ПАРАЗИТОЛОГИЯ, 2022, том 56, № 2, с. 126-138.
УДК [591.69-9+591.531.213]:639.1.081.12
К ВОПРОСУ О ПОТЕРЕ ЭКТОПАРАЗИТОВ
МЕЛКИХ МЛЕКОПИТАЮЩИХ
ПРИ ОТЛОВЕ ЛОВУШКАМИ ГЕРО
© 2022 г. Н. А. Кочерова a, Л. А. Беспятова a, С. В. Бугмырин a,*
a Институт биологии КарНЦ РАН,
Пушкинская ул., д. 11, Петрозаводск, 185910 Россия
* e-mail: sbugmyr@mail.ru
Поступила в редакцию 11.02.2022 г.
После доработки 08.03.2022 г.
Принята к публикации 11.03.2022 г.
Для разных систематических групп эктопаразитов проведена сравнительная оценка времени
покидания мелких млекопитающих при их отлове в стандартные ловушки типа Геро. Ловушки,
установленные в линию, были оборудованы датчиками для автоматической регистрации вре-
мени поимки животного. В целом, наши результаты подтверждают основное правило о том,
что эктопаразиты со временем покидают тело погибшего хозяина. Наиболее существенное
и быстрое снижение зараженности животного после гибели отмечено для Ixodes persulcatus
Schulze, 1930. Блохи и паразитические гамазовые клещи покидают хозяина после его гибели
в первые 2-4 ч, вши - через 9-13 ч. Для I. (Exopalpiger) trianguliceps Birula, 1895 не показано
существенного снижения численности в течение длительного времени.
Ключевые слова: время поимки, блохи, вши, гамазовые клещи, иксодовые клещи, Ixodes
persulcatus, Ixodes trianguliceps
DOI: 10.31857/S003118472202003X, EDN: FFBHYC
Мелкие млекопитающие являются резервуарными хозяевами многих опасных для
человека и животных природно-очаговых инфекций, поддержание и распространение
которых, в первую очередь, обеспечивается массовым паразитированием различных
видов эктопаразитов.
При изучении биологии мелких млекопитающих традиционно широко использу-
ется метод отловов животных ловушками Геро (Карасева, Телицына, 1996; Шефтель,
2018). Этот метод прост и универсален, его можно применять для решения широко-
126
го круга задач, в том числе для изучения таксономического состава и численности
паразитов этих млекопитающих. Одним из недостатков метода является сложность
отслеживания времени поимки животного. Эктопаразиты после гибели хозяина по-
кидают его, а отсутствие сведений о длительности пребывания животных в ловушке
затрудняет интерпретацию результатов по их зараженности, в том числе и видами,
имеющими важное эпидемиологическое значение, например, иксодовыми клещами.
Личинкам и нимфам иксодовых клещей посвящено основное число работ, оцени-
вающих их потерю после гибели хозяина (Наумов, 1958; Шилова и др., 1958; Колонин
и др., 1977; De Pelsmaeker et al., 2020). Исследований, касающихся паразитов других
систематических групп, немного (Борисова и др., 1982; Ельшин, 1992). Основные под-
ходы к решению этого вопроса базируются как на результатах непосредственных на-
блюдений за сползанием клещей (Наумов, 1958), так и на сравнении степени заражен-
ности животных, добытых разными методами отлова (Колонин и др., 1977). Главным
образом, полученные результаты свидетельствуют о покидании эктопаразитами тела
погибшего животного хозяина спустя какой-то промежуток времени. Вместе с тем,
в некоторых работах при сравнении мелких млекопитающих, добытых живоловками
и ловушками Геро, не наблюдали различия в их зараженности иксодовыми клещами
(Шилова и др., 1958; De Pelsmaeker et al., 2020).
Целью нашего исследования была сравнительная оценка времени покидания
у разных систематических групп эктопаразитов мелких млекопитающих при их от-
лове в стандартные ловушки типа Геро. Основное внимание было уделено двум видам
иксодовых клещей - Ixodes persulcatus Schulze, 1930 и I. (Exopalpiger) trianguliceps
Birula, 1895, имеющим важное эпидемиологическое значение и преобладающим по
численности в сборах эктопаразитов в данном районе. В основу анализа положены
данные, полученные с помощью линии ловушек с автоматической регистрацией вре-
мени поимки животного.
МАТЕРИАЛ И МЕТОДИКА
Материал по паразитам мелких млекопитающих был собран в период с 2013 по 2021 гг.
в районе Гомсельского научного стационара ИБ КарНЦ РАН (62.068° с. ш., 33.961° в. д.). Район
исследования расположен в 60 км севернее г. Петрозаводск на границе Южного и Средне-
карельского зоогеографических подрайонов. Отловы мелких млекопитающих производились
в разнотипных биотопах с помощью ловушко-линий. Давилки Геро выставляли по 25-50 штук
на расстояние 3-5 м друг от друга на несколько суток. Линии проверяли один раз (обычно
в 8 утра) или два раза (около 8 и 18 ч) в сутки. Погибших животных помещали в отдельные
мешочки. Осмотр зверьков, сбор и определение эктопаразитов проведены по общепринятым
методикам (Брегетова, 1956; Высоцкая, Кирьянова, 1970; Беспятова, Бугмырин, 2012).
127
Для определения точного времени гибели животного была использована линия ловушек
с автоматической регистрацией времени срабатывания (Яковлев и др., 2020; Бугмырин и др.,
2021). Устройство разработано для широко распространенных и недорогих ловушек Геро, где
фиксация и запоминание времени поимки животных на всех ловушках линии осуществляется
с использованием стандартного микроконтроллера. В исходном заряженном состоянии ловушки
Геро цепь замкнута. При срабатывании ловушки происходит размыкание цепи, что фиксируется
микроконтроллером и сохраняется на карту памяти с указанием номера ловушки и време-
ни ее срабатывания. Линия из 14 ловушек с регистрацией времени поимки была выставлена
в осиново-березовом травяном приручейном лесу. Описание данного биотопа приводилось ранее
(Беспятова и др., 2019). Линия работала в 2013-2021 гг. в июне, июле и августе по 3-4 сут
в течение одного учета; отработано около 1000 ловушко-суток. Всего в экспериментальную
линию было поймано 84 особи мелких млекопитающих: Neomys fodiens - 1, Sorex araneus - 28,
S. minutes - 1, S. isodon - 4, Myodes glareolus - 46, Microtus agrestis - 4 экз.
Сравнительную оценку вклада различных факторов (длительность нахождения животного
в ловушке, сезон, год, вид, пол и возраст хозяина) на численность эктопаразитов проводили
с помощью многофакторного дисперсионного анализа. Сила влияния (%) оценена как отноше-
ние факториальной суммы квадратов к общей сумме квадратов.
Для определения периода покидания паразитами животного после его гибели использовали
метод моделирования «точки изменения» (the change point analysis), который был разработан для
обнаружения разрывов в многомерных геохимических данных в скважине или образце керна
(Gallagher et al., 2011). Положение «точки» было оценено методом Монте-Карло с Марковскими
цепями, количество интераций составляло 100000. Максимальное количество точек изменения
выбрано 10 по умолчанию. Все вычисления и построение графиков реализованы в программе
PAST v4.05 (Hammer et al., 2001).
Для остальных животных, отловленных на других линиях, по внешним признакам дава-
ли субъективную оценку длительности их пребывания в ловушке - давно (например, были
частично поедены другими животными) и недавно (например, зверек был живым или снят
при вечерней проверке линий) попавшиеся. Соответствующую информацию регистрировали в
журнале. Всего было обследовано 931 экз. мелких млекопитающих двух отрядов: Eulipotyphla -
обыкновенная кутора (Neomys fodiens) 3 экз., обыкновенная бурозубка (Sorex araneus) 416,
малая бурозубка (S. minutus) 28, равнозубая бурозубка (S. isodon) 12, средняя бурозубка
(S. caecutiens) 6 экз. и Rodentia - рыжая полевка (Myodes glareolus) 443 экз., пашенная полевка
(Microtus agrestis) 18, лесная мышовка (Sicista betulina) 4 экз. Все животные были разнесены
по группам, для которых рассчитывали показатели относительной численности эктопаразитов:
встречаемость (ИВ), индекс обилия (ИО) и их доверительные интервалы. Расчеты индексов
и оценка значимости их различий (Fisher's Exact Test для ИВ и bootstrap ANOVA для ИО)
выполнены в программе Quantitative Parasitology (Reiczigel et al., 2019).
128
РЕЗУЛЬТАТЫ
Численность эктопаразитов на мелких млекопитающих,
оцененных по внешним признакам как «давно» или «недавно» попавшиеся
Все животные по внешним признакам были условно разделены на три группы:
недавно попались (или живые), давно попались и остальные (табл. 1). Межгрупповые
различия встречаемости и/или индекса обилия были получены для блох и иксодовых
клещей (p ≤ 0.05). Ожидаемо, что эти различия определяются более низкими показа-
телями заражения у группы давно попавшихся животных. Для личинок и нимф двух
видов клещей Ixodes trianguliceps и I. persulcatus рассчитаны ИВ и ИО. Значимые
групповые различия (p ≤ 0.05) были получены для I. trianguliceps (табл. 1).
Численность эктопаразитов на мелких млекопитающих, отловленных
ловушками с регистраторами точного времени поимки животного
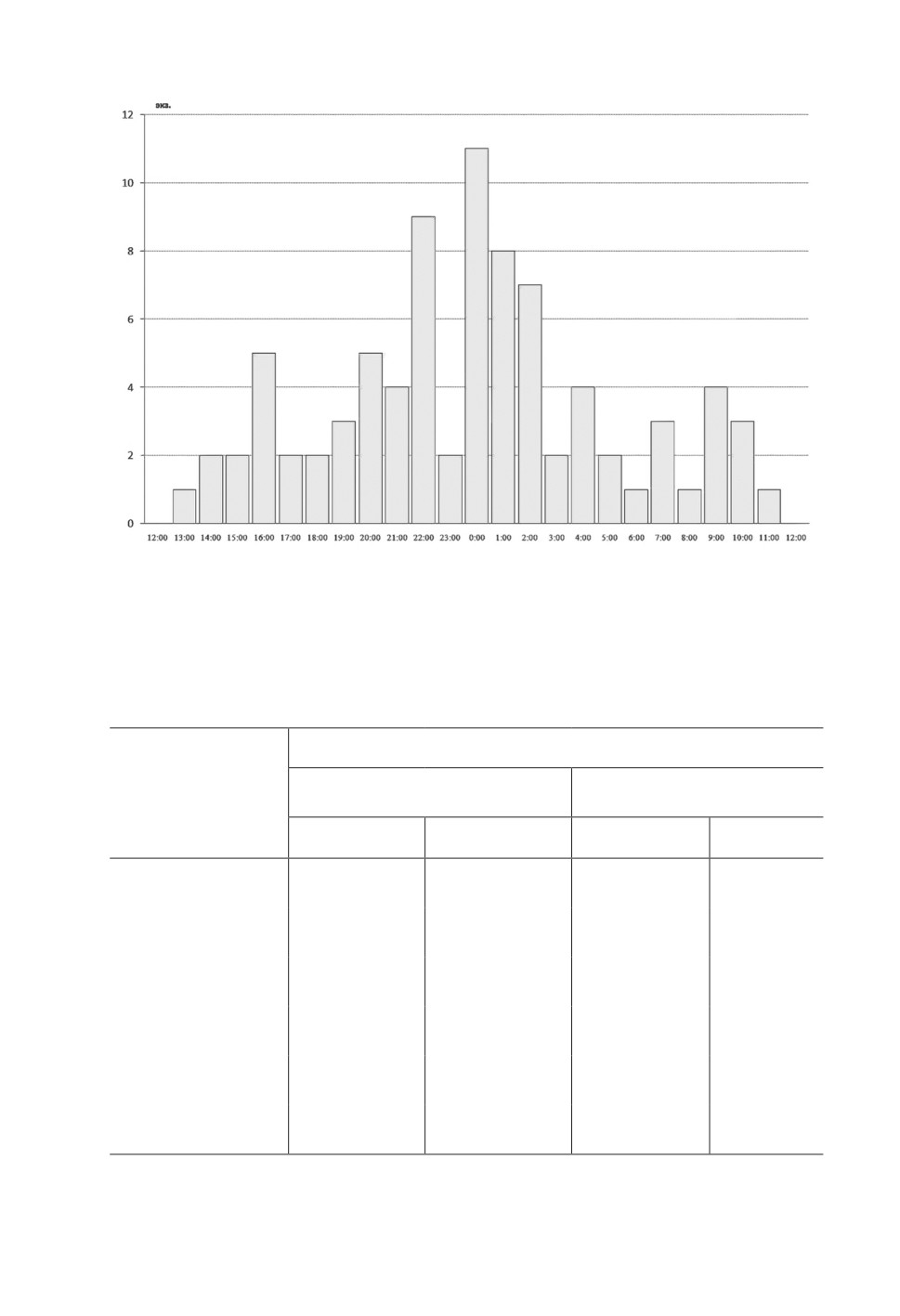
По данным, обобщенным за весь период работы линии, наибольшее число мелких
млекопитающих было поймано в интервале с 22.00 до 03.00 (рис. 1). Сравнение дан-
ных по численности эктопаразитов у двух групп мелких млекопитающих, различаю-
щихся по длительности пребывания в ловушке (до 8 ч и более 11 ч после поимки),
не выявило существенных различий в их зараженности (табл. 2).
По результатам дисперсионного анализа сила влияния фактора времени - длитель-
ности пребывания животного в ловушке - на численность паразитов варьировала от
9 (вши) до 24% (гамазовые и иксодовые клещи) (рис. 2).
По результатам моделирования «точки изменения» наиболее существенное сниже-
ние зараженности в первые часы после гибели животного отмечено для I. persulcatus
(рис. 3A, 3B). Для блох также характерно покидание хозяина после его поимки в те-
чение первых часов. Численность вшей заметно снижается через 9-13 ч после гибели
животного. Для гамазовых клещей отмечены две «точки перехода»: первая (через
2 ч) определяется снижением численности, вторая (через 18-21 ч) - увеличением
численности (рис. 3). Для I. trianguliceps не наблюдается существенных отклонений
от средней численности в течение длительного времени пребывания хозяев в ловушке.
ОБСУЖДЕНИЕ
Численность паразитов - величина стохастическая, вместе с тем эта стохастич-
ность во многом является результатом воздействия комплекса факторов, например
условий биотопа, сезона года, характеристик хозяина (вид, пол, возраст) и др. Фактор
времени, в сравнении с вышеперечисленными, вносит существенный вклад в наблю-
даемые вариации численности эктопаразитов (рис. 2). В большей мере это относится
к блохам, иксодовым и гамазовым клещам.
129
Таблица 1. Численность эктопаразитов на мелких млекопитающих в зависимости от
длительности их пребывания в ловушках (субъективная оценка состояния животного
по внешним признакам)
Table 1. Infestation of small mammals differing in the duration of staying in traps
(evaluation of animals by external signs)
Недавно попались
Давно попались
Остальные
Систематическая
(32)
(80)
(819)
группа паразитов
ИВ, %
ИО
ИВ, %
ИО
ИВ, %
ИО
Вши
12.5
1.2
13.8
0.4
17.7
0.9
[3.5-29]
[0.1-5.1]
[7.1-23.3]
[0.2-0.8]
[15.1-20.5]
[0.7-1.1]
Блохи
37.5
1.1
17.5
0.3
38.6*
1
[21.1-56.3]
[0.6-1.9]
[9.9-27.6]
[0.1-0.5]
[35.2-42]
[0.9-1.2]
Гамазовые клещи
40.6
0.9
37.5
1.0
42.6
1.1
[23.7-59.4]
[0.5-1.4]
[26.9-49]
[0.6-2.0]
[39.2-46.1]
[0.9-1.3]
Иксодовые клещи
40.6
4.4
22.5
2.3
37.6*
1.3*
[23.7-59.4]
[1.9-8.5]
[13.9-33.2]
[1.4-4.4]
[34.3-41]
[1.1-1.6]
I. persulcatus
15.6
2.8
15
0.4
19.7
0.7
[5.3-32.8]
[0.7-7]
[8-24.7]
[0.2-1.0]
[17-22.5]
[0.6-1]
личинки
15.6
2.5
12.5
0.4
15.8
0.6
[0.5-32.8]
[0.5-6.8]
[6.2-21.8]
[0.2-1.0]
[13.3-18.4]
[0.5-0.8]
нимфы
9.4
0.3
2.5
0.03
8.1
0.15
[0.2-25]
[0.03-1]
[0.3-8.7]
[0-0.06]
[6.3-10.1]
[0.11-0.21]
I. trianguliceps
37.5
1.6
7.5
0.09
25.5*
0.6
[21.1-56.3]
[0.7-4.3]
[2.8-5.6]
[0.03-0.2]
[22.6-28.7]
[0.5-0.7]
личинки
18.8
0.8
2.5
0.03
17*
0.4*
[7.2-36.4]
[0.1-3.5]
[0.3-8.7]
[0-0.06]
[14.5-19.7]
[0.3-0.5]
нимфы
28.1
0.7
3.8
0.05
11*
0.2
[13.7-46.7]
[0.3-1.5]
[0.8-10.6]
[0-0.13]
[8.9-13.3]
[0.13-0.22]
Примечания. В круглых скобках - число исследованных особей. Для каждого таксона
приведены значения индекса встречаемости (ИВ), индекса обилия (ИО) и их доверительные
интервалы (в квадратных скобках); * - Значимые (p < 0.05) межгрупповые различия
ИВ и ИО.
130
Рисунок 1. Распределение числа поимок мелких млекопитающих
в течение суток (обобщенные данные).
Figure 1. Distribution of small mammal captures during the day (generalized data).
Таблица 2. Численность эктопаразитов на мелких млекопитающих,
в зависимости от длительности их пребывания в ловушках
Table 2. Infestation of small mammals differing in the duration of staying in traps
Время нахождения животного в ловушке
Систематическая
от 0 до 8 ч
от 11 до 23 ч
группа паразитов
(39)
(35)
ИВ, %
ИО
ИВ, %
ИО
Блохи
46.2
1.0
37.1
0.9
[30.1-62.8]
[0.6-1.6]
[20.2-59.4]
[0.5-1.7]
Вши
28.2
2.3
20
0.5
[15-44.9]
[1.1-5.5]
[8.4-36.9]
[0.1-1.8]
Гамазовые клещи
56.4
1.2
42.9
1.2
[39.6-72.2]
[0.7-1.9]
[26.3-60.6]
[0.7-2.1]
Иксодовые клещи
48.7
1.9
54.3
1.8
[32.4-65.2]
[1.1-3]
[36.6-71.2]
[1-3.3]
I. trianguliceps
35.9
0.95
28.6
1.2
[21.9-52.6]
[0.5-1.5]
[14.6-46.3]
[0.4-2.8]
I. persulcatus
23.1
0.9
31.4
0.5
[11.1-39.3]
[0.4-2.5]
[16.9-49.3]
[0.3-0.9]
Обозначения см. табл. 1.
131
Рисунок 2. Относительный вклад различных факторов в наблюдаемые вариации
численности основных систематических групп эктопаразитов мелких млекопитающих.
Figure 2. Relative contribution of various factors to the observed variations
in the counts of ectoparasites on small mammals:
1 - time after being caught in a snap trap, 2 - season, 3 - year, 4 - host species,
5 - host sex, 6 - host age, 7 - residual.
В нашем исследовании с помощью метода оценки резких изменений (Gallagher
et al., 2011) мы попытались определить время, после которого наблюдается наиболее
существенное снижения числа эктопаразитов различных систематических групп.
Значительное снижение численности отмечено для Ixodes persulcatus с наибольшей
вероятностью в первые часы после гибели животного (рис. 3). Полученные результаты
согласуются с данными прямых полевых наблюдений, где было показано, что около
40% личинок I. persulcatus покидает труп прокормителя в течение первых трех часов
после поимки (Наумов, 1958). По результатам лабораторных опытов сход с хозяина
питающихся личинок I. persulcatus начинали регистрировать через три часа после его
гибели, и на следующий день эксперимента прикрепившиеся личинки отсутствовали
(Nakao, Sato, 1996).
132
Рисунок 3. Изменение численности эктопаразитов мелких млекопитающих
в течение времени после поимки в ловушки:
А - изменение численности эктопаразитов разных систематических групп в течение
времени после гибели животного, B - вероятность наступления события (точки перехода) -
существенного изменения численности в течение времени для тех же групп эктопаразитов.
Figure 3. Changes in the number of ectoparasites of small mammals
over time after being caught in a snap trap (the change point analysis):
A - changes in the abundance of ectoparasites of different taxonomic groups over time
after the host’s death, B - probability of a tangible change in abundance over time
for the same groups of ectoparasites.
133
Для другого вида иксодовых клещей - специфичного паразита мелких млекопита-
ющих I. trianguliceps - мы не получили резкого снижения численности в течение дли-
тельного времени после гибели хозяина (рис. 3), хотя данные по давно и недавно по-
павшимся животным показывают значительные различия в их зараженности (табл. 1).
По-видимому, покидание I. trianguliceps животного растянуто во времени и не имеет
столь резкого характера как у I. persulcatus.
Гамазовые клещи, собранные с мелких млекопитающих в районе исследования,
имеют в своем составе паразитов (облигатных и факультативных), а также хищников и
схизофагов (Bespyatova, Bugmyrin, 2006). При длительном нахождении животных в ло-
вушке происходит постепенное снижение численности паразитических и значительное
возрастание свободноживущих видов. Этим можно объяснить некоторое увеличение
зараженности мелких млекопитающих гамазовыми клещами, которое наблюдается,
по нашим данным, спустя 18 ч после поимки животного (рис. 3).
Блохи - одни из самых активных эктопаразитов, поэтому было ожидаемо, что со
временем происходит снижение их численности (табл. 1). По нашим данным, наи-
более существенное падение численности блох у мелких млекопитающих происходит
в течение первых 4 ч после отлова (рис. 3). Ранее было показано (Борисова и др.,
1982), что у разных видов блох тенденции к схождению с трупа зверька выражены
по-разному (что справедливо и для паразитов других систематических групп). Так,
блохи Megabothris (Gebiella) rectangulatus (Wahlgren, 1903) начинают покидать живот-
ное через 10 мин после его гибели и через 12 ч полностью уходят в подстилку, за тот
же период времени только около 40% Amalareus peniciliger (Grube, 1851) покидают
своих хозяев (Борисова и др., 1982). Вши - постоянные безотрывные эктопаразиты-
гематофаги млекопитающих (Балашов, 2009). Относительный вклад фактора времени
в наблюдаемые вариации численности у вшей ниже по сравнению с аналогичным
показателем у других групп эктопаразитов мелких млекопитающих (рис. 2). Вши на-
чинают покидать тело хозяина в среднем через 9 ч после поимки животного (рис. 3).
Основным фактором, определяющим начало схода эктопаразитов с прокормителя,
является понижение температуры тела хозяина. Скорость этого понижения зависит как
от размера самого животного, так и от температуры окружающей среды. Следователь-
но, в разные сезоны проведения исследования темпы снижения числа эктопаразитов
при длительном нахождении животного в ловушке могут различаться. Ранее было
отмечено, что потери иксодовых клещей увеличивались с мая по август, что объясня-
лось ростом среднесуточной температуры окружающей среды (Колонин и др., 1977).
134
В целом, наши результаты подтверждают основное правило о том, что эктопара-
зиты со временем покидают тело погибшего хозяина. Часто при первичном осмотре
животного мы можем сделать заключение о длительности его пребывания в ловушке.
Сравнительный анализ наших данных показывает, что группа животных, которых мы
отнесли к категории «давно попавшиеся», имеет более низкие показатели заражен-
ности эктопаразитами (табл. 1). С другой стороны, если доля этих животных в общей
выборке низкая (в нашем случае около 10%), то ошибка, связанная с недоучетом
паразитов, не должна быть высокой.
Суточная активность мелких млекопитающих в районе исследования приходится
на вечерне-ночной период (с 22:00 до 03:00) вследствие чего значительное число от-
ловленных животных находилось в ловушках по 5-8 ч до паразитологического обсле-
дования. Соответственно данные по относительной численности эктопаразитов будут
занижены и в первую очередь за счет личинок и нимф I. persulcatus. Использование
вместо ловушек Геро живоловок не будет в полной мере компенсировать потерю иксо-
довых клещей (Шилова и др., 1958; De Pelsmaeker et al., 2020). Более частая проверка
линии (в том числе и ночная) позволит снизить потери эктопаразитов, но потребует
значительных усилий и, главное, регулярности (повторяемости) для корректного со-
поставления данных, например разных биотопов или периодов. При географической
(биотопической) характеристике или многолетнем мониторинге видового состава
и численности паразитов мелких млекопитающих можно игнорировать фактор дли-
тельности пребывания животного в ловушке, если, стандартизировать метод отлова
и кратность сбора материала. Вместе с тем, потери эктопаразитов следует учитывать
при построении прогностических моделей, в которых зараженность мелких млекопи-
тающих - важная составляющая расчетов.
БЛАГОДАРНОСТИ
Авторы выражают благодарность В.В. Яковлеву (КарНЦ РАН, Петрозаводск) за
техническое сопровождение проводимых научных исследований. Мы признательны
рецензентам за полезные комментарии к рукописи статьи.
Финансовое обеспечение осуществлялось из средств федерального бюдже-
та на выполнение государственного задания КарНЦ РАН (FMEN-2022-0005,
№ г. р. 122032100130-3).
135
СПИСОК ЛИТЕРАТУРЫ
Балашов Ю.С. 2009. Паразитизм клещей и насекомых на наземных позвоночных. СПб., Наука, 357 с.
[Balashov Yu.S. 2009. Acari and insect parasitism on terrestrial vertebrates. St. Petersburg, Nauka, 357
pp. (in Russian)].
Беспятова Л.А., Бугмырин С.В. 2012. Иксодовые клещи Карелии (распространение, экология, клещевые
инфекции). Петрозаводск, Карельский научный центр РАН, 100 с. [Bespyatova L.A., Bugmyrin S.V.
2012. Ixodid ticks of Karelia (expantion, ecology, the main tick-borne infect). Petrozavodsk, Karelian
Research Centre of RAS, 100 pp. (in Russian)].
Беспятова Л.А., Бугмырин С.В., Кутенков С.А., Никонорова И.А. 2019. Численность иксодовых клещей
(Acari: Ixodidae) на мелких млекопитающих в лесных биотопах среднетаежной подзоны Карелии.
Паразитология 53 (6): 463-473. [Bespyatova L.A., Bugmyrin S.V., Kutenkov S.A., Nikonorova I.A. 2019.
The abundance ixodid ticks (Acari: Ixodidae) on small mammals in forest biotopes of the middle taiga
subzone of Karelia. Parazitologiya 53 (6): 463-473. (in Russian)].
Борисова В.И., Назарова И.В., Орехов Е.М. 1982. Значение техники отлова грызунов при количественном
изучении эктопаразитов и нидиколов. В сб.: Петров В.С. (ред.) Наземные и водные экосистемы:
Сборник научных трудов. Горький, Изд-во ГГУ, 112-117. [Borisova V.I., Nazarova I.V., Orekhov E.M.
1982. Znachenie tekhniki otlova gryzunov pri kolichestvennom izuchenii ektoparazitov i nidikolov. In:
Petrov V.S. (eds). Nazemnye i vodnye ekosistemy: Sbornik nauchnyh trudov. Gor'kij, Izdatelstvo GGU,
112-117. (in Russian)].
Брегетова Н.Г. 1956. Гамазовые клещи (Gamasoidea). Определители по фауне СССР, издаваемые Зоологи-
ческим институтом АН СССР. Вып. 61. М.-Л., АН СССР, 246 с. [Bregetova N.G. 1956. Gamazovyye
kleshchi (Gamasoidea). Opredeliteli po faune SSSR, izdavayemyye Zoologicheskim institutom AN SSSR,
Vol. 61, M.-L., AS USSR, 246 pp. (in Russian)].
Бугмырин С.В., Яковлев В.В., Беспятова Л.А. 2021. Линия ловушек для отлова мелких млекопитающих с
регистрацией времени срабатывания. Труды КарНЦ РАН. Экспериментальная биология (3): 103-108.
[Bugmyrin S.V., Yakovlev V.V., Bespyatova L.A. 2021. Small mammal trap line with capture time logging.
Высоцкая С.О., Кирьянова А.Н. 1970. Методы сбора и изучения блох и их личинок. Л. Наука, 83 с.
[Vysockaya S.O., Kirʼyanova A.N. 1970. Metody sbora i izucheniya bloh i ih lichinok. L., Nauka, 83 pp.
(in Russian)].
Ельшин С.В. 1992. Зависимость количественных показателей зараженности грызунов Ямала эктопаразитами
от метода отлова. Паразитология 26 (4): 338-340. [Elshin S.V. 1992.The dependence of the quantitative
indices of the infection of rodents from Yamal with ectoparasites on the method of catching. Parazitologiya
26 (4): 338-340. (in Russian)].
Карасева Е.В., Телицына А.В. 1996. Методы изучения грызунов в полевых условиях: Учеты численности
и мечения. М., Наука, 227 с. [Karaseva E.V., Telitsyna A.Yu. 1996. The methods of studying rodents in
the wild nature. M., Nauka, 227 pp. (in Russian)].
136
Колонин Г.В., Матюшина О.А., Болотин Е.И., Петрова Н.К. 1977. К оценке численности иксодовых клещей,
паразитирующих на мелких млекопитающих. Медицинская паразитология и паразитарные болезни
46 (5): 569-571. [Kolonin G.V., Matyushina O.A., Bolotin E.I., Petrova N.K. 1977. On estimation of the
abundance of ixodid ticks parasitizing small mammals. Meditsinskaya Parazitologiya i Parazitarnye Bolezni
46 (5): 569-571. (in Russian)].
Наумов Р.Л. 1958. О сползании клещей с грызунов. Зоологический журнал 37 (7): 1100-1101. [Naumov
R.L. 1958. On the slipping dawn of ticks from the rodents those latter being caught in killing traps.
Zoologicheskiy zhurnal 37 (7): 1100-1101. (in Russian)].
Шефтель Б.И. 2018. Методы учета численности мелких млекопитающих. Russian Journal of Ecosystem
Ecology 3 (3): 1-21. [Sheftel B.I. 2018. Metods for estimating the abundence of small mammals. Russian
Шилова С.А., Троцкий Б.Б., Мальков Г.Б., Белькович В.М. 1958. Значение подвижности лесных мышевид-
ных грызунов в распределение клещей Ixodes persulcatus P.Sch. в очагах весенне-летнего энцефалита.
Зоологический журнал 37 (6): 931-938. [Shilova S.A., Troitsky V.B., Malkov G.B., Belkovich V.M. 1958.
The role the mobility of forest mouse-like rodents in the distribution of Ixodes persulcatus P. Sch. in the
nidi of spring summer encephalitis. Zoologicheskiy zhurnal 37 (6): 931- 938. (in Russian)].
Яковлев В.В., Бугмырин С.В., Беспятова Л.А. 2020. Устройство для отлова мелких млекопитающих. Па-
тент на полезную модель RU 195297 U1, 22.01.2020. [Yakovlev V.V., Bugmyrin S.V., Bespyatova
L.A. 2020. Ustroystvo dlya otlova melkikh mlekopitayushchikh. Patent na poleznuyu modelʼ].
Bespyatova L.A., Bugmyrin S.V. 2006. Species diversity of gamasid mites (Parasitiformes, Gamasina) on small
mammals from the middle taiga of Karelia. Acarina 14 (2): 209-214.
De Pelsmaeker N., Korslund L., Steifetten Ø. 2020. Do bank voles (Myodes glareolus) trapped
in live and lethal traps show differences in tick burden? PLoS ONE 15 (9): e0239029.
Gallagher K., Bodin T., Sambridge M., Weiss D., Kylander M., Large D. 2011. Inference of abrupt changes in
noisy geochemical records using trans dimensional change point models. Earth and Planetary Science
Hammer Ø., Harper D.A.T., Ryan P.D. 2001. PAST: paleontological statistics software package for education and
data analysis. Paleontological Electronica 4 (1): 1-9.
Nakao M., Sato Y. 1996. Refeeding activity of immature ticks of Ixodes persulcatus and transmission
of Lyme disease spirochete by partially fed larvae. The Journal of Parasitology 82 (4): 669-672.
Reiczigel J., Marozzi M., Fabian I., Rozsa L. 2019. Biostatistics for parasitologists - a primer to Quantitative
137
ON THE LOSS OF ECTOPARASITES OF SMALL MAMMALS CAPTURED
IN SNAP TRAPS
N. A. Kocherova, L. A. Bespyatova, S. V. Bugmyrin
Keywords: snap traps, fleas, lice, mites, ticks, Ixodes persulcatus, Ixodes trianguliceps
SUMMARY
This study is aimed to comparatively estimate host abandonment times for different taxonomic
groups of small mammal ectoparasites after host capture in standard snap traps. The input for the
analysis was the data obtained in experiments with specially designed trap line with automatic log-
ging of animal capture instant. Overall, our results support the basic rule that ectoparasites abandon
the dead host after some time. The most tangible and rapid reduction in infection rates after host’s
death was detected for Ixodes persulcatus. Fleas and parasitic gamasid mites abandon the host during
the first 2-4 hours; lice - 9-13 hours after snap-trapping. Ixodes trianguliceps displayed no sharp
decline in abundance for a prolonged period of time.
138