ПАРАЗИТОЛОГИЯ, 2022, том 56, № 2, с. 108-125.
УДК 576.895
Гельминтофауна утиных
северо-запада россии и эстонии
© 2022 г. А. А. Виноградоваa, b, *, В. В. Скворцовb
a Зоологический институт РАН,
Университетская наб., 1, Санкт-Петербург, 199034 Россия
b РГПУ им. А.И. Герцена, факультет биологии,
наб. р. Мойки, 48б, Санкт-Петербург, 191186 Россия
⁕e-mail: gennadyeva@yandex.ru
Поступила в редакцию 20.01.2022 г.
После доработки 23.02.2022 г.
Принята к публикации 26.02.2022 г.
На территории Северо-Запада России (разные районы Ленинградской, Калининградской и
Псковской областей и Лоухского района Карелии) и Эстонии (Калласте) проведены исследо-
вания гельминтофауны речных уток. Всего обнаружено 43 вида гельминтов, принадлежащих
к Trematoda, Cestoda, Acanthocephala и Nematoda. Выявленные различия в видовом составе
гельминтов уток в разных районах и в разные сезоны в основном объясняются различиями
в доступности для них кормовых объектов - беспозвоночных, в некоторых случаях даже рыб,
которые служат промежуточными хозяевами гельминтов. Большинство из гельминтов, обнару-
женных в кишечниках уток, связаны своими жизненными циклами с пресными водами. Только
на побережье Белого моря гельминтофауна кряквы состоит исключительно из «морских» видов,
заражение которыми происходит при питании птиц на литорали во время отлива.
Ключевые слова: Anatinae, Trematoda, Cestoda, Acanthocephala, Nematoda
DOI: 10.31857/S0031184722020028, EDN: FESNNG
Бóльшая часть исследований гельминтофауны уток приходится на вторую поло-
вину XX века. Среди них выделяются монографии и статьи Быховской-Павловской
(1962), Скрябина (1951, 1953), Смогоржевской (1976), Спасской (1966), Хохловой
(1986), Мак Дональда (Key to trematodes reported in waterfowl. 1981) и Лапажа (Lapage,
1961). В этих работах приведены материалы по основным видам гельминтов, встреча-
ющимся у водоплавающих с различных территорий. Большинство из этих гельминтов
108
паразитирует в органах пищеварительной системы, преимущественно в кишечнике.
Наиболее разнообразна фауна трематод и цестод, тогда как нематоды и скребни менее
богаты видами, хотя и обычны для уток.
Несмотря на давнюю историю исследования паразитов утиных, приходится кон-
статировать, что сведения по гельминтам уток, а также по промежуточным хозяевам
с территории Северо-Запада России в большей степени известны с территории
Карелии (Фролова, 1975; Яковлева и др., 2012, 2018; Lebedeva et al., 2015, 2017),
а в других районах фрагментарны. Между тем, эта территория представляет суще-
ственный интерес, поскольку через нее проходит Восточно-Атлантический пролетный
путь (Davidson, Stroud, 2001), по которому весной идет массовая миграция водопла-
вающих с мест зимовок в Африке и Европе к местам гнездовий на Северо-Востоке
Европы и в Западной Сибири, а осенью - в обратном направлении. Некоторые из
этих мигрантов останавливаются для отдыха на территории Северо-Запада России и
сопредельных стран, а часть из них остается здесь для гнездования и линьки (Носков
и др., 2016). Соответственно, в состав сообществ гельминтов уток этого региона могут
быть включены виды, циркулирующие на его территории, а также виды, приносимые
утками с мест зимовок («южные» виды по классификации Догеля (1962)), северных
гнездований («северные» виды) и приобретаемые в ходе весенних и осенних миграций
(«миграционные» виды). Недостаточность информации по составу гельминтов уток
в обсуждаемом регионе, а также по путям реализации их жизненных циклов побу-
дило нас выполнить данное исследование. Цель исследования - определение состава
сообществ гельминтов у ряда видов уток и оценка возможности их трансмиссии
в некоторых районах Ленинградской, Псковской и Калининградской областей, а также
Эстонии.
Материал и методика
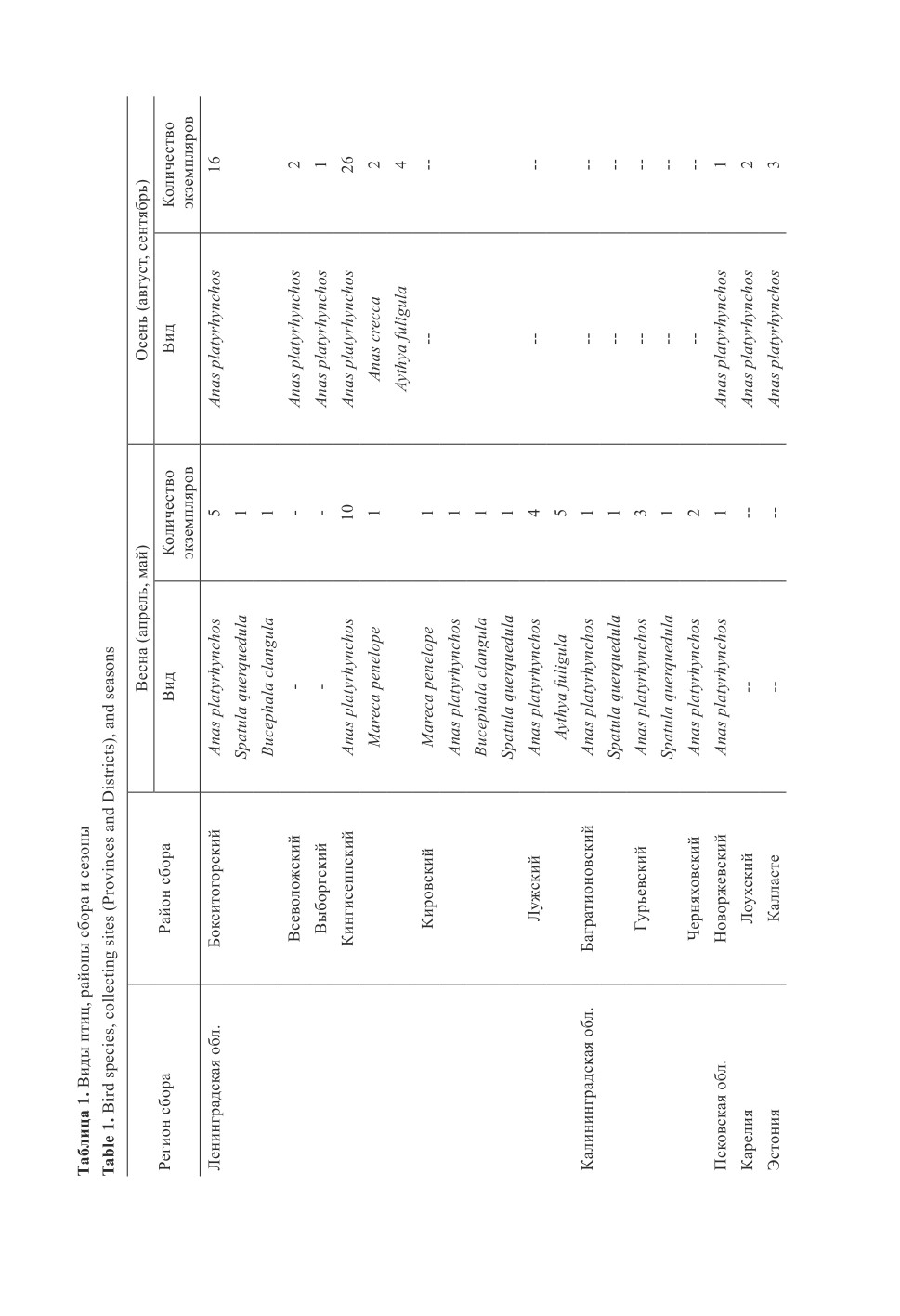
Гельминтологический материал собран от 99 экз. уток, добытых во время весенних и осен-
них охот с 2010 по 2019 гг. в разных районах Северо-Запада России, а также в восточной
части Эстонии (сроки и районы приведены в табл. 1). Основную часть обследованных птиц
составили благородные утки: кряква обыкновенная (Anas platyrhynchos) 78 экз., чирок-свистунок
(A. crecca) 2 экз., свиязь (Mareca penelope) 2 экз., чирок-трескунок (Spatula querquedula) 4 экз.
Нырковые утки были представлены только двумя видами: хохлатая чернеть (Aytya fuligula)
11 экз., гоголь обыкновенный (Bucephala clangula) 2 экз. Вскрытие птиц производили не позд-
нее суток с момента добычи охотниками в период открытия охот в регионах.
У всех птиц извлекали печень и кишечник и сразу их замораживали (примерно - 18°С).
У трех крякв сохранился желчный пузырь. В лаборатории эти органы размораживали и под-
вергали паразитологическому обследованию в соответствии с рекомендациями Быховской-
Павловской (1985) и Дубининой (1971). Обнаруженных плоских червей и скребней фиксировали
в 70% этаноле, а нематод - в 4% горячем формалине (примерно 90°С).
109
Изготовление тотальных препаратов проводили по стандартной методике. Для окраски гель-
минтов использовали различные красители: квасцовый кармин (для трематод), гематоксилин
Эрлиха (для цестод), гематоксилин Бемера (для скребней). После обезвоживания в спиртах
восходящей концентрации объекты заключали в монтирующую среду (Tissue-Mount™ Mounting
Medium, Sakura). Нематод помещали в смесь глицерина и воды (1:1) для просветления, посте-
пенно добавляя глицерин в раствор, делая его более концентрированным, по методике, пред-
ложенной Рыссом (Ryss, 2017).
Видовую идентификацию обнаруженных гельминтов проводили по оригинальным работам
и определителям «Key to trematodes reported in Waterfowl» (McDonald, 1981); «Keys to the
Trematoda» Vol. 1 (Gibson et al., 2002); «Цестоды птиц СССР. Гименолепидиды» (Спасская,
1966); «Акантоцефалы наземных позвоночных фауны СССР» (Хохлова, 1986).
Расчёт интенсивности инвазии (ИИ), экстенсивности инвазии (ЭИ) и доверительных ин-
тервалов (ДИ) средних значений для 95% уровня значимости проводили с использованием
программы Quantitative Parasitology (QP3). Расчет точного критерия Фишера выполнен в про-
грамме Microsoft Excel 2013.
Результаты
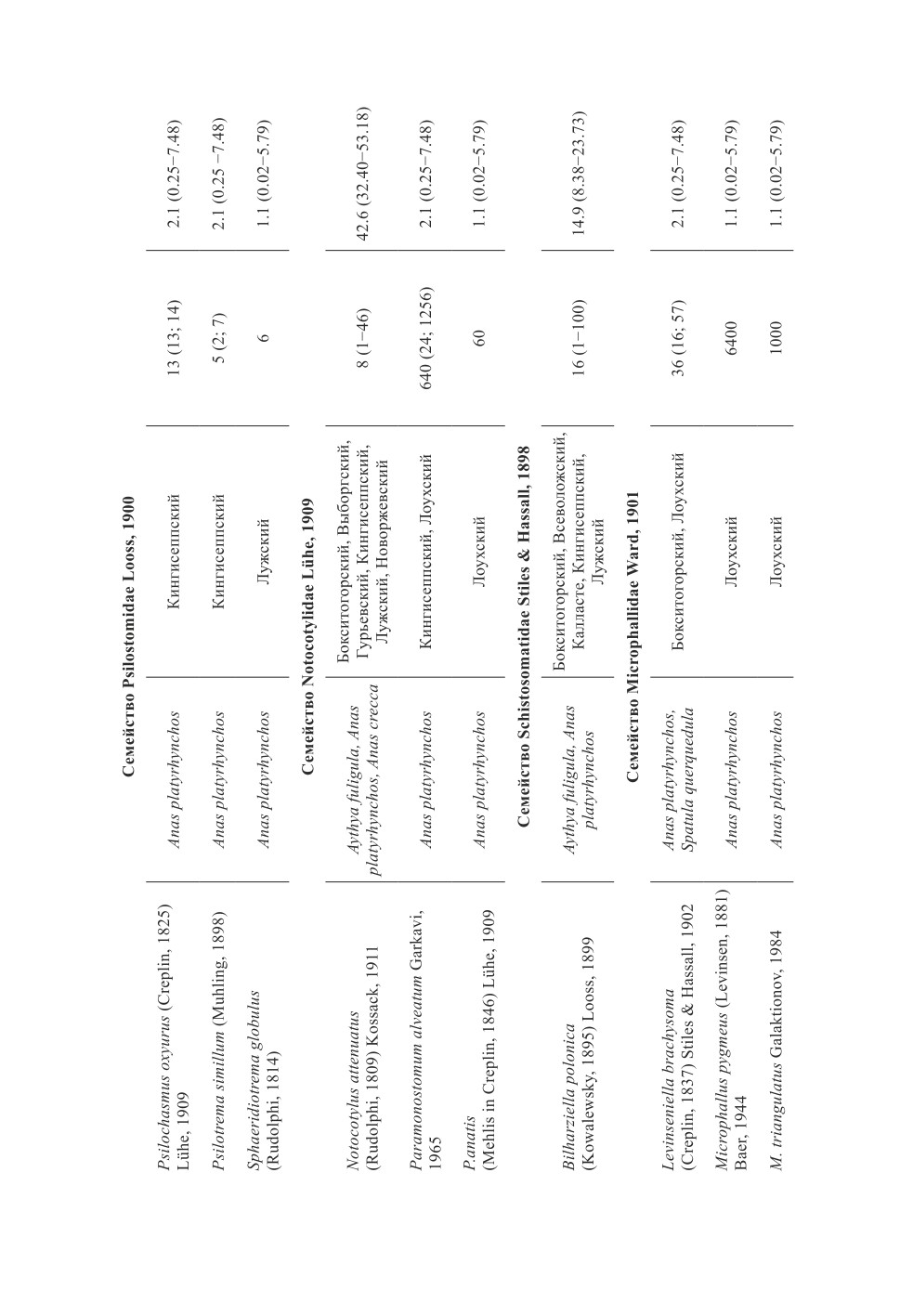
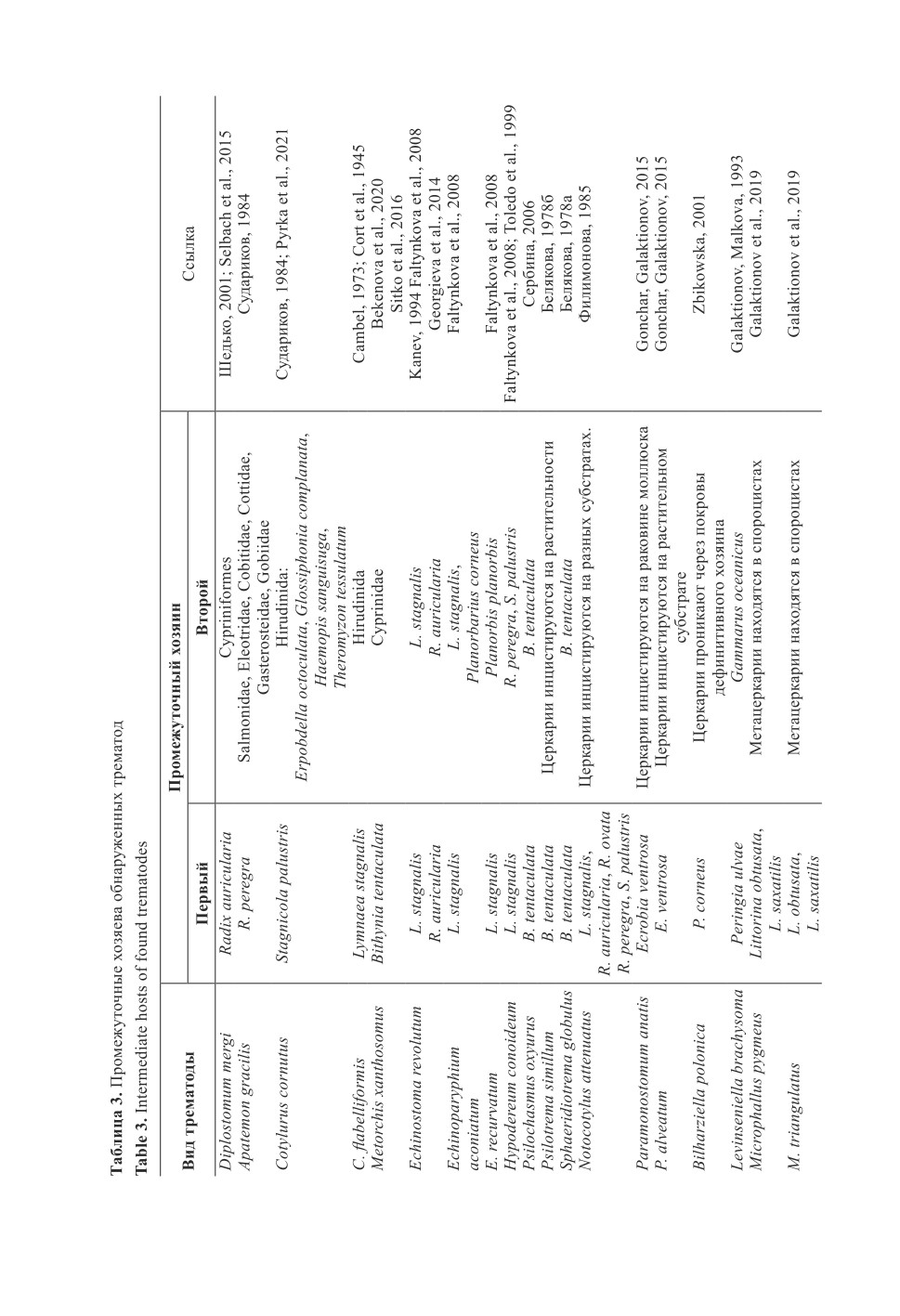
В обследованных утках в общей сложности обнаружены 43 вида гельминтов (табл. 2).
В кровеносных сосудах и протоках печени отмечены только два вида трематод:
Bilharziella polonica и Metorchis xanthosomus. Остальные гельминты локализовались
в кишечнике. Желчные пузыри были изучены только у трёх экземпляров кряквы -
гельминты в них отсутствовали.
Практически все обнаруженные гельминты, кроме Retinometra macracanthos и
Sobolevicanthus gracilis, выявлены у кряквы обыкновенной (табл. 2). Только у ныр-
ковых уток отмечен R. macracanthos. И у нырковых, и у благородных уток зареги-
стрированы B. polonica, Cotylurus cornutus, Echinoparyphium aconiatum, Notocotylus
attenuatus, Cloacotaenia megalops и Polymorphus minutus.
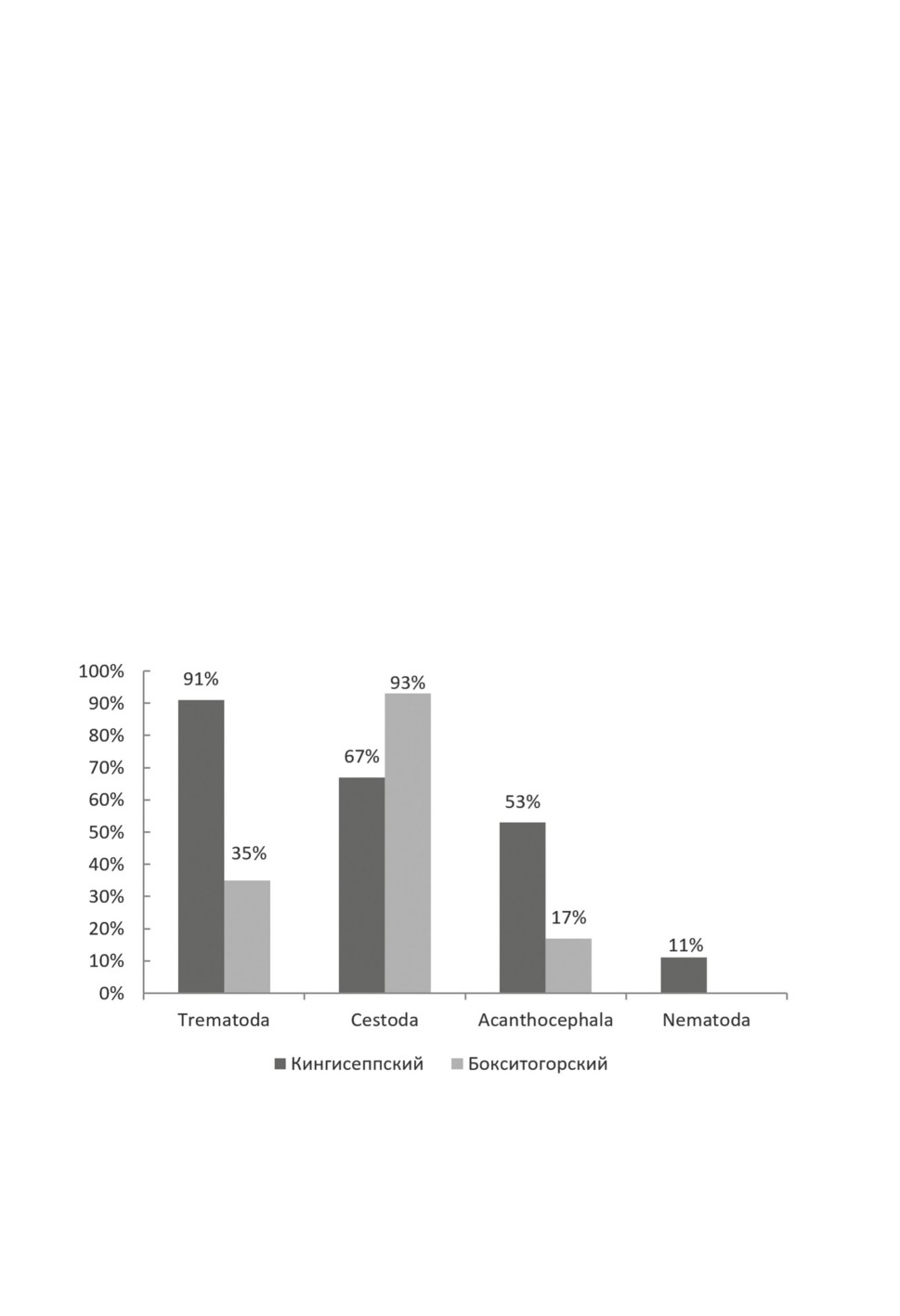
Наибольшее количество изученных птиц приходится на Кингисеппский (45 экз.)
и Бокситогорский (23 экз.) районы Ленинградской обл. В сборах из уток Кингисепп-
ского р-на выявлено наибольшее число видов трематод, а у утиных в Бокситогорском
р-не наибольшее число видов приходилось на цестод (рис. 1).
У крякв из Бокситогорского р-на отмечены трематоды Bilharziella polonica, Echi-
nostoma revolutum, Echinoparyphium recurvatum, Hypodereum conoideum, Notocotylus
attenuates. Также у одного чирка-трескунка зарегистрированы E. recurvatum и Levin-
seniella brachysoma. Фауна трематод уток Кингисеппского р-на более разнообразна.
В ее составе присутствуют те же виды, что и в Бокситогорском р-не. Кроме них,
у крякв также отмечены Apatemon gracilis, Cotylurus cornutus, Cotylurus flabelliformis,
Echinoparyphium aconiatum, Paramonostomum anatis, Psilochasmus oxyurus, Psilotrema
simillimum. Diplostomum mergi встречен у одной особи свиязи.
111
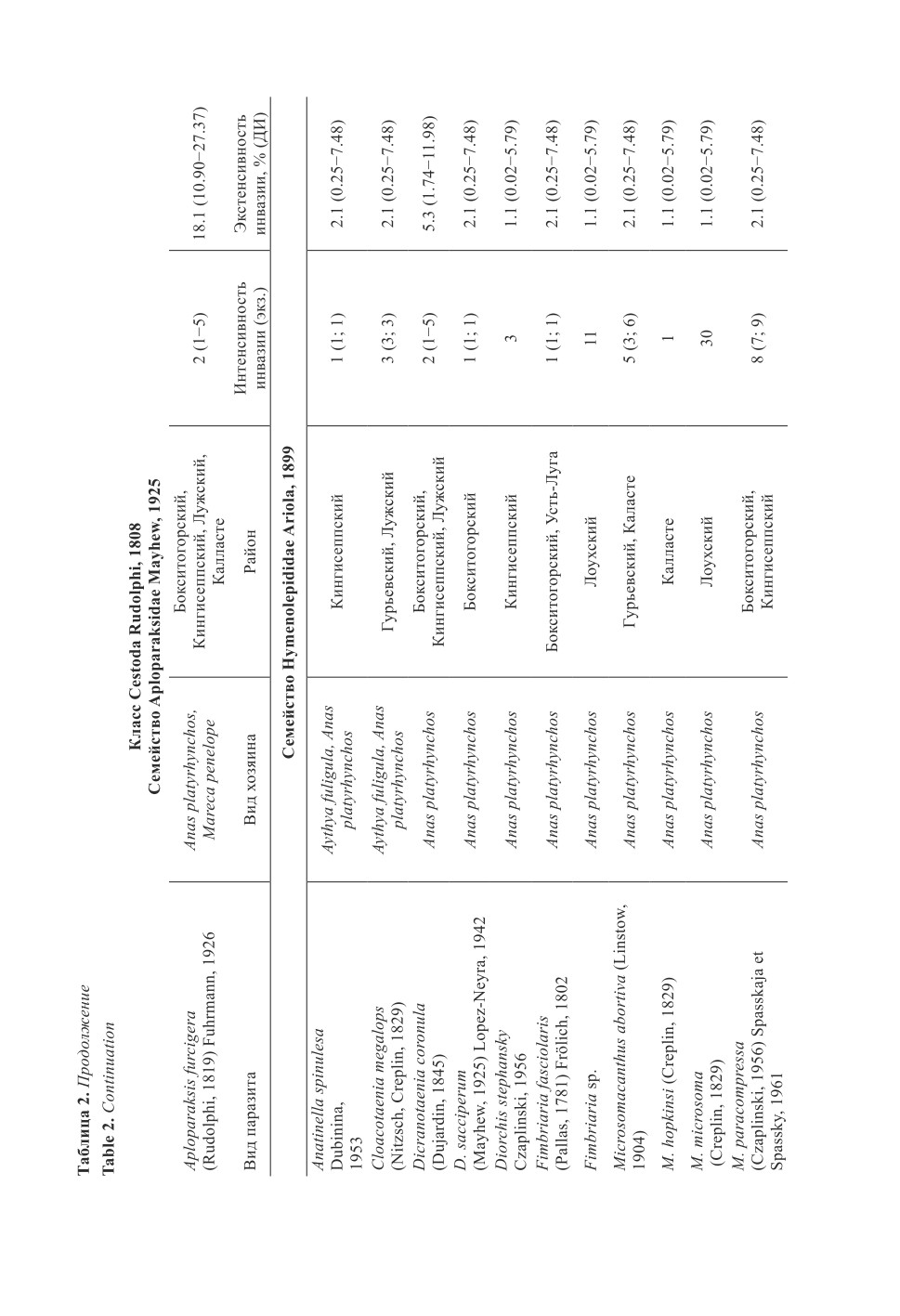
Цестоды утиных Кингисеппского р-на представлены 9 видами, из них у кряквы
обыкновенной обнаружены: Anatinella spinulesa, Aploparaksis furcigera, Dicranotaenia
coronula, Diorchis stephansky, Fimbriaria fasciolaris, Microsomacanthus paracompressa,
Retinometra venusta. Только у хохлатой чернети выявлен R. macrocanthos и у свистунка -
Sobolevicanthus gracilis. В выборке из Бокситогорского р-на присутствует 12 видов
цестод. Из них Dicranotaenia sacciperum, Parabisaccanthes philactes, Parabisaccan-
thes kazachstanica, Sobolevivanthus octacantha и Wardoides nyroca отмечаются только
у крякв в Бокситогорском р-не. Остальные черви, за исключением Anatinella spinulesa,
Diorchis stephansky и R. venusta, присутствуют в обеих выборках.
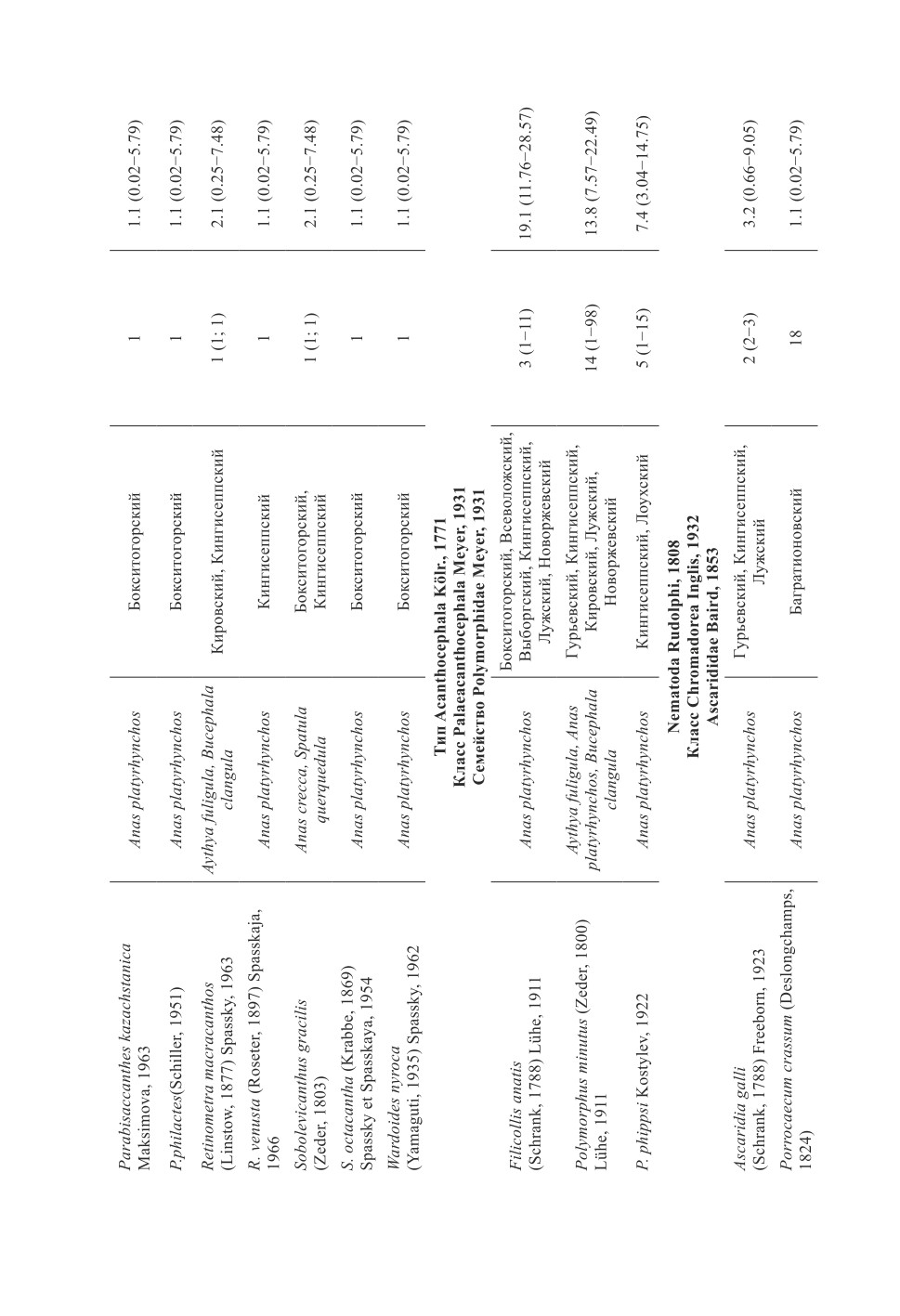
У уток выявлено три вида скребней в Кингисеппском р-не: Filicollis anatis,
Polymorphus phippsi и P. minutus. В Бокситогорском р-не зарегистрирован только
F. anatis.
Нематоды были представлены только одним видом Ascaridia galli в Кингисеппском
р-не, в Бокситогорском р-не представители этих паразитов не обнаружены.
Использование точного критерия Фишера для сравнения экстенсивности инвазии
для общих видов позволило выявить статистически значимые различия только для
Bilharziella polonica (p=0.037) и Notocotylus attenuatus (p=0.000). Зараженность этими
паразитами оказалась выше в Кингисеппском р-не.
В Калининградской и Псковской областях, Калласте (Эстония) и в других ука-
занных нами районах Ленинградской обл. зарегистрированы единичные находки
гельминтов, не отмеченные в других районах. В сборах из Калласте обнаружен
Microsomacanthus hopkinsi. Только в Лужском р-не отмечен вид Sphaeridiotrema
globulus. В выборках из Калласте и Гурьевского р-на Калининградской обл. встречался
M. abortiva. Только в Багратионовском р-не отмечен Porrocaecum crassum.
Для всех районов, кроме Лоухского, характерны общие виды: Aploparaksis
furcigera, Bilharziella polonica, Filicollis anatis, Hypodereum conoideum, Notocotylus
attenuatus, Polymorphus minutus. Наиболее часто встречаемым видом оказался N. at-
tenuatus (табл. 2).
В Лоухском р-не на побережье Белого моря были добыты две особи кряквы обык-
новенной. У них были выявлены трематоды Levinseniella brachysoma, Microphallus
pygmeus, M. triangulatus, Paramonostomum anatis, P. alveatum, цестоды Microsomacan-
thus microsoma, Fimbriaria sp. и скребень Polymorphus phippsi. Среди этих гельминтов
присутствуют только «морские» виды, реализующие свои жизненные циклы в при-
брежных экосистемах Белого моря. Это отличает Лоухский р-н от всех остальных,
так как в других районах преобладают гельминты, в трансмиссии которых участвуют
пресноводные беспозвоночные, а также некоторые виды рыб (табл. 3).
116
Обсуждение
Полученные результаты гельминтологических вскрытий уток дополняют имею-
щуюся информацию по гельминтофауне утиных на севере Палеарктики. В том числе
впервые получены сведения по гельминтам уток Ленинградской обл.
В целом, видовой состав обнаруженных нами гельминтов совпадает с видовыми
составами, приводимыми в исследованиях по паразитическим червям утиных Чехии,
Польши (Kavetska et al., 2008) и Украины (Syrota et al., 2018). По сравнению с этими
работами, в нашем материале встречается меньше видов. Отсутствуют трематоды че-
тырех родов: Parastrigea, Opisthorchis, Plagiorchis и Echinochasmus, мариты которых
регистрировались в вышеперечисленных регионах у разных видов благородных уток.
Трематоды рода Echinochasmus характерны для поганок, но в материале некоторых
авторов (Kavetska et al., 2008; Lapage, 1961) они отмечены и для кряквы. Заражение
утиных трематодами родов Echinochasmus и Opisthorchis могут происходить при
поедании различных видов рыб, которые служат для них вторыми промежуточными
хозяевами; Parastrigea - через головастиков, Plagiorchis - через личинки водных бес-
позвоночных (поденки, стрекозы) и ракообразных (Heneberg et al., 2018). Среди цестод
не были выявлены представители семейства Dilepididae. Так же гораздо меньше видов
родов Diorchis, Microsomacanthus, Sobolevicanthus, которые могут встречаться у ути-
ных (Спасская, 1966; Толкачева, 1991). Мы предполагаем, что описанные выше раз-
личия в составе гельминтофауны утиных Северо-Запада России и европейских стран,
где проводились аналогичные исследования (Kavetska et al., 2008; Lapage, 1961), могут
быть связаны с малой зараженностью промежуточных хозяев, сезоном, характером и
спектром питания. Трематоды из материала, полученного в нашем исследовании, реа-
лизуют свои жизненные циклы (табл. 3), преимущественно, через гастропод (первые
и вторые промежуточные хозяева) и пиявок (вторые промежуточные хозяева). Для
цестод ту же роль промежуточных хозяев играют люмбрикулиды (Бондаренко и др.,
2006) и остракоды (Доброхотова, 1985). Можно предположить, что на территории
Северо-Запада эти беспозвоночные составляют существенную долю в диете уток,
обеспечивая их заражение обнаруженными в исследовании видами гельминтов. Не
выявлены нами и трематоды родов Prostogonimus и Leucochloridiomorpha - паразиты
Фабрициевой сумки утиных. В нашем материале представлены только взрослые утки,
поэтому эти виды нами не обнаружены.
Благородные и нырковые утки отличаются по способу добывания пищи (Исаков,
1953; Носков и др., 2016). Такие виды как хохлатая чернеть и гоголь обыкновен-
ный ныряют в поисках корма, а кряква, свиязи и другие благородные утки ― нет.
Соответственно, последние могут захватывать корм только с неглубокой части при-
брежной зоны. Нырковые и благородные утки являются смешанноядными птицами.
118
У благородных уток преобладает корм растительного происхождения, а у нырков -
животного (Искова, 1970). Учитывая это, мы предположили, что гельминтофауна
нырковых и благородных уток может различаться. Однако при сравнении видово-
го состава гельминтов кряквы, чернети и гоголя в нашем материале существенных
различий обнаружить не удалось, что объясняется, скорее всего, небольшим числом
вскрытых нырков. И у благородных, и у нырковых уток в равной мере присутствовали
гельминты, которыми птицы могли заразиться при поедании промежуточных хозяев
или адолескарий трематод (N. attenuatus, P. alveatum, Ps. simillimum) и яиц (A. galli),
которые могут аккумулироваться на различных субстратах во внешней среде.
При сравнении состава фауны гельминтов из разных районов выявлены различия
между Кингисеппским, где большая часть утиных заражена трематодами, и Бокси-
тогорским, где большинство уток были заражены цестодами (рис. 1). Обследование
водоемов в местах добычи уток в Бокситогорском р-не показало практически полное
отсутствие в них гастропод (наши данные). Видимо, этим и объясняется небольшое
число видов трематод, зарегистрированных нами в утках из Бокситогорского р-на.
Следует отметить, что некоторыми паразитами, которых мы обнаружили у обследо-
ванных птиц, они заразились в местах зимовок или в миграционный период. На это
указывают находки Levinseniella brachysoma в чирке с весенней охоты в Боксито-
горского р-не, а также Paramonostomum anatis и Polymorphus phippsi в двух кряквах
с осенней охоты в Кингисеппском. В трансмиссии этих видов участвуют морские
беспозвоночные, поэтому реализация циклов в пресноводных экосистемах невозмож-
на. Пролетные же пути утиных захватывают Белое и Балтийское моря, в прибрежье
которых они останавливаются и питаются морскими беспозвоночными (Galaktionov
et al., 2019).
Сезонная доступность того или иного корма служит основной причиной, опреде-
ляющей состав сообществ гельминтов утиных. Об этом, в том числе, свидетельствуют
находки Porrocaecum crassum в Калининградской обл. Известны случаи, когда в поис-
ках мест для гнездования утки прилетают на поля, где при таянии снега образуются
временные водоемы (Мальчевский, Пукинский, 1983). В таких местах птицы могут
заразиться Por. crassum, питаясь дождевыми червями, которые служат для этой не-
матоды промежуточными хозяевами. Доступность для уток этого корма ранней весной
и определяет их заражение в этот период Por. crassum (Birova et al., 1990; Lebedeva
et al., 2017).
Анализируя состав гельминтов крякв, добытых в Лоухском р-не, можно предпо-
ложить, что благородные утки заражаются при поедании морских беспозвоночных
в момент обнажения литорали во время отлива. Видимо, в составе корма крякв при-
119
сутствуют бокоплавы, через которых происходит трансмиссия Levinseniella brachysoma
(Gammarus duebeni и G. oceanicus), Polymorphus phippsi (G. setosus) (Галактионов,
Атрашкевич, 2015), а также литоральные гастроподы, такие как Littorina spp., служа-
щие промежуточными хозяевами для Microphallus pygmaeus и M. triangulatus (Galak-
tionov et al., 2019). В предпочитаемых утками заиленных бухтах Белого моря обитают
моллюски-гидробииды Ecrobia ventrosa и Peringia ulvae, которые служат первыми
промежуточными хозяевами нотокотилидным трематодам Paramonostomum anatis и
P. alveatum, церкарии которых инцистируются на подводных субстратах почти сразу
после выхода из зараженных моллюсков и превращаются в инвазионных для птиц
метацеркарий (адолескарий) (Gonchar, Galaktionov, 2015). Эти данные отличаются от
аналогичных исследований в Карелии (Лебедева и др., 2015, 2017; Яковлева и др.,
2012, 2018). В материале этих авторов присутствовали гельминты, чьи жизненные ци-
клы реализуются только с участием пресноводных беспозвоночных и рыб. Очевидно,
в районах, приближенных к Белому морю, утки предпочитают питаться на литорали,
где кормовая база доступнее и обильнее, чем в обедненных северных озерах.
Рисунок 1. Зараженность основными группами гельминтов Кингисеппского
и Бокситогорского района.
Figure 1. Infestation of the main helminth groups in Kingisepp and Boksitogorsk Districts.
120
В целом, можно сказать, что фауна гельминтов утиных Северо-Запада России и
фауна стран Европы, расположенных в пределах Восточно-Атлантического мигра-
ционного пути (Чехия, Польша, Нидерланды), сходны. Скорее всего, это связано
с миграционной активностью утиных и сходством в видовом составе промежуточных
хозяев (беспозвоночных и рыб) в водоемах данных регионов. В районах, приближен-
ных к Белому морю, доминируют гельминты, реализующие свои жизненные циклы
с участием морских беспозвоночных. Однако большая часть обнаруженных в нашем
исследовании гельминтов осуществляют свои жизненные циклы в пресных водоемах.
Таким образом, пресноводные речные утки участвуют в трансмиссии не только «пре-
сноводных» гельминтов, но и «морских».
Благодарности
За предоставленный материал благодарим Г.Л. Атаева, Л.О. Пяйта, О.Е. Белкина,
К.В. Авво, Г.С. Слюсарева.
Выражаем благодарность за помощь в обработке материала К.В. Регель - сотруд-
нику лаборатории экологии гельминтов ИБПС ДВО РАН, а также коллективу лабора-
тории экспериментальной зоологии РГПУ им. А.И. Герцена и коллективу лаборатории
по изучению паразитических червей и протистов ЗИН РАН.
Работа выполнена при финансовой поддержке РФФИ (грант № 19-0400384)
и в рамках темы Государственных заданий № 122031100260-0.
Список литературы
Белякова Ю.В. 1978а. Новые данные по циклу развития Sphaeridiotrema globulus Rud., 1819 (Trematoda:
Psilostomidae). Жизненные циклы, экология и морфология гельминтов животных Казахстана. Алма-
Ата, Наука, 40-47. [Belyakova Y.V. 1978. Novye dannye po cyklu razvitiya Sphaeridiotrema globulus
Rud., 1819 (Trematoda: Psilostomidae). Zhiznennye cykly, ecologia i morphologiya gelmintov zhivotnykh
Kazakhstana. Alma-Ata, Nauka, 40-47. (in Russian)].
Белякова Ю.В. 1978б. Жизненный цикл Psilotrema simillimum (Muhling, 1898) (Trematoda: Psilostomidae).
Паразитология 12 (1): 62-67. [Belyakova Y.V. 1978. The life cycle of Psilotrema simillimum (Muhling,
1898) (Trematoda: Psilostomidae). Parazitologiya 12 (1): 62-67. (in Russian)].
Бондаренко С.К. 2006. Аплопараксиды диких и домашних птиц. Основы цестодологии. Т. 14. /Бондаренко
С.К., Контримавичус В.Л.; отв. ред. С.О. Мовсесян. М., Наука, 443 с. [Bondarenko S.K. 2006. Aplo-
paraksidae of wild and domesticated birds. Osnovy cestodologiy. Vol. 14. /Bondarenko S.K., Kontrimavi-
chus V.L. Resp. Ed. S.O. Movsesyan. M., Nauka, 443 рр. (in Russian)].
Быховская-Павловская И.Е. 1962. Трематоды птиц СССР: эколого-географический обзор. М.-Л., Изд-во
АН СССР, 321 с. [Bykhovskaya-Pavlovskaya I.E. 1962. Trematody ptits SSSR: ecologo-geographicheskiy
obzor. M.-L., Izd-vo AN SSSR, 321 рр. (in Russian)].
Быховская-Павловская И.Е. 1985. Паразиты рыб: руководство по изучению. JI., Наука, 123 с. [ Bykhovskaya-
Pavlovskaya I.E. 1985. Parazity ryb: rukovodstvo po izucheniyu. L., Nauka, 123 рр. (in Russian)].
121
Галактионов К.В., Атрашкевич Г.И. 2015. Специфика циркуляции паразитов морских птиц в высокой
Арктике на примере паразитарной системы скребня Polymorphus phippsi (Palaeacanthocephala, Poly-
morphidae). Паразитология 49 (6): 393-411. [Galaktionov К.V., Atrashkevich G.I. 2015. Patterns in cir-
culation and transmission of marine bird parasites in high arctic: a case of acanthocephalan Рolymorphus
phippsi (Palaecanthocephala, Polymorphidae). Parazitologiya 49 (6): 393-411. (in Russian)].
Доброхотова О.В. 1985. Биоценотические связи остракод с гименолепидидами водно-болотных птиц в
водоемах Казахстана. Гельминты животных в экосистемах Казахстана. Алма-Ата, Наука, 22-45. [Do-
brokhotova O.V. 1985. Biocenothicheskie svyazi s hymenolepididami vodno-bolotnykh ptits v vodoemakh
Kazakhstana. Helminty zhivotnykh v ecosistemakh Kazakhstana. Alma-Ata, Nauka, 22-45. (in Russian)].
Догель В.А. 1962. Общая паразитология. Л., ЛГУ, 464 с. [Dogel V.A. 1962. Obshchaya parasitologia. L.,
LGU, 464 рр. (in Russian)].
Дубинина М.Н. 1971. Паразитологическое исследование птиц: методы паразитологических исследований.
Вып. 4, Л., Наука, 139 с. [Dubinina M.N. 1971. Parasitologicheskoe issledovanie ptits: metody parasito-
logicheskikh issledovaniy. Vyp. 4, L., Nauka, 139 рр. (in Russian)].
Исаков Ю.А. 1953. Птицы Советского Союза. Под ред. Дементьева Г.П., Гладкова Н.А.. Т. 4. М., Советская
наука, 362-518. [Isakov Y.A. 1953. Ptitsy Sovetskogo Soyuza. Pod red. Dementeva G.P., Gladkova N.A.
T. 4. M., Sovetskaya nauka, 362-518. (in Russian)].
Искова Н.И. 1970. Зависимость трематодофауны водно-болотных птиц от питания. Вестник зоологии 3:
61-68. [Iskova N.I. 1970. Dependence of trematode fauna of water-bog birds on nutrition. Vestnik zoologii
3: 61-68. (in Russian)].
Мальчевский А.С., Пукинский Ю.Б. 1983. Птицы Ленинградской области и сопредельных территорий.
История, биология, охрана, Т. 1. Л., ЛГУ, 480 с. [Malchevskiy A.S., Pukinskyi Y.B. 1983. Ptitsy Lenin-
gradskoi oblasti I sopredelnykh territoriy. Istoria, biologia, okhrana, T. 1. L., LGU, 480 рр. (in Russian)].
Носков Г.А., Рымкевич Т.А., Гагинская А.Р. 2016. Миграции птиц Северо-Запада России. Неворобьиные.
СПб., «Профессионал», 656 с. [Noskov G.A., Rymkevich T.A., Gaginskaya A.R. 2016. Migration of birds
of Northwest Russia. Non-passerines. St.Petersburg, «Professional», 656 рр. (in Russian)].
Сербина Е.А. 2006. Распространение трематод семейства Psilostomatidae Odhner, 1913 в Западной Сибири.
Сибирский экологический журнал 4: 409-418. [Serbina E.A. 2006. Prevalence of Trematodas Family
Psilostomatidae Odhner, 1913 in the South of West Siberia. Siberian Journal of Ecology 4 (13): 409-418.
(in Russian)].
Скрябин К.И. 1951. Трематоды животных и человека. Основы трематодологии. Т. 5. Schistosomata. М.,
Изд-во АН СССР, 624 с. [Skryabin K.I. 1951. Trematodes of animals and man. Fundamentals of Trema-
todology. Schistosomata, Vol. 5, M., AN SSSR, 624 рр. (in Russian)].
Скрябин К.И. 1953. Трематоды животных и человека. Основы трематодологии. Т. 8. Notocotylidae. М.,
АН СССР, 618 с. [Skryabin K.I. 1953. Trematodes of animals and man. Fundamentals of Trematodology.
Notocotylidae, Vol. 8, M., AN SSSR, 618 рр. (in Russian)].
Смогоржевская Л.А. 1976. Гельминты водоплавающих и болотных птиц фауны Украины. Киев, Наукова
Думка, 416 с. [Smogorzhevskaya L.A. 1976. Helminty vodoplavayushchikh I bolotnikh ptits fauny Ukrainy.
Kiev, Naukova dumka, 416 рр. (in Russian)].
Спасская Л.П. 1966. Цестоды птиц СССР. Гименолепидиды. М., Наука, 698 с. [Spasskaya L.P. 1966. Cestody
ptits SSSR. Hymenolepididy. M., Nauka, 698 рр. (in Russian)].
122
Судариков В.Е. 1984. Трематоды фауны СССР. Стригеиды. М., Наука, 168 с. [Sudarikov V.E. 1984. Trematoy
fauny SSSR. Strigeidy. M., Nauka, 168 рр. (in Russian)]
Толкачева Л.М. 1991. Цестоды фауны СССР: Род Diorchis. М., Наука, 181. [Tolkacheva L.M. 1991. Cestody
fauny SSSR. Rod Diorchis. M., Nauka, 181 рр. (in Russian)].
Фролова Е.Н. 1975. Личинки трематод в моллюсках озер Южной Карелии. Л.: Наука, 182 с. [Frolova E.N.
1975. Lichinki trematode v molluskakh ozer Yuzhnoi Karelii. L., Nauka, 182 рр. (in Russian)].
Хохлова И.Г. 1986. Акантоцефалы наземных позвоночных фауны СССР. М., Наука, 277. [Hohlova I.G. 1986.
Akantocephaly nazemnykh pozvonochnykh fauny SSSR. M., Nauka, 277 рр. (in Russian)].
Шедько М.Б. 2001.Распространение метацеркарий трематод семейства Diplostomidae (in Russian)в пресно-
водных экосистемах Приморья. Чтения памяти Владимира Яковлевича Леванидова. Вып. 1: 96-104.
[Schedko M.B. 2001 Rasprostranenie metacerkarii trematod semeistva Diplostomidae v presnovodnikh eko-
sistemakh Primorya. Chtenia pamyati Vladimira Yakovlevicha Levanidova. Vyp. 1: 96-104 (in Russian)].
Яковлева Г.А., Лебедева Д.И., Иешко Е.П. 2012. Фауна трематод водоплавающих птиц Карелии. Паразито-
логия 46 (2): 98-110. [Yakovleva G.A., Lebedeva D.I., Ieshko E.P. 2012. Trematodes fauna of waterfowl
birds in Karelia. Parazitologiya 46 (2): 98-110. (in Russian)].
Яковлева Г.А., Лебедева Д.И., Иешко Е.П. 2018. Акантоцефалы - паразиты кряквы (Anas platyrhynchos
Linnaeus, 1758) акватории Ладожского озера. Вестник охотоведения 15 (4): 335-338. [Yakovleva G.A.,
Lebedeva D.I., Ieshko E.P. 2018. Acantocephala of mallard (Anas platyrhynchos L.) from of the lake
Ladoga region. Vestnik okhotovedeniya 15 (4): 335-338. (in Russian)].
Andrew S., Bell L., Sommerville L., Gibson D. 2002. Multivariate analyses of morphometrical features from
Apatemon gracilis (Rudolphi, 1819) Szidat, 1928 and A. annuligerum (v. Nordmann, 1832) (Digenea:
Bekenova A., Smagulova A., Katokhin A., Borovikov S., Kiyan V. 2020. Molecular differential diagnosis between
Opisthorchis felineus and Metorchis bilis. Advances in animal and veterinary sciences 8(3): 27-32. doi.
org/10.17582/journal.aavs/2020/8.s3.27.32
Birova V., Spakulova M., Macko J.K. 1990. Seasonal dynamics of the invasive cycle of nematodes and acan-
thocephalans in the wild (Anas platyrhynchos L.) and domestic duck (Anas platyrhynchos f. dom.). Hel-
mintologia 27: 291-301.
Campbell R.A. 1973. Studies on the biology of the life cycle of Cotylurus flabelliformis (Trematoda: Strigeidae).
Transactions of the American Microscopical Society 92 (4): 629-640.
Cort W.W., Brackett S., Olivier L., Nolf L.O. 1945. Influence of Larval Trematode Infections in snails on their
second intermediate host relations to the strigeid trematode, Cotylurus flabelliformis (Faust, 1917). The
Journal of Parasitology 31 (1): 61-78.
Davidson N.C., Stroud D.A. 2001. Waterbird Flyways and History of International Cooperation for Waterbird
Conservation. An Atlas of Wader Populations in Africa and Western Eurasia. Delany S., Scott D. doi.
org/10.1007/978-94-007-6172-8_104-1
Faltynkova A., Nasincova V., Kablaskova L. 2008. Larval trematodes (Digenea) of planorbid snails (Gastropoda:
Pulmonata) in Central Europe: a survey of species and key to their identification. Systematic Parasitology
69 (3): 155-178. doi.org/10.1007/s11230-007-9127-1
Galaktionov K.V., Malkova I.I. 1993. Development of the alimentary tract during morphogenesis of the metacercar-
iae of Levinseniella brachysoma. Journal of Helminthology 67: 87-94. doi.org/10.1017/S0022149X00012943
123
Galaktionov K.V., Nikolaev K.E., Aristov D.A., Levakin I.A., Kozminsky E.V. 2019. Parasites on the edge: pat-
terns of trematode transmission in the Arctic intertidal at the Pechora Sea (South-Eastern Barents Sea).
Polar Biology 42: 1719-1737. doi.org/ 10.1007/s00300-018-2413-3
Georgieva S., Faltynkova A., Brown R., Blasco-Costa I., Soldanova M., Sitko J., Scholz T., Kostadinova A. 2014.
Echinostoma 'revolutum' (Digenea: Echinostomatidae) species complex revisited: species delimitation based
on novel molecular and morphological data gathered in Europe. Parasites & Vectors 7 (1): 520 2-18. doi.
org/ 10.1186/PREACCEPT-1112350941442199
Gonchar A., Galaktionov K.V. 2015. Substratum preferences in two notocotylid (Digenea, Notocotylidae) cercariae
from Hydrobia ventrosa at theWhite Sea. Journal of Sea Research. doi.org/ 10.1016/j.seares.2015.07.006
Heneberg P., Sitko J., Tesinskya M., Rzad I., Bizosa J. 2018. Central European Strigeidae Railliet, 1919 (Trema-
toda: Strigeidida): Molecular and comparative morphological analysis suggests the reclassification of Para-
strigea robusta Szidat, 1928 into Strigea Abildgaard, 1790. Parasitology International 67 (6): 688-701.
doi.org/10.1016/j.parint.2018.07.003
Kanev I. 1994. Life-cycle, delimitation and redescription of Echinostoma revolutum (Froelich, 1802) (Trematoda:
Echinostomatidae). Systematic Parasitology 28: 125-144.
Kavetska K., Rząd I., Sitko J. 2008. Taxonomic structure of Digenea in wild ducks (Anatinae) from West Pomera-
nia. Wiadomości Parazytologiczne 54 (2): 131-136.
Key to trematodes reported in waterfowl. 1981. M. McDonald. University of California Libraries. 168 рр.
Keys to the Trematoda. 2002.Gibson D.I., Jones A., Bray R.A. Vol. 1. 544 рр.
Lapage G. 1961. A list of the parasitic Protozoa, Helminths and Arthropoda recorded from species of the Family
Anatidae (Ducks, Geese and Swans). Parasitology 51 (1-2): 1-109. doi.org/10.1017/s0031182000068517
Lebedeva D.I., Yakovleva G.A., Ieshko E.P. 2015. Nematodes in the mallard (Anas platyrhynchos Linnaeus,
1758) and the common goldeneye (Bucephala clangula Linnaeus, 1758) (Anatidae) from Northern Europe.
Parasitology Research 114: 3935-3937. doi.org/ 10.1007/s00436-015-4697-3
Lebedeva D.I., Yakovleva G.A., Ieshko E.P. 2017. Nematodes of the Eurasian wigeon (Anas penelope) and the
common teal (A. сrecca) in northwestern Russia. Паразитология 51 (3): 206-212.
Pyrka E., Kanarek G., Zaleśny G., Hildebrand J. 2021. Leeches as the intermediate host for strigeid trematodes:
genetic diversity and taxonomy of the genera Australapatemon Sudarikov, 1959 and Cotylurus Szidat,
1928. Parasites & Vectors 14 (44): 1-17. doi.org/ 10.1186/s13071-020-04538-9
Ryss A.Y. 2017. A Simple Express Technique to Process Nematodes for Collection Slide Mounts. Journal of
Nematology 49 (1): 27-32. doi.org/ 10.21307/jofnem-2017-043
Selbach C., Soldanova M., Georgieva S., Kostadinova A., Sures B. 2015. Integrative taxonomic approach to the
cryptic diversity of Diplostomum spp. in lymnaeid snails from Europe with a focus on the ‘Diplostomum
mergi’ species complex. Parasites & Vectors 8: 300. doi.org/ 10.1186/s13071-015-0904-4
Sitko J., Bizos J., Sherrard-Smith E., David W., Stanton G., Komorova P., Heneberg P. 2016. Integrative tax-
onomy of European parasitic flatworms of the genus Metorchis Looss, 1899 (Trematoda: Opisthorchiidae).
Parasitology International 65 (3): 258-267. doi.org/ 10.1016/j.parint.2016.01.011
Syrota Y.Y., Greben O.B., Poluda A.M., Maleha O.M., Lisitsyna O.I., Kornyushin V.V. 2018. Helminths of the
Mallard, Anas platyrhynchos (Aves Anatidae), in Ukraine: Analysis of the Diversity in Mixed Forest Zone
and the Black Sea Region. Vestnik Zoologii 52 (4): 267-278. doi.org/ 10.2478/vzoo-2018-0028
124
Toledo R., Muñoz-Antoli C., Esteban J-G. 1999. Metacercarial encystment of Hypoderaeum conoideum (Bloch,
1782) (Trematoda: Echinostomatidae) in several freshwater gastropod species. Folia Parasitologica 73:
157-158.
Zbikowska E. 2001. Bird schistosomes in Lymnaea stagnalis (L.) and Planorbarius corneus (L.) from selected
lakes in Poland. Helminthologia 38: 244.
Duck HELMINTS
of the NorthWestern Russia and Estonia
A. A. Vinogradova, V. Skvortsov
Keywords: Anantinae, Trematoda, Cestoda, Acanthocephala, Nematoda
SUMMARY
The helminth fauna of dabbling and diving ducks (Anatinae) was studied in the territory of
Northwestern Russia (Leningrad, Kaliningrad, and Pskov Provinces; Loukhi District of Karelia) and
Estonia (Kallaste). In total, 43 helminth species belonging to Trematoda, Cestoda, Acanthocephala,
and Nematoda were found. The revealed differences in the species composition of duck helminths
in different regions and different seasons are explained mainly by differences in the availability of
trophic objects - invertebrates and fish, which serve as intermediate hosts for helminths. Most of the
helminths recorded in ducks by their life cycles are associated with fresh waters. Only in the White
Sea coast the helminth fauna of the mallard duck consists exclusively of "marine" species that can
be explained by feeding on the intertidal zone.
125