ПАРАЗИТОЛОГИЯ, 2023, том 57, № 4, с. 337-343.
УДК 576.895.133:595.133
Неполная Фрагментация
гигантских ядер тегумента у акантелл скребня
Acanthocephalus tenuirostris
(Palaeacanthocephala, Echinorhynchida)
© 2023 г. Т. В. Давыденкоa, , В. П. Никишинa
a Институт биологических проблем Севера ДВО РАН,
ул. Портовая, 18, Магадан, 685000 Россия
e-mail: sesha17@mail.ru
Поступила в редакцию 27.06.2023 г.
После доработки 13.07.2023 г.
Принята к публикации 17.07.2023 г.
Гигантские ядра тегумента скребня Acanthocephalus tenuirostris на стадии поздней акантел-
лы подвергаются неполной фрагментации. Этому процессу предшествует трансформация ядер
и приобретение ими формы дерева. Между фрагментами ядер тегумента как у цистакантов,
так и у взрослых скребней сохраняются тонкие связи. Предполагается, что такая организация
ядерного аппарата обеспечивает жизнедеятельность тегумента, объем которого многократно
увеличивается в период последующего роста скребня.
Ключевые слова: скребни, фрагментация ядер, тегумент, акантелла, цистакант,
Acanthocephalus tenuirostris
DOI: 10.31857/S003118472304004X, EDN: KIAQYZ
Многочисленные ядра тегумента взрослых скребней некоторых семейств из класса
Palaeacanthocephala, по распространенному мнению, являются ядерными фрагментами,
производными предшествующих гигантских ядер (Петроченко, 1956; Хохлова, 1986;
Van Cleave, 1928; Miller, Dunagan, 1985; и др.). По данным разных авторов, фрагмен-
тация гигантских ядер тегумента происходит на стадиях средней и/или поздней акан-
телл. Например, Nicholas (1967) указывает, что на стадии поздней акантеллы (виды
не указаны) кортекс трансформируется в тегумент, сходный с тегументом взрослой
особи. Van Cleave (1928) отмечает, что у представителей классов Palaeacanthocephala
337
и Archiacantocephala на этой же стадии кортикальные ядра увеличиваются в размере
и числе, а также становятся древовидными или делятся амитозом. Schmidt (1985),
суммируя известные к тому времени данные и не уточняя таксономического положе-
ния скребней, заключает, что на стадии акантеллы ядра приобретают форму амебы
с длинными отростками, которые распадаются на фрагменты. Эти фрагменты затем
перемещаются в свои надлежащие места. У Echinorhynchus lageniformis ядра стано-
вятся дендритными к 22-му дню развития в промежуточном хозяине, но распадаются
к 25-му дню, в то время как полностью сформированный цистакант наблюдается
к 30-му дню (Olson, Pratt, 1971). К сходному выводу пришла Butterworth (1969) при
изучении Polymorphus minutus с более протяженным жизненным циклом: на стадии
средней акантеллы (84-100 дней развития) у гигантских кортикальных ядер появля-
ются древовидные отростки, а на стадии поздней акантеллы (100-150 дней развития)
ядра фрагментируются и их отростки образуют множество мелких ядер. При этом
автор отмечает, что между ядрами сохраняются тонкие связи, т.е. фрагментация не
является полной. При изучении Acantocrephalus parksidei из изопод, инвазированных
естественным образом, Amin (1982) пришел к выводу, что фрагментация ядер может
начаться еще до дифференцировки хоботкового аппарата и репродуктивной системы.
Таким образом, сведения об одном из значительных преобразований, маркирую-
щих процесс органогенеза скребней, о фрагментации гигантских ядер тегумента не-
однозначны. Неясен и характер тонких связей, сохраняющихся между фрагментами
ядер. Для подтверждения имеющихся данных о характере и полноте фрагментации
гигантских ядер тегумента мы проследили этот процесс в условиях эксперимента на
завершающих этапах развития скребня Acanthocephalus tenuirostris в промежуточном
хозяине.
Материал и методика
Материалом для исследований являлись акантеллы и цистаканты Acanthocephalus tenuirostris,
полученные экспериментальным путем, а также взрослые скребни этого вида. Взрослые
скребни для исследования были извлечены из хариусов Thymallus arcticus pallasii, пойманных
в р. Эльген (правый приток р. Буюнда, бассейн р. Колыма). Для экспериментов использовались
водяные ослики Asellus hilgendorphi, отловленные в р. Aмутты (приток р. Эльген, бассейн
р. Буюнда) в качестве промежуточного хозяина, и зрелые яйца самки скребня, извлеченной
из ленка Brachymystax lenok, пойманного в р. Буюнда.
Эксперимент проводился при температуре 14-15ᴼС. Водяных осликов содержали в контей-
нерах с водопроводной водой с добавлением мха рода Calliergon и кормили сухими листья-
ми листопадных деревьев. Исследуемые акантеллы были получены через 30 дней (средние
акантеллы) и 39 дней (поздние акантеллы) со дня начала эксперимента, а цистаканты - через
60 и 65 дней.
338
Скребней фиксировали в 2% глутаровом альдегиде на фосфатном буфере (pH 7.4). После
альдегидной фиксации образцы постфиксировали в 1% тетраоксиде осмия на таком же буфере,
обезвоживали и заключали в смесь эпона и аралдита. В процессе обезвоживания образцы
проводили через 1% раствор уранилацетата на 70% спирте в течение ночи. Полутонкие срезы
окрашивали смесью 1% метиленового синего и 1% кристаллического фиолетового в разных со-
отношениях и исследовали с помощью светового микроскопа Olympus CX41 с использованием
фотокамеры Olympus E-420. Ультратонкие срезы дополнительно контрастировали 1% раство-
ром уранилацетата на 70% спирте в течение двух часов. Исследования проводили с помощью
трансмиссионного электронного микроскопа JEM1400PLUS с использованием фотокамеры SIS
Veleta. Полученные фотографии обрабатывали с помощью комплекта программ CorelDRAW 20.
Результаты и обсуждение
На основании классификации Butterworth (1969) мы дифференцировали исследуе-
мых акантелл на средних (без дополнительного разделения) и поздних.
На поверхности средних акантелл располагается рыхлый слой мелкогранулярно-
го и аморфного материала, толщиной 0.6 мкм, представляющий собой формирую-
щуюся цисту. Тегумент средних акантелл (или кортекс по терминологии Nicholas,
1967) представляет собой пласт цитоплазмы толщиной от 7 до 30 мкм на разных
стадиях развития. Этот пласт не дифференцирован на слои, характерные для взрос-
лых скребней (поперечно-полосатый, войлочно-волокнистый, радиально-волокнистый
и трубочковый). Ярко выражен только везикулярный слой, образованный расширен-
ными в форме ампул инвагинатами наружной плазматической мембраны (криптами
по терминологии Miller, Dunagan, 1985). В этих расширениях заметны скопления рых-
лого материала, впоследствии выделяющегося на поверхность тегумента и входящего
в состав цисты. Похожий материал обнаруживается в электронно-светлых включени-
ях, которые в большом количестве находятся в цитоплазме под везикулярным слоем.
Гигантские ядра (по разным терминологиям кортикальные, подкожные, гиподер-
мальные) располагаются в толще тегумента. В тегументе средних акантелл были от-
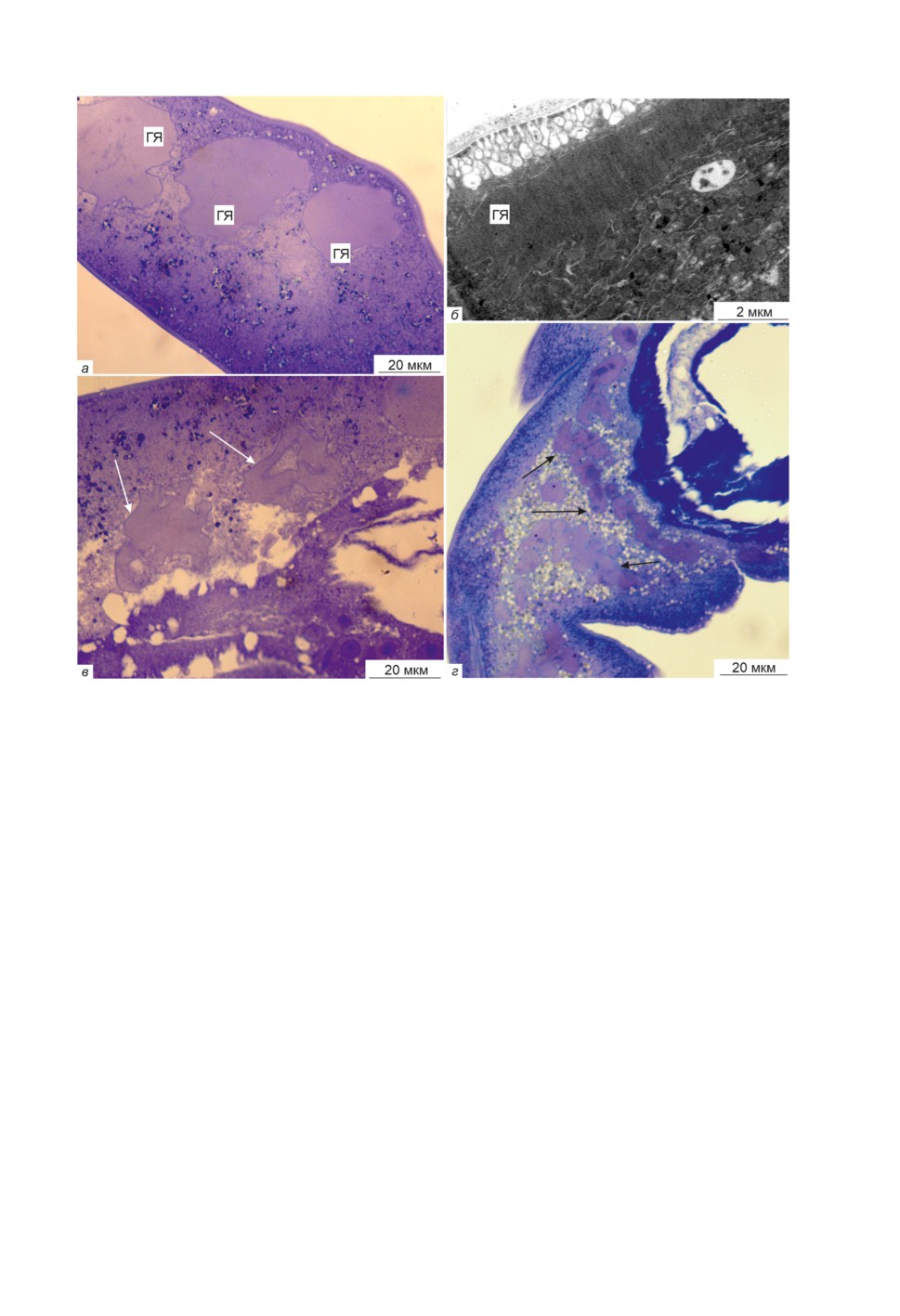
мечены преимущественно гигантские ядра вытянутой формы, размерами до 35 мкм
по большей оси (рис. 1а). Ядрышек в гигантских ядрах не обнаружено, кариоплазма
гомогенная. Помимо этого, были найдены ядра, которые отличались от вышеописан-
ных неправильной формой и имели многочисленные лопастные отростки. Наличие
таких ядер, по нашему мнению, свидетельствует о начавшемся процессе фрагмента-
ции (рис. 1б, 1в).
В тегументе поздних акантелл угадываются формирующиеся слои тегумента бу-
дущего цистаканта, при этом войлочно-волокнистый слой окрашивается интенсив-
нее радиально-волокнистого. На рис. 1г видны отдельные фрагменты ядер, которые
уже отделились от вытянутого «родительского» древовидного ядра и имеют длину
до 60 мкм.
339
Рисунок 1. Ядра тегумента акантеллы Acanthocephalus tenuirostris на разных этапах
фрагментации: а-в - средняя акантелла, г - поздняя акантелла. ГЯ - гигантские ядра
тегумента. Белыми стрелками показаны древовидные ядра тегумента, черными -
фрагментирующиеся. а, в, г - световая микроскопия, б - электронная микроскопия.
Figure 1. Nuclei of the tegument of the acanthella Acanthocephalus tenuirostris at different stages
of fragmentation.
Тегумент цистаканта практически не отличается от такового взрослого скребня
за исключением наличия везикулярного слоя (у взрослых скребней образующие его
элементы не расширены) и размерных характеристик (Давыденко, Никишин, 2023).
Волокна войлочно-волокнистого слоя, а также лакуны и множество липидных капель
в радиально-волокнистом слое хорошо просматриваются даже в световой микроскоп.
Ядерные фрагменты различной формы обычно находятся друг от друга на некотором
расстоянии, но встречаются и близко расположенные. В некоторых из них обнаружи-
ваются от 1 до 4 ядрышек (рис. 2а, 2б). В ряде случаев во фрагментах ядер наблю-
даются удлинение ядрышек и образование своеобразной центральной «перетяжки»
между двумя их частями (рис. 2а), что может свидетельствовать о начинающемся
процессе деления или фрагментации ядрышек.
340
Рисунок 2. Фрагментированные ядра тегумента скребня Acanthocephalus tenuirostris
(световая микроскопия): а, б - цистакант; в, г - взрослый скребень. Белой стрелкой показано
ядрышко, располагающееся в двух ядерных фрагментах. Черными стрелками показаны связи
между фрагментами ядер тегумента.
Figure 2. Fragmented tegument nuclei of the Acanthocephalus tenuirostris (light microscopy).
Тегумент взрослых скребней по сравнению с таковым цистакантов претерпевает
некоторые преобразования: увеличивается его толщина, элементы везикулярного слоя
сужаются и приобретают вид, обычный для инвагинатов наружной цитоплазмати-
ческой мембраны тегумента, уменьшается количество липидных капель, волокна и
лакуны становятся более выраженными. Фрагменты ядер тегумента по своей морфо-
логии и размерам не отличаются от таковых тегумента цистаканта, и многие из них
связаны друг с другом тонкими отростками.
Таким образом, фрагментация ядер у исследованных скребней начинается с про-
цесса приобретения ими формы дерева на стадии средней акантеллы, продолжается
в виде собственно фрагментации на стадиях средней и поздней акантеллы и завер-
341
шается на стадии цистаканта. При этом фрагменты ядер, по крайней мере, некоторые
из них, остаются связанными друг с другом тонкими отростками.
Полученные результаты в совокупности с данными литературы ставят под сомне-
ние правомерность использования по отношению к ядрам тегумента скребней термина
«фрагментация», поскольку, как минимум, между некоторыми из них сохраняются
связи. По этой же причине невозможно рассматривать процесс увеличения количества
ядерных элементов как форму амитоза. Тем не менее, учитывая отсутствие данных
о точном распределении генетического материала между дочерними фрагментами
ядер, а также однозначных доказательств наличия амитоза, и принимая во внимание
наличие связей, по крайней мере, между некоторыми ядерными фрагментами, мы
полагаем возможным сохранение термина «фрагментация» с дополнением «неполная».
Как уже отмечалось выше, тонкие связи между ядерными фрагментами ра-
нее наблюдались у поздних акантелл Polymorphus minutus (Butterworth, 1969). На-
шими недавними исследованиями показано, что многие фрагменты ядер у взрос-
лых Macracanthorhynchus catulinus (Archiacantocephala) и ювенильных Corynosoma
strumosum (Palaeacanthocephala) также остаются связанными между собой тонкими
отростками (Никишин, Пономарев, 2023). Было сделано предположение, что посред-
ством этих связей фрагменты ядер объединяются в своеобразные сети, количество
которых соответствует количеству изначальных гигантских ядер (Никишин, Понома-
рев, 2023). Такая организация ядерного аппарата, вероятно, позволяет обеспечивать
жизнедеятельность тегумента, объем которого многократно увеличивается в период
роста скребня в организме окончательного хозяина. Полученные здесь результаты не
противоречат этому предположению. Анализ имеющихся данных позволяет заклю-
чить, что сохранение связей между ядерными фрагментами в тегументе скребней не
зависит от размера паразитов. Возможно, эта особенность характерна для тегумента
многих, если не всех, представителей класса Palaeacanthocephala, а также некоторых
Archiacanthocephala.
ФИНАНСИРОВАНИЕ РАБОТЫ
Исследования проведены в ходе выполнения государственного задания по теме:
«Гельминты в биоценозах северо-восточной Азии: биоразнообразие, морфология и
молекулярная филогенетика» (№ регистрации: 1021060307693-0).
Список литературы
Давыденко Т.В., Никишин В.П. 2023. Особенности организации тегумента у цистаканта и взрослого скребня
Acanthocephalus tenuirostris. Биология внутренних вод (2): 164-172. DOI 10.31857/S0320965223020043.
[Davydenko T.V., Nikishin V.P. 2023. Features of the organization of the tegument in cystacanth and adult
acanthocephalan Acanthocephalus tenuirostris (Achmerov et Dombrowskaja - Achmerova, 1941). Inland
Water Biology 16 (2) 185-192. DOI 10.1134/S1005082923020049].
342
Никишин B.П., Пономарев Д.В. 2023. Об организации ядер в тегументе некоторых скребней классов
Palaeacanthocephala и Archiacanthocephala. Паразитология 57 (1): 3-9. [Nikishin V.P., Ponomarev
D.V. 2023. Ob organizatsii yader v tegumente nekotoryh skrebney klassov Palaeacanthocephala i
Archiacanthocephala. Parazitologiya 57 (1): 3-9. (In Russian)].
Петроченко В.И. 1956. Акантоцефалы (скребни) домашних и диких животных. Т. I. М., Изд-во АН СССР,
435 с. [Petrochenko V.I. 1956. V. I. Acanthocephala of Domestic and Wild Animals. Moscow, Izdatel’stvo
Akademii Nauk SSSR, 435 pp. (In Russian)].
Хохлова И.Г. 1986. Акантоцефалы наземных позвоночных фауны СССР. М., Наука, 278 с. [Khokhlova
I.G. 1986. Acanthocephalans of Terrestrial Vertebrates of the USSR Fauna. Moscow, Nauka, 278 pp. (In
Russian)].
Amin O.M. 1982. Description of larval Acanthocephalus parksidei Amin, 1975 (Acanthocephala: Echinorhynchidae)
from its isopod intermediate host. Proceedings of the Helminthological Society of Washington 49 (2):
235-245.
Butterworth P. 1969. The development of the body wall of Polymorphus minutus (Acanthocephala) in its
intermediate host Gammarus pulex. Parasitology 59 (2): 373-388.
Miller D.M., Dunagan T.T. 1985. Functional morphology. In: Crompton D.W.T., Nickol B.B. (eds). Biology of
the Acanthocephala. Cambridge, Cambridge University Press, P. 73-124.
Nicholas W.L. 1967. The biology of the Acanthocephala. Advances in Parasitology 5: 205-246.
Olson R.E., Pratt I. 1971. The life cycle and larval development of Echinorhynchus lageniformis Ekbaum, 1938
(Acanthocephala: Echinorhynchidae). Journal of Parasitology 57 (1): 143-149.
Schmidt G.D. 1985. Development and life cycles. In: Crompton D.W.T., Nickol B.B. (eds). Biology of the
Acanthocephala. Cambridge, Cambridge University Press, P. 237-286.
Van Cleave H.J. 1928. Nuclei of the subcuticula in the Acanthocephala. Zeitschrift für Zellforschung und
Mikroscopische Anatomie 7 (1): 109-113.
INCOMPLETE Fragmentation of giant tegument nuclei in
Acanthocephalus tenuirostris (Palaeacanthocephala,
Echinorhynchida)
T. V. Davydenko, V. P. Nikishin
Keywords: acanthocephalans, tegument, fragmentation of nuclei, acanthella, cystacanth,
Acanthocephalus tenuirostris
Summary
The giant nuclei of the Acanthocephalus tenuirostris tegument at the late acanthella stage show
incomplete fragmentation. This process is preceded by the transformation of the nuclei, and their
acquisition of a tree-like shape. Thin connections between the nuclear fragments of the tegument is
retained both at the stage of cystacanth and in the adult acanthocephalans. It is assumed that such an
organization of the nuclear structure ensures the vital activity of the tegument, the volume of which
increases repeatedly during the subsequent growth of the acanthocephalan.
343