ПАРАЗИТОЛОГИЯ, 2022, том 56, № 5, с. 385-417.
УДК 576.895.775:574.9/579.842.23
ПАЛЕАРКТИЧЕСКИЕ ВИДЫ БЛОХ
РОДА XENOPSYLLA (SIPHONAPTERA: PULICIDAE),
ПАРАЗИТИРУЮЩИЕ НА ПЕСЧАНКАХ (RHOMBOMYS, MERIONES),
И ИХ РОЛЬ В ПРИРОДНЫХ ОЧАГАХ ЧУМЫ
© 2022 г. С. Г. Медведевa,*, Д. Б. Вержуцкийb,**, Б. К. Коттиc, d,***
a Зоологический институт РАН,
Университетская наб., 1, Санкт-Петербург, 199034 Россия
b Иркутский научно-исследовательский противочумный институт Роспотребнадзора,
ул. Трилиссера, 78, Иркутск, 664047 Россия
cСеверо-Кавказский федеральный университет,
ул. Пушкина, 1, Ставрополь, 355009 Россия
d Ставропольский противочумный институт Роспотребнадзора,
ул. Советская, 13, Ставрополь, 355035 Россия
*e-mail: smedvedev@zin.ru; sgmed@mail.ru
**e-mail: verzh58@rambler.ru
***e-mail: boris_kotti@mail.ru
Поступила в редакцию 10.09.2022 г.
После доработки 19.09.2022 г.
Принята к публикации 20.10.2022 г.
В обзоре проанализированы таксономическое разнообразие, особенности распростране-
ния и паразито-хозяинных связей блох рода Xenopsylla (Pulicidae). Показано, что шесть видов
и два подвида блох рода Xenopsylla, паразитирующих на песчанках на территории России
и сопредельных стран, известны как основные, второстепенные или случайные переносчики
чумной инфекции.
Ключевые слова: блохи, Siphonaptera, переносчики возбудителя чумы, таксономическое
разнообразие, Xenopsylla
DOI: 10.31857/S0031184722050039, EDN: fhbcct
Данная статья является очередной в серии публикаций, посвященных анализу
особенностей распространения и паразито-хозяинных связей видов блох - активных
переносчиков возбудителя чумы. В предыдущих работах (Котти, Жильцова, 2019;
Медведев и др., 2019, 2020, 2021, 2022; Медведев, Вержуцкий, 2019; Вержуцкий и др.,
385
2021) рассмотрены видовой состав, распространение, биоценотические связи и эпи-
зоотологическое значение представителей родов Citellophilus, Oropsylla, Rhadinopsylla,
Neopsylla, Frontopsylla и Paradoxopsyllus. Целью настоящей работы является обобще-
ние данных по блохам рода Xenopsylla, многие представители которого, паразитируя
на песчанках, также известны как основные переносчики чумной инфекции. Как и
ранее, в качестве основных задач поставлена оценка особенностей распространения,
паразито-хозяинных связей палеарктических видов данного рода в целом и их от-
ношения к чумной инфекции.
Блохи рода Xenopsylla:
особенности распространения и паразито-хозяинных связей
По числу видов род Xenopsylla - один из крупных в фауне блох Старого Света.
Он насчитывает 75 видов и 10 подвидов, распространенных преимущественно в Аф-
ротропической области и субтропической зоне Палеарктической области. Несколько
видов обитают в Индо-Малайской и Австралийской областях. В фауне России и со-
предельных стран Центральной и Средней Азии род Xenopsylla, тяготеющий к по-
лупустынным и пустынным регионам, известен по 10 видам и трем подвидам.
Представители рода паразитируют на широком круге хозяев, включающем сусли-
ков, хомячков рода Cricetulus, песчанок, тушканчиков и мышиных (табл. 1). Ряд его
видов относятся к синантропным. Так, как и крысиная блоха Nosopsyllus (N.) fasciatus
(Bosc, 1800), всесветное распространение имеет обитающая на черной и серой крысах
в отапливаемых помещениях блоха Xenopsylla cheopis (Rothschild, 1903). Космополи-
тическое распространение присуще также X. brasiliensis (Baker, 1904).
В общей сложности блохи рода Xenopsylla отмечены на 114 видах грызунов
60 родов из шести семейств, из которых виды хомяковых составляют половину
(56 видов из 17 родов). Всего же виды этого рода блох были отмечены на 178 видах
из 85 родов 19 семейств 10 отрядов млекопитающих. Имеется ряд находок блох это-
го рода с 18 видов птиц из 15 родов 13 семейств, среди которых, кроме нескольких
видов хищных, отмечались воробьинообразные, и, в частности, виды рода Hirundo
из ласточковых и рода Oenanthe из дроздовых. Среди видов рода Xenopsylla пред-
ставлены как узкоспецифичные моноксенные виды, так и виды с очень широким
кругом хозяев. В качестве примера последних можно указать блоху X. conformis
с широким европейско-азиатским (турано-ирано-центральноазиатским) ареалом, кото-
рая, кроме девяти видов песчанок рода Meriones, была в общей сложности обнаружена на
62 видах 38 родов восьми отрядов хозяев, включая даже таких случайных как летучие
мыши и птицы (табл. 2).
386
Таблица 1. Типы ареалов и паразито-хозяинные связи видов рода Xenopsylla фауны
Афротропической (АФ), Палеарктической (ПА), Индо-Малайской (ИМ) и Австралийской
(АВС) зоогеографических областей
Table 1. Types of ranges and host-parasite relations of fleas of the genus Xenopsylla
in Afrotropical (AT), Palearctic (PA), Indo-Malayan (IM), and Australian (AUS)
zoogeographical realm
Зоогеографические области
Типы ареалов
Виды блох
Космо-
АФ
ПА
ИМ
АВС
политы
Отр. RODENTIA - Грызуны
Сем. Cricetidae - Хомяковые
Род Tatera - Гололапые песчанки, татэры
Восточно-Африканские
Xenopsylla conformis coppensi,
11
X. cornigera, X. difficilis, X. humilis,
X. jorgei, X. lobengulai, X. pestanai,
X. raybouldi, X. silvai, X. syngenis
и X. tanganyikensis
Капские
X. frayi, X. geldenhuysi,
5
X. hipponax, X. mulleri
и X. phyllomae
Индийские
X. hussaini
1
Род Gerbillus - Карликовые песчанки
Восточно-Африканские
X. bantorum, X. debilis
2
Капские
X. demeilloni, X. h. hirsute,
4
X. hirsuta multisetosa, X. placidia
Восточно-
X. dipodilli
1
Средиземноморский
Западно-
X. blanci
1
Средиземноморский
Род Meriones - Малые, или гребенщиковые, песчанки
Иранский
X. buxtoni
1
Турано-Иранский
X. regis
1
Род Gerbillurus
Капский
X. davisi, X. sulcata
2
Род Desmodillus
Капско-Восточно-
X. trifaria
1
Африканский
Род Rhombomys - Большие песчанки
Туранский
X. nuttalli
1
387
Зоогеографические области
Типы ареалов
Виды блох
Космо-
АФ
ПА
ИМ
АВС
политы
Род Ellobius - Слепушонки
Туранский
X. magdalinae
1
Род Hypogeomys
Мадагаскарский
X. petteri
1
Сем. Dipodidae - Тушканчиковые
Род Dipus - Мохноногие тушканчики
Туранский
X. conformis dipodilis
1
Центрально-Азиатский
X. tarimensis
1
Сем. Gliridae - Соневые
Род Graphiurus - Африканские сони
Капский
X. hamula
1
Сем. Muridae - Мышиные
Род Aethomys - Акациевые крысы
Восточно-Африканский
X. cuisancei, X. georychi, X. torta
3
Восточно-Африканско-
X. versuta
1
Капский
Капский
X. scopulifer, X. zumpti
2
Род Saccostomus
Восточно-Африканский
X. angolensis, X. sarodes
4
manyarensis, X. s. sarodes,
X. sarodes serengetiensis
Капский
X. bechuanae
1
Род Mus - Домовые мыши
Трансавстралийский
X. australiaca
1
Малайский
X. nesiotes
1
Западно-
X. guancha
1
Средиземноморский
Род Acomys
Восточно-Африканский
X. morgandaviesi
1
Восточно-
X. acomydis
1
Средиземноморский
Род Parotomys - Иглистые мыши
Капский
X. occidentalis
1
388
Род Cricetomys
Восточно-Африканский
X. crinita
1
Род Thallomys
Восточно-Африканский
X. graingeri
1
Род Lemniscomys
Восточно-Африканский
X. robertsi
1
Род Melomys
Папуасский
X. papuensis
1
Сем. Sciuridae - Беличьи
Род Xerus
Капский
X. cryptonella
1
Сем. Bathyergidae - Землекоповые
Род Cryptomys
Капский
X. philoxera
1
Отр. LAGOMORPHA - Зайцеообразные
Сем. Leporidae - Зайцевые
Род Oryctolagus - Кролики
Западно-
X. cunicularis
1
Средиземноморский
Отр. CARNIVORA - Хищные
Сем. Canidae - Псовые
Род Vulpes - Лисицы
Восточно-Африканский
X. nubica
1
Капский
X. eridos, X. piriei
2
Сахаро-Аравийский
X. taractes
1
Западно-
X. conformis mycerini
1
Средиземноморско-
Сахаро-Аравийский
Туранский
X. g. gerbilli, X. gerbilli minax
2
Турано-Иранский
X. nesokiae, X. persica
2
Турано-Ирано-Сахаро-
X. astia
1
Аравийско-Индийский
Центрально-Азиатско-
X. c. conformi
1
Турано-Иранский
Род Canis - Волки
Космополитический
X. cheopis
1
389
Зоогеографические области
Типы ареалов
Виды блох
Космо-
АФ
ПА
ИМ
АВС
политы
Сем. Felidae - Кошачьи
Род Felis - Кошки
Восточно-Африканский
X. aequisetosa
1
Западно-
X. ramesis
1
Средиземноморско-
Сахаро-Аравийский
Сем. Herpestidae - Мангустовые
Род Cynictis - Желтые мангусты
Капский
X. erilli
1
Космополитический
X. brasiliensis
1
Сем. Mustelidae - Куньи
Род Mustela - Ласки и хорьки
Туранский
X. skrjabini
1
Сем. Viverridae - Виверровые
Род Genetta - Генеты
Восточно-Африканский
X. nilotica
1
Отр. ARTIODACTYLA - Парнокопытные
Сем. Bovidae - Полорогие
Род Damaliscus - Лиророгие бубалы
Космополитический
X. c. cheopis
1
Отр. PRIMATES - Приматы
Сем. Hominidae - Гоминиды
Род Homo - Люди
Сахаро-Аравийский
X. gratiosa
1
Отр. DASYUROMORPHIA - Хищные сумчатые
Сем. Dasyuridae - Сумчатые куницы
Род Dasycercus - Гребнехвостые сумчатые мыши
Малайско-Папуасско-
X. vexabilis
1
Центрально-
австралийский
390
Отр. APODIFORMES - Стрижеобразные
Сем. Apodidae - Стрижиные
Род Apus - Стрижи
Туранский
X. hirtipes
1
Отр. PROCELLARIIFORMES - Буревестникообразные
Сем. Procellariidae - Буревестниковые
Род Puffinus - Настоящие буревестники
Сахаро-Аравийский
X. gratiosa
1
Отр. PASSERIFORMES - Воробьинообразные
Сем. Hirundinidae - Ласточковые
Род Hirundo - Ласточки
Восточно-Африканский
X. trispinis tenuis
1
Капский
X. t. trispinis
1
Сем. Turdidae - Дроздовые
Род Oenanthe - Каменки
Туранский
X. gerbilli caspica
1
Род Xenopsylla был выделен в начале прошлого века (Glinkiewicz, 1907) (типо-
вой вид: X. pachyuromyidis (Glinkiewicz, 1907) = X. cheopis (Rothschild, 1903)). Боль-
шая часть видов и подвидов данного рода (59 из 85) была описана пятью авторами
в период с 1903 по 1960 г. Так, К. Джордан (K. Jordan) и Н. Ротшильд (N. Roth-
schild) по отдельности и совместно описали в общей сложности 21 вид и два подвида
из Афротропической, Индо-Малайской и Палеарктической областей, Де Мелон (De
Meillon) - 12 видов и один подвид из Афротропической области, Ф. Смит (F. Smit) -
шесть видов из Афротропической и Палеарктической областей. Отечественный автор
И.Г. Иофф (I. Ioff) описал пять видов и два подвида с территории бывшего СССР.
При этом на территории Палеарктической области из 18 видов и шести подвидов рода
Xenopsylla большая часть форм (15 видов и шесть подвидов) были описаны в период
с 1903 по 1960 годы; на территории Афротропической области из 44 видов и девяти
подвидов 46 форм (39 видов и семь подвидов) - с 1901 по 1964 год; с территории
Индо-Малайской области - четыре вида были описаны с 1908 по 1933 год. Следует
также отметить, что большая часть подвидов (восемь из 10) были описаны позднее -
во второй половине прошлого века, в период с 1950 по 1990 год.
391
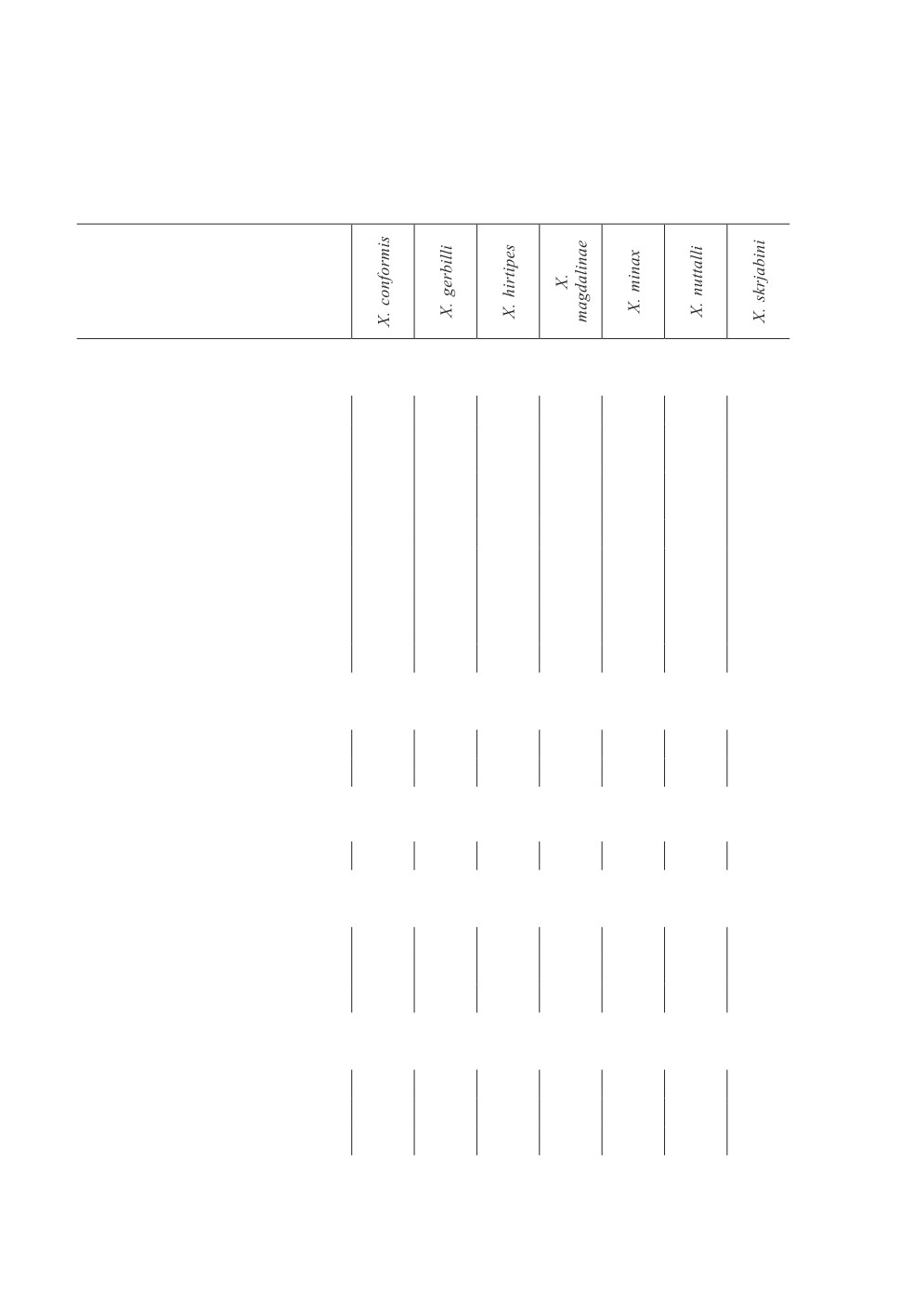
Таблица 2. Типы ареалов видов блох рода Xenopsylla и количество таксонов их хозяев -
млекопитающих и птиц - в различных зоогеографических областях мира
Table 2. Types of ranges of fleas of the genus Xenopsylla and the number of taxa of mammals
and birds in different zoogeographical regions
Количество таксонов хозяев
№
Типы ареалов и виды блох
Виды
Роды
Семейства
Отряд
Австралийская область
Трансавстралийские
1.
Xenopsylla australiaca
10
5
1
1
Афротропическая область
Восточно-Африканские
2.
X. nubica
34
21
9
4
3.
X. bantorum
12
9
4
2
4.
X. pestanai
5
5
2
1
5.
X. morgandavies
4
4
2
2
6.
X. nilotica
4
4
3
2
7.
X. syngenis
4
4
2
1
8.
X. aequisetosa
3
3
3
2
9.
X. debilis
3
3
2
1
10.
X. robertsi
3
3
1
1
11.
X. torta
3
3
1
1
12.
X. cornigera
2
2
2
1
13.
X. crinita
2
2
1
1
14.
X. georychi
2
1
1
1
15.
X. humilis
2
1
1
1
16.
X. jorgei
2
2
2
1
17.
X. angolensis
1
1
1
1
18.
X. conformis coppensi
1
1
1
1
19.
X. cuisancei
1
1
1
1
20.
X. difficilis
1
1
1
1
21.
X. graingeri
1
1
1
1
22.
X. lobengulai
1
1
1
1
23.
X. raybouldi
1
1
1
1
24.
X. sarodes manyarensis
1
1
1
1
25.
X. s. sarodes
1
1
1
1
26.
X. sarodes serengetiensis
1
1
1
1
27.
X. silvai
1
1
1
1
28.
X. tanganyikensis
1
1
1
1
392
29.
X. trispinis tenuis
1
1
1
1
Восточно-Африканско-Капские
30.
X. versuta
5
5
2
1
Капские
31.
X. philoxera
9
8
5
2
32.
X. piriei
7
6
3
2
33.
X. erilli
6
6
4
2
34.
X. eridos
5
4
3
2
35.
X. hipponax
5
5
2
1
36.
X. h. hirsuta
5
4
2
1
37.
X. hirsuta multisetosa
4
4
2
1
38.
X. hirsuta placidia
4
4
2
1
39.
X. zumpti
4
4
1
1
40.
X. frayi
3
2
2
1
41.
X. scopulifer
3
3
1
1
42.
X. davisi
2
2
1
1
43.
X. hamula
2
2
2
1
44.
X. sulcata
2
2
1
1
45.
X. bechuanae
1
1
1
1
46.
X. cryptonella
1
1
1
1
47.
X. demeilloni
1
1
1
1
48.
X. geldenhuysi
1
1
1
1
49.
X. mulleri
1
1
1
1
50.
X. occidentalis
1
1
1
1
51.
X. phyllomae
1
1
1
1
52.
X. trispinis trispinis
1
1
1
1
Капско-Восточноафриканские
53.
X. trifaria
3
3
1
1
Мадагаскарские
54.
X. petteri
1
1
1
1
Индо-Малайская область
Индийские
55.
X. hussaini
1
1
1
1
Малайские
56.
X. nesiotes
2
2
1
1
Малайско-Папуасско-Центральноавстралийские
57.
X. vexabilis
17
10
3
2
393
Количество таксонов хозяев
№
Типы ареалов и виды блох
Виды
Роды
Семейства
Отряд
Папуасские
58.
X. papuensis
4
4
1
1
Палеарктическая область
Сахаро-Аравийские
59.
X. taractes
8
7
5
3
60.
X. gratiosa
4
4
3
2
Восточно-Средиземноморские
61.
X. dipodilli
6
3
1
1
62.
X. acomydis
1
1
1
1
Западно-Средиземноморские
63.
X. blanci
6
3
2
1
64.
X. guancha
2
2
1
1
65.
X. cunicularis
1
1
1
1
Западно-Средиземноморско-Сахаро-Аравийские
66.
X. ramesis
25
8
8
4
67.
X. conformis mycerini
21
13
6
3
Иранские
68.
X. buxtoni
2
1
1
1
Туранские
69.
X. skrjabini
18
12
5
2
70.
X. gerbilli gerbilli
15
11
8
5
71.
X. gerbilli minax
13
9
6
3
72.
X. hirtipes
12
7
5
3
73.
X. magdalinae
4
3
2
1
74.
X. conformis dipodilis
3
3
2
1
75.
X. nuttalli
3
3
2
1
76.
X. gerbilli caspica
2
2
2
2
Турано-Иранские
77.
X. nesokiae
8
6
4
2
78.
X. persica
2
2
2
2
79.
X. regis
2
2
1
1
Турано-Иранско-Сахаро-Аравийско-Индийские
80.
X. astia
18
13
5
3
Центрально-Азиатские
81.
X. tarimensis
2
2
1
1
394
Центрально-Азиатско-Турано-Иранские
82.
X. conformis conformis
46
29
15
7
Космополитические
83.
X. brasiliensis
24
20
6
2
84.
X. cheopis
15
12
6
3
85.
X. cheopis cheopis
53
42
24
11
Всего
196
100
34
23
Блоху X. cheopis можно отнести к одному из наиболее изученных представителей
отряда блох. На примере этого вида были подробно изучены особенности строения
скелета и мускулатуры груди (Lewis, 1961), а также механизм прыжка блох (Rothschild,
Schlein, 1975). Наиболее детальные исследование и описание ультраструктуры пиги-
дия были также выполнены у блохи X. cheopis (Иванов, 1993). Пигидий - уникальная
структура, присущая только представителям отряда блох. Он находится на дорсальной
поверхности 10-го сегмента брюшка, а его нитевидные рецепторные волоски - трихо-
ботрии, и хордотональные сенсиллы способны воспринимать механические стимулы
небольшой интенсивности и, в частности, улавливать слабые потоки воздуха и зву-
ковые колебания воздушной среды. В целом, блохи рода Xenopsylla характеризуются
чертами высокой специализации: компактным, округлым и относительно коротким
телом с длинными прыгательными ногами. Блохи этого рода способны очень активно
перемещаться по телу хозяина и среди частиц субстрата гнезда и нор.
Следует указать особенности ряда видов рода Xenopsylla, распространенных
в других регионах мира. Так, например, один вид рода Xenopsylla обнаружен на труб-
коносых (буревестникообразные отряда Procellariformes) птицах. В частности, на бело-
лицем буревестнике (Puffinus (=Calonectris) leucomelas (Temminck, 1836)) паразитирует
Xenopsylla gratiosa Jordan et Rothschild, 1923. Ареал этого вида блох охватывает об-
ширную территорию от Канарских островов до побережья Туниса. На представителе
семейства ласточковых (Hirundinidae), кроме многих видов блох из других родов и
семейств, обнаружены блохи Xenopsylla trispinis Waterston, 1911. Этот вид, в частно-
сти, собран с гигантской (Hirundo senegalensis (L., 1766)) и южноафриканской горной
(H. spilodera Jerdon, 1841) ласточек в Южной Африке.
В Афротропической области различные виды рода Xenopsylla указываются
как постоянные паразиты мышиных, среди которых представители таких энде-
мичных родов как Thommomys, Oenomys, Arvicanthis и Rhabdomys, а также родов
395
Pelomys, Lemniscomys (в частности, барбарийской полосатой мыши (L. barbarus
(L., 1766)), Aethomys, Praomys и Lophuromys (желтоточечная жесткошерстная мышь
L. flavopunctatus Thomas, 1888). Афротропические пышнохвостые крысы (подсем. Cri-
cetomyinae) являются основными хозяевами для блох Xenopsylla scopulifer (Rothschild,
1905), X. aequisetosa (Enderlein, 1901) и X. crinite Jordan et Rothschild, 1922, а предста-
витель соневых (Gliridae) - африканская саванная соня (Graphiurus murinus (Desmarest,
1822)) - известна как специфический хозяин блохи X. hamula Jordan, 1925.
В целом, видам рода Xenopsylla присущ 21 тип ареалов (табл. 2). Среди них по-
ловину - 10 типов - составляют ареалы палеарктического типа, что обусловливает-
ся размерами и ландшафтно-зональным разнообразием этой части Евразии. Ареалы
Палеарктического типа присущи 18 видам и шести подвидам, распространенными
в аридных зонах. У 10 видов и пяти подвидов ареалы охватывают Турано-Иранскую
подобласть (девять из них эндемичные), и у восьми видов и одного подвида Среди-
земноморскую и, в меньшей степени, Сахаро-Аравийскую. В Сахаро-Аравийской,
наиболее бедной по составу фауны блох в Палеарктике, род Xenopsylla насчитывает
пять видов.
Африканские ареалы видов рода Xenopsylla менее разнообразны, но в этой области
распространена их большая часть - 44 вида и девять подвидов, которым присущи
пять типов распространения. В Индо-Малайской области представлено четыре вида
рода Xenopsylla, каждый из которых имеет свои тип ареала.
Из 44 афротропических видов рода Xenopsylla 25 имеют восточноафриканские
ареалы. Эти виды принадлежат к шести группами, среди которых наиболее полно
представлены группы «brasiliensis» (15 видов, из них девять - восточноафриканские)
и «nilotica» (12 видов, из них семь - восточноафриканские). Обе эти группы, а так-
же группы «hirsuta» и «trispinus», являются афротропическими эндемиками. Блохи
группы «nilotica» тесно связаны с песчанками рода Tatera, а группы «brasiliensis» -
с африканским мышиными.
Из 19 видов рода Xenopsylla представители группы «erilli» (насчитывает два вида)
и «eridos» (восемь видов) распространены в Капской подобласти. Эндемиками данной
подобласти являются также четыре вида группы «hirsuta». Еще один вид из этой
группы (X. petteri Lumaret, 1962) обитает за пределами Капской подобласти на Мада-
гаскаре, где его блохи паразитируют на громадном хомяке, или воалаво (Hypogeomys
antimena A. Grandidier, 1869). Такие виды из группы «brasiliensis» как X. bechuanae
De Meillon, 1947, X. hamula Jordan, 1925, X. scopulifer (Rothschild, 1905) и X. zumpti
Haeselbarth, 1963 распространены от Южной до Центральной Африки, а X. mulleri De
Meillon, 1947 из группы «nilotica» обнаружен во всех подобластях Афротропической
области. Виды группы «brasiliensis» паразитируют на таких мышиных как мешотча-
396
тые хомяки рода Saccostomus, акациевые крысы родов Aethomys и Thallomys, афри-
канские мыши рода Thammomys, африканские сони рода Graphiurus и кустарниковые
белки рода Paraxerus. Прокормителями блох из других, перечисленных выше групп,
служат песчанки родов Gerbillus, Tatera и ряда других.
В фауне Индо-Малайской подобласти представлены паразит индийской песчанки
(Tatera indica (Hardwicke, 1807)) - блоха Xenopsylla hussaini Sharif, 1930. На хищ-
ных сумчатых и кольцехвостых кускусах отмечены блохи X. vexabilis Jordan, 1925
с Малайско-Папуасско-Центральноавстралийским ареалом. Этот вид отмечен также
на различных видах мышиных. Хозяевами блох X. astia в этой области служат пес-
чанки родов Tatera, Meriones и Gerbillus. Из других видов группы «astia» Малайский
тип ареала имеет также блоха мышиных - X. nesiotes (Jordan et Rothschild, 1908),
а папуасский - паразит мышиных - блоха X. papuensis (Jordan, 1933).
В Австралийской области трансавстралийский ареал имеет паразит мышиных -
блоха X. australiaca Mardon et Dunnet, 1971. Этот же вид известен с тушканчиковых
мышей. Два вида рода Xenopsylla известны в Австралийской области с девяти ви-
дов крыс. Наиболее широко представлен на крысах (восемь видов) указанный выше
X. vexabilis.
С песчанками тесно связаны около 16 видов рода Xenopsylla. Подсемейство пес-
чанок (подсем. Gerbillinae) относится к сем. Muridae. Песчанки распространены
в полупустынях, пустынях и саваннах Старого Света: Передней, Средней, Малой,
Центральной и Южной Азии, Северном Прикаспии, Аравии и почти по всей Африке.
Они представлены 17 родами (Павлинов и др., 1990). Виды и подвиды рода Xeno-
psylla в общей сложности отмечались на песчанках, относимых к 10 родам (Кучерук,
Дарская, 1981): Tatera, Gerbilliscus, Gerbilurus, Desmodillus, Gerbillus, Pachyuromys,
Sekeetamys, Meriones, Psammomys и Rhombomys. При этом в Палеарктике они пара-
зитируют на песчанках родов Rhombomys и Meriones, в фауне Афротропической об-
ласти - на татерах (Tatera). Средиземноморско-Сахаро-Аравийское распространение
имеют паразиты песчанок Xenopsylla ramesis (Rothschild, 1904) и паразит мелких
млекопитающих - Xenopsylla astia Rothschild, 1911.
В фауне России и сопредельных стран из 10 видов рода Xenopsylla на песчанках
паразитируют семь. Это, в частности, X. с. conformis (Wagner, 1903), X. g. gerbilli
(Wagner, 1903), X. g. caspica Ioff, 1950, X. g. minax Jordan, 1926, X. hirtipes Rothschild,
1913, X. nuttalli Ioff, 1930, X. persica Ioff, 1946, X. regis (Rothschild, 1903) и X. skrjabini
Ioff, 1930. Все они относятся к преимущественно палеарктической группе видов «con-
formis». Виды X. nubica, X. conformis dipodis и X. skrjabini также часто отмечаются
на различных видах тушканчиков (Dipodidae) родов Jaculus, Dipus и Paradipus в от-
крытых ландшафтах Северной Африки, Передней и Средней Азии.
397
На территории Палеарктики распространен ряд небольших по объему семейств
мышиных, ведущих подземный образ жизни. Так, слепушонок рода Ellobius отно-
сят к подсем. Microtinae или выделяют в самостоятельное подсем. Ellobiinae, или
слепушонковых. В роде Ellobius выделяют 3-4 вида, распространенных в степной
зоне Евразии от Южной Украины до Тувы, Монголии и Северо-Западного Китая.
Специфическими паразитами этих зверьков являются Xenopsylla magdalinae Ioff, 1935.
На территории Кавказа X. magdalinae паразитирует на обыкновенной слепушонке
(Ellobius talpinus (Pallas, 1770)), которая в регионе является единственным предста-
вителем рода Ellobius. Однако на остальной части своего ареала блоха X. magdalinae
паразитирует на других видах этого рода.
Представители рода Xenopsylla, паразитирующие на песчанках, значительно разли-
чаются по частоте питания и яйцекладки в разные сезоны. В экспериментах выявлено,
что имаго X. conformis при температуре 5-7°C не размножаются и питаются 0.7 раз
в сутки, а при повышении температуры до 18-22°C питание учащается до 3.5 раза
в сутки (Дарская, 1970). При этом самки совершают 2.0-3.3 кладок яиц. Метаморфоз
протекает в норах за счет радиационного прогрева почвы (Дарская, 1977).
Виды рода Xenopsylla, паразитирующие на песчанках, -
переносчики возбудителя чумы
Ниже приведены сведения о распространении, специфичности паразито-хозяинных
связей видов рода Xenopsylla - паразитов песчанок Палеарктики и данные об обна-
ружении зараженными чумой особей блох в естественных условиях и их эпизоото-
логическом значении (Ралль, 1960; Каримова, Неронов, 2007; Гончаров и др., 2013;
Слудский, 2014).
Xenopsylla buxtoni Jordan, 1949
Иранский, Центральнопалеарктический, Азиатский (несибирский) тип ареала.
Ареал: Иран.
Хозяева: малые песчанки рода Meriones - персидская (M. persicus (Blanford, 1875)),
Виноградова (M. vinogradovi Heptner, 1931), ливийская, или краснохвостая (M. libycus
(Lichtenstein, 1823)), переднеазиатская (M. tristrami (Thomas, 1892)) (Hopkins et Roth-
schild, 1953; Klein et al., 1963; Фаранг-Азад, 1972).
Инфицированность возбудителем чумы: обнаружен зараженным в естественных
условиях на территории Ирана (Гончаров и др., 2013). Наряду со Stenoponia insperata
Weiss, 1930 является одним из основных переносчиков в Курдо-Иранском горно-
степном очаге (основные носители - песчанки: персидская, малоазийская, краснохво-
стая и Виноградова), расположенном на северо-востоке Ирака, западной части Ирана
и юго-востоке Турции (Варшавский, Козакевич, 1984). Высказано предположение
(Каримова, Неронов, 2007), что Xenopsylla buxtoni может быть одним из основных
переносчиков в Анатолийско-Армянском природном очаге чумы (с основными носи-
398
телями персидской песчанкой и анатолийским сусликом (Spermophilus xanthoprymnus
(Bennett, 1835)). В Турецко-Сирийском равнинном полупустынно-пустынном очаге
чумы, где, вероятно, основными носителями являются несколько видов песчанок,
основным переносчиком, скорее всего, также является Xenopsylla buxtoni (Каримова,
Неронов, 2007).
Xenopsylla conformis (Wagner, 1903)
Сахаро-Аравийско-Турано-Ирано-Центральноазиатский, Северо-Африканско-
Азиатский (несибирский), Южно-Транспалеарктический тип ареала. Ареал: Алжир,
Тунис, Египет, север Саудовской Аравии, Иордания, Сирия, Иран, Ирак, страны Закав-
казья, Нижнее Поволжье, Казахстан, пустыни Средней Азии, Афганистан, Внутренняя
Монголия до восточной части пустыни Гоби.
Хозяева: малые песчанки рода Meriones - Сундавелла (M. crassus (Sundevall,
1842)), ливийская, переднеазиатская, Виноградова, тамарисковая (M. tamariscinus
(Pallas, 1773)), когтистая, или монгольская (M. unguiculatus (Milne-Edwards, 1867)).
Другие песчанковые (подсем. Gerbillinae) - дневные песчанки рода Psammomys, жир-
нохвостые песчанка рода Pachyuromys, карликовые песчанки рода Gerbillus, пуши-
стохвостые песчанка рода Sekeetamys, а также индийская голопалая песчанка (Tatera
indica (Hardwicke, 1807)) (Wagner, Wassilief, 1933; Hoogstraal, Traub, 1965; Фаранг-
Азад, 1972; Lewis, 1972, 1967; Cooreman, 1973; Hastriter, Tipton, 1975; Klein et al., 1975
a, b; Misonne, 1977; Beaucournu, Hellal, 1977; Beaucournu, Kowalski, 1985; Beaucournu,
Launay, 1990). В общей сложности блохи обнаружены на 38 видах млекопитающих
21 рода из восьми семейств. Кроме случайных находок на птицах и хищных, блохи
этого вида отмечались также на ряде других семейств грызунов и, в частности, среди
тушканчиковых (Dipodidae) отмечались как хозяева тушканчик Хотсона (Allactaga hot-
soni Thomas, 1920), тушканчик Северцова (A. severtzovi Vinogradov, 1925), тушканчик-
прыгун (A. sibirica (Forster, 1778)), мохноногий тушканчик (Dipus sagitta (Pallas,
1773)), длинноухий тушканчик (Euchoreutes naso Sclater, 1891), тушканчик Бланфор-
да (Jaculus blanfordi (Murray, 1884)), а также обыкновенный емуранчик (Scirtopoda
telum (Lichtenstein, 1823)). Среди беличьих блохи Xenopsylla conformis были собраны
с тонкопалого суслика (Spermophilopsis leptodactylus (Lichtenstein, 1823)), даурского
(Spermophilus dauricus (Brandt, 1843)) и желтого (Spermophilus fulvus (Lichtenstein,
1823)) сусликов.
Подвиды: X. c. dipodis- на тушканчиках в Средней Азии. X. c. mycerini (Rothschild,
1904) - на песчанке (Gerbillus campestris (Loche, 1867)) в Египте, Алжире, Тунисе,
Сирии. X. c. conformis - на песчанках в Северной Африке, Передней (Сирийская пу-
стыня и север Саудовской Аравии), Средней и Центральной Азии, Казахстане (Тифлов
и др., 1977; Lewis, Lewis, 1990).
399
Среднегодовые индексы обилия этого подвида на краснохвостой песчанке в За-
кавказском равнинно-предгорном очаге чумы составляют 0.2-1.3 (Природные…, 2004).
Изучение многолетней динамики численности этого вида в Закавказском равнинно-
предгорном очаге чумы (Ширанович и др., 1977) позволило установить, что в наи-
большей степени на блох влияли погодные условия - в теплые и сухие годы числен-
ность имаго X. c. conformis была достоверно выше, чем в дождливые и холодные.
При этом для развития эпизоотий чумы в данном природном очаге численность рас-
сматриваемого вида оказалась одним из определяющих факторов.
Блохи этого вида в условиях эксперимента питались на белых мышах в среднем
один раз в двое суток, что значительно реже, чем у крысиных блох (Xenopsylla cheopis
и Nosopsyllus fasciatus (Bosc, 1800)). У большинства имаго проводили второе кровосо-
сание в течение одних суток после первого, но существенно чаще, чем у блох мало-
го суслика (через четверо суток у Citellophilus tesquorum (Wagner, 1898)) пившими
были 70.0% особей, а у Neopsylla setosa (Wagner, 1898) - 42.3%) (Новокрещенова и
др., 1968б).
Инфицированность возбудителем чумы: данный подвид считается основным пере-
носчиком чумы в Закавказье - в Приараксинском низкогорном очаге (основной но-
ситель - песчанка Виноградова). В Копетдагском пустынном очаге чумы (основные
носители - большая и краснохвостая песчанки) этот подвид является одним из двух
основных переносчиков инфекции, наряду с X. nuttalli Ioff, 1930. В Волго-Уральском
пустынном (основные носители - полуденная и гребенщиковая песчанки), а также
в Закавказском равнинно-предгорном очаге (основной носитель - краснохвостая пес-
чанка) X. c. conformis также один из двух основных переносчиков, вместе с блохой
Nosopsyllus laeviceps (Wagner, 1909) (Природные…, 2004; Каримова, Неронов, 2007).
В Урало-Эмбинском, Каракумском, Муюнкумском и Таукумском пустынных и Прика-
спийском песчаном природных очагах чумы X. c. conformis является второстепенным
переносчиком инфекции. В Прибалхашском пустынном очаге блохи этого подвида
паразитируют преимущественно на полуденной песчанке и относятся к случайным
переносчикам (Природные…, 2004; Каримова, Неронов, 2007). В Волго-Уральском
песчаном очаге чумы при изучении особенностей течения эпизоотий в 1997-1999 г.
установлено, что индексы доминирования и индексы обилия блохи X. conformis на
эпизоотических участках были статистически выше, чем на участках, где эпизоотий
не выявляли (Сараев и др., 2019).
В Ирано-Афганском низкогорном пустынном очаге чумы (основные носители -
большая и краснохвостая песчанки) данный подвид, вероятно, является одним из двух
основных переносчиков инфекции, наряду с X. nuttalli (Варшавский, Козакевич, 1984;
Каримова, Неронов, 2007).
400
В экспериментальных условиях, приближенным к естественным в норах большой
песчанки (температура +4°С, относительная влажность 80-95%), в серии опытов до-
казана способность большей части взятых в эксперимент блох X. conformis сохранять
в своем организме чумной микроб до 122 дней (срок наблюдений) и передавать воз-
будитель интактным зверькам (Акиев и др., 1968). В лабораторных условиях показана
способность этих блох образовывать блок преджелудка у 54-86% от всех зараженных
особей (Мокриевич и др., 1983).
В экспериментах при содержании инфицированных блох, проведенных в разные
сезоны года при температуре от 12 до 22°С, уровень блокообразования у данного
вида составлял: при кормлении на белых мышах 50-84.6%, на тамарисковых песчан-
ках - 27.2-37.0%; на краснохвостых песчанках - 36.0-65.3%. Средний срок жизни
блокированных блох составлял 2.3 суток (Загнибородова и др., 1979; Ващенок, 1984).
Xenopsylla gerbilli (Wagner, 1903)
Туранский, Центральнопалеарктическо-туранский тип ареала. Ареал: Передняя,
Средняя и Центральной Азии от Каспия до Южной Монголии и Северного Китая.
Паразитирует на песчанках, главным образом, на большой песчанке (Rhombomys
opimus (Lichtenstein, 1823)) (Hopkins, Rothschild, 1953; Иофф и др., 1965). Блохи Xe-
nopsylla gerbilli обнаружены на малом тушканчике и тушканчике Северцова, а также
на тонкопалом и желтом суслике.
Подвиды: Xenopsylla gerbilli (Wagner, 1903) - южные районы Туркмении и Узбе-
кистана, Иран; X. caspica Ioff, 1950 - Средняя Азия; X. minax Jordan, 1926 - Средняя
Азия, Казахстан, Китай.
Xenopsylla gerbilli caspica Ioff, 1950
Ареал: Средняя Азия, центральная часть пустынных ландшафтов, на севере и на
юге региона почти не встречается (Иофф и др., 1965).
В Северных Кызылкумах прослежены закономерности жизнедеятельности этой
блохи в пределах микробиотопа большой песчанки (Климова, 1972). Всего использо-
ваны данные по раскопке 210 нор большой песчанки и очесу 807 зверьков, с которых
собрано было 64579 имаго X. gerbilli caspica. Установлено, что сезонный ход чис-
ленности характеризуется двумя пиками - в июне-начале июля и в октябре-ноябре.
Размножение начинается в марте, первые молодые блохи появляются в мае. При
первом пике численности количество имаго на одну нору большой песчанки дости-
гает 1085-3010 насекомых (в среднем 671.4). В это же время молодые блохи массово
мигрируют ко входам нор и на поверхность, где их скопления достигают нескольких
сотен особей. Далее, в течение лета, численность имаго снижается, достигая годо-
вого минимума в сентябре. В октябре-ноябре размножающиеся самки встречаются
единично, подавляющее большинство имеет развитое жировое тело и реже питается.
401
В это время наблюдается второй пик численности, с увеличением количества имаго
на одну колонию в 1057-2134 особей (в среднем 520 блох на нору).
Показательно сезонное распределение блох по микробиотопу. Летом, при крайне
высоких наружных температурах воздуха и некомфортной температуре в нижних
горизонтах норы (на глубине 320 см этот показатель колеблется в пределах 11-16°С)
большинство блох концентрируется в кормовых камерах большой песчанки (72.5%
особей). На хозяевах находится 25.1% блох и лишь 1.7% встречаются в верхних хо-
дах норы. Осенью, когда глубинные горизонты почвы прогреваются до 17.2-17.6°С,
песчанки подготавливают норы к зимнему сезону. В это время основная часть блох
(88.5%) находится в кормовых камерах, остальные распределялись между нижними
ходами (2.4%), зимовочными гнездами (5%) и хозяевами (4.1%). Зимой, в гнездовых
камерах, расположенных в нижнем глубинном поясе, сосредотачивается 75.6% всех
имаго, в кормовых камерах, находящихся ближе к поверхности, находится только
13.6% этих насекомых. В весенний период поверхность почвы нагревается, и основная
часть блох мигрирует в кормовые камеры (66.9%), остальные по большей части рас-
пределяются между хозяевами (11.6%) и выводковыми гнездами (8.3%). Специально
подчеркивается, что подобная сложность поведения блох этого вида в плане измене-
ния предпочитаемой части используемого микробиотопа должна учитываться при про-
ведении учетных работ по оценке численности блох рода Xenopsylla (Климова, 1972).
В сериях масштабных опытов показано, что в апреле около 15% блох данного под-
вида единовременно находится в шерсти больших песчанок, в осенний период этот
показатель снижается до 7%. Средний период нахождения блохи в шерсти зверька
составляет в апреле 7.2 часа, в октябре - 16.8 часа (Солдаткина, Левошина, 1968).
Инфицированность возбудителем чумы: обнаружен зараженным в естественных
условиях (Приаралье) (Гончаров и др., 2013). В Каракумском пустынном очаге чумы
подвид является одним из трех основных переносчиков, наряду с X. g. gerbilli и
X. hirtipes (Каримова, Неронов, 2007). По другим данным данный подвид выступает
в роли основного переносчика только на Сарыкамышском участке Каракумского пу-
стынного очага и на большинстве участков (кроме крайнего севера) Кызылкумского
пустынного очага (Природные…, 2004).
В условиях эксперимента при кормлении на белых мышах уровень блокообразо-
вания у этой блохи варьировал при 18-20°С от 8.8 (осенью) до 28.8% (весной). При
содержании в условиях более высокой температуры (22-24°С) данный показатель
составил 41.0% (Ващенок, 1984).
Были изучены особенности блокообразования и инфекционного потенциала у блох
X. gerbilli caspica, взятых из разных популяций в Кызылкумском природном очаге
чумы (Сержанов и др., 1979). Для опытов использован местный вирулентный штамм
402
чумы, подкормки проводили на белых мышах. Блохи для экспериментов собраны
с колоний большой песчанки из аллювиальной равнины старого русла Сыр-Дарьи
(урочище Достанбек), грядово-ячеистых песков центральной части очага (урочище
Жалаулы) и побережья Аральского моря (урочище Шеймахан). Первый участок ха-
рактеризовался короткими межэпизоотическими периодами (3-4 года) и стойким со-
хранением эпизоотий чумы, во втором эпизоотии протекали лишь периодически и
имели эфемерный характер, на третьем участке возбудитель чумы не обнаруживался.
В результате проведенных опытов было установлено, что инфекционный потенциал
у блох из разных популяций неодинаков. У блох достанбекской популяции интен-
сивность блокообразования оказалась значительно выше (54%), чем у насекомых из
жалаулинской популяции (40%) и шейхаманской (21%). Блохи X. gerbilli caspica из
достанбекской и жалаулинской популяций оказались способными к длительному со-
хранению возбудителя чумы в своих организмах. Так, на 32-е сутки эксперимента
(срок наблюдения) у достанбекской группы блокированными оказалось 12.5% блох,
у жалаулинской - 2.5%, у шейхаманской популяции образование блоков полностью
прекратилось на 28-е сутки.
Xenopsylla gerbilli gerbilli (Wagner, 1903)
Ареал: южные и западные районы Туркмении и Узбекистана, Иран (Иофф и др.,
1965; Фаранг-Азад, 1972).
В Центральных Кызылкумах имаго паразитирует на больших песчанках в течение
всего года. Численность вида значительно варьирует по сезонам и годам - от 1.0
до 8.3 на одну песчанку и от 1.0 до 96.2 на одну колонию. Максимум численности
приходится на период с февраля по апрель. Размножение происходит в течение всего
года. Массовый выплод молодых блох регистрируется с июня по декабрь. В сравнении
с X. hirtipes проявляют несколько большую привязанность к хозяину (Загнибородова,
1968).
Инфицированность возбудителем чумы: обнаружен зараженным в естественных
условиях на территории Туркмении (Гончаров и др., 2013). Является основным пере-
носчиком на участке юго-востока Каракумов в Каракумском пустынном очаге чумы
(Природные…, 2004). Относится к второстепенным переносчикам в Копетдагском
пустынном очаге чумы и случайным, в силу низкой численности, в Таукумском пу-
стынном очаге (Природные…, 2004).
Xenopsylla gerbilli minax Jordan, 1926
Ареал: Средняя Азия, Казахстан, Китай (Иофф и др., 1965).
В Бетпакдалинском природном очаге чумы (Южно-Казахстанская и Карагандин-
ская области Республики Казахстан) основными носителями считаются два вида -
большая и краснохвостая песчанка (Природные…, 2004). Среди 110 штаммов воз-
403
будителя чумы, полученных от грызунов за период с 1983 по 2013 г., от этих видов
изолировано 51 и 54 культуры чумного микроба, соответственно. Из 90 штаммов, вы-
деленных от блох, 67% получено от блох большой песчанки. Среди трех выделяемых
ландшафтно-эпизоотологических районов (ЛЭР) в Шолакеспинском основными пере-
носчиками считаются X. gerbilli minax и Coptopsylla lamellifer (Wagner, 1895); в За-
падном Бетпакдалинском Xenopsylla gerbilli minax, Coptopsylla lamellifer и Nosopsyllus
laeviceps; в Каракойынском - Xenopsylla skrjabini и Coptopsylla lamellifer, а Xenopsylla
gerbilli minax на его территории вообще не встречается (Сажнев и др., 2018).
Подвид является основным переносчиком на участке песков Арыскумы (Заараль-
ский песчаный очаг) и в Муюнкумском песчаном очаге чумы. В Прибалхашском
пустынном очаге данный подвид играет роль одного из трех основных переносчиков
чумы, наряду с X. skrjabini и X. hirtipes (Природные…, 2004).
При экспериментальном заражении 61 особи больших песчанок блохами данного
подвида, инфицированными чумой, 41 грызун погиб, в 17 случаях от павших зверь-
ков удалось выделить чистую культуру чумного микроба (Бибикова и др., 1968а).
При кормлении инфицированных блох этого вида на белых мышах при температуре
22-24°С уровень блокообразования составил 50.0%. Инфицированные блохами боль-
шие песчанки погибали от генерализованной инфекции в 20.0-28.3%, полуденные
песчанки - в 7.2% случаев (Ващенок, 1984).
Xenopsylla hirtipes Rothschild, 1913
Туранский, Азиатско (несибирские), Центральнопалеарктическо-туранский тип
ареала. Ареал: Средняя Азия и Китай (Синьцзян-Уйгурский автономный район).
Хозяева: паразитирует, преимущественно, на большой песчанке, реже встречаясь
на других мелких млекопитающих - представителях пустынной фауны Средней и
Центральной Азии. В частности, был собран с таких видов песчанок как Meriones
chengi Wang, 1964 и M. erythrourus (Gray, 1842), полуденной (M. meridianus (Pallas,
1773)), тамарисковой и когтистой песчанок, а также с тонкопалого и желтого сусликов.
Предпочитает биотопы с закрепленными и слабо закрепленными песками, на
участках с твердым грунтом встречается редко (Иофф и др., 1965; Гончаров и др.,
1989; The Atlas…, 2000; Гончаров и др., 2013).
В Центральных Кызылкумах имаго паразитирует на больших песчанках в течение
всего года. Численность вида значительно варьирует по сезонам и годам - от 0.8 до
8.5 на одну песчанку и от 0.8 до 122.2 на одну колонию. Максимум численности
приходится на период с февраля по апрель. Размножение происходит в течение всего
года. Массовый выплод молодых блох регистрируется с июня по декабрь (Загнибо-
родова, 1968).
404
Инфицированность возбудителем чумы: основной переносчик на большинстве
участков Каракумского и Каракумского песчаных очагов чумы и второстепенный
в Муюнкумском песчаном очаге. В Таукумском пустынном очаге чумы данный вид
является одним из двух основных переносчиков инфекции, наряду с X. skrjabini.
В Прибалхашском пустынном очаге X. hirtipes - одни из трех основных переносчиков
чумы, наряду с блохами X. skrjabini и X. gerbilli minax (Природные…, 2004; Каримова,
Неронов, 2007).
В условиях эксперимента инфицированные чумой блохи этого вида выживали при
температуре 8-10°С около одного месяца, при 20-22°С - 17.3 дня, при 25-28°С -
7.2 дня. При этом блохи, содержащиеся при низкой температуре, не образовывали
блока преджелудка, а среднее число микробных клеток на одну блоху составляло
391755. При температуре 20-22°С за подкормку блокировалось 2% взятых в опыт
блох с содержанием возбудителя в одном насекомом в среднем 811627 микробных
клеток (максимальный показатель - 22.6 млн м. к.). При культивировании в условиях
высоких температур блокировалось 5.2% всех блох, но число микробных тел на одну
особь составляло лишь 19645 (Бибикова и др., 1968а).
В нескольких экспериментах, при содержании при температуре 11-22°С и под-
кормках на белых мышах, инфицированные чумой блохи X. hirtipes образовывали
блоки от 51.2 до 75.5% случаев, при подкормках на больших песчанках - от 51.0
до 65.0% (Загнибородова и др., 1979). Это свидетельствует о незначительном влия-
нии фактора прокормителя в сезонном аспекте формирования агрегированных форм
чумного микроба. Средний срок жизни блокированных блох этого вида составлял
2.1 суток.
Xenopsylla hussaini Sharif, 1930
Ирано-Индийский, Центрально-Палеарктическо-Западно-Индо-Малайский тип
ареала. Ареал: Иран, Северная Индия.
Основным прокормителем этой блохи является индийская голопалая песчанка
(Hopkins et Rothschild, 1953; Фаранг-Азад, 1972).
Инфицированность возбудителем чумы: не установлена.
Xenopsylla nuttalli Ioff, 1930
Туранский, Азиатский (несибирский), Центральнопалеарктическо-Туранский тип
ареала. Ареал: Передняя, Средняя Азия и Казахстан. От предгорий западной части
Копет-Дага до южных частей Устюрта и от побережья Каспия до Заузбойского плато
на востоке (Иофф и др., 1965).
Хозяева: на большой песчанке в западной части ее ареала, а также с тонкопалого
и желтого сусликов.
Инфицированность возбудителем чумы: обнаружен зараженным в естественных
условиях на территории Туркмении (Гончаров и др., 2013). На участке Красноводского
405
плато Каракумского пустынного очага чумы является одним из двух основных пере-
носчиков вместе с X. hirtipes (Природные…, 2004). В Копетдагском пустынном очаге
чумы данный вид является одним из двух основных переносчиков инфекции, наряду
с X. conformis (Природные…, 2004). В Ирано-Афганском низкогорном пустынном
очаге чумы (основные носители - большая и краснохвостая песчанки) данный вид,
вероятно, один из двух основных переносчиков инфекции, наряду с X. c. conformis
(Варшавский, Козакевич, 1984; Каримова, Неронов, 2007).
В нескольких экспериментах при содержании при температуре 11-22°С и под-
кормках на белых мышах инфицированные чумой блохи X. nuttalli образовывали
блоки от 47.5 до 78.0% случаев, при подкормках на больших песчанках - от 28.3
до 67.0% (Ващенок, 1984). В опытах, проведенных в летний период с подкормками
насекомых на белых мышах, при достаточно большой выборке (200 особей) у этого
вида был получен уровень блокообразования в 100% (Загнибородова и др., 1979).
Продолжительность блокообразования у блох X. nuttalli составила от 2 до 12 суток.
Блокированные блохи жили в среднем 2.7 суток.
Xenopsylla persica Ioff, 1946
Турано-Иранский, Азиатский (несибирский), Центральнопалеарктический тип ареа-
ла. Ареал: Туркмения и Афганистан - горы Большой Балхан, Копет-Даг, предгорья
Паропамиза.
Хозяева: основной хозяин - краснохвостая персидская песчанка (Иофф и др.,1965;
Фаранг-Азад, 1972).
Инфицированность возбудителем чумы: не установлена.
Xenopsylla regis (Rothschild, 1903)
Турано-Иранский, Азиатский (несибирский), Центральнопалеарктический тип
ареала. Ареал: Передняя и Средняя Азия - Йемен, Туркмения.
Хозяева: преимущественно на королевской песчанке (Meriones rex Yerbury et Thom-
as, 1895). Известен также с большой песчанки (Иофф и др., 1965; Lewis, Lewis, 1990).
Инфицированность возбудителем чумы: не установлена.
Xenopsylla skrjabini Ioff, 1930
Турано-Иранский, Азиатско (несибирский), Центральнопалеарктический тип ареа-
ла. Ареал: Средняя и Центральная Азия - от Каспийского моря до Восточной Мон-
голии, Синьцзян-Уйгурского автономного района и провинции Внутренняя Монголия
в Северном Китае.
Хозяева: преимущественно на большой песчанке, попадается в сборах с других ви-
дов мелких млекопитающих, свойственных фауне бореальных пустынь и полупустынь
Палеарктики. Находки также имеются с хомяка Эверсмана (Cricetulus eversmanni
(Brandt, 1859)) и серого хомячка (C. migratorius (Pallas, 1773)), полуденной, тамари-
406
сковой и когтистой песчанок, а также большой песчанки и тарбаганчика (Pygeretmus
pumilio (Kerr, 1792)), малого суслика (Alactagulus pygmaeus (Pallas, 1779)), гобийского
тушканчика (A. bullata G. Allen, 1925), тушканчика-прыгуна, пятипалого карликового
тушканчика (Cardiocranius paradoxus Satunin, 1903), мохноногого тушканчика, обык-
новенного емуранчика и желтого суслика (Иофф и др., 1965; Гончаров и др., 1989;
The Atlas…, 2000).
В Урало-Эмбинском природном очаге чумы среднемноголетняя численность оцени-
вается в 1000-1200 имаго на 1 га, в Приаральско-Каракумском - в 500-600 имаго на
1 га (Природные…, 2004). В условиях Приаральских Каракумов показано (Золотова
и др., 1975), что средняя численность этого вида по годам значительно варьирует.
Так, в 1969 г. она составляла 2490 блох на 1 га, в 1970 г. она снизилась почти вдвое
(1796 особей на 1 га), а в 1971 г. сохранилась почти на уровне предыдущего года
(1431 блоха). При наблюдениях за генеративной активностью данного вида установ-
лено, что ранняя и теплая весна вызывает массовое включение блох в размножение.
Холодная и затяжная весна сдвигает начало размножения на более поздние сроки.
Откладка яиц продолжается в течение всего теплого периода года - с апреля по
сентябрь, максимальные показатели отмечаются в мае, в сентябре откладывают яйца
только единичные блохи (Золотова и др., 1975).
Характерными особенностями Xenopsylla skrjabini в условиях Урало-Эмбинского
природного очага чумы были достаточно стабильная, по сравнению с другими пред-
ставителями рода Xenopsylla, численность и преимущественная концентрация в верх-
них горизонтах нор большой песчанки (Старожицкая, 1970). По представленным дан-
ным, подавляющее большинство имаго этого вида встречается по ходам на глубине
20-50 см от поверхности и даже зимой этот вид не обнаруживается глубже 70 см.
Инфицированность возбудителем чумы: в Урало-Эмбинском, Северо-Приаральском,
Приаральско-Каракумском, Зааральском (участок Дарьялыктакыр), Мангышлакском,
Устюртском и Предустюртском пустынных очагах чумы является единственным
основным переносчиком инфекции. В Кызылкумском пустынном очаге считается
основным переносчиком в северной его части (в пределах Республики Казахстан).
В Таукумском пустынном очаге чумы данный вид является одним из двух основ-
ных переносчиков инфекции, наряду с X. hirtipes. В Прибалхашском пустынном
очаге данный подвид является одним из трех основных переносчиков чумы, наряду
с X. gerbilli minax и X. hirtipes. В силу невысокой численности отмечен как второ-
степенный переносчик в Муюнкумском песчаном очаге чумы (Природные…, 2004;
Каримова, Неронов, 2007; Гончаров и др., 2013).
В Монголии, среди 46 описанных природных очагов чумы (Вержуцкий, Адъя-
сурэн, 2019), в подавляющем большинстве эпизоотический процесс обеспечивается
407
сурками и их блохой Oropsylla silantjewi (Wagner, 1898). Только два очага полностью
связаны с песчанками - Южно-Гобийский (основной носитель - большая песчанка) и
Замын-Удский (основной носитель - монгольская песчанка). В первом единственным
основным переносчиком является блоха Xenopsylla skrjabini, во втором - к основным
переносчикам относят X. skrjabini, Nosopsyllus laeviceps и Neopsylla pleskei Ioff, 1928.
Замын-Удский природный очаг чумы является трансграничным, на территории
Китая он обозначается как Эрлянский. Считается, что перечисленные для монголь-
ской части очага виды блох также являются основными переносчиками на китайской
стороне, но по соотношению выделяемых культур на первом месте стоит Nosopsyllus
laeviceps (34.4% от всех выделенных от эктопаразитов штаммов чумного микроба),
на втором - Xenopsylla conformis (24.7%) и на третьем - Neopsylla pleskei (6.0%).
Среди других природных очагов чумы в Китае имеется еще один природный очаг
песчаночьего типа - Джунгарский песчаный, где наиболее массовый вид грызунов
(большая песчанка) является и основным носителем чумы. Здесь единственным основ-
ным переносчиком считается Xenopsylla skrjabini. На большую песчанку и этот вид
блох приходилось 92.3% от всех штаммов возбудителя чумы, выделенных в этом
очаге (Вержуцкий, 2022). Кроме того, в Китае имеется ряд очагов чумы крысиного
типа, с основным переносчиком X. cheopis, эти очаги будут рассмотрены в отдельной
публикации.
На территории Приаральско-Каракумского природного очага чумы, численность
данного вида стабильна и оценивается в 35-70 тысяч имаго на 1 км2 (Айсауэтов
и др., 2019).
В опытах при содержании 238 блох этого вида при температуре 14-16°С и корм-
лении через 3-4 дня на больших песчанках и малых сусликах уровень блокообразо-
вания составил 24.7%, при ежедневных (или через день) подкормках 132 блох этот
показатель достиг 54.0%. При содержании в условиях более высокой температуры
(20-24°С) картина изменилась - при подкормках через три дня в опыт взято 212 блох)
блокировалось 62.0% особей, при ежедневных или через день в опыте задействовано
131 блоха) - уровень блокообразования снизился до 45.9% (Бибиковой и др., 1968б).
В опытах показано, что уровень блокообразования у блох этого вида при содер-
жании от 16.0 до 21.0°С в существенной степени зависел от длительности подкормок
насекомых - при увеличении продолжительности подкормки с 3 до 18-24 часов и со-
кращении интервала между подкормками с двух до одних суток число блокированных
особей возрастало с 20-40 до 80%. При условиях питания, близких к природным,
этот показатель был близок к 50%, практически не меняясь по сезонам. При этом при
кормлении на белых мышах уровень блокообразования в разных опытах варьировал
от 50.0 до 98.5%, при подкормках на больших песчанках - от 16.4 до 81.5% (Ново-
крещенова и др., 1968а).
408
Обсуждение
Роль каждого вида блох в природном очаге чумы может быть оценена по совокуп-
ности данных. Наибольшее значение при этом придают паразитированию имаго на
основных носителях чумы, частоте и регулярности встреч зараженных возбудителем
особей в природе, сезонности существования имаго, активности их нападения на
хозяев, питания и размножения, способности трансмиссии возбудителя чумы.
Сведения о представителях рода Xenopsylla, паразитирующих на песчанках, осно-
ваны, главным образом, на данных о распределении имаго между телом хозяина и
его убежищем. В природе в холодное время года имаго Xenopsylla неактивны и на-
ходятся в ходах нор песчанок, а весной с наступлением тепла начинают интенсивно
нападать на хозяина.
В 18 природных очагах чумы песчаночьего типа на территории России и сопредель-
ных стран блохи рода Xenopsylla являются основными переносчиками (табл. 3). Это,
в частности, блоха X. skrjabini в Урало-Эмбинском пустынном, Предустюртском пу-
стынном, Устюртском пустынном, Северо-Приаральском пустынном, Арыскумско-
Дарьялыктакырском (Зааральском) пустынном, Мангышлакском пустынном,
Приаральско-Каракумском пустынном, Прибалхашском пустынном, Таукумском
пустынном очагах; X. hirtipes - в Каракумском пустынном очаге; X. g. gerbilli -
в Копетдагском пустынном очаге; X. g. minax - в Бетпакдалинском и Мойынкумском
пустынном очагах; X.conformis - в Волго-Уральском песчаном, Приараксинском низ-
когорном и Закавказском равнинно-предгорном очагах; X. gerbilli - в Арыскумско-
Дарьялыктакырском пустынном очаге. Только в Прикаспийском песчаном очаге основ-
ной переносчик - это Nosopsyllus laeviceps (Кадастр…, 2016).
Естественная зараженность возбудителем чумы отмечена для 13 видов рода Xeno-
psylla - паразитов песчанок: Xenopsylla astia, X. buxtoni, X. c. conformis, X. eridos
(Rothschild, 1904), X. gerbilli caspica, X. g. gerbilli, X. g. minax, X. h. hirsuta Ingram,
1928, X. hirtipes, X. nubica (Rothschild, 1903), X. nuttalli, X. philoxera Hopkins, 1949,
X. piriei Ingram, 1928, X. skrjabini, X. versuta Jordan, 1925.
Способность передавать возбудителя чумы в эксперименте установлена для
X. astia, X. skrjabini, X. hirtipes, X. nuttalli, X. conformis, X. gerbilli caspica и X. minax.
Они относятся к высокоактивным переносчикам, за исключением активного перенос-
чика X. astia (Ващенок, 1988, 1997). В песчаночьих очагах, для которых характерна
наибольшая интенсивность эпизоотий весной и осенью, роль основного перенос-
чика отводят блохам рода Xenopsylla, в соответствии с их высокой численностью,
круглогодичным паразитированием на основном носителе и способностью активно
передавать возбудителя.
Таким образом, блохи рода Xenopsylla участвуют в эпизоотическом процессе во
многих природных очагах чумы (всего их описано более ста), входя в пул основных
переносчиков, обеспечивая непрерывность эпизоотического процесса.
409
Таблица 3. Значение видов рода Xenopsylla как основных (О), второстепенных (В),
случайных (С) переносчиков возбудителей чумы в природных очагах разных типов
на территории России и сопредельных стран
Table 3. Significance of species of the genus Xenopsylla as the main (М), secondary (В),
and occasional (С) vectors of plague pathogens in natural foci in the territory of Russia
and adjacent countries
Название очага
Кавказ и Закавказье
Песчаночьи очаги
Бозчёльский равнинно-предгорный
О
Гянджа-Казахский равнинно-
О
предгорный
Джейранчельский равнинно-
О
предгорный
Иорский равнинно-предгорный
О
Кобыстанский равнинно-
О
предгорный
Мильско-Карабахский равнинно-
О
предгорный
Приараксинский низкогорный
С, О
С
Северо-Западный и Северный Прикаспий
Песчаночьи очаги
Волго-Уральский песчаный
О
Прикаспийский песчаный
С
Сусликовые очаги
Зауральский степной
С
С
Казахстан
Песчаночьи очаги
Бетпакдалинский пустынный
С
С
С
Илийский межгорный
С
О
О
О
Приалакольский низкогорный
С
О
О
Средняя Азия
Песчаночьи очаги
Зааральский пустынный
С
С
С
О
О
Каракумский пустынный
С
С
С
С
Копетдагский пустынный
О
С
О
410
Кызылкумский пустынный
С
В, С
В, С
С
В, С
Мангышлакский пустынный
В
С
О
Муюнкумский пустынный
С
С
С
О
С
Е
(=Мойынкумский)
Предустюртский пустынный
С
О
Приаральско-Каракумский
С
С
С
О
пустынный
Прибалхашский пустынный
С
С
О
О
О
Северо-Приаральский пустынный
С
С
С
Таукумский пустынный
С
О
О
О
Урало-Эмбенский пустынный
В
О
Устюртский пустынный
С
О
О
Подводя итоги, можно заключить, что блохи рода Xenopsylla выполняют ведущую
роль в эпизоотическом процессе во многих природных очагах чумы на территории
Средней и Центральной Азии, Ближнего Востока. Наиболее древние связи с возбу-
дителем чумы, вероятно, имеют блохи сурков рода Oropsylla (Медведев, Вержуцкий,
2019). Представители рода Xenopsylla столкнулись с этим патогеном в более позднее
время, но их экологические и физиологические особенности оказались исключительно
благоприятными для существования чумного микроба, что, в значительной степени
определило широкое распространение возбудителя путем транспортировки заражен-
ных блох этого рода и их прокормителей по многим регионам планеты во время
третьей пандемии.
БЛАГОДАРНОСТИ
Работа выполнена на базе коллекции Зоологического института РАН (ЗИН РАН)
(УФК ЗИН рег. № 2-2.20) и Ставропольского противочумного института при финан-
совой поддержке темы Министерства науки и высшего образования «Современные
основы систематики и филогенетики паразитических и кровососущих членистоногих»
(Гос. Регистрационный номер 122031100263-1).
Список литературы
Айсауытов Б.Н., Толенбай Г.К., Бекжан Г.Е., Жадырасын С.Д., Суйндиков Е.А., Сердалы Ш.Ш., Айхожаев
А.Т., Жасмамбет М.Б. 2019. Данные многолетнего мониторинга за чумой Казалинским ПЧО. Каран-
тинные и зоонозные инфекции в Казахстане 1 (38): 42-45. [Ajsauytov B.N., Tolenbaj G.K., Bekzhan
G.E., Zhadyrasyn S.D., Sujndikov E.A., Serdaly Sh.Sh., Ajhozhaev A.T., Zhasmambet M.B. 2019. Dannye
mnogoletnego monitoringa za chumoj Kazalinskim PCHO. Karantinnye i zoonoznye infekcii v Kazahstane.
1 (38): 42-45. (in Russian)].
411
Акиев А.К., Деревянченко К.И., Лалазаров Г.А., Новикова Е.И. 1968. Сохранение возбудителя чумы в бло-
хах Xenopsylla conformis в зимний межэпизоотический сезон. Грызуны и их эктопаразиты. Саратов,
211-216. [Akiev A.K., Derevyanchenko K.I., Lalazarov G.A., Novikova E.I. 1968. Sohranenie vozbuditelya
chumy v blohah Xenopsylla conformis v zimnij mezhepizooticheskij sezon. Gryzuny i ih ektoparazity.
Saratov, 211-216 (in Russian)].
Бибикова В.А., Анисимова Т.И., Егорова Р.П., Айкимбаев М.А., Волохов В.А. 1968а. К вопросу об эпи-
зоотологической роли блох песчанок. Сообщение 2. Xenopsylla hirtipes et X. gerbilli minax. Грызуны
и их эктопаразиты. Саратов, 222-227. [Bibikova V.A., Anisimova T.I., Egorova R.P., Ajkimbaev M.A.,
Volohov V.A. 1968a. K voprosu ob epizootologicheskoj roli bloh peschanok. Soobshchenie 2. Xenopsylla
hirtipes et X. gerbilli minax. Gryzuny i ih ektoparazity. Saratov, 222-227. (in Russian)].
Бибикова В.А., Медведевских В.И., Данков С.С. 1968б. К вопросу об эпизоотологической роли блох пес-
чанок. Сообщение 4. Xenopsylla skrjabini. Грызуны и их эктопаразиты. Саратов, 228-232. [Bibikova
V.A. Medvedevskih V.I., Dankov S.S. 1968b. K voprosu ob epizootologicheskoj roli bloh peschanok.
Soobshchenie 4. Xenopsylla skrjabini. Gryzuny i ih ektoparazity. Saratov, 228-232. (in Russian)].
Варшавский С.Н., Козакевич В.П. 1984. Биоценотическая структура и ландшафтные особенности зарубеж-
ных очагов чумы в Передней и Юго-Западной Азии. Бюллетень МОИП, отд. биол. 89 (1): 13-20.
[Varshavskij S.N., Kozakevich V.P. 1984. Biocenoticheskaya struktura i landshaftnye osobennosti zarube-
zhnyh ochagov chumy v Perednej i Yugo-Zapadnoj Azii. Byulleten’ MOIP, otd. biol. 89 (1): 13-20. (in
Russian)].
Ващенок В.С. 1984. Блохи и возбудители бактериальных болезней человека и животных. Паразитологиче-
ский сборник 32: 79-123. [Vashchenok V.S. 1984. Blohi i vozbuditeli bakterial’nyh boleznej cheloveka
i zhivotnyh. Parazitologicheskij sbornik 32: 79-123. (in Russian)].
Ващенок В.С. 1988. Блохи - переносчики возбудителей болезней человека и животных. Л., Наука, 163 с.
[Vashchenok V.S. 1988. Fleas - vectors of human and animal diseases. L.: Nauka, 163 ss. (in Russian)].
Ващенок В.С. 1997. Экология блох группы видов conformis (Siphonaptera; Pulicidae: Xenopsylla) фауны
России и сопредельных стран (обзор). Паразитология 31 (6): 492-513. [Vashchenok V.S. 1997. The
ecology of the fleas conformis species group (Siphonaptera: Pulicidae; Xenopsylla of Russia and neighbour-
ing countries fauna (review). Parasitologiya 31 (6): 492-513. (in Russian)].
Вержуцкий Д.Б. 2022. Природные очаги чумы Китая: аннотированный список. Байкальский зоологический
журнал 2 (32): 135-145. [Verzhuckij D.B. 2022. Prirodnye ochagi chumy Kitaya: annotirovannyj spisok.
Bajkal’skij zoologicheskij zhurnal 2 (32): 135-145. (in Russian)].
Вержуцкий Д.Б., Адъясурэн З. 2019. Природные очаги чумы в Монголии: аннотированный список. Бай-
кальский зоологический журнал 2 (25): 92-103. [Verzhuckij D.B., Ad”yasuren Z. 2019. Prirodnye ochagi
chumy v Mongolii: annotirovannyj spisok. Bajkal’skij zoologicheskij zhurnal 2 (25): 92-103. (in Russian)].
Вержуцкий Д.Б., Вержуцкая Ю.А., Холин А.В., Медведев С.Г. 2021. Граница ареалов двух подвидов блох
- паразитов сусликов (Citellophilus tesquorum sungaris и Citellophilus tesquorum altaicus). Байкальский
зоологический журнал 1 (29): 116-120. [Verzhutsky D.B., Verzhutskaya Ju.A., Kholin A.V., Medvedev
S.G. 2021. The boundary of the areas of two subspecies of fleas - parasites of Ground squirrels (Citel-
lophilus tesquorum sungaris and Citellophilus tesquorum altaicus). Baikalskij zoologičeskij žurnal 1 (29):
116-120. (in Russian)].
Гончаров А.И., Ромашева Т.П., Котти Б.И., Баваасан А., Жигмид С. 1989. Определитель блох Монгольской
Народной Республики. Улан-Батор, 417 с. [Goncharov A.I., Romasheva T.P., Kotti B.I., Bavaasan A.,
ZHigmid S. 1989. Opredelitel' bloh Mongol’skoj Narodnoj Respubliki. Ulan-Bator, 417 s. (in Russian)].
412
Гончаров А.И., Тохов Ю.М., Плотникова Е.П., Артюшина Ю.С. 2013. Список видов и подвидов блох, об-
наруженных зараженными возбудителем чумы в естественных условиях. Ставрополь, РИО ИДНК,
34 с. [Goncharov A.I., Tohov Yu. M., Plotnikova E.P., Artyushina Yu. S. 2013. Spisok vidov i podvidov
bloh, obnaruzhennyh zarazhennymi vozbuditelem chumy v estestvennyh usloviyah. Stavropol’, RIO IDNK,
34 s. (in Russian)].
Дарская Н.Ф. 1970. К изучению годовых циклов блох рода Xenopsylla Roths., 1903. Переносчики особо
опасных инфекций и борьба с ними. Ставрополь, 108-131. [Darskaya N.F. 1970. K izucheniyu go-
dovich ziklov bloch roda Xenopsylla Roths., 1903. Peerenoschiki osobo opasnych infekziy i borba s nimi.
Stavropol, 108-131. (in Russian)].
Дарская Н.Ф. 1977. Особенности образа жизни блох песчанок. Экология и медицинское значение песчанок
фауны СССР. M., 226-230. [Darskaya N.F. 1977. Features of the of gerbil fleas life style. Ecology and
medical significance of gerbils of the fauna of the USSR. M., 226-230. (in Russian)].
Иофф И.Г., Микулин М.А., Скалон О.И. 1965. Определитель блох Средней Азии и Казахстана. М., Ме-
дицина, 371 с. [Ioff I.G., Mikulin M.A., Skalon O.I. 1965. Opredelitel’ bloh Srednej Azii I Kazahstana.
M., Medicina, 371 s. (in Russian)].
Загнибородова Е.Н. 1968. Многолетнее изучение экологии блох большой песчанки на юге Центральных
Каракумов. Грызуны и их эктопаразиты. Саратов, 78-86. [Zagniborodova E.N. 1968. Mnogoletnee
izuchenie ekologii bloh bol’shoj peschanki na yuge Central’nyh Karakumov. Gryzuny i ih ektoparazity.
Saratov, 78-86. (in Russian)].
Загнибородова Е.Н., Русакова Л.В., Бурлаченко Т.А., Старожицкая Г.С., Адаменко В.И., Сачеев М.Т. 1979.
Сравнительные данные по образованию чумного блока у блох большой и краснохвостой песчанок и
желтого суслика. Проблемы особо опасных инфекций 68 (4): 35-39. [Zagniborodova E.N., Rusakova
L.V., Burlachenko T.A., Starozhickaya G.S., Adamenko V.I., Sacheev M.T. 1979. Sravnitel’nye dannye
po obrazovaniyu chumnogo bloka u bloh bol’shoj i krasnohvostoj peschanok i zheltogo suslika. Problemy
osobo opasnyh infekcij 68 (4): 35-39. (in Russian)].
Золотова С.И., Хохлова С.А., Филипченко В.Е. 1975. Материалы к размножению блох Xenopsylla skrjabini
в Приаральских Каракумах. Проблемы особо опасных инфекций 43-44 (3-4): 97-102. [Zolotova S.I.,
Hohlova S.A., Filipchenko V.E. 1975. Materialy k razmnozheniyu bloh Xenopsylla skrjabini v Priaral’skih
Karakumah. Problemy osobo opasnyh infekcij 43-44 (3-4): 97-102. (in Russian)].
Иванов В.П. 1993. Электронномикроскопическое исследование пигидиального рецепторного органа у блохи
Xenopsylla cheopis Roths., 1903 (Siphonaptera). Энтомологическое обозрение 72 (3): 507-518. [Iva-
nov V.P. 1993. Electronnomicroscopicheskoe issledovanie pigidialnogo organa u blohi Xenopsylla cheopis
Roths., 1903 (Siphonaptera). Entomologicheskoe obozrenie 72 (3): 507-518. (in Russian)].
Иофф И.Г., Микулин М.А., Скалон О.И. 1965. Определитель блох Средней Азии и Казахстана. М., Ме-
дицина, 371 с. [Ioff I.G., Mikulin M.A., Skalon O.I. 1965. Opredelitel’ bloh Srednej Azii i Kazahstana.
M., Medicina, 371 s. (in Russian)].
Кадастр эпидемических и эпизоотических проявлений чумы на территории Российской Федерации и стран
Ближнего Зарубежья (с 1876 по 2016 год). 2016. Саратов, Амирит, 248 с. [Kadastr ehpidemicheskii
ehpizooticheskikh proyavlenij chumy na territorii Rossijskoj Federacii i stran Blizhnego Zarubezh’ya (s
1876 po 2016 god). 2016. Saratov, Amirit, 248 s. (In Russian)].
Каримова Т.Ю., Неронов В.М. 2007. Природные очаги чумы Палеарктики. М., Наука, 199 с. [Karimova
T.Yu., Neronov V.M. 2007. Prirodnye ochagi chumy Palearktiki. M., Nauka, 199 s. (in Russian)].
Климова З.И. 1972. Xenopsylla gerbilli caspica - компонент норовых микробиоценозов большой песчанки
в Северных Кызылкумах. Проблемы особо опасных инфекций 24 (2): 94-102. [Klimova Z.I. 1972.
Xenopsylla gerbilli caspica - komponent norovyh mikrobiocenozov bol’shoj peschanki v Severnyh Ky-
zylkumah. Problemy osobo opasnyh infekcij 24 (2): 94-102. (in Russian)].
413
Котти Б.К., Жильцова М.В. 2019. Значение блох (Siphonaptera) в природных очагах чумы. Паразитология
53 (6): 506-517. [Kotti B.K., Zhilzova M.V. 2019. A value of fleas (Siphonaptera) in the natural foci of
plague. Parasitologiya 53 (6): 504-514. (in Russian)].
Кучерук В.В., Дарская Н.Ф. 1981. Блохи песчанок: хозяева, распространение, родственные связи. Экология
и медицинское значение песчанок фауны СССР. М, 198-203. [Kucheruk V.V., Darskaya N.F. 1981.
Blochi peschanok: chozyaeva, rasprostraneniye, rodstvenniye svyazi. Ekologiya imedizinkoye znacheniye
peschanok fauny SSSR. M, 198-203. (in Russian)].
Медведев С.Г., Вержуцкий Д.Б. 2019. Разнообразие блох - переносчиков возбудителей чумы: паразит сус-
ликов - блоха Oropsylla silantiewi (Wagner, 1898) (Siphonaptera, Ceratophyllidae). Паразитология 53
(4): 267-282. [Medvedev S.G., Verzhutsky D.B. 2020. Diversity of Fleas, Vectors of Plague Pathogens:
the Flea Oropsylla silantiewi (Wagner, 1898) (Siphonaptera, Ceratophyllidae). Entomological Review 100
(1): 45-57.].
Медведев С.Г., Котти Б.К., Вержуцкий Д.Б. 2019. Разнообразие блох (Siphonaptera) - переносчиков возбуди-
телей чумы: паразит сусликов - блоха Citellophilus tesquorum (Wagner, 1898). Паразитология 53 (3):
179-197. [Medvedev S.G., Kotti B.K., Verzhutsky D.B. 2019. Diversity of Fleas (Siphonaptera), Vectors
of Plague Pathogens: the Flea Citellophilus tesquorum (Wagner, 1898), a Parasite of Ground Squirrels of
the Genus Spermophilus. Entomological Review 99 (5): 565-579.].
Медведев С.Г., Вержуцкий Д.Б., Котти Б.К. 2020. Разнообразие переносчиков возбудителя чумы: полиго-
стальные паразиты - блохи рода Rhadinopsylla Jordan et Rothschild, 1911 (Siphonaptera: Hystrichopsyl-
lidae). Паразитология 54 (3): 205-231. [Medvedev S.G., Verzhutsky D.B., Kotti B.K. 2020. Diversity of
Vectors of Plague Pathogens: Polyhostal Parasites, Fleas of the Genus Rhadinopsylla Jordan et Rothschild,
1911 (Siphonaptera, Hystrichopsyllidae). Entomological Review 100 (9): 1218-1235.].
Медведев С.Г., Вержуцкий Д.Б., Котти Б.К. 2021. Разнообразие переносчиков чумы: блохи рода Fronto-
psylla Wagner et Ioff, 1926 (Siphonaptera, Pulicidae). Паразитология 55 (6): 476-495. [Medvedev S.G.,
Verzhutsky D.B., Kotti B.K. 2021. Diversity of Plague Vectors: Fleas of the Genus Frontopsylla Wagner
et Ioff, 1926 (Siphonaptera, Pulicidae). Entomological Review 101 (9): 1-14.].
Медведев С.Г., Вержуцкий Д.Б., Котти Б.К. 2022. Блохи рода Paradoxopsyllus Miyajima et Koidzumi, 1909
(Siphonaptera, Leptopsyllidae) их роль в природных очагах чумы. Паразитология 56 (3): 226-251.
[Medvedev S.G., Verzhutsky D.B., Kotti B.K. 2022. Fleas of the Genus Paradoxopsyllus Miyajima et
Koidzumi, 1909 (Siphonaptera, Leptopsyllidae). Parazitologiya 56 (3): 226-251. (in Russian)].
Мокриевич Н.А., Рябцева Н.Л., Булах О.С., Талыбов А.Н. 1983. К оценке блокообразующей и заражающей
способности блох - основных переносчиков чумы в Закавказском равнинно-предгорном очаге. Про-
филактика природноочаговых инфекций. Ставрополь, 250-252. [Mokrievich N.A., Ryabceva N.L., Bulah
O.S., Talybov A.N. 1983. K ocenke blokoobrazuyushchej i zarazhayushchej sposobnosti bloh - osnovnyh
perenoschikov chumy v Zakavkazskom ravninno-predgornom ochage. Profilaktika prirodnoochagovyh
infekcij. Stavropol, 250-252. (in Russian)].
Новокрещенова Н.С., Кочетов А.Х., Кузнецова К.А., Старожицкая Г.С. 1968а. Влияние особенностей пита-
ния блох на их активность в передаче чумы (в эксперименте). Грызуны и их эктопаразиты. Саратов,
245-255. [Novokreshchenova N.S., Kochetov A.H., Kuznecova K.A., Starozhickaya G.S. 1968a. Vliyanie
osobennostej pitaniya bloh na ih aktivnost’ v peredache chumy (v eksperimente). Gryzuny i ih ektoparazity.
Saratov, 245-255. (in Russian)].
Новокрещенова Н.С., Солдаткин И.С., Левошина А.И. 1968б. Сравнительная частота питания различных
видов блох, определенная в лабораторных условиях с применением радиоактивных индикаторов.
Грызуны и их эктопаразиты. Саратов, 49-54. [Novokreshchenova N.S., Soldatkin I.S., Levoshina A.I.
1968b. Sravnitel’naya chastota pitaniya razlichnyh vidov bloh, opredelennaya v laboratornyh usloviyah s
primeneniem radioaktivnyh indikatorov. Gryzuny i ih ektoparazity. Saratov, 49-54. (in Russian)].
414
Природные очаги чумы Кавказа, Прикаспия, Средней Азии и Сибири. 2004. Под ред. Г.Г. Онищенко, В.В.
Кутырева. М., Медицина, 192 с. [Prirodnye ochagi chumy Kavkaza, Prikaspiya, Srednej Azii i Sibiri.
2004. Pod red. G.G. Onishchenko, V.V. Kutyreva. M., Medicina, 192 ss. (in Russian)].
Павлинов И.Я., Дубровский Ю.А., Россолимо О.Л., Потапова Е.Г. 1990. Песчанки мировой фауны. Москва,
Наука, 368 с. [Pavlinov I.Ya., Dubrovski U.A., Rossolimo O.L., Potapova E.G. 1990. Peschanki mirovoi
fauny. Moskva, Nauka, 368 pp. (in Russian)].
Ралль Ю.М. 1960. Грызуны и природные очаги чумы. М., Медгиз, 224 с. [Rall Yu.M. 1960. Gryzuny I
prirodnye ochagi chumy. M., Medgiz, 224 pp. (in Russian)].
Сажнев Ю.С., Рапопорт Л.П., Кулемин М.В., Сайлаубекулы Р. 2018. Материалы по ландшафтно-
эпизоотологическому районированию западной части Бетпакдалинского автономного очага чумы.
Карантинные и зоонозные инфекции в Казахстане. 1-2 (36-37): 31-38. [Sazhnev Yu.S., Rapoport
L.P., Kulemin M.V., Sajlaubekuly R. 2018. Materialy po landshaftno-epizootologicheskomu rajonirovaniyu
zapadnoj chasti Betpakdalinskogo avtonomnogo ochaga chumy. Karantinnye i zoonoznye infekcii v Ka-
zahstane 1-2 (36-37): 31-38. (in Russian)].
Сараев Ф.А., Хамзин Т.Х., Козулина И.Г., Башмакова А.А., Тегисбаева А.У., Башмаков А.А. 2019. Особен-
ности эпизоотии чумы 1997-1999 годов в южной части Волго-Уральских песков. Карантинные и
зоонозные инфекции в Казахстане 1 (38): 109-117. [Saraev F.A., Hamzin, T.H., Kozulina I.G., Bash-
makova A.A., Tegisbaeva A.U., Bashmakov A.A. 2019. Osobennosti epizootii chumy 1997-1999 godov v
yuzhnoj chasti Volgo-Ural’skih peskov. Karantinnye i zoonoznye infekcii v Kazahstane 1 (38): 109-117.
(in Russian)].
Сержанов О.С., Хрусцелевская Н.М., Чумаченко В.Д., Асенов Г.А., Матаков М.И. 1979. Блокообразование у
блох Xenopsylla gerbilli caspica из различных ландшафтно-экологических участков Кызылкумов. Про-
блемы особо опасных инфекций 68 (4): 58-60. [Serzhanov O.S., Hruscelevskaya N.M., CHumachenko
V.D., Asenov G.A., Matakov M.I. 1979. Blokoobrazovanie u bloh Xenopsylla gerbilli caspica iz razlichnyh
landshaftno-ekologicheskih uchastkov Kyzylkumov. Problemy osobo opasnyh infekcij 68 (4): 58-60. (in
Russian)].
Слудский А.А. 2014. Эпизоотология чумы (обзор исследований и гипотез). (Деп в ВИНИТИ 11.08.2014.
№ 231-В 2014 Саратов, 313 с. [Sludskyi AA. 2014. Epizootology of plague (review of studies and hy-
potheses). Deposited in Russian Institute for Scientific and Technical Information of Russian Academy of
Sciences, 08.11.2014, № 231, 2014), Saratov, 2014. 313 pp.].
Солдаткин И.С., Левошина А.И. 1968. К вопросу о продолжительности непрерывного пребывания блох в
шерсти больших песчанок. Грызуны и их эктопаразиты. Саратов, 55-58. [Soldatkin I.S., Levoshina
A.I. 1968. K voprosu o prodolzhitel’nosti nepreryvnogo prebyvaniya bloh v shersti bol’shih peschanok.
Gryzuny i ih ektoparazity. Saratov, 55-58. (in Russian)].
Старожицкая Г.С. 1970. Экологические особенности блох большой песчанки на участке стойкой очаговости
чумы. Проблемы особо опасных инфекций 16 (6): 157-163. [Starozhickaya G.S. 1970. Ekologicheskie
osobennosti bloh bol’shoj peschanki na uchastke stojkoj ochagovosti chumy. Problemy osobo opasnyh
infekcij 16 (6): 157-163. (in Russian)].
Тифлов В.Е., Скалон О.И., Ростигаев Б.А. 1977. Определитель блох Кавказа. Ставрополь, Ставроп. книжн.
изд-во, 280 с. [Tiflov V.E., Skalon O.I., Rostigaev B.A. 1977. Opredelitel' bloh Kavkaza. Stavropol',
Stavrop. knizhn. izd-vo, 280 s. (in Russian)].
Фаранг-Азад А. 1972. Материалы по фауне блох Ирана. Паразитология 6 (6): 513-521. [Farhang-Azad A.
1972. Materials on the fauna of fleas of Iran. Parazitologiya 6 (6): 513-521. (in Russian)].
415
Ширанович П.И., Рабинович Б.К., Эйгелис Ю.К., Ахундов М.А., Тимофеева М.Е., Васильченко А.П., Ко-
стенко И.С. 1977. О многолетних изменениях численности краснохвостой песчанки и ее блох в цен-
тральной части Прикуринской низменности. Проблемы особо опасных инфекций 54 (2): 28-32. [Shi-
ranovich P.I., Rabinovich B.K., Ejgelis Yu.K., Ahundov M.A., Timofeeva M.E., Vasil’chenko A.P., Kostenko
I.S. 1977. O mnogoletnih izmeneniyah chislennosti krasnohvostoj peschanki i ee bloh v central’noj chasti
Prikurinskoj nizmennosti. Problemy osobo opasnyh infekcij 54 (2): 28-32. (in Russian)].
Beaucournu J.C. 1977. Les Puces (Siphonaptera) du Maroc. Bulletins de l'Institut Scientifique, Rabat 2: 85-86.
Beaucournu J.C., Hellal H. 1977. Liste annotee des Siphonateres de Tunisie. Bulletin de la Société de pathologie
exotique 70: 524-537.
Beaucournu J.C., Kowalski K. 1985. Donnees nouvelles sur les puces (Insecta, Siphonaptea dAlgerie. Bulletin
de la Société de pathologie exotique 78: 378-392.
Beaucournu J.C., Launay H. 1990. Les Puces (Siphonaptera) de France et du Bassin Mediterraneen occidenta.
548 pp.
Cooreman J. 1973. Siphonaptera recuellis en Libye par la mission X. Misonne (1972-1973). Bulletin de l’Institut
royal des sciences naturelles de Belgique, Entomologie 49 (5): 1-11.
Hastriter M.W., Tipton V.J. 1975. Fleas (Siphonaptera) associated with small mammals of Morocco. Journal of
the Egyptian Public Health Association 50 (2): 79-169.
Hoogstraal H., Traub R. 1965. The fleas of Egypt. Host-parasite relationhips of rodens of Cricetid rodents (Family
Cricetidae, subfamily Gerbillinae). The Journal of the Egyptian public health association 40 (3): 141-145.
Hopkins G.H.E., Rothschild M. 1953. An illustrated catalogue of the Rothschild collection of fleas (Siphonaptera)
in the British Museum. Vol. 1. London, University Press, Cambridge, XV+361 p.
Glinkiewicz A. 1907. Ergebnisse der mit Subvention aus der Erbschaft Treitl untemommenen zoologischen Forsc-
hungsreise Dr. Franz Werner’s nach dem aegyptischen Sudan und Nord-Uganda. X. Parasiten von Pachy-
uromys duprasi Lat. Sber. Akad. Wiss. Wien 116, Abt. I: 381-386.
Klein J., Mofidi C., Chamas M., Karimi Y., Bahmanyar M., Seydian B. 1963. Les puces (Insecta, Siphonaptera)
de l’Iran. Bulletin de la Société de pathologie exotique 56: 533-550.
Klein J.-M., Alonso J. M., Baranton G., Poulet A.R., Mollaret H.H. 1975a. La Peste en Mauritanie. Re. Médecine
et Maladies Infectieuses 5 (4): 198-207.
Klein J.-M., Simonkovich E., Alonso J. M., Baranton G. 1975b. Observations écologiques dans une zone enzoo-
tique de peste en Mauritanie: 2. Les puces de rongeurs (Insecta, Siphonaptera). Cahiers ORSTOM. Série
Entomologie Médicale et Parasitologie 13 (1): 29-39.
Lewis R.E. 1967. The fleas (Siphonaptera) of Egypt. An illustrated and annotated key. The Journal of Parasitol-
ogy 53 (4): 863-885.
Lewis R.E. 1972. Notes on the geographical distribution and host preferences in the order Siphonaptera. Part 1.
Pulicidae. Journal of Medical Entomology 9 (6): 511-520.
Lewis R.E., Lewis J.H. 1990. Catalogue of invalid genus-group and species-group names in Siphonaptera (Insecta).
Koenigstein, Koeltz Scientific Books, 264 pp.
Misonne X. 1977. Un foyer naturel de peste in Libye. Annales Societe Belge Medicne Tropicale 57 (3): 163-168.
Rothschild N.C. 1903. New species of Siphonaptera from Egypt and the Soudan. Entomologist’s monthly maga-
zine 39 (467): 83-87.
Rothschild M., Schlein Y. 1975. The jumping mechanism of Xenopsylla cheopis. 1. Exoskelet structures and
musculature. Philosophical Transactions of the Royal Society. London. Ser. B. 271 (914): 457-490.
Lewis R.E. 1961. The thoracic musculature of the Indian rat flea, Xenopsylla cheopis (Siphonaptera). Annals of
the Entomological Society of America 54: 387-397.
The Atlas of Plague and Its Environment in the People`s Republic of China. 2000. Beijing, Science Press, 221 pp.
Wagner J., Wassilief A. 1933. Tables analytiques pour la determination des puces rencontrees en Algerie et Tunisie.
Archives de l’Instiut Pasteur de Tunis 21 (3): 431-467.
416
PALEARCTIC FLEA SPECIES
OF THE GENUS XENOPSYLLA (SIPHONAPTERA: PULICIDAE),
PARASITIZING ON GERBILS (RHOMBOMYS, MERIONES),
AND THEIR ROLE IN NATURAL PLAGUE FOCI
S. G. Medvedev, D. B. Verzhutsky, B. K. Kotti
Keywords: fleas, Siphonaptera, species vectors of plague pathogen, taxonomic diversity,
Xenopsylla
SUMMARY
Taxonomic diversity, characters of distribution and host-parasite relations of fleas of the genus
Xenopsylla (Pulicidae) are analyzed. It is demonstrated that six species and two subspecies of fleas
of the genus Xenopsylla, parasitizing on gerbils in the territory of Russia and adjacent countries are
known as the main, secondary, or occasional vectors of the plague.
417