ПАРАЗИТОЛОГИЯ, 2022, том 56, № 2, с. 91-107.
УДК 597. 552.5: 591.69 (282.247.212)
ФЕНОТИПИЧЕСКОЕ РАЗНООБРАЗИЕ ПОПУЛЯЦИОННЫХ
ГРУППИРОВОК PROTEOCEPHALUS LONGICOLLIS (ZEDER 1800)
(CESTODA: PROTEOCEPHALIDAE) -
ПАРАЗИТА СИГА COREGONUS LAVARETUS (L.)
© 2022 г. Л. В. Аникиеваa, *, Е. П. Иешкоa
aОбособленное подразделение Федерального государственного бюджетного учреждения
науки Институт биологии Карельского научного центра РАН
ул. Пушкинская, 11 Петрозаводск, Р. Карелия 185910 Россия
*e-mail: lva-45@mail.ru
Поступила в редакцию 16.02.2022 г.
После доработки 11.03.2022 г.
Принята к публикации 15.03.2022 г.
Изучено фенотипическое разнообразие популяции цестоды Proteocephalus longicollis (Zeder
1800) из сига Coregonus lavaretus (Linnaeus 1758) оз. Каменное (заповедник Костомукшский,
Северная Карелия). Дана характеристика изменчивости внутрипопуляционных группировок
P. longicollis, выделенных по дискретным признакам прикрепительного и трофико-
репродуктивного комплексов. Оценены разнообразие, сходство и различие морфометрических
показателей и роль отдельных группировок в фенотипическом разнообразии популяции. По-
казано, что основу популяции P. longicollis составляет фенотип с булавовидным сколексом.
Такой фенотип хорошо дифференцируется от ядровидного и ланцетовидного фенотипов более
крупными размерами сколекса и более низкими показателями изменчивости. Сделан вывод
о том, что важным фактором формирования фенотипического разнообразия и структуры по-
пуляции гельминта является экологический тип поведения хозяина, определяющий занимаемую
популяцией нишу. Адаптивный ответ популяции P. longicollis на изменение вида хозяина обе-
спечивается за счет изменения частот фенотипов, различающихся признаками прикрепления.
Ключевые слова: популяция, структура, фенотипическое разнообразие, обыкновенный сиг
Coregonus lavaretus, цестода Proteocephalus longicollis
DOI: 10.31857/S0031184722020016, EDN: FEOVCZ
Изучение внутривидовой изменчивости и популяционной организации вида -
одна из основных задач популяционной биологии, теории эволюции и систематики.
Определение роли изменчивости в процессе внутривидовой дифференциации, а также
91
в освоении видом различной среды обитания позволяет понять пути приспособления
вида к изменяющимся условиям среды (Майр, 1974; Тимофеев-Ресовский и др., 1977).
Изучение изменчивости и популяционной структуры вида у паразитических ор-
ганизмов представляет особенный интерес. В отличие от свободноживущих видов
животных и растений популяционная структура вида формируется как взаимодей-
ствие популяций паразита и хозяина в процессе их адаптации к условиям среды.
Двойственный характер среды обитания паразитов, в которой средой первого порядка
является живой организм, принципиально отличает адаптацию паразитов от адапта-
ции свободноживущих организмов. Специфичность как встречаемость в определен-
ном круге хозяев отражает определенный этап специализации и эволюции паразитов
и проявляется в исторически сложившейся степени соответствия определенного вида
паразита определенному виду или группе хозяев (Шульман, 1958; Добровольский
и др., 1994).
Анализ работ показывает, что спектр гельминтов, вовлекаемых в исследования
с позиций популяций, сравнительно широк и охватывает разные систематические
группы: моногеней, трематод, цестод, скребней. Популяционные группировки пара-
зитов, формирующиеся в разных видах хозяев, характеризуются специфическими па-
раметрами морфологических признаков, различаются набором дискретных признаков
и частотами их встречаемости (Шульман-Альбова, 1952; Фортунато, 1987; Пугачев,
1988; Евланов, 1992; Гиченок, 1995; и др.).
Цестода Proteocephalus longicollis (Zeder 1800) (син. P. exiguus) - широко рас-
пространенный паразит лососеобразных рыб. Для вида характерны широкий размах
морфологической изменчивости (Фрезе, 1965; Scholz, Hanzelova, 1998), высокая ге-
нетическая изменчивость (Král’ová, 1996; Král’ová, Spakulova, 1996; и др.) и морфо-
логический полиморфизм (Иешко, Аникиева, 1980; Hanzelova et al., 1995). Основные
хозяева P. longicollis - сиговые рыбы. В водоемах Европейского Севера из семейства
сиговых обитают два вида - европейская ряпушка Coregonus albula (Linnaeus 1758)
и обыкновенный сиг Coregonus lavaretus (Linnaeus 1758).
Экологическая изменчивость и фенотипическое разнообразие популяций P. lon-
gicollis наиболее подробно изучены в озерах Карелии. Выявлено два несвязанных
комплекса дискретных признаков: прикрепления (форма сколекса и апикальной при-
соски, расположение боковых присосок) и трофико-репродукции (форма члеников и
их внутренних структур). По форме сколекса выделено три вариации: ланцетовидная,
ядровидная и булавовидная, по форме половозрелых члеников три вариации типа
стробил: с короткими широкими члениками, с квадратными члениками и длинными
узкими члениками (Аникиева и др., 2004; Аникиева, Барская, 2008). Показано, что
в популяции P. longicollis, формируемой в европейской ряпушке C. albula, доминирует
92
только одна группировка с ядровидной формой сколекса и стробилой с половозрелы-
ми члениками квадратной формы. Группировки с другими фенотипами редки и мало-
численны (Аникиева и др., 2004; Аникиева, Иешко, 2007). Прослежены изменения
в структуре популяции P. longicollis в ареале европейской ряпушки (Аникиева, 2008).
В данной работе предпринято изучение популяционной морфологии P. longicol-
lis из сига Coregonus lavaretus оз. Каменного. Изучали дискретные группировки
P. longicollis для оценки их разнообразия, сходства и различий по морфометрическим
показателям и роли в фенотипическом разнообразии популяции.
МАТЕРИАЛ И МЕТОДЫ
Оз. Каменное (заповедник Костомукшский, бассейн р. Кемь, Белое море) - олиготрофный
водоем с чистой по химическому составу водой и разнообразной флорой и фауной. Водоем
представляет интерес как одна из немногих экосистем, сохраняющих естественное состояние и
близких к ненарушенным природным экосистемам. Рыбное население представлено 13 видами.
Среди них преобладают ряпушка, сиг, налим, а также окунь и щука (Первозванский, 1986).
Материалом для изучения морфологической изменчивости P. longicollis послужили сборы
цестод из обыкновенного сига. Методом неполного гельминтологического вскрытия в июне
2009-2010 гг. исследовано 33 экз. сигов. Цестоды были расслаблены в воде, зафиксированы
в спирте и окрашены кармином (Быховская-Павловская, 1985). Микроскопирование и изме-
рение червей выполнены с использованием оборудования Центра коллективного пользования
ИБ КарНЦ РАН. Измеряли только строго дорсо-вентрально расположенные на препаратах
особи. Для анализа трофико-репродуктивных признаков выбирали членики, находящиеся на
одной и той же стадии развития (со сформированными репродуктивными органами, но без яиц
в матке). Анализировали пластические признаки (ширина сколекса, диаметр боковых присосок,
длина и ширина апикальной присоски, длина и ширина половозрелых члеников, длина бурсы
цирруса, размах крыльев яичника) и один счетный признак (число семенников). Всего изучено
37 экз. половозрелых гельминтов. Анализ морфологической изменчивости выполнен с исполь-
зованием метода главных компонент Past 4.06 (Hammer et al., 2001) и пакета программ Statistica
5.0. Значимость различий и их величину определяли по t- и F- критериям (Лакин, 1990).
РЕЗУЛЬТАТЫ
Выборка цестод из сига оз. Каменное представлена особями, различающимися по
форме сколекса - ланцетовидной, ядровидной и булавовидной, которые были объеди-
нены в три группировки. Наиболее часто встречались особи с булавовидным ско-
лексом - 55% от числа исследованных, с ланцетовидным и ядровидным сколексами
соответственно 24 и 21%. Все три группировки имели широкий размах изменчивости
показателей ширины сколекса и различались их минимальными и максимальными
значениями. У группировки с ядровидным сколексом оба показателя были сдвинуты
влево, у группировки с булавовидным сколексом - вправо, а группировка с ланцето-
93
видным сколексом занимала промежуточное положение (табл. 1). Диапазон значений
ширины сколекса особей, принадлежащих к разным группировкам, перекрывался на
28%. При попарном сравнении группировок коридор перекрытия значений был шире
и варьировал от 50 до 65%. По средним значениям и дисперсии ширина сколекса
у ланцетовидной и ядровидной группировок была сходна, а булавовидная достоверно
отличалась от них более крупными размерами и меньшими значениями дисперсии
(табл. 1) (t
= 5.2, P>0.05; t
= 2.9, P>0.05; F
= 1.9, P>0.05; F
= 1.9,
Sc3-Sc2
Sc3-Sc1
Sc3-Sc2
Sc3-Sc1
P>0.05). Частотное распределение значений показало изменение распределения зна-
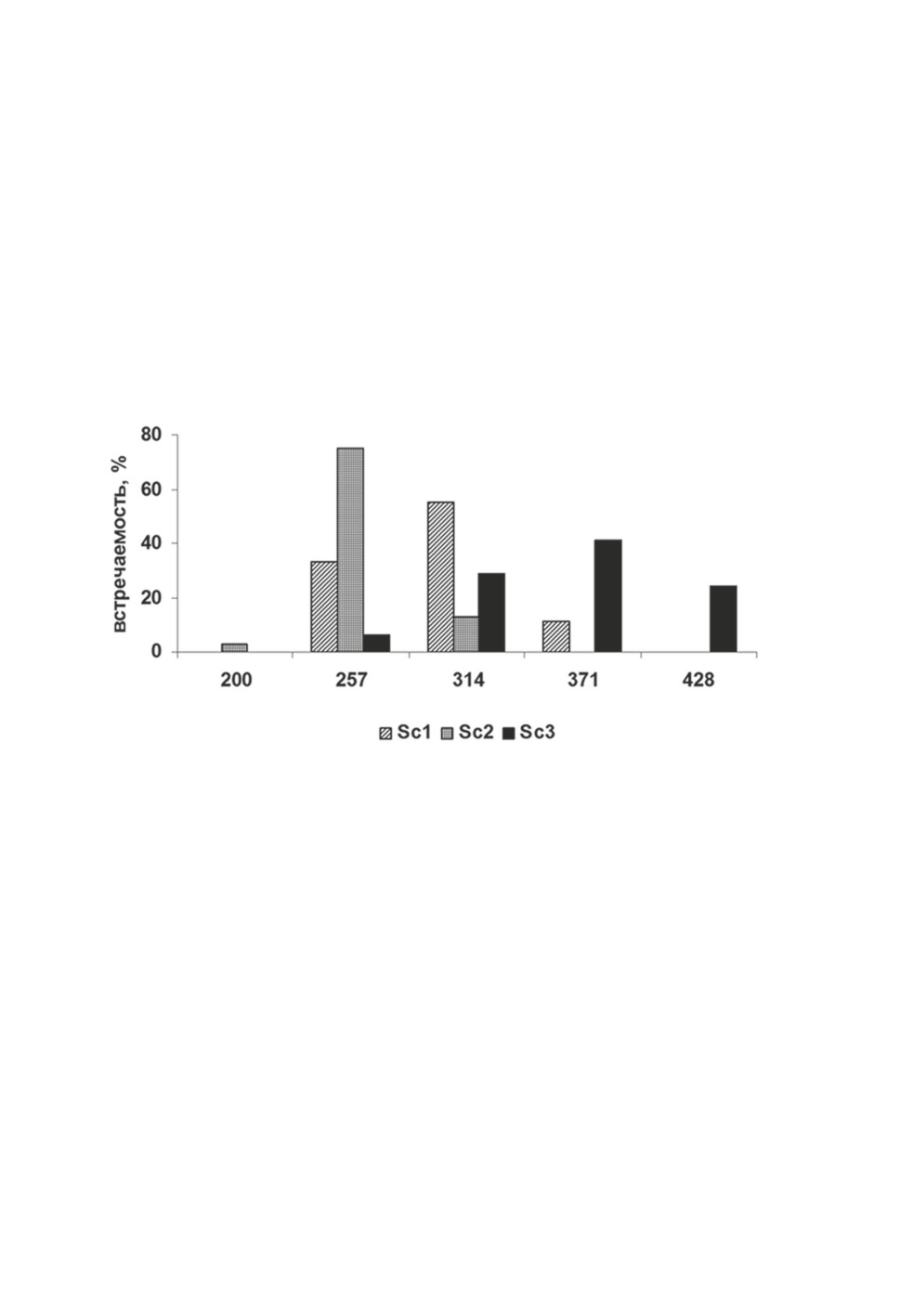
чений ширины сколекса по частотным классам у разных группировок (рис. 1).
Рисунок 1. Частотное распределение значений ширины сколекса (мкм)
в разных группировках P. longicollis из сига Coregonus lavaretus оз. Каменное.
Здесь и на рис. 2 и 3 Sc1 - ланцетовидная группировка, Sc2 - ядровидная группировка,
Sc3 - булавовидная группировка.
Figure 1. Frequency distribution of scolex width values across groupings of P. longicollis
in the whitefish Coregonus lavaretus from Lake Kamennoye, µm.
Note - here and below Sc1 is the lanceolate grouping, Sc2 is the nucleiform grouping,
and Sc3 is the clavate grouping.
Диаметр боковых присосок в выделенных группировках, так же как и ширина
сколекса, значительно варьировал. Колебания размеров боковых присосок ланцетовид-
ной и булавовидной группировок полностью входили в диапазон значений боковых
присосок ядровидной группировки. Общий диапазон перекрытия значений диаметра
боковых присосок у выделенных группировок составил 65%. По средним значениям
боковые присоски изучаемых группировок были сходны, но различались характером
изменчивости (табл. 1) (F
= 2.2, P>0.05; F
= 2.8, P>0.05).
Sc2-Sc1
Sc2-Sc3
94
Таблица 1. Морфометрические показатели (мкм) прикрепительного комплекса признаков
Proteocephalus longicollis из сига оз. Каменного
Table 1. Morphometric parameters (µm) of the set of attachment features
of Proteocephalus longicollis in whitefish from Lake Kamennoye
Признак
Группировка
Пределы
М±m
Дисперсия
Ширина сколекса
Sc1
228-370
293±14
1877
Sc2
194-342
251±15
1860
Sc3
285-399
339±8
993
Диаметр боковой присоски
Sc1
85.5-119.7
108.0±3.6
117
Sc2
79.8-131.1
98.2±5.4
258
Sc3
85.5-119.7
96.9±2.5
93
Длина апикальной присоски
Форма A1
45.6-85.5
62.7±2.9
159
Ширина апикальной присоски
17.1-39.9
28.5±1.9
65
Длина апикальной присоски
Форма A2
57.0-85.5
66.5±3.2
152
Ширина апикальной присоски
17.0-39.0
29.0±2.5
81
Высота мышечного валика
5.7-22.8
14.8±1.1
18
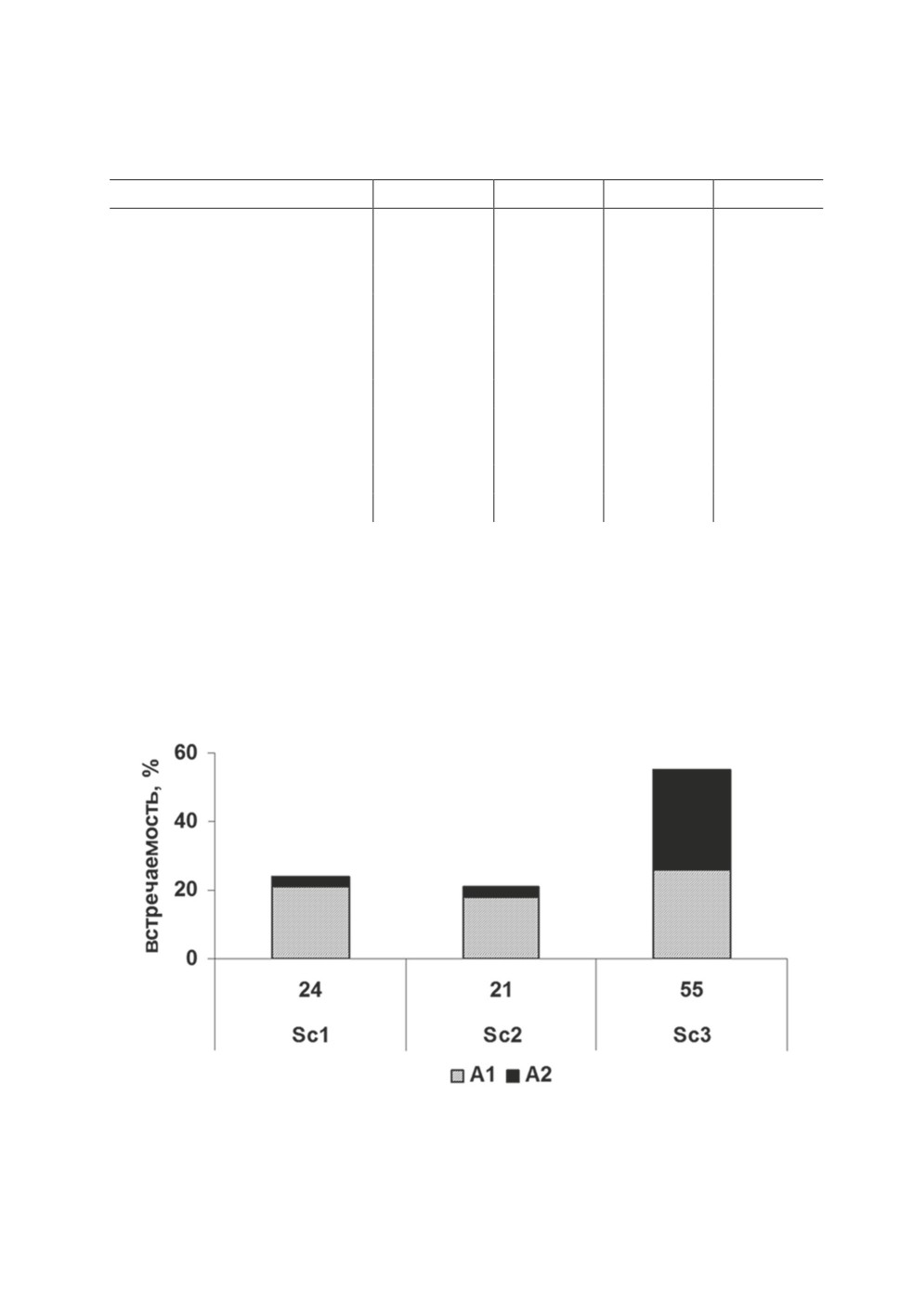
Вариации формы апикальной присоски (простая А1 и сложная - с мышечным
валиком А2) были обнаружены во всех трех группировках. У ланцетовидной и ядро-
видной группировок чаще встречалась простая форма апикальной присоски, у бу-
лавовидной группировки обе вариации формы апикальной присоски встречались со
сходной частотой (рис. 2).
Рисунок 2. Встречаемость (%) вариаций формы апикальной присоски в группировках
P. longicollis из сига Coregonus lavaretus оз. Каменное.
Figure 2. Prevalences (%) of apical sucker shapes in P. longicollis groupings in the whitefish
Coregonus lavaretus from Lake Kamennoye.
95
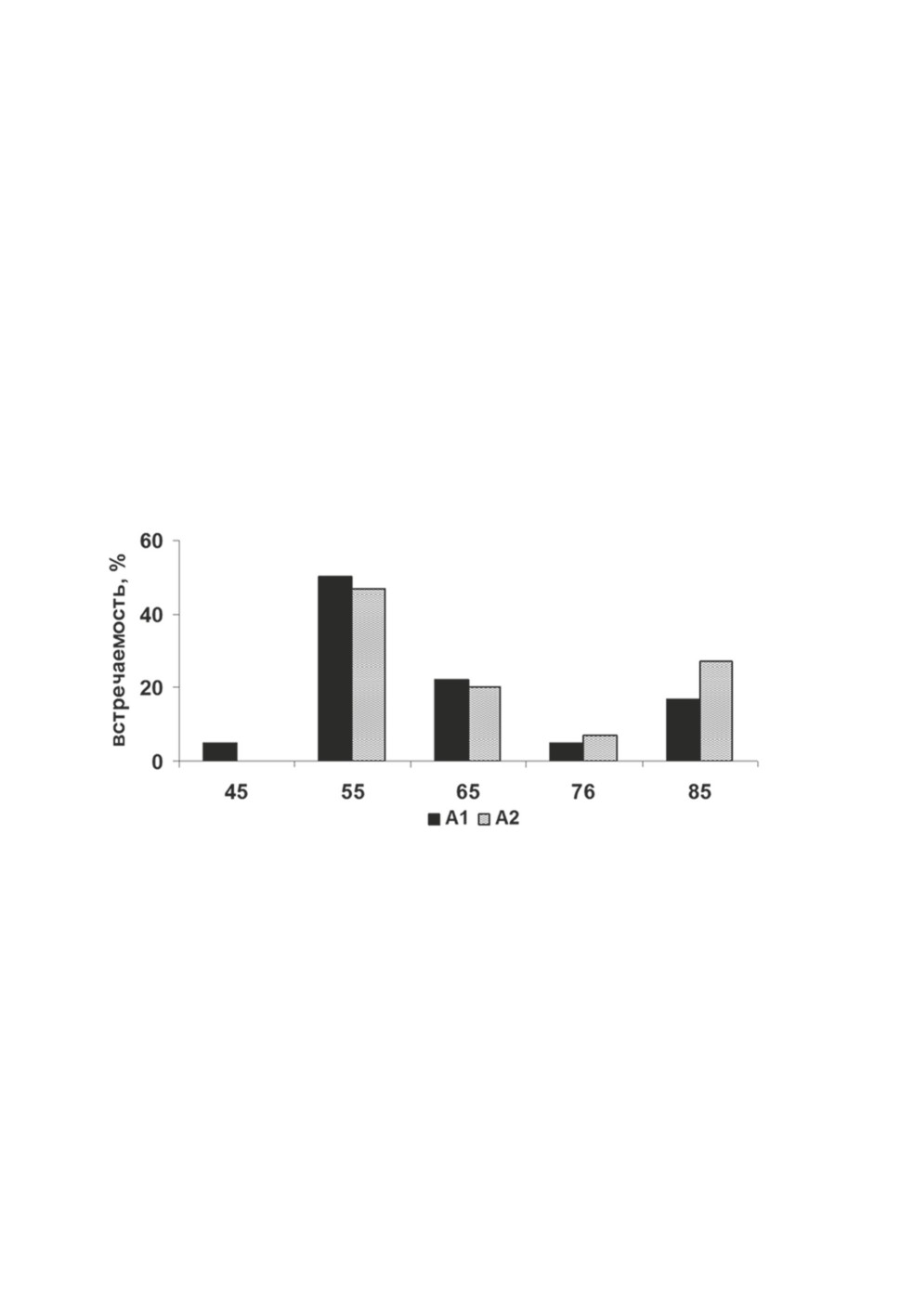
Размеры апикальной присоски как А1, так и А2 широко варьировали и были
сходны по пределам, средним значениям и характеру их варьирования. Анализ соот-
ношения длины и ширины апикальной присоски показал, что ее форма высоко из-
менчива и менялась от уплощенно продолговатой (45-65 мкм × 17 мкм) до округло
продолговатой (45 мкм × 40 мкм). При одних и тех же значениях длины присоски
ее ширина варьировала от минимальных значений до максимальных. Чаще встреча-
лась апикальная присоска овальной формы (78 %). Крайние варианты были редки,
с плоской формой - 17%, округлой - 5%. Доминирующие размеры мышечного валика
апикальной присоски А2 находились в пределах 17-23 мкм (90%). Крайние варианты
встречались редко. Частотное распределение значений длины апикальной присоски
имело двувершинную кривую, при этом обе формы имели сходный характер варьи-
рования этого признака (рис. 3).
Рисунок 3. Распределение значений длины (мкм) апикальной присоски
Proteocephalus longicollis из сига Coregonus lavaretus оз. Каменное.
Figure 3. Distribution of apical sucker length (µm) values of Proteocephalus longicollis
in the whitefish Coregonus lavaretus from Lake Kamennoye.
Изучение встречаемости формы половозрелых члеников P. longicollis, которая
определяет экстерьер стробилы, показало, что в общей выборке цестод наиболее рас-
пространена группировка с половозрелыми члениками квадратной формы Р2 (66%),
группировка с короткими и широкими члениками Р1 составила 23%, группировка
с длинными и узкими члениками Р3 - 11%. При сравнении морфометрических показа-
телей особей по комплексу трофико-репродуктивных признаков было установлено, что
96
группировки различались направленностью границ изменчивости трех положительно
связанных признаков - ширины членика, длины бурсы цирруса и размаха крыльев
яичника. Границы изменчивости каждого из этих трех признаков представляли собой
цепь последовательно увеличивающихся значений от группировки Р3 к группировке
Р2 и затем к группировке Р1. Группировка Р1 имела хиатус с группировкой Р3 по
ширине половозрелых члеников, очень узкие границы пересечения значений по длине
бурсы цирруса (11%) и размаху крыльев яичника (4%). Группировки Р1 и Р3 также
имели хиатус по длине половозрелых члеников, а группировка Р2 занимала проме-
жуточную (центральную) позицию и перекрывалась с группировками Р1 и по Р3 по
длине и ширине половозрелых члеников, длине бурсы цирруса и размаху крыльев
яичника. По средним значениям признаков выявлены достоверные различия между
группировками Р1 и Р2 в размерах половозрелых члеников. Половозрелые членики
в группировке Р1 были короче и шире, чем в группировке Р2. Критерий Стьюдента
по длине члеников составил 5.8, по ширине - 3.2; P>0.05. Длина члеников в груп-
пировке Р1 отличалась меньшей изменчивостью, чем в группировке Р2 (F = 5.0,
P>0.05). Признаки внутренних структур - число семенников, длина бурсы цирруса
и размах крыльев яичника - не имели достоверных различий по средним значениям
показателей, но различались характером варьирования числа семенников F = 3.16,
P>0.05. По длине бурсы цирруса и размаху крыльев яичника различия были недо-
стоверны (табл. 2).
Таблица 2. Морфометрические показатели Proteocephalus longicollis из сига оз. Каменного
по комплексу трофико-репродуктивных признаков, мкм
Table 2. Morphometric parameters of the set of trophic and reproductive features
of Proteocephalus longicollis in whitefish from Lake Kamennoye, µm
P1
P2
P3
Признак
Пределы
М±m
Дисперсия
Пределы
М±m
Дисперсия
Пределы
Длина членика
339-509
427±24
4441
452-960
658±32
22253
678-1356
Ширина членика
734-1243
999±74
43375
452-1073
743±28
17161
395-678
Число семенников
42-94
63.6±9
550
38-86
57.3±3
174
53-60
Длина бурсы
226-350
288±18
2444
214-361
268±8
1428
159-249
цирруса
Размах крыльев
452-847
680±57
22959
283-757
526±24
13106
249-475
яичника
97
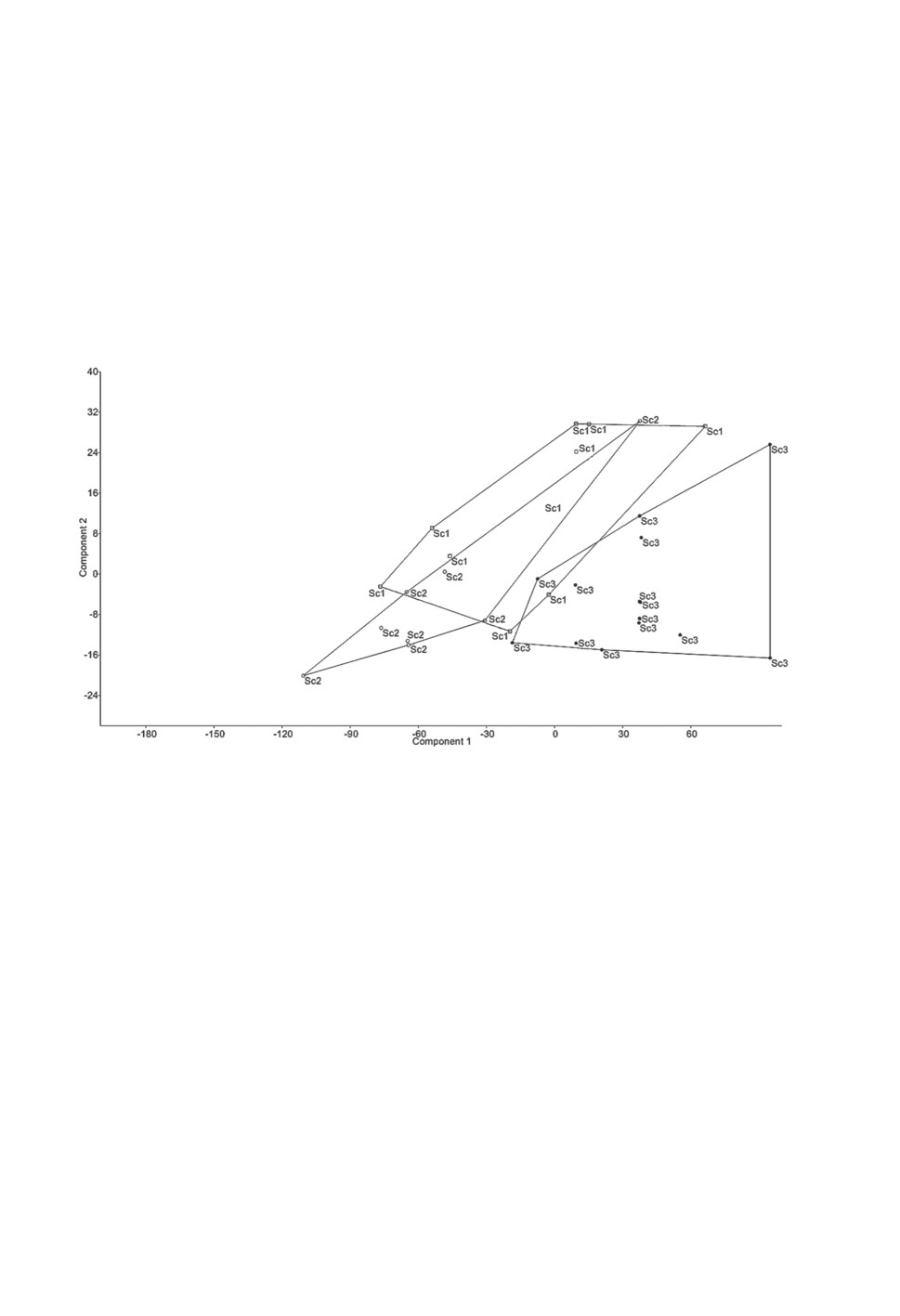
Исследование выборки методом главных компонент показало, что по морфо-
метрическим показателям комплекса признаков прикрепления группировка особей
с булавовидным сколексом хорошо отличается от двух других по компоненте 2 (ши-
рина сколекса). Фенотипические группировки Sc1 и Sc2 перекрываются, образуя
единое поле в координатах компоненты 1 (процент объясненной дисперсии 88.28) и
компоненты 2 (процент объясненной дисперсии 7.14). Сходство объясняется высоким
вкладом и близостью значений признаков апикальной присоски формы A1 и боковых
присосок (рис. 4).
Рисунок 4. Популяционная структура фенотипических группировок Proteocephalus longicollis
по признакам сколекса, на основе анализа главных компонент (PC).
Figure 4. Population structure of Proteocephalus longicollis phenotypic groupings by scolex
features, based on principal component (PC) analysis.
По комплексу трофико-репродуктивных признаков выборка распалась на три
отдельные группировки, в координатах главных компонент (рис. 5). Показано, что
фенотипические группировки Р1, Р2 и Р3 представлены тремя обособленными груп-
пировками в координатах компоненты 1 (процент объясненной дисперсии 66.43) и
компоненты 2 (процент объясненной дисперсии 31.33). Различия выявленных фе-
нотипов члеников цестод объясняются высоким вкладом значений длины члеников,
которые выделяют группировку фенотипа Р3, и признаков ширины половозрелых
члеников и размаха крыльев яичника, вклад которых определяет выделение группи-
ровки фенотипов Р1. Как видно на рис. 5 наиболее массовый фенотип Р2 занимает
центральное положение.
98
Рисунок 5. Популяционная структура фенотипических группировок Proteocephalus longicollis
по признакам половозрелых члеников, на основе анализа главных компонент (PC).
Figure 5. Population structure of Proteocephalus longicollis phenotypic groupings by mature
proglottid features, based on principal component (PC) analysis.
ОБСУЖДЕНИЕ
Согласно современным представлениям под изменчивостью понимается явление
разного индивидуального воплощения эпигенотипа в фенотипе, реализация обуслов-
ленного развитием законов возможного (допустимого) преобразования отдельных
признаков. Фенотип является результатом совместного действия гена и условий внеш-
ней среды. Основополагающее влияние на наследственность оказывает не геном,
а эпигенетическая система - совокупность факторов, воздействующих на онтогенез
(Шишкин, 1988; Васильев и др., 2007).
Онтогенез цестод в процессе эволюции претерпел глубокую трансформацию. Она
связана с возникновением интеркалярной церкоидной стадии и способностью раз-
виваться с участием разного числа хозяев в жизненном цикле. Поливариантность
траекторий жизненного цикла и полигостальность являются механизмом поддержания
динамической устойчивости популяций цестод. На их основе в различных экологи-
ческих условиях возникает большое разнообразие форм (Гуляев, 1998).
Проведенные нами исследования показали, что популяция P. longicollis, форми-
руемая в обыкновенном сиге, разнородна по дискретным признакам, выполняющим
важные функции жизнедеятельности гельминтов: прикрепления и трофики, и пред-
ставлена особями, различающимися формой сколекса, апикальной присоски и по-
ловозрелых члеников. По форме сколекса P. longicollis встречаются 3 вариации, по
99
форме апикальной присоски - 2, по форме половозрелых члеников - 3. Объединение
особей со сходными вариациями дискретных признаков в группировки позволило
оценить характер их изменчивости популяционными методами. Полученные данные
показали широкие границы изменчивости признаков в дискретных группировках
P. longicollis, выявили различия в средних значениях и варьирования, а также в ха-
рактере частотного распределения показателей. Широкий диапазон разнообразия по-
ловозрелых особей P. longicollis позволяет оценить их свойства как «нормальные»
для данных условий среды (Шмальгаузен, 1982, 1983) и определяет широкую норму
реакции как сложившейся адаптивной возможности организмов (Северцов, 1990).
Важным аспектом характера изменчивости является распределение значений при-
знаков в вариационном ряду. На материале по индивидуальной изменчивости темпов
роста личинок Rana temporaria (Linnaeus 1758) в разных условиях среды Северцов
и Сурова (1981) показали, что распределение по признаку с широкой нормой реак-
ции отражает генотипическое разнообразие популяции. Полученные нами данные
о высоком фенотипическом разнообразии особей в группировках P. longicollis со-
гласуются с генетическими исследованиями, проведенными с помощью RAPD-PCR
анализа (Král’ová, Spakulova, 1996). Так, в RAPD профиле 20 экз. взрослых цестод
P. longicollis, полученном с помощью OPA 04 праймера, только три фрагмента были
постоянны и обнаружены в каждом из исследованных экземпляров цестод, 6 фраг-
ментов встречались с максимальной частотой, один фрагмент был очень редким.
Применение метода главных компонент наглядно продемонстрировало высокую
фенотипическую изменчивость выделенных группировок и позволило выделить их
наиболее существенные особенности. По морфометрическим показателям комплекса
признаков прикрепления булавовидная группировка имеет более крупные размеры
сколекса, меньший диапазон изменчивости его ширины и диаметра боковых присо-
сок, высокие показатели встречаемости апикальной присоски с вариацией А2. Две
другие группировки имеют большýю площадь перекрытия параметров признаков, но
различаются их крайними значениями. По комплексу трофико-репродуктивных при-
знаков группировки четко различаются длиной половозрелых члеников. Группировка
с квадратными члениками занимает центральное место. Две другие располагаются
по разные стороны от нее. Распределение особей в координатах главных компонент
показывает, что по совокупности значений признаков каждая особь занимает свое
место в группировке, так же как и группировки в структуре популяции P. longicollis.
При этом каждая из особей иерархически соотносится с группировкой, к которой она
принадлежит, соответственно каждая субпопуляционная группировка иерархически
соотносится с популяцией P. longicollis. Все вместе они создают эпигенетический
100
ландшафт популяции, который инвариантен для входящих в нее особей (Васильев
и др., 2007).
Выделенные группировки представляют собой элементарную структурную еди-
ницу популяции P. longicollis. По сочетанию двух признаков - формы сколекса и
формы апикальной присоски - можно выделить несколько фенотипов. На их основе
обеспечивается адаптивный ответ популяции на изменение среды. Анализ распреде-
ления фенотипов по форме сколекса и форме апикальной присоски выявил сложную
структуру популяции по признакам прикрепления. Фенотип с булавовидным сколексом
и двумя формами апикальной присоски составляет основу популяции P. longicollis.
Относительно многочисленны фенотипы с ланцетовидным и ядровидным сколексом,
и простой апикальной присоской. Два фенотипа - с ланцетовидным и ядровидным
сколексами и сложной апикальной присоской малочисленны. Доминирующий фенотип
с булавовидным сколексом хорошо дифференцируется от них более крупными раз-
мерами сколекса и более низкими показателями его изменчивости.
Преобладание в структуре популяции P. longicollis фенотипа с булавовидной фор-
мой сколекса и фенотипа стробилы с квадратной формой половозрелых члеников
позволяет считать их более приспособленными к условиям среды обитания и опреде-
лить булавовидный фенотип сколекса и квадратный фенотип половозрелых члеников
стробилы как ядро популяции - устойчивую внутрипопуляционную структуру, обе-
спечивающую эффективное воспроизводство популяции. Другие фенотипы составляют
периферию популяции. Их высокая изменчивость способствует расширению границ
разнородности и повышает пластичность и устойчивость популяции.
Ранее было показано, что популяция P. longicollis, формируемая в европейской
ряпушке, обладает высоким фенотипическим разнообразием, которое проявляется
в широком наборе дискретных вариаций признаков и их сочетаний. Однако в структу-
ре разнообразия доминирует одна группировка по признакам прикрепления, имеющая
сколекс ядровидной формы, и одна группировка по трофико-репродуктивным при-
знакам стробилы - с половозрелыми члениками квадратной формы. Остальные мало-
численны и редки, что свидетельствует о жестком регулировании хозяином нормы
реакции паразита (Аникиева и др., 2004).
Сиг и ряпушка - представители одного рода Coregonus Linnaeus 1758. Как и всем
сиговым им свойственны высокая генетическая изменчивость и фенотипическая пла-
стичность. Обыкновенный сиг Coregonus lavaretus sensu lato имеет циркумполярное
распространение. На протяжении ареала он образует множество экологических форм,
которые занимают разные экологические ниши, имеют разные типы питания, различа-
ются темпом роста, местами и сроками нереста. В озере Каменное сиги представлены
101
двумя экологическими формами: малотычинковой и среднетычинковой. Наиболее
многочислен малотычинковый сиг (Первозванский, 1986). Обе экологические формы
обыкновенного сига из оз. Каменное пластичны по размерно-весовым признакам,
темпу роста, продолжительности жизни, плодовитости. Анализ молекулярного раз-
нообразия (AMOVA) выявил высокое внутрипопуляционное разнообразие (90% от
всей генетической изменчивости) сигов в оз. Каменное. Анализ изменчивости гена
ND1 ДНК показал, что преобладающие гаплотипы обеих экологических форм сигов
достаточно хорошо дифференцированы друг от друга (Ильмаст и др., 2016).
Ареал европейской ряпушки охватывает бассейны Северного, Балтийского, Барен-
цева и Белых морей. В пределах вида выделяют 2 формы европейской ряпушки -
мелкую длиной 10-12 см, массой 50-70 г и крупную массой до 200 г (Потапова,
1978). Из них самыми крупными являются ладожский рипус и онежский килец (Рыбы
в заповедниках России, 2010).
Изменчивость и многообразие форм сиговых рыб определяются особенностями
их питания в водоемах Севера (Решетников, 1980). Европейская ряпушка - специали-
зированный планктофаг. Типичная мелкая ряпушка в течение всей жизни питает-
ся планктоном. Крупная ряпушка, оставаясь планктофагом, способна использовать
в пищу амфибиотических и воздушных насекомых, нектобентические и бентические
организмы. Спектр и интенсивность питания ряпушки зависят от температурного
режима водоема, от состава и величины запасов корма и физиологического состояния
рыбы (Потапова, 1978; Решетников, 1980). Сиги оз. Каменное относятся к бентофагам
с широким спектром питания. Ведущими группами являются моллюски, личинки
хирономид, поденок, ручейников, веснянок и бокоплавы. Встречаемость веслоногих
ракообразных, в число которых входят и промежуточные хозяева P. longicollis, не
превышает 10%. Спектр питания сигов разный в разные годы, у рыб разного раз-
мера и возраста, а также у особей с крайними вариантами числа жаберных тычинок
в пределах одной популяции (Первозванский, 1986; Ильмаст, Стерлигова, 2012).
Популяции сига и ряпушки, несмотря на то, что они могут обитать в одном во-
доеме, всегда в большей или меньшей степени изолированы друг от друга. Места
обитания сига в период открытой воды приурочены к песчано-илистым участкам
литоральной зоны, где он нагуливается в летний период. Ряпушка обитает в цен-
тральной части озера. Различия обнаруживаются и в местах нереста сига и ряпушки
(Решетников, 1980).
Известно, что внутрипопуляционный отбор является реакцией различных гено-
типов на воздействие любых локальных условий среды и приводит к постепенному
смещению частот различных генотипов в направлении средней приспособленности
102
популяции в целом. На этом пути формируются локальные адаптации и протека-
ет формирование популяционно-генетической основы локальных дифференцировок
(Тимофеев-Ресовский и др., 1977). Особенности биологии хозяев P. longicollis и
разделение их популяций по экологической нише формируют степень изоляции по-
пуляционных гостальных группировок паразита и накопление между ними генети-
ческих и морфологических различий. Разнообразие экологических условий обитания
P. longicollis приводит к тому, что в разных популяциях направленность естественного
отбора оказывается различной.
ЗАКЛЮЧЕНИЕ
Полученные нами данные по фенотипической изменчивости и структуре популя-
ции Р. longicollis, формируемой в сиге, и сопоставление с известными материалами
Р. longicollis из европейской ряпушки показывают, что важным фактором формиро-
вания фенотипического разнообразия и структуры гельминта является экологический
тип поведения хозяина, определяющий занимаемую популяцией нишу.
БЛАГОДАРНОСТИ
Финансовое обеспечение исследований осуществлялось из средств федерального
бюджета на выполнение государственного задания КарНЦ РАН (FMEN-2022-0005,
№ г.р. 122032100130-3).
СПИСОК ЛИТЕРАТУРЫ
Аникиева Л.В. 2008. Популяционная изменчивость Proteocephalus longicollis (Cestoda: Proteocephalidae)
из европейской ряпушки Coregonus albula озер Карелии. Паразитология 42 (1): 3-12. [Аnikieva L.V.
2008. Population variability of Proteocephalus longicollis (Cestoda: Proteocephalidae) from the European
vendace Coregonus albula from lakes of Karelia. Parazitologiya 42 (1): 3-12. (in Russian)].
Аникиева Л.В., Барская Ю.Ю. 2008. Морфологическое разнообразие паразита лососевидных рыб цестоды
Proteocephalus longicollis (Zeder, 1800) из сига Coregonus lavaretus малых водоемов национального
парка “Паанаярви” // Труды КарНЦ РАН. Вып. 13. Петрозаводск, КарНЦ РАН, 12-16. [Anikieva L.V.,
Barskaya Yu.Yu. 2008. Morphological diversity of salmon parasite Proteocephalus longicollis (Zeder, 1800)
from whitefish Coregonus lavaretus in small water bodies of the Paanajärvi National Park // Transactions
of Karelian Research Center of Russian Academy of Science. Issue. 13. Petrozavodsk, KarRC RAS,
12-16. (in Russian)].
Аникиева Л.В., Иешко Е.П. 2007. Морфологический полиморфизм цестоды Proteocephalus longicollis (Ces-
toda: Proteocephalidae) в онтогенезе. Паразитология 41 (2): 103-112. [Anikieva L.V., Ieshko E.P. 2007.
Morphological polymorphism of the cestode Proteocephalus longicollis (Cestoda: Proteocephalidae) in
ontogeny. Parazitologiya 41 (2): 103-112. (in Russian)].
103
Аникиева Л.В., Харин В.Н., Спектор Е.Н. 2004. Полиморфизм и структура популяции Proteocephalus
longicollis (Zeder, 1800) (Cestoda: Proteocephalidae) из европейской ряпушки Coregonus albula L.
Паразитология 38 (5): 438-447. [Anikieva L.V., Kharin V.N., Spector E.N. 2004. Polymorphism and
population structure of Proteocephalus longicollis (Zeder, 1800) (Cestoda: Proteocephalidae) from the
European vendace Coregonus albula L. Parazitologiya 38 (5): 438-447. (in Russian)].
Быховская-Павловская И.Е. 1985. Паразиты рыб. Руководство по изучению. Л.: Наука. 121 с. [Bykhovskaya-
Pavlovskaya I.E. 1985. Fish parasites. Study guide. L.: Science. 121 pp. (in Russian)].
Васильев А.Г., Васильева И.А., Большаков В.Н. 2007. Феногенетическая изменчивость и методы ее изуче-
ния. Учебное пособие. Отв. ред. докт. биол. наук И.М Хохуткин. Екатеринбург, изд-во Уральского
университета, 279 с. [Vasilyev A.G., Vasilyeva I.A., Bolshakov V.N. 2007. Phenogenetic variability and
methods of its study. Tutorial. Managing ed. Doctor of Biological Sciences I.M. Khokhutkin. Ekaterinburg,
publishing house of the Ural University, 279 pp. (in Russian)].
Гиченок Л.А. 1995. Изменчивость и фенотипическое разнообразие скребня Echinorhynchus gadi (Acantho-
cephala) из двух видов беломорских рыб. Зоологический журнал 74 (8): 15-26. [Gichenok L.A. 1995.
Variability and phenotypic diversity of the acanthocephalan Echinorhynchus gadi (Acanthocephala) from
two White Sea fish species. Zoologicheskii Zhurnal 74 (8): 15-26. (in Russian)].
Гуляев В.Д. 1998. Становление организации цестод и основные направления ее преобразования в эволю-
ции. Автореф. дис. … докт. биол. наук. М. 46 с. [Gulyaev V.D. 1998. Formation of the organization
of cestodes and the main directions of its transformation in evolution. Abstract dis. …d. b. s. M., 46 pр.
(in Russian)].
Добровольский А.А., Евланов И.А., Шульман С.С. 1994. Паразитарные системы: анализ структуры и
стратегии, определяющей их устойчивость. Отв. ред. С.С. Шульман. Экологическая паразитология.
Петрозаводск: Карельский научный центр РАН: 3-43. [Dobrovolsky A.A., Evlanov I.A., Shulman S.S.
1994. Parasitic systems: an analysis of the structure and strategy that determines their resilience. Rep. ed.
S.S. Shulman. Ecological parasitology. Petrozavodsk, Karelian Research Center RAS, 3-43. (in Russian)].
Евланов И.А. 1992. Внутрипопуляционный полиморфизм плероцеркоидов Digramma interrupta (Cestoda,
Ligulidae) и его роль в функционировании паразитарной системы. Журнал общей биологии 53 (3):
368-372. [Evlanov I.A. 1992. Intrapopulation polymorphism of plerocercoids Digramma interrupta (Ces-
toda, Ligulidae) and its role in the functioning of the parasitic system. Zhurnal obshchey biologii 53 (3):
368-372. (in Russian)].
Иешко Е.П., Аникиева Л.В. 1980. Полиморфизм Proteocephalus exiguus - массового паразита сиговых рыб.
Паразитология 14 (5): 422-426. [Ieshko E.P., Anikieva L.V. 1980. Polymorphism of Proteocephalus
exiguus, a mass parasite of whitefish. Parazitologiya 14 (5): 422-426. (in Russian)].
Ильмаст Н.В., Сендек Д.С., Титов С.Ф., Абрамов С.А., Зуйкова Е.И., Бочкарев Н.А. 2016. К вопросу о
дифференциации экологических форм/ подвидов сига Coregonus lavaretus озера Каменное. Ученые
записки Петрозаводского гос. университета. Общая биология 4 (157): 42-53. [Ilmast N.V., Sendek
D.S., Titov S.F., Abramov S.A., Zuikova E.I., Bochkarev N.A. 2016. On the issue of differentiation of
ecological forms/subspecies of the whitefish Coregonus lavaretus of Lake Kamennoye. Scientific notes of
the Petrozavodsk state. University. General biology 4 (157): 42-53. (in Russian)].
104
Ильмаст Н.В., Стерлигова О.П. 2012. Экологическая характеристика водоема, не затронутого влиянием
деятельности человека (оз. Каменное). Рыбное население. Под общей ред. Н.Н. Немовой и др.
Биота северных озер в условиях антропогенного воздействия. Петрозаводск, изд-во Карельский
научный центр РАН, 21-26. [Ilmast N.V., Sterligova O.P. 2012. Ecological characteristics of a water
body not affected by human activities (Lake Kamennoe). fish population. Under the general editorship.
N.N. Nemovoi et al. Biota of northern lakes under anthropogenic impact. Petrozavodsk, publishing house
Karelian Scientific Center of the Russian Academy of Sciences, 21-26. (in Russian)].
Лакин Г.Ф. 1990. Биометрия. М., Высшая школа, 352 с. [Lakin G.F. 1990. Biometrics. M., Higher School,
352 pр. (in Russian)].
Майр Э. 1974. Популяции, виды и эволюция. Пер. с англ. М., Мир, 460 с. [Mayr E. 1970. Populations, spe-
cies, and evolution. Cambridge, Harvard University Press, 453 pp.].
Первозванский В.Я. 1986. Рыбы водоемов района Костомукшского железорудного месторождения (экология,
воспроизводство, использование). Петрозаводск: Карелия. 216 с. [Pervozvansky V.Ya. 1986. Fishes
of reservoirs of the area of the Kostomuksha iron ore deposit (ecology, reproduction, use). Petrozavodsk,
Karelia, 216 pр. (in Russian)].
Потапова О.И. 1978. Крупная ряпушка Coregonus albula L. Л.: Наука. 132 с. [Potapova O.I. 1978. Large
vendace Coregonus albula L. L., Nauka, 132 pр. (in Russian)].
Пугачев О.Н. 1988. Феногеографический анализ Dactylogyrus phoxini Malewitzkaja, 1949. В сб.: С.С. Шуль-
ман, Е.П. Иешко (ред.). Эколого-популяционный анализ паразито-хозяинных отношений. Петроза-
водск, Изд-во КарНЦ РАН, 103-109. [Pugachev O.N. 1988. Phenogeographical analysis of Dactylogyrus
phoxini Malewitzkaja, 1949. In: S.S. Shulman, E.P. Ieshko (eds). Ecological-population analysis of host-
parasite relations. Petrozavodsk, KarRC RAS, 103-109. (in Russian)].
Решетников Ю.С. 1980. Экология и систематика сиговых рыб. М., Наука, 300 с. [Reshetnikov Yu.S. 1980.
Ecology and systematics of whitefishes. M., Nauka, 300 pр. (in Russian)].
Рыбы в заповедниках России. 2010. (Под ред. Ю.С. Решетникова). Т. 1. М., Товарищество научных из-
даний КМК, 627 с. [Fish in the reserves of Russia. 2010. (Edited by Yu.S. Reshetnikov). T. 1. M.: KMK
Scientific Press Ltd., 627 pр. (in Russian)].
Северцов А.С. 1990. Направленность эволюции. М., Изд-во МГУ, 272 с. [Severtsov A.S. 1990. Direction of
evolution. M., Moscow State University, 272 pр. (in Russian)].
Северцoв А.С., Сурова Г.С., 1981. Индивидуальная изменчивость нормы реакции и адаптация популяции.
Журнал общей биологии 42 (2): 181-192. [Severtsov A.S., Surova G.S. 1981. Individual variability in
reaction rate and population adaptation. Zhurnal obshchey biologii 42 (2): 181-192. (in Russian)].
Тимофеев-Ресовский Н.В., Воронцов Н.Н., Яблоков А.В. 1977. Краткий очерк теории эволюции. М.., Наука,
297 с. [Timofeev-Resovsky N.V., Vorontsov N.N., Yablokov A.V. 1977. Brief outline of the theory of
evolution. M., Nauka, 297 pp. (in Russian)].
Фортунато М.Э. 1987. Выделение неметрических вариаций и характеристика некоторых группировок Dac-
tylogyrus vastator Nyb., 1924 (Monogenea), паразита карповых рыб. Труды Зоологического института
АН СССР 161: 51-62. [Fortunato M.E. 1987. Isolation of nonmetric variations and characterization of
some groups of Dactylogyrus vastator Nyb., 1924 (Monogenea), a parasite of cyprinids. Proceedings of
the Zoological Institute of the Academy of Sciences of the USSR, 161: 51-62. (in Russian)].
105
Фрезе В.И. 1965. Протеоцефаляты - ленточные гельминты рыб, амфибий и рептилий. М., Наука, 538 с.
[Frese V.I. 1965. Proteocephalates - tapeworms of fish, amphibians and reptiles. M., Nauka, 538 pp. (in
Russian)].
Шишкин М.А. 1988. Эволюция как эпигенетический процесс. В кн.: Макридин В.П., Меннер В.В. (ред.)
Современная палеонтология. Т. 2. М., Недра, 142-168. [Shishkin M.A. 1988. Evolution as an epigenetic
process. In: Makridin V.P., Menner V.V. Modern paleontology. T.2. M., Nedra, 142-168. (in Russian)].
Шмальгаузен И.И. 1982. Организм как целое в индивидуальном и историческом развитии Избранные труды.
М., Наука, 383 с. [Shmalgauzen I.I. 1982. Organism as a whole in individual and historical development
Selected Works. M., Nauka, 383 pp. (in Russian)].
Шмальгаузен И.И. 1983. Пути и закономерности эволюционного процесса. Избранные труды. Отв. ред.
М.С. Гиляров. М., Наука, 360 с. [Shmalgauzen I.I. 1983. Ways and laws of the evolutionary process.
Selected works. Rep. ed. M.S. Gilyarov. M., Nauka, 360 pp. (in Russian)].
Шульман С.С. 1958. Специфичность паразитов рыб. В сб.: Полянский Ю.И. (отв. ред.). Основные про-
блемы паразитологии рыб. Л., Изд-во ЛГУ, 109-121. [Shulman S.S. 1958. Specificity of fish parasites.
In Polyansky Yu.I. (ed.) Main problems of fish parasitology. L., Leningrad State University, 109-121.
(in Russian)].
Шульман-Альбова Р.Е. 1952. К вопросу об изменчивости дигенетического сосальщика рыб Podocotyle
atomon (Rud.) Odhner. Ученые записки Ленинградского университета, серия биологическая 141 (28):
110-126. [Shulman-Albova R.E. 1952. On the question of the variability of the digenetic fish fluke Podo-
cotyle atomon (Rud.) Odhner. Uchenyye zapiski Leningradskogo universiteta, seriya biologicheskaya 141
(28): 110-126. (in Russian)].
Hammer Ø., Harper D.A.T., Ryan P.D. 2001. PAST: Paleontological Statistics Software Package for Education
and Data Analysis. Palaeontologia Electronica V. 4 (issue1): 9 pp.
Hanzelova V., Snabel V., Spakulova M., Kralova I., Fagerholm H-P. 1995. A comparative study of the fish parasites
Proteocephalus exiguus and P. percae (Cestoda: Proteocephalidae): morphology, isoenzymes and karyotype.
Canadian Journal of Zoology 73 (7): 1191-1198.
Král’ová I. 1996. A total DNA characterization in Proteocephalus exiguus and P. percae (Cestoda: Proteoce-
phalidae): random amplified polymorphic DNA and hybridization techniques. Parasitology Research 82
(8): 668-671.
Král’ová I.,·Spakulová M. 1996. Intraspecific variability of Proteocephalus exiguus La Rue, 1911 (Cestoda:
Proteocephalidae) as studied by the random amplified polymorphic DNA method. Parasitology Research
82 (6): 542-545.
Scholz T., Hanzelova V. 1998. Tapeworms of the genus Proteocephalus Weinland, 1858 (Cestoda: Proteocephali-
dae), parasites of fishes in Europe. Praha, 118 pр.
106
PHENOTYPIC DIVERSITY OF POPULATION GROUPINGS
OF PROTEOCEPHALUS LONGICOLLIS (ZEDER 1800)
(CESTODA: PROTEOCEPHALIDAE),
A PARASITE OF THE WHITEFISH COREGONUS LAVARETUS (L.)
L. V. Anikieva , E. P. Ieshko
Keywords: population, structure, phenotypic diversity, common whitefish Coregonus
lavaretus, cestode Proteocephalus longicollis
SUMMARY
The population diversity of the cestode Proteocephalus longicollis population in whitefish Core-
gonus lavaretus from Lake Kamennoye (Kostomukshsky State Nature Reserve, northern Karelia)
was studied. Variability of groupings identified in the P. longicollis population according to discrete
parameters of attachment and trophic-reproductive features is characterized. The diversity, similarity,
and distinctions of the morphometric parameters and the contributions of specific groupings to the
phenotypic diversity are assessed. The P. longicollis population is represented mostly by the phe-
notype with a clavate scolex, which is easily differentiated from the nucleiform and the lanceolate
phenotypes by larger scolex dimensions and lower variability. We conclude that an essential factor
behind the phenotypic diversity and structure of the helminth population is the ecological type of
host’s behavior, which determines the niche occupied by the population. The adaptive response of
the P. longicollis population to change of the host species is manifested in a modified frequency
distribution of phenotypes with different attachment features.
107