужском
районе 26 декабря. Кроме представителей рода Lipoptena, других эктопаразитов об-
наружено не было.
ОБСУЖДЕНИЕ
Свободноживущие L. cervi массово приурочены к местообитаниям, связанным
с жизнедеятельностью прокормителей, поскольку, из-за особенностей анатомического
строения, держатся близко к местам своего появления. Численность лётных форм
L. cervi напрямую зависит от плотности хозяев и интенсивности использования ими
тех или иных стаций, о чем свидетельствуют и литературные источники (Балашов,
1996; Meier et al., 2014; Samuel et al., 2012). Пространственный аспект их распреде-
ления связан, в частности, с местами осенне-зимней концентрации прокормителеи.
Численность остальных хозяев (Capreolus capreolus (Linnaeus, 1758); Cervus elaphus
Linnaeus, 1758; Cervus nippon Temminck, 1838) на территории Северо-Запада, в сравне-
нии с численностью основного прокормителя - европейского лося - крайне мала, что-
бы значительно влиять на биотопическое распределение кровососок. Обнаруженные
места концентрации окрыленных форм могут служить индикатором зимних местоо-
битаний лосей (зимние кормовые угодья, богатые лиственной и сосновой порослью,
места стойбищ, жировок и лёжек), а также количественным индексом использования
разных участков геопространства лосями. При этом пространственное распределение
зимних кучек экскрементов, которое в свою очередь служит достоверным индикато-
ром использования территории крупными растительноядными (Månsson et al., 2011),
по неопубликованным данным одного из авторов данной статьи, в максимальных
значениях представлено в биотопах, аналогичных тем, в которых зафиксированы
максимальные показатели лета L. cervi. Однако между показателями встречаемости
следов жизнедеятельности лосей (экскрементов) осенне-зимнего периода и плотно-
стью летных форм L. cervi в аналогичных биотопах существуют некоторые разли-
чия, исходя из следующих особенностей жизненного цикла паразита и хозяина. Во-
первых, множество кровососок в среднем погибают раньше того периода, когда лоси
перестают потреблять в пищу древесно-веточный корм. Такое заключение сделано
на основании литературных данных и подтверждено тенденцией к уменьшению ин-
тенсивности заражения по исследованным образцам шкур, и на примере опытного
иммобилизованного лося (самец, 3.5-4.5 лет), у которого в начале апреля в шкуре не
обнаружилось ни одной кровососки. В то же время использование стаций лосями во
второй половине зимы и в весенний период может сильно отличаться от осеннего и
раннезимнего (Юргенсон и др., 1935; Тимофеева, 1974; Верещагин, Русаков, 1979),
а распределение экскрементов будет, в свою очередь, характерно для всего периода,
в который лоси употребляют древесно-веточный корм. Во-вторых, пупарии больше
всего выпадают из шкуры хозяев при активном движении и отряхивании, поэтому
наибольшая плотность может быть отмечена в местах жировок, лёжек и на основных
переходах между этими стациями, в то время как наибольшее количество экскремен-
тов можно будет обнаружить в кормовых угодьях. В-третьих, выживаемость пупариев
43
в условиях естественной среды, гипотетически, зависит от многих факторов, которые
в настоящее время досконально не изучены. По мнению авторов, два данных метода
оценки плотностей «следов жизнедеятельности», при использовании их для изучения
пространственно-временного использования территории представителями Cervidae,
могут значительно дополнять друг друга.
Неоднородность встречаемости кровососок в одинаковых биотопических условиях
может быть объяснена разницей в численности основных прокормителей и различной
интенсивностью использования ими стаций. Помимо этого приуроченность окрылен-
ных форм к местам с избыточным увлажнением свидетельствует не только о том, что
данные биотопы используют лоси, но и о том, что выживаемость пупариев выше на
увлажненных участках геопространства. Кроме этого, по данным Бахтушкиной (2018),
влажность воздуха 60-80% положительно влияет на продолжительность жизни имаго,
тем самым в более увлажненных биотопах в единицу времени может накапливаться
больше имагинальных форм, нежели в сухих биотопах. Существует также предпо-
ложение, что в сформированных биоценозах с высоким биоразнообразием множество
пупариев могут быть съедены консументами высоких порядков.
Также стоит отметить, что актуальные данные плотности летных форм превышают
некоторые ранее отмеченные в литературе. Так, по данным Балашова Ю.С., плот-
ность кровососок в 90-е годы в Ленинградской области на постоянных маршрутах не
превышала 33.5 экз./км. В Новгородской области максимальная плотность достигала
44.6 экз./км маршрута. При пересчете отловленных особей L. cervi на 1 км марш-
рута на некоторых участках плотность достигает 181 экз./км маршрута. Актуаль-
ные значения более чем в 4 раза превышают литературные. Однако другие данные
с Северо-Запада, а именно из Вологодской области, свидетельствуют о плотности
в 75-120 экз./мин (Варнаков, 1977), что, наоборот, значительно превышает данные
настоящего исследования. Вероятно, такая большая разница объясняется различиями
методики сбора и учета окрыленных форм. По представлениям авторов, если плот-
ность достигает 120 экз./мин, то у учетчика физически нет возможности подсчитать
всех кровососок в такое короткое время, тем более, что в таких биотопах при наличии
на учетчике множества кровососок возникает непреодолимое желание покинуть эти
места как можно быстрее. Для таких данных требуется проверка, однако авторы не
отрицают, что плотность кровососок в угодьях с высокой численность лося может
превышать рассчитанные показатели.
Наши данные о зараженности основных прокормителей оленьей кровосоской
(количество на 1 дм2) можно корректно сравнить с опубликованными данными от-
ечественных, финских и норвежских исследователей. В настоящий момент данные,
представленные в настоящей статье, - единственные сведения о зараженности лосей
оленьей кровосоской на Северо-Западе РФ, полученные по апробированной методике.
Данных выборки для представления достоверных средних значений по половозраст-
ным группам хозяев недостаточно, однако, возможно, средние значения не покажут
реальной картины, так как зараженность, по мнению авторов, может быть разной на
разных обособленных участках.
В исследовании финского коллеги Пакконена (Paakkonen, 2012) плотность оленьей
кровососки была максимальной на передней части спины и шее - здесь находилась
половина всех особей, зарегистрированных на теле хозяина. Вероятным объяснением
44
их скопления в этой части является максимальная длина шерсти, которая использу-
ется кровососками в качестве наилучшего убежища. Еще одним предположением,
возможно объясняющим такую высокую плотность, является положительный фото-
таксис. Последнее в свою очередь требует проверки образцами с остальной части
спины. Возможно, также кровососок привлекает большое количество капилляров и
кровеносных сосудов в шейном отделе, хотя на холке зарегистрирована максимальная
толщина кожи (Sokolov, Chernova, 1987).
По данным того же автора, плотность заражения на шее варьировала в диапазоне
26.0-97.6 экз./дм2. Полученные данные настоящего исследования имеют больший
диапазон - 17.9-459.0 экз./дм2. При этом единичные особи лосей могут иметь зара-
женность кровососками, в несколько раз превышающую зараженность в Финляндии
(по данным Пакконена (2012), средняя зараженность 10616 ± 1375 экз./особь, мак-
симальная 17490 экз./особь; по предположению авторов данной статьи, максималь-
ная зараженность лосей кровососками в Ленинградской области может достигать
30 000 экз./особь). Согласно данным советских авторов, средняя численность кровосо-
сок, паразитирующих на лосе, составляла 200-300 экз./особь, максимальная - порядка
1 000 экз./особь. Полуторагодовалый самец мог иметь максимальную плотность кро-
вососок (459 экз./дм2) в связи с высокой активностью перемещении в период массо-
вого лёта, который совпадает с периодом гона. Молодые лоси, которые не участвуют
в полной мере в процессе спаривания, но подвергаются иерархической конкуренции,
и которые не имеют устоявшегося участка обитания, могут совершать хаотичные
перемещения, тем самым увеличивая интенсивность заражения. Частично эти данные
подтверждаются данными из Фенноскандии (Madslien et al., 2012), но эта гипотеза
требует более серьезного подтверждения.
ЗАКЛЮЧЕНИЕ
Проведенное авторами исследование позволило получить новые данные о дина-
мике лётной активности, о биотопическом распределении окрыленных форм L. cervi
и новейшие сведения о зараженности хозяев на территории Ленинградской области.
По мнению авторов данной статьи, эти сведения могут быть полезны для предупре-
ждения нежелательных встреч людей, работающих и отдыхающих в лесу, с активно
нападающими паразитами, а также для формирования представления об инвазии на
самых крупных представителях Cervidae - лосях. Наряду с этим данные о плотности
не стоит считать точными «до экземпляра» и абсолютно информативными, а стоит
рассматривать только как относительные показатели обилия исследуемого объекта
в различных лесорастительных условиях. Использованная методика маршрутных уче-
тов позволяет охватить максимальное число биотопов и выявить места концентрации
окрыленных форм, что совместно с другими методиками может довольно точно сви-
детельствовать об активном использовании этих стаций прокормителями.
Новые данные о зараженности свидетельствуют о высокой инвазии лосей оленьей
кровосоской. Авторы рекомендуют проводить постоянные наблюдения за численно-
стью и распространением этих и других эктопаразитов копытных в Ленинградской
области для сохранения продуктивности популяций диких оленьих. Появившиеся
при обсуждении полученных результатов вопросы являются предметом дальнейших
исследований по данному направлению.
45
СПИСОК ЛИТЕРАТУРЫ
Балашов Ю.С. 1996. Изменения численности оленьей кровососки Lipoptena cervi (Hippobosci-
dae) в лесах Северо-Запада России. Паразитология
30
(2): 182-184. [Balashov Yu.S.
1996. The fluctuations of abundance of the deer louse-fly Lipoptena cervi (Hippobosci-
dae) in forests of the North-West Russia. Parazitologiya 30 (2): 182-184. (in Russian)].
Балашов Ю.С. 2009. Паразитизм клещей и насекомых на наземных позвоночных. Санкт-Петербург, Наука,
357 c. [Balashov Yu.S. 2009. Parasitism of ticks and insects on terrestrial vertebrates. Saint Petersburg,
Nauka, 357 pp. (in Russian)].
Бахтушкина А.И. 2018. Выживаемость имагинальных форм оленьих кровососок (Diptera, Hippoboscidae).
Российский паразитологический журнал 12 (3): 23-26. [Bakhtushkina A.I. 2018. Survivability of ima-
ginal forms of deer louse flies (Diptera, Hippoboscidae). Russian Journal of Parasitology 12 (3): 23-26.
(in Russian)].
Буракова О.В. 2002. Стоит ли опасаться оленьей кровососки Lipoptena cervi L. (Diptera, Hippoboscidae).
РЭТ-Инфо 2 (42): 16-19. [Burakova O.V. 2002. Should we be afraid of the deer ked Lipoptena cervi L.
(Diptera, Hippoboscidae)? RAT-Info 2 (42): 16-19. (in Russian)].
Варнаков А.П. 1977. Оленья кровососка в европейской тайге. Труды Всесоюзного сельскохозяйственного
института заочного образования 138: 78-81. [Varnakov A.P. 1977. Deer louse-fly in the European
taiga. Trudy Vsesoyuznogo sel'skohozyajstvennogo instituta zaochnogo obrazovaniya 138: 78-81. (in
Russian)].
Верещагин Н.К., Русаков О.С. 1979. Копытные Северо-Запада СССР (история, образ жизни и хозяйственное
использование). Ленинград, Наука, 309 с. [Vereshchagin N.K., Rusakov O.S. 1979. Ungulates of the
North-West of the USSR (history, lifestyle and economic use). Leningrad, Nauka, 309 pp. (in Russian)].
Иванов В.И. 1974. О вредоносности оленьей кровососки Lipoptena cervi L. (Diptera, Hippoboscidae)
в Белоруссии. Паразитология 8 (3): 252-253.
[Ivanov V. I. 1974. On the damage done by Lipop-
tena cervi L. (Diptera, Hippoboscidae) in Byelorussia. Parazitologiya 8 (3): 252-253. (in Russian)].
Иванов В.И. 1975. К антропофилии оленьей кровососки Lipoptena cervi L. (Diptera, Hippoboscidae). Ме-
дицинская паразитология и паразитарные болезни 4: 491-495.
[Ivanov V.I. 1975. On the attacks on
humans by Lipoptena cervi L. (Diptera, Hippoboscidae). Medical parasitology and parasitic diseases 4:
491-495. (in Russian)].
Колесников В.В., Макарова Д.С. 2014. Определение возраста самцов европейского лося (Alces alces alces
L.) по рогам. Вестник охотоведения 11 (2): 317-322. [Коlеsnikоv V.V., Маkагоvа D.S. 2014. Age
dеtermination of males of the European moose (Alces alces alces L.) by their horns. Vestnik okhotove-
deniya 11 (2): 317-322. (in Russian)].
Кнорре Е.П., Шубин Г.Г. 1959. Определение возраста лося. Труды Печоро-Илычского заповедника 7:
123-132. [Knorre E.P., Shubin G.G. 1959. Determining the age of the elk. Trudy Pechoro-Ilychskogo
gosudarstvennogo zapovednika 7: 123-133. (in Russian)].
Попов А.В. 1965. Жизненный цикл мух-кровососок Lipoptena cervi L. и Stenepteryx hirundinis L. (Diptera,
Hippoboscidae). Энтомологическое обозрение 44 (3): 573-583. [Popov A.V. 1965. The life cycle of louse-
flies Lipoptena cervi L. and Stenepteryx hirundinis L. (Diptera, Hippoboscidae). Entomological Review
44 (3): 573-583. (in Russian)].
Тимофеева Е.К. 1974. Лось (экология, распространение, хозяйственное значение). Ленинград: издательство
Ленинградского университета, 167 с. [Timofeeva E.K. 1974. Elk (ecology, distribution, economic value).
Leningrad, Publisher: Leningrad State University, 167 pp. (In Russian)].
Юргенсон П.Б., Капланов Л.Г., Книзе А.А. 1935. Лось и его промысел (распространение, экология и про-
мысел лосей). Москва, Типография «Гудок», 155 c. [Jurgenson P.B., Kaplanov L.G., Knise A.A. 1935.
Elk and its hunting (ecology, distribution and elk hunting). Moscow, Typography «Gudok», 155 pp. (In
Russian)].
Kaunisto S., Niemela P., Harkonen L., Roininen H., Ylonen H. 2011. Northward invasion of the parasitic deer
ked (Lipoptena cervi), is there geographical variation in pupal size and development duration. Parasitology
Kaunisto S., Ylönen H., Kortet R. 2015. Passive sinking into the snow as possible survival strategy during the
Madslien K., Ytrehus B., Vikøren T., Malmsten J., Isaksen K., Olav Hygen H., J. Solberg E. 2011. Hair-loss
epizootic in moose (Alces alces) associated with massive deer ked (Lipoptena cervi) infestation. Journal
46
Madslien K., Ytrehus B., Viljugrein H., Solberg E.J., Bråten K.R., Mysterud A. 2012. Factors affecting deer ked
(Lipoptena cervi) prevalence and infestation intensity in moose (Alces alces) in Norway. Parasites and
Månsson J., Andrén H., Sand H. 2011. Can pellet counts be used to accurately describe habi-
tat selection by moose Alces alces? European Journal of Wildlife Research 57: 1017-1023.
Meier C.M., Bonte D., Kaitala A., Ovaskainen O. 2014. Invasion rate of deer ked depends on spa-
tiotemporal variation in host density. Bulletin of Entomological Research 104: 314-322.
Paakkonen T. 2012. Ecophysiology of the deer ked (Lipoptena cervi) and its hosts. In: Pasanen P., Vornanen
M., Peiponen K. (eds). Dissertations in Forestry and Natural Sciences № 66. Joensuu, Publications of the
University of Eastern Finland, 83 pp.
Samuel B., Madslien K., Gonynor-McGuire J. 2012. Review of deer ked (Lipoptena cer-
vi) on moose in Scandinavia with implications for North America. Alces 48: 27-33.
Sokolov V.E., Chernova O.F. 1987. Morphology of the skin of moose (Alces alces L.). Swedish Wildlife Research.
Supplement. Vol. 1, 367-375.
DENSITY OF WINGED FORMS OF THE DEER KED
LIPOPTENA CERVI (LINNAEUS, 1758) IN DIFFERENT BIOTOPES
AND INTENSITY OF INFESTATION OF THE MAIN HOST
IN THE TERRITORY OF LENINGRAD PROVINCE
N. V. Sedikhin, A. V. Dmitryukov
Key words: deer ked, Lipoptena cervi, density, invasion, European moose, Alces alces
SUMMARY
The article presents the results of actual research to determine the population density
and distribution of the deer ked Lipoptena cervi (Linnaeus, 1758) flight forms in various
biotopes of Leningrad Province, and also contains information on the infestation of its main
host, the European moose Alces alces (Linnaeus, 1758). At the current stage of research,
the density determination of winged forms was carried out by a method of route registra-
tion with reference to biotopes. Mass flight starts in August 8-10 and lasts until September
18-20. The flight period lasts over 91 days (from July 18 to October 17). The density of
bloodsuckers >50 pcs/10 min was recorded from August 31 till September 16. The main
biotopes in which a significant number of flight forms were observed included mixed blue-
berry spruce forests with sphagnum in the ground cover (max = 35 pcs/10 min); natural
glades (max = 81.4 pcs/10 min) and linear objects (Power lines, Gas/Oil pipelines) (max =
31.1 pcs/10 min) overgrown with young stands; sphagnum and moistened forb pine forests
(max = 83.3 pcs/10 min); the outskirts of bogs and fens (max = 120.9 pcs/10 min); rich
herb and moistened birch forests (max = 155 pcs/10 min); young stands renewable after
clear and gradual cuts (max = 213.3 pcs/10 min); aspen sorrel forests with a high natural
mortality of aspen as a result of a windblow (max = 264.2 pcs/10 min). Even taking into
account rather small size of sample of examined animals, quantitative data on the number of
deer keds parasitizing moose in autumn - winter, obtained for the first time for Leningrad
Province, indicates an intense infection of various sex and age groups (min - 17 psc/dm2
for adult female, 26.12; max - 455 psc/dm2 for yearling male, 21.11).
47