ПАРАЗИТОЛОГИЯ, 2022, том 56, № 4, с. 303-313.
УДК 576.895.421
ЭКОЛОГО-ФАУНИСТИЧЕСКИЙ КОМПЛЕКС
ВИДОВ ИКСОДОВЫХ КЛЕЩЕЙ (PARASITIFORMES, IXODIDAE)
НА ОСТРОВЕ ПУТЯТИНА (ПРИМОРСКИЙ КРАЙ)
© 2022 г. Т. В. Звереваa, А. Я. Никитинb, *, Н. С. Солодкаяa,
Ю. А. Вержуцкаяb, Н. С. Гордейкоa, С. В. Балахоновb
aПриморская противочумная станция Роспотребнадзора,
ул. Дзержинского, 46, Уссурийск, 692512 Россия
bИркутский научно-исследовательский противочумный институт Роспотребнадзора,
ул. Трилиссера, 78, Иркутск, 664047 Россия
* e-mail: nikitin_irk@mail.ru
Поступила в редакцию 29.06.2022 г.
После доработки 08.07.2022 г.
Принята к публикации 08.07.2022 г.
На островах Японского моря (Приморский край) выявлены би- и полидоминантные типы
населения пастбищных иксодид. Остров Путятина (площадь 27.9 км²) единственный из остро-
вов, исследованных в 1982-1983 гг., где наблюдался монодоминантный тип населения с вы-
раженным преобладанием Haemaphysalis concinna Koch, 1844. Цель работы - описать совре-
менную структуру населения гемипопуляций имаго иксодовых клещей на о. Путятина. Сбор
клещей с растительности на флаг проведен 17.05-21.05.2021 г. с различных ландшафтных
участков острова. В результате зарегистрированы (в порядке убывания): Dermacentor silvarum
Olenev, 1932, H. concinna Koch, 1844, Ixodes persulcatus Schulze, 1930, H. japonica douglasi
Nuttall et Warburton, 1915, I. pavlovskyi pavlovskyi Pomerantzev, 1946. Всего собрано 354 особи
и установлен низкий индекс обилия клещей. На острове перестал встречаться H. longicornis
Neumann, 1901, резко возросла в сборах доля D. silvarum, в меньшей степени I. persulcatus,
впервые зарегистрирован I. pavlovskyi. Обсуждаются возможные причины изменения струк-
туры сообщества иксодид. Показано, что на о. Путятина отсутствует монодоминирование,
произошло формирование гемафизалисно-дермаценторного типа населения (D. silvarum 45.8%
и H. concinna 33.3%).
Ключевые слова: иксодовые клещи, острова Приморья, видовое разнообразие
DOI: 10.31857/S0031184722040044, EDN: Fgpnfk
303
Приморский край характеризуется значительным разнообразием видов иксодо-
вых клещей, которое по последним данным насчитывает 22 представителя семейства
(Леонова, 2020). Вместе с тем на материке в пихтово-еловых, смешанных и широ-
колиственных лесах преимущественно доминирует Ixodes persulcatus Schulze, 1930
(таежный клещ), в отдельных небольших антропогенно нарушенных локациях - Der-
macentor silvarum Olenev, 1932, на участках с повышенной влажностью - Haemaphy-
salis concinna Koch, 1844, а в хвойно-широколиственных лесах южной прибрежной
части содоминантом таежного клеща изредка является Haemaphysalis japonica douglasi
Nuttall et Warburton, 1915 (Худяков, 1968; Беликова, 1969; Болотин, 1980; Колонин,
1986; Леонова, 1997). Лишь на островах Залива Петра Великого в Японском море
(Приморский край) в полной мере проявляется видовое богатство семейства (Коло-
нин, 1986; Леонова, 1997; Никитин и др., 2018; Гордейко, 2019). В частности, ни на
одном из обследованных островов Приморья не зафиксировано абсолютное доми-
нирование таежного клеща, на большинстве из них наблюдаются, по терминологии
Алифанова и др. (1973), би- и полидоминантные типы населения пастбищных видов
иксодовых клещей. На сегодняшний день единственным островом Приморья, где об-
наружен монодоминантный тип населения (с преобладанием H. concinna), является
о. Путятина (Колонин, 1986). Однако описание фауны гемипопуляций половозрелых
клещей на его территории сделано в 1983 г., 40 лет назад, и структура видов вполне
могла измениться. Кроме того, есть данные, что в конце 50-х начале 60-х годов на
о. Путятина содоминантами являлись H. japonica и Haemaphysalis longicornis Neu-
mann, 1901 (прежнее название H. neumanni) (Худяков, 1968), что косвенно подтверж-
дает отсутствие на островах монодоминатных типов населения и возможность до-
статочно быстрых преобразований структуры сообществ иксодид (Гордейко, 2019).
Цель работы - описать современную эколого-фаунистическую структуру населения
гемипопуляций имаго иксодовых клещей на о. Путятина.
МАТЕРИАЛ И МЕТОДИКА
Сбор взрослых клещей на о. Путятина проведен на флаг с растительности 17.05-21.05.2021
г. Всего отработано 26.5 флаго-часов и собрано 354 особи иксодовых клещей пяти видов.
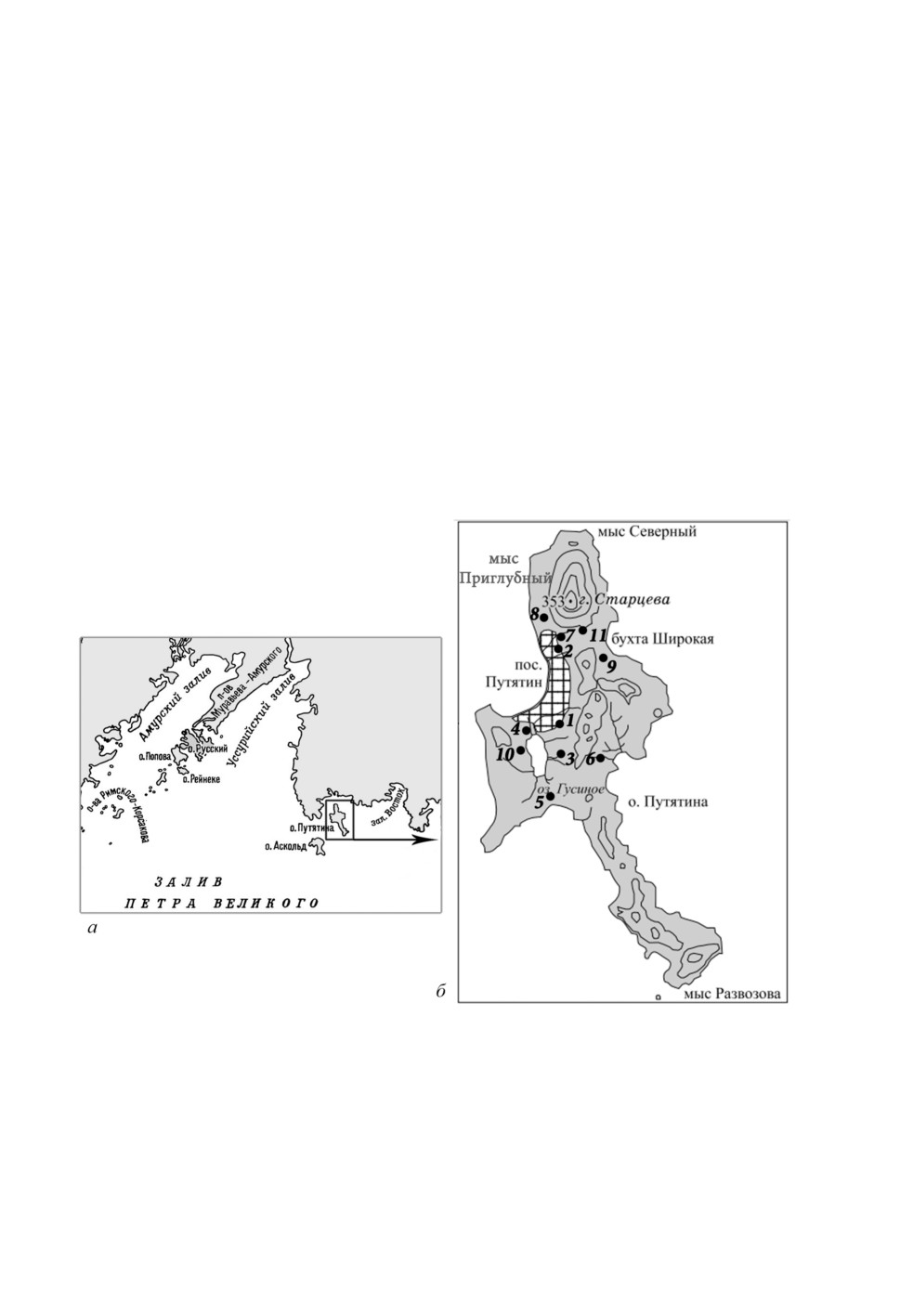
Расположение о. Путятина в Японском море (рис. 1а) и маршруты (точки) сбора иксодовых
клещей отражены на картосхеме (рис. 1б). Географические координаты, примерно соответствую-
щие центрам маршрутов, приведены в табл. 1. Ниже дано краткое описание климатических и
ландшафтных особенностей о. Путятина, в том числе на маршрутах, обследованных при сборе
иксодовых клещей.
Остров Путятина, входящий в состав северо-восточной группы архипелага залива Петра
Великого Японского моря, является вторым по величине (площадь 27.9 км²) и входит в число
четырех с постоянно проживающим населением.
Климат о. Путятина типичен для юга Приморья и относится к муссонной области умерен-
ных широт (Алисов, 1956). Средняя температура января -12.0°C, августа +21.0°C, среднегодо-
вая - +4.9°С. Рельеф острова холмистый, берега возвышенные и во многих местах прорезаны
долинами и оврагами. Самая высокая точка - сопка Старцева (353 м над ур. м.) расположена
в северной части острова. На о. Путятина имеется несколько пресных озер, самое большое
из них - Гусиное.
304
Территория о. Путятина относится к зоне хвойно-широколиственных лесов Маньчжурской
провинции Дальневосточной области (Колесников, 1961). В его современной ландшафтной
структуре доминирующее положение (более 70%) занимают пологие и средней крутизны скло-
ны, покрытые вторичными широколиственными лесами: дуба монгольского (Quercus mongolica),
липы амурской (Tilia amurensis), клена мелколистного (Acer mono) и ложнозибольдова
(A. pseudosieboldianum), берез даурской (Betula davurica) и плосколистной (B. platyphylla), ольхи
волосистой (Alnus hirsuta) (Лящевская и др., 2014; Лящевская, 2016). Через описанную зону
проходили маршруты № 7, 9, частично 2 и 6 (рис. 1б, табл. 1).
На склонах сопки Старцева сохранились высокосомкнутые дубовые леса. Их нижние по-
логие части заняты кустарниково-разнотравными сообществами из леспедечника (Lespedeza
bicolor), лещины маньчжурской (Corylus mandshurica), полыни Гмелина (Artemisia gmelinii).
Сборы иксодовых клещей с этих участков соответствуют маршрутам № 8, 11, частично 7
(рис. 1б, табл. 1).
Комплекс равнинных ландшафтов (около 12% площади острова) в основном формирует его
центральную часть в районе озер Гусиное и Цаплинное, а также вдоль восточного побережья
(маршрут № 5 и частично 3). Здесь преобладают урочища прибрежных низменностей, с раз-
нотравными болотами и влажными разнотравными лугами (рис. 1б, табл. 1).
В нижней части западного склона центральной части острова распространены черемухово-
ивовые леса из черемухи Максимовича (Padus maximowiczii), ив Шверина (Salix schwerinii) и
козьей (S. caprea), дуба монгольского, клена зеленокорого (Acer tegmentosum), ольхи волосистой
(маршрут № 10).
Наибольшему антропогенному влиянию на о. Путятина подвергается территория в районе
поселка на побережье бухты Назимова (маршрут № 4, частично № 1-3), где наблюдаются
значительное обеднение и адвентизация растительных комплексов (рис. 1б, табл. 1). Влияние
деятельности человека, особенно сильное в 70-80 гг. ХХ века, привело к увеличению площади
лугово-кустарниковых сообществ постпирогенного происхождения, преимущественно пред-
ставленных ивой козьей и малиной боярышниколистной (Rubus crataegifolius).
Особенностью островных территорий является отсутствие многих обычных для материка
видов диких прокормителей иксодовых клещей. На о. Путятина зарегистрированы: Vulpes vulpes
Limxams, 1758 (лисица), Erinaceus amurensis Schrenk, 1859 (амурский еж), Apodemus agrarius
Pallas, 1771 (полевая мышь), Microtus fortis Buchner, 1889 (дальневосточная полевка), Rattus
norvegicus Веrkenhout, 1769 (серая крыса), Сrоcidura suaveolens Раllas, 1811 (малая белозуб-
ка) (Шереметьев, 2001). Из прокормителей клещей, связанных с деятельностью человека, на
острове присутствуют домашние и сельскохозяйственные животные.
Видовую принадлежность клещей определяли по морфологическим признакам (Померан-
цев, 1950; Филиппова, 1977, 1997) с использованием стереомикроскопов в отраженном свете
(увеличение ×80, МС-2 «Биомед» и ×84, МБС-10, ЛОМО, Россия).
Сравнение фаунистических структур комплекса видов иксодид, обитавших ранее (Колонин,
1986), с современным комплексом проведено по показателю процентного сходства и индек-
су Пиелу, который также известен как показатель выравненности Шеннона (Уиттекер, 1980).
Границы 95% доверительного интервала возможной изменчивости встречаемости отдельных
видов в сборах клещей рассчитаны с применением F распределения (Животовский, 1991). Все
расчеты выполнены в компьютерной программе Excel.
305
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
За время обследования (17.05-21.05.2021 г.) на о. Путятина собраны (в порядке
убывания): D. silvarum (112 самок, 50 самцов), H. concinna (58 самок, 60 самцов),
I. persulcatus (27 самок, 20 самцов, 9 нимф), H. japonica (7 самок, 7 самцов), Ixodes
pavlovskyi pavlovskyi Pomerantzev, 1946 (4 самки) (табл. 2). Всего 354 особи. Таким
образом, на острове наблюдается эколого-фаунистический комплекс видов иксодид,
который характерен для окрестностей населенных пунктов и формируется под влия-
нием человека (Беликова, 1969).
Наибольший индекс обилия (ИО) клещей выявлен на маршрутах № 1 и 10 (табл. 1,
рис. 1б). Первый из них проходил непосредственно по окраине пос. Путятина и в рай-
оне кладбища (6.0 особей на флаго-час); второй маршрут - по склону вдоль за-
падного берега оз. Гусиное через дубово-мелколиственный лес и ольхово-березовые
колки (7.5 особей флаго-час). Т. е. оба маршрута находятся в местах, часто посещае-
мых местным населением и туристами. На остальных обследованных территориях
о. Путятина ИО клещей значительно ниже. Отметим, что ИО клещей на о. Путятина
ниже не только среднемноголетнего показателя на материке, но и на самом большом
Рисунок 1. Картосхема района проведенных исследований: а - расположение о. Путятина
в заливе Петра Великого, б - места сбора иксодовых клещей на о. Путятина. Номера
точек сбора на картосхеме о. Путятина соответствуют номерам, приведенным в табл. 1.
Штриховкой показана зона наиболее полного обследования.
Figure 1. Figure. Scheme of the location of Putyatin Island in Peter the Great Bay (a)
and the collecting sites of ixodid ticks in its territory (b). The numbers of collecting sites
in the map of Putyatin Island are given in Table. 1. The shaded area is the area of the most
complete examination.
306
в Приморье о. Русский и на относительно небольшом о. Рейнеке (Никитин и др.,
2018; Гордейко, 2019).
Таблица 1. Описание точек сбора иксодовых клещей на о. Путятина
Table 1. Description of collecting sites of ixodid ticks in Putyatin Island
Номера точек
Географические
маршрутов,
Наименование
координаты середины
Собрано клещей /
приведенных
исследуемой
маршрута
число клещей
на картосхеме
локации
сбора клещей
на флаго-час
острова (рис. 1б)
(с.ш., в.д.)
1
Поселок Путятин:
42.858054°, 132.420642°
125 / 6.0
ул. Арсеньева, ул. Озерная,
район кладбища
2
Окраина пос. Путятин
42.876452°, 132.417007°
32 / 1.5
(ул. Набережная)
3, 4
Окрестности оз. Гусиное
42.851747°, 132.421356°
43 / 1.3
42.857723°, 132.409078°
5
Окрестности бухты Ступени
42.840354°, 132.420052°
4 / 1.0
6
Окрестности бухты Петух
42.850500°, 132.433094°
5 / 1.0
7
Подножье сопки Старцева
42.879384°, 132.418641°
12 / 3.0
(южный склон)
8
Подножье сопки Старцева
42.8885308°, 132.412903°
18 / 3.0
(западный склон)
9
Окрестности бухты Широкая
42.875576°, 132.434159°
5 / 1.0
10
Склон вдоль западного
42.852749°, 132.406840°
105 / 7.5
берега оз. Гусиное
11
Подножье сопки Старцева
42.881780°, 132.426119°
5 / 0.3
(юго-восточный склон)
Данные проведенного обследования о. Путятина подтверждают малую распростра-
ненность на нем клещей из рода Ixodes (табл. 2). Вместе с тем, если ранее территория
этого острова - единственная из изученных островов Приморья - характеризовалась
монодоминантным типом населения, то теперь и на о. Путятина подтверждено на-
личие бидоминантного дермаценторно-гемофизалисного комплекса видов: D. silvarum
45.8% и H. concinna 33.3%. По сравнению с описаниями, сделанными ранее (Худяков,
1968; Колонин, 1986), на острове исчез H. longicornis (не выявлен среди 354 клещей),
зато резко возросла встречаемость D. silvarum (с 0.4 до 45.8%), в меньшей степени
возросла встречаемость I. persulcatus, впервые выявлен I. pavlovskyi.
Исчезновение H. longicornis, вероятно, обусловлено истреблением браконьерами
на о. Путятина пятнистого оленя, встречавшегося здесь до начала-середины 90-х
годов ХХ столетия. Несмотря на достаточно широкий круг прокормителей у этого
307
вида клещей (Белов и др., 2019), ряд авторов особую роль отводят пятнистому оленю
(Беликова, 1969; Колонин, 1986; Филиппова, 1997). Отмеченный характер измене-
ния фаунистического комплекса населения иксодид немаловажен по двум причинам.
Первое, в последнее время эпидемиологическое значение H. longicornis рассматрива-
ется как чрезвычайно высокое, особенно в соседних Китае, Южной Корее и Японии
(Андаев и др., 2021; Zhao et al., 2021). Второе, в 2021 г. в южной части о. Путятина
вновь создано частное оленеводческое хозяйство, куда завезены пятнистые олени.
Не исключено, что после появления оленей структура видов населения клещей на о.
Путятина вновь обогатится H. longicornis, и будет важно оценить особенности его
расселения по территории острова и вероятный характер связанного с этим эпиде-
миологического риска.
Таблица 2. Сравнение встречаемости видов иксодовых клещей при сборах с растительности
на о. Путятина: 27.05-29.05.1983 г. (Колонин, 1986) и 17.05-20.05.2021 г.
Table 2. Comparison of the occurrence of ixodid tick species in Putyatin Island in collections
of May 27-May 29, 1983 (Kolonin, 1986) and our collections of May 17-20, 2021
Границы 95%
Число
Доля от суммы
доверительного
Вид клещей
особей
особей, %
интервала изменения
доли вида
Сбор 1983 г.
I. persulcatus
12
4.7
2.8-8.3
H. japonica
28
10.9
8.0-15.1
H. concinna
202
78.6
78.3-78.7
H. longicornis
14
5.4
3.5-9.0
D. silvarum
1
0.4
0-13.2
Сумма
257
100
-
Индекс Пиелу
0.47
Сбор 1921 г.
I. persulcatus
56
15.8
15.6-16.1
I. pavlovskyi
4
1.1
0.4-4.0
H. japonica
14
4.0
2.5-6.6
H. concinna
118
33.3
33.2-33.5
D. silvarum
162
45.8
45.6-45.9
Сумма
354
100
-
Индекс Пиелу
0.71
Значение показателя
42.3
процентного сходства
между двумя сборами
308
Менее понятна причина роста встречаемости D. silvarum на территории остро-
ва (табл. 2). По имеющимся данным он всегда был на острове малочисленным
(Худяков, 1968; Колонин, 1986). Возможно, повышение обилия вида стало следствием
значительного роста числа заброшенных и зарастающих участков земли, возникших
в результате сокращения населения поселка в 90-е годы XX века и вывода с его
территории воинских частей. Такие локации в настоящее время являются местами
постоянного самовыпаса скота, что, по нашим наблюдениям, обычно приводит к уве-
личению обилия D. silvarum. В подтверждение этого предположения отметим, что
100 особей этого вида из 162 (61.7%) собрано непосредственно вблизи поселка: на
маршрутах № 1, 2, 4, 7 (рис. 1б). Подобный тип населения иксодид на территории
Приморья Беликова рассматривает как целиком поддерживаемый деятельностью че-
ловека (Беликова, 1969).
Высокое видовое разнообразие, формирование би- и полидоминантных типов на-
селения клещей (табл. 2) характерны и для других островов Приморья (Гордейко,
2019). Причиной этого могут быть особенности растительных формаций островных
территорий в Японском море. Так, по данным Гуреминой (2005), «Характерными
чертами растительности островов в настоящее время являются фрагментарность («ми-
ниатюрность») сообществ, высокая гетерогенность и пестрота видового состава …»,
которые обусловлены расчленённостью рельефа, различиями в крутизне и направле-
нии склонов, близостью моря, разнообразием почв «… создают на небольших про-
странствах разнообразные условия местообитания» (с. 95). И далее: «Влияние климата
и почвы усугубляется деятельностью человека и в результате создаются очень пестрые
разнообразные насаждения» (с. 112).
Именно антропогенное преобразование территорий, прилегающих к ряду населен-
ных пунктов Западной Сибири, исследователи рассматривают как одну из возмож-
ных причин расширения ареала I. pavlovskyi, что привело к появлению локальных
бидоминантных типов населения этого вида с таежным клещом и поливекторности
природных очагов трансмиссивных инфекций (Ливанова и др., 2011; Романенко, 2011;
Малькова и др., 2012; Якименко и др., 2013). Таким образом, причина формирования
би- или полидоминантных типов населения иксодовых клещей связана с гетероген-
ностью среды их обитания.
Считается, что величина индекса Пиела характеризует благоприятность условий
обитания видов (Сообщества и популяции …, 2010). В случае, когда этот показатель
находится в границах от 0.50 до 0.79, виды имеют неравномерное распределение и
для большинства из них характерно относительно угнетенное существование. Если
индекс Пиелу < 0.5 - условия неблагоприятны для всех видов, кроме доминанта.
Согласно полученным данным (табл. 2), значение индекса Пиелу увеличивается, сле-
довательно, условия обитания на о. Путятина в современный период становятся бо-
лее выравненными и благоприятными для сосуществования разных видов иксодовых
клещей.
Величина процентного сходства видового состава клещей из двух сборов, про-
веденных с территории одного острова с 40-летним разрывом, составила около 42%
(табл. 2). Значение этого показателя между отдельными сборами иксодид с одних
309
и тех же территорий на материке колеблется от 69 до 94%, а на о. Русский составило
88% (Гордейко, 2019), т.е. было значительно большим. Однако во всех этих случаях
период времени между повторными наблюдениями был около трех лет.
Таким образом, в настоящее время на о. Путятина выявлен бидоминатный
гемафизалисно-дермаценторный тип населения иксодовых клещей с низким ИО для
каждого представителя семейства. Следовательно, эпидемиологический риск для жи-
телей острова и туристов по инфекциям, передающимся клещами, невысок и, как и
на других островах с таким видовым составом населения, преимущественно связан
с клещевыми риккетсиозами (Никитин и др., 2018). Для более точной оценки эпи-
зоотологического и эпидемиологического потенциалов островной фауны иксодид
собранные клещи переданы в специализированную лабораторию для изучения их
инфицированности патогенными для человека возбудителями.
СПИСОК ЛИТЕРАТУРЫ
Алисов Б.П. 1956. Климат СССР. М., Издательство Московского университета, 127 с. [Alisov B.P. 1956.
Climate of the USSR. Moscow, Moscow University Publishing House, 127 pp. (In Russian)].
Алифанов В.И., Богданов И.И., Нецкий Г.И., Мальков Г.Б. 1973. Типы населения иксодовых клещей (Ixodi-
dae Murr.) и их зараженность возбудителями природноочаговых болезней на территории Западной
Сибири. Эпидемиологическая география клещевого энцефалита, Омской геморрагической лихорадки
и клещевого риккетсиоза Азии в Западной Сибири (вопросы инфекционной патологии). Омск, Изд-
во Омский научно-исследовательский институт природноочаговых инфекций, 15-26. [Alifanov V.I.,
Bogdanov I.I., Netskii G.I., Malkov G.B. 1973. Population types of ixodid ticks (Ixodidae Murr.) and
their infestation with naturofocal disease agents in West Siberia. Epidemiological geography of tick-borne
encephalitis, Omsk hemorrhagic fever and tick-borne rickettsiosis in Asia in Western Siberia (issues of
infectious pathology). Omsk, Izdatelstvo Omskii nauchno-issledovatelskii Institute prirodnoochagovykh
infektsii 15-26. (In Russian)].
Андаев Е.И., Адельшин Р.В., Балахонов С.В. 2021. Тяжелая лихорадка с тромбоцитопеническим синдро-
мом: современная эпидемиологическая ситуация. Эпидемиология и Вакцинопрофилактика 20 (4):
akhonov S.V. 2021. Severe Fever with Thrombocytopenic Syndrome: Current Epidemiological Situation.
4-114-122 (In Russian)].
Беликова Н.П. 1969. Основные эколого-фаунистические комплексы иксодид, поддерживающие существова-
ние природных очагов инфекций в Приморском крае. Медицинская паразитология и паразитарные
болезни 4: 401-405. [Belikova N.P. 1969. The main ecological and faunal complexes of Ixodid ticks
supporting the existence of natural foci of infections in Primorsky Territory. Medtsinsaya parazitologiya i
parazitarnye bolezni 4: 401-405. (In Russian)].
Белов Ю.А., Москвина Т.В., Щелканов Е.М., Бурухина Е.Г., Волков Ю.Г., Попов И.А., Какарека Н.Н.,
Галкина И.В., Панкратов Д.В., Суровый А.Л., Щелканов М.Ю. 2019. К вопросу о северной грани-
це ареала и хозяевах клеща Haemaphysalis longicornis (Acari: Ixodidae) в Приморском крае. В кн.:
Чтения памяти Алексея Ивановича Куренцова 30: 177-182. [Belov Yu.A., Moskvina T.V., Shchelkanov
E.M., Burukhina E.G., Volkov Yu.G., Popov I.A., Kakareka N.N., Galkina I.V., Pankratov D.V., Surovyi
A.L., Shchelkanov M.Yu. 2019. About the northern boundary of distribution and the host species of the
ticks Haemaphysalis longicornis (Acari: Ixodidae) in Primorsky Territory. A.I. Kurentsov's Annual Me-
morial Meetings 30: 177-182. (In Russian)]. https://doi.org/10.25221/kurentzov.30.16 http://zoobank.org/
References/6E48FF36-C541-4034-A8D6-049179C7C0CF
310
Болотин Е.И. 1980. Эколого-фаунистический обзор иксодовых клещей Приморского края. Владивосток,
Деп. в ВИНИТИ 22.04.80, № 1906. [Bolotin E.I. 1980. Ecological and faunistic review of ixodid ticks
in Primorsky Territory. Vladivostok, Deposited at VINITI on April 22, 1980, № 1906. (In Russian)].
Гордейко Н.С. 2019. Клещи семейства Ixodidae Приморья: типы населения, паразито-хозяинные связи,
инфицированность патогенами (на примере материковых и островных сообществ). Автореф. дис.
… канд. биол. наук. Иркутск, 22 с. [Gordeyko N.S. 2019. Ticks of the family Ixodidae of Primorye:
population types, parasite-host relationships, infection with pathogens (on the example of mainland and
island communities). Abstract of the dissertation of the candidate of biological Sciences. Irkutsk, 22 pp.
(In Russian)].
Гуремина Н.В. 2005. Ландшафтная характеристика и рекреационное освоение островов залива Петра Ве-
ликого (Японское море). Дис
канд. геогр. наук. Владивосток, 185 с. [Guremina N.V. 2005. Land-
scape characteristics and recreational development of the islands of Peter the Great Bay (Sea of Japan):
Dissertation for the degree of Candidate of Geographical Sciences. Vladivostok, 185 pp. (In Russian)].
Животовский Л.А. 1991. Популяционная биометрия. М., Наука, 271 с. [Zhivotovsky L.A. 1991. Population
biometry. Moscow, Nauka, 271 pp. (In Russian)].
Колесников Б.П. 1961. Растительность. Дальний Восток: физико-географическая характеристика. М., Изд-во
АН СССР, 182-245. [Kolesnikov B.P. 1961. Vegetation. Far East: physical and geographical characteristics.
Moscow, Izdatelstvo Akademii nauk SSSR, 182-245. (In Russian)].
Колонин Г.В. 1986. Материалы по фауне иксодовых клещей юга Приморского края. Паразитология 20 (1):
15-18. [Kolonin G.V. 1986. Materials on the fauna of ixodid ticks from the south of Primorye Territory.
Parazitologiya 20 (1): 15-18. (In Russian)].
Леонова Г.Н. 1997. Клещевой энцефалит в Приморском крае: вирусологические и эколого-
эпидемиологические аспекты. Владивосток, Дальнаука, 190 с. [Leonova G.N. 1997. Tick-borne en-
cephalitis in Primorsky Territory: virological and environmental and epidemiological aspects. Vladivostok,
Dalnauka, 190 pp. (In Russian)].
Леонова Г.Н. 2020. Клещевой энцефалит в Дальневосточном очаговом регионе евразийского континента.
Журнал микробиологии, эпидемиологии и иммунобиологии 97 (2): 150-158. [Leonova G.N. 2020.
Tick-borne encephalitis in the Far East focal region of the Eurasian continent. Zhurnal mikrobiologii,
2020-97-2-150-158
Ливанова Н.Н., Ливанов С.Г., Панов В.В. 2011. Особенности распределения клещей Ixodes persulcatus
и Ixodes pavlovskyi на границе лесной и лесостепной зон Приобья. Паразитология 45 (2): 94-103.
[Livanova N.N., Livanov S.G., Panov V.V. 2011. Characteristics of the distribution of ticks Ixodes persul-
catus and Ixodes pavlovskyi at the border between the forest and forest-steppe zones in the territory near
Ob river. Parazitologiya 45 (2): 94-103. (In Russian)].
Лящевская М.С., Киселёва А. Г., Родникова И.М., Пшеничникова Н.Ф., Ганзей К.С. 2014. Развитие
почвенно-растительного покрова острова Путятина в позднем голоцене (Японское море). География
и природные ресурсы 1: 124-133. [Lyashchevskaya M.S., Kiseleva A.G., Rodnikova I.M., Pshenichnikova
N.F., Ganzey K.S. 2014. Soil-vegetation cover development in Putyatin Island in the Late Holocene (Sea
of Japan). Geografiya i prirodnye resursy 1: 124-133. (In Russian)].
Лящевская М.С. 2016. Ландшафтно-климатические изменения на островах залива Петра Великого (Японское
море) за последние 20 000 лет. Успехи современного естествознания 11 (2): 372-379. [Lyaschevskaya
M.S. 2016. Landscape-climate changes of the Islands of Peter the Great Bay (Sea of Japan) during last 20
000 years. Uspekhi sovremennogo estesvoznaniya 11(2): 372-379. (In Russian)].
Малькова М.Г., Якименко В.В., Танцев А.К. 2012. Изменение границ ареалов пастбищных иксодовых кле-
щей рода Ixodes Latr., 1795 (Parasitiformes, Ixodidae) на территории Западной Сибири. Паразитология
46 (5): 369-383. [Malkova M.G., Yakimenko V.V., Tancev A.K. 2012. Changes in the ranges of pasture
311
ixodid ticks of the genus Ixodes Latr., 1795 (Parasitiformes, Ixodinae) in Western Siberia. Parazitologiya
46 (5): 369-383. (In Russian)].
Никитин А.Я., Морозов И.М., Андаев Е.И., Алленов А.В., Сидорова Е.А., Яковчиц Н.В., Бондаренко Е.И.,
Гордейко Н.С., Балахонов С.В. 2018. Видовой состав и возможное эпидемиологическое значение
иксодовых клещей (Ixodidae) на острове Рейнеке (Приморский край). Медицинская паразитология
и паразитарные болезни 1: 48-52. [Nikitin A.Ya., Morozov I.M., Andaev E.I., Allenov A.V., Sidorova
E.A., Yakovchits N.V., Bondarenko E.I., Gordeiko N.S., Balakhonov S.V. 2018. The species composition
and possible epidemiological importance of ticks (Ixodidae) on Reyneke Island (The Primorye Territory).
Medtsinsaya parazitologiya i parazitarnye bolezni 1: 48-52. (In Russian)].
Померанцев Б.И. 1950. Иксодовые клещи (Ixodidae). Фауна СССР. Паукообразные. М., Л., Изд-во АН
СССР, 4 (2): 224 с. [Pomerancev B.I. 1950. Ixodid ticks (Ixodidae). Fauna of the USSR. Arachnoidea.
Moscow-Leningrad, Nauka, 4 (2): 224 pp. (In Russian)].
Романенко В.Н. 2011. Многолетняя динамика численности и видового состава иксодовых клещей (Ixodidae)
на антропогенно нарушенных и естественных территориях. Паразитология 45 (5): 384-391. [Ro-
manenko V.N. 2011. Long-term dynamics of abundance and species composition of ixodid ticks (Ixodidae)
in anthropogenically disturbed and natural areas. Parasitology 45 (5): 384-391. (In Russian)].
Сообщества и популяции животных: экологический и морфологический анализ. 2010. Новосибирск-Мо-
сква, Товарищество научных изданий КМК, 256 с. [Animal communities and populations: ecological
and morphological analysis. 2010. Novosibirsk-Moscow, KMK Association of Scientific Publications, 256
pp. (In Russian)].
Уиттекер Р. 1980. Сообщества и экосистемы. Пер. с англ. М., Прогресс, 326 с. [Whittaker R. 1980. Com-
munities and ecosystems. Moscow, Progress, 326 p. (In Russian)]. (Whittaker R.H. 1975. Communities
and Ecosystems. 2nd Revise Edition. New-York, Macmillan Publishing Co, 385 p.).
Филиппова Н.А. 1977. Иксодовые клещи. Подсемейство Ixodinae. Фауна СССР. Паукообразные. Л., Наука,
4 (4): 396 с. [Filippova N.A. 1977. Ixodid ticks. Subfamily Ixodinae. Fauna of the USSR. Arachnoidea.
Leningrad, Nauka, 4 (4): 396 pp. (In Russian)].
Филиппова Н.А. 1997. Иксодовые клещи подсем. Amblyomminae. Фауна России и сопредельных стран.
Паукообразные. СПб., Наука, 436 с. [Filippova N.A. Ixodid ticks of the subfamily Amblyomminae.
Fauna of Russia and bordering countries. Arachnidae. SPb., Nauka, 436 pp.].
Худяков И.С. 1968. К материалам по изучению иксодовых клещей в южном Приморье Дальнего Востока
и на островах Японского моря. Известия Иркутского государственного научно-исследовательского
противочумного института Сибири и Дальнего Востока 27: 323-331. [Khudyakov I.S. 1968. To the
materials on the study of ixodid ticks in the southern Primorye of the Far East and in the islands of the
Sea of Japan. Izvestiya Irkutskogo gosudarstvennogo naucho-issledovatelskogo protivochumnogo institute
Sibiri i Dalnego Vostoka 27: 323-331. (In Russian)].
Шереметьев И.С. 2001. Наземные млекопитающие островов залива Петра Великого (Японское море). Дис.
… канд. биол. наук. Владивосток, 148 с. [Sheremetiev I.S. 2001. Terrestrial mammals of the Islands
of Peter the Great Bay (Sea of Japan). Dissertation for the degree of Candidate of Biological Sciences.
Vladivostok, 148 pp. [(In Russian)].
Якименко В.В., Малькова М.Г., Шпынов С.Н. 2013. Иксодовые клещи Западной Сибири: фауна, экология,
основные методы исследования. Омск, Изд-во ООО ИЦ «Омский научный вестник», 240 с. [Yaki-
menko V.V., Malkova M.G., Shpynov S.N. 2013. Ixodid ticks of Western Siberia: fauna, ecology, main
research methods. Omsk, Publishing house of LLC IC "Omsk scientific Bulletin", 240 pp. (in Russian)].
Zhao G.-P., Wang Y.-X., Fan Z.-W., Ji Y., Liu M.-J., Zhang W.-H, Li X.-L., Zhou S.-X., Li H., Liang S., Liu W.,
Yang Y., Fang L.-Q. 2021. Mapping ticks and tick-borne pathogens in China. Nature Communications 12
312
ECOLOGICAL-FAUNISTIC COMPLEX
OF IXODID TICK SPECIES (PARASITIFORMES, IXODIDAE)
IN PUTYATIN ISLAND (PRIMORSKY TERRITORY)
T. V. Zvereva, A. Ya. Nikitin, N. S. Solodkaya,
Yu. A. Verzhutskaya, N. S. Gordeyko, S. V. Balakhonov
Keywords: Ixodid ticks, Primorye Islands, species diversity
SUMMARY
Ixodid ticks communities in Primorsky Territory are characterized by a significant diversity of
species. The species richness of the family is more completely manifested in the islands of Primo-
rye than in the mainland. Bi- and polydominant ixodid communities are observed only in islands.
Monodominant tick community was, however observed only in Putyatin Island of the eight islands
surveyed in 1982-1983; the goal of the present work is to describe contemporary structure of hemi-
populations of adult ixodid ticks in Putyatin Island. Ticks were collected from vegetation by flagging
in May 17-21, 2021 at different locations in the island. The following species have been recorded in
the island (in descending order of abundance): Dermacentor silvarum Olenev, 1932, Haemaphysalis
concinna Koch, 1844, Ixodes persulcatus Schulze, 1930, H. japonica douglasi Nuttall et Warburton,
1915, I. pavlovskyi pavlovskyi Pomerantzev, 1946. In total, 354 individuals of ixodid ticks were col-
lected. A low index of tick abundance was revealed. As a result of survey work in Putyatin Island in
2021, H. longicornis Neumann, 1901 was not found, I. pavlovskyi was registered for the first time, the
abundance of D. silvarum increased significantly, and the abundance of I. persulcatus increased to a
lesser extent. Possible causes of changes in the structure of ixodid tick communities are discussed. It
has been established that bidominant complex of ixodid tick species is currently observed on Putyatin
Island (D. silvarum 45.8% и H. concinna 33.3%), as well as in other islands of Primorsky Territory.
313