ПАРАЗИТОЛОГИЯ, 2023, том 57, № 6, с. 504-520.
УДК 595.122.1
РАСПРЕДЕЛЕНИЕ МОНОГЕНЕЙ РОДА LIGOPHORUS
НА ЖАБРАХ ПИЛЕНГАСА PLANILIZA HAEMATOCHEILUS
© 2023 г. Н. В. Пронькинаa,*, Е. В. Дмитриеваa
aФИЦ Институт биологии южных морей имени А.О. Ковалевского РАН,
пл. Нахимова, 2, Севастополь, 299011 Россия
*e-mail: natalya-pronkina@yandex.ru
Поступила в редакцию 14.09.2023 г.
После доработки 29.10.2023 г.
Принята к печати 01.11.2023 г.
Изучены особенности локализации на жабрах хозяев трех видов моногеней рода Ligophorus
Еuzet et Suriano, 1977, паразитирующих у пиленгаса Planiliza haematocheilus (Temminck &
Schlegel, 1845) в Черном и Азовском морях. Особи L. pilengas и L. llewellyni предпочитали
локализоваться на одних и тех же участках жабр, а именно, на I-III жаберных дугах и их
медиальных секторах. Третий вид - L. kaohsianghsieni - распределялся на жабрах более ску-
ченно, около 80% его особей локализовалось на II жаберной дуге, встречаясь в медиальном
и вентральном секторах в равной степени. В целом, большинство лигофорусов найдено на
бóльших по площади и хорошо омываемых водой II-III дугах, а также в медиальном и вен-
тральном секторах передней полужабры, но здесь моногенеи предпочитали прикрепляться
к проксимальным участкам филаментов с меньшим током воды. Присутствие других видов рода
на жабрах не влияло на распределение L. pilengas. Рост численности инфрапопуляций приводил
к более равномерному распределению моногеней на участках жабр, а при увеличении размера
рыб значимо возрастала их доля на быстрее растущей передней полужабре.
Ключевые слова: моногенеи, Ligophorus, виды одного рода, распределение по жабрам,
численность инфрапопуляции, размер рыб
DOI: 10.31857/S0031184723060054 ; EDN: RXVQTK
Неравномерное и неслучайное распределение моногеней на жабрах их хозяев
было отмечено еще в XIX веке (Cerfontaine, 1896). Позднее В.А. Догель (1949) писал
о большом теоретическом интересе, который представляет явление одновременного
паразитирования видов одного рода в одном и том же хозяине.
Представители разных видов Ligophorus Euzet et Suriano, 1977 (Monopisthocotylea:
Ancyrocephalidae) часто встречаются на одних и тех же рыбах совместно, составляя
сообщества от 2 до 6 видов на одном хозяине (Dmitrieva et al., 2012). Однако только
для трех пар видов - L. parvicirrus Euzet et Suriano, 1977 и L. imitans Euzet et Suriano,
1977, паразитирующих на Liza ramada в Средиземном море (Euzet, Sanfilippo, 1983;
Abu Samak, Hassan, 1998), а также L. vanbenedeni Euzet et Suriano, 1977 и L. szidati
Euzet et Suriano, 1977, и L. acuminatus Euzet et Suriano, 1977 и L. szidati, встречающих-
ся на Chelon auratus (Risso, 1810) и C. saliens (Risso, 1810), соответственно, в Черном
504
море (Пронькина и др., 2010), - были проведены исследования их распределения по
жабрам хозяев. Авторы еще одной публикации, посвященной распределению видов
этого рода на жабрах Liza macrolepis (Kumar et al., 2016), не определили моногеней
до вида, анализируя их как один сборный вид - «Ancyrocephalus vanbenedenii».
Данная работа посвящена изучению особенностей локализации на жабрах хозяев
трех видов моногеней рода Ligophorus Еuzet et Suriano, 1977, паразитирующих на
пиленгасе Planiliza haematocheilus (Temminck & Schlegel, 1845) в Черном и Азовском
морях, и влиянию на их распределение присутствия моногеней других видов, числен-
ности их инфрапопуляций, а также размера рыб.
МАТЕРИАЛ И МЕТОДИКА
Материал собран в 2001-2016 гг. от 51 экз. Planiliza haematocheilus, общей длиной от 13
до 66 см, из четырех районов крымского прибрежья Черного и Азовского морей (прибрежные
акватории г. Севастополь, п. Набережное, п. Юркин, п. Щелкино).
Моногеней собирали живыми и фиксировали в глицерин-желатине (Гусев, 1983), идентифи-
цировали под микроскопом Olympus CX41 с фазово-контрастным устройством при увеличении
×800. Найдены три вида моногеней: Ligophorus pilengas Sarabeev et Balbuena, 2004 (2250 экз.),
L. llewellyni Dmitrieva, Gerasev et Pronkina, 2007 (1387 экз.), L. kaohsianghsieni (Gusev, 1962)
Gusev, 1985 (50 экз.).
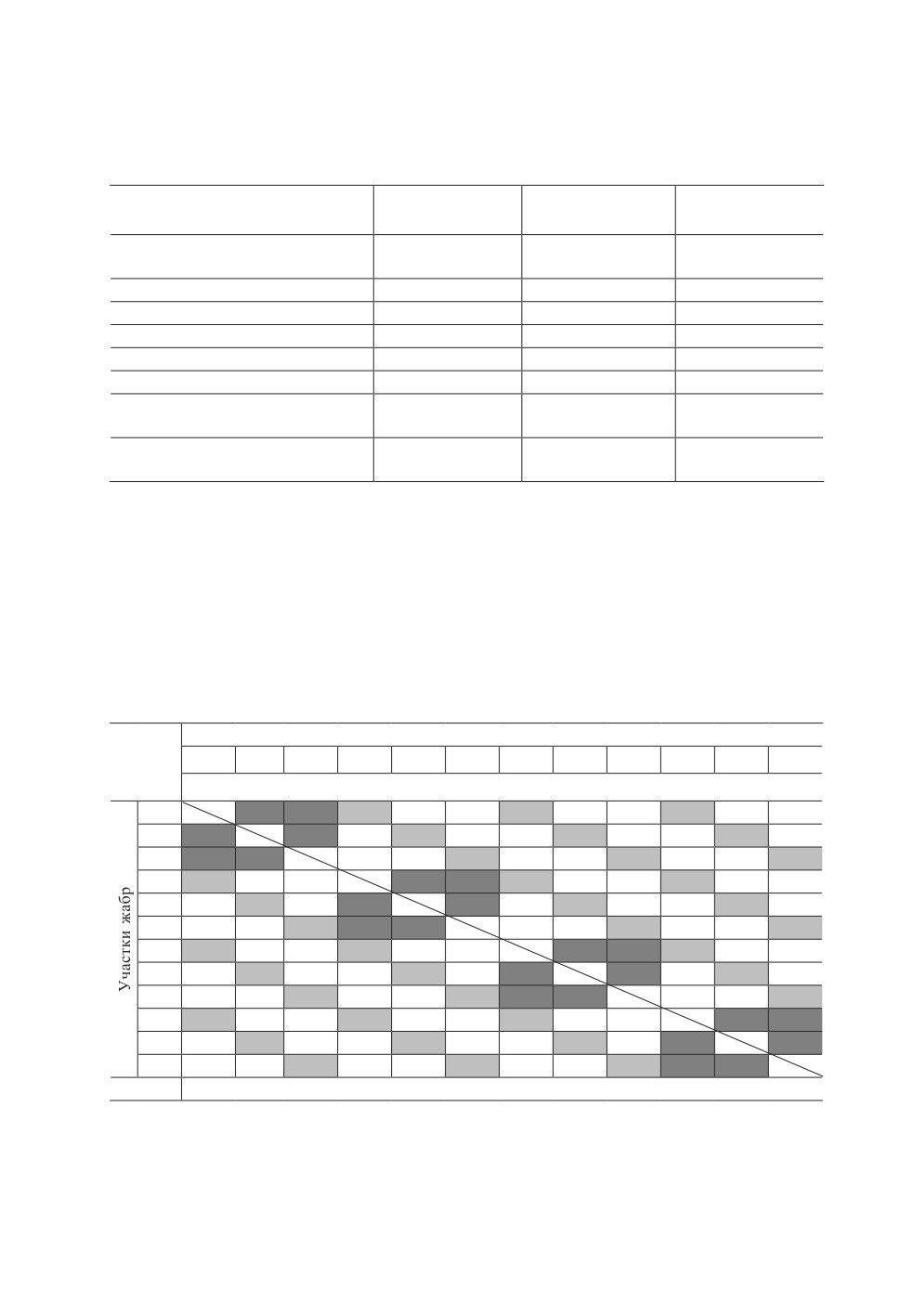
Жабры делили (рис. 1) на правую и левую половины, состоящие из четырех жаберных дуг
(I-IV), каждую дугу разделяли на переднюю и заднюю полужабры , на которых выделяли по
три сектора (1 - дорсальный, 2 - медиальный и 3 - вентральный) и две зоны вдоль жаберных
лепестков (дистальная и проксимальная половины).
Рисунок 1. Схема деления жабр Planiliza haematocheilus на участки. Обозначения: I-IV -
четыре жаберные дуги, П - передняя полужабра, З - задняя полужабра, 1 - дорсальный
сектор, 2 - медиальный сектор, 3 - вентральный сектор, Д - дистальная часть жаберных
лепестков, Пр - проксимальная часть жаберных лепестков.
Figure 1. Schem of the division of Planiliza haematocheilus gills into sections. Keys: I-IV - four
gill arches, П - anterior hemibranchies, З - posterior hemibranchies, 1 - dorsal sector, 2 - medial
sector, 3 - ventral sector, Д - distal part of gill filaments, Пр - proximal part of gill filaments.
505
Для характеристики распределения моногеней в пределах жаберного биотопа рассчитана
доля червей, локализующихся на выделенных участках жабр, от общего количества червей на
рыбе. Выборки долей характеризовались их средними значениями и стандартными ошибками.
Численность моногеней характеризовалась интенсивностью инвазии - ИИ, экз./особь,
и индексом обилия - ИО, экз./особь, встречаемость - экстенсивностью инвазии, ЭИ (%). Для
анализа влияния численности инфрапопуляций на распределение L. pilengas и L. llewellyni
были выделены 4 класса по ИИ, для первого вида: 1 - ≤ 20 экз./особь (20 рыб, 216 экз. моно-
геней), 2 - 21-60 экз./особь (11 рыб, 365 экз.), 3 - 61-100 экз./особь (10 рыб, 701 экз.) и 4 -
101-200 экз./особь (8 рыб, 968 экз.); для второго вида: 1 - ≤ 10 экз./особь (14 рыб, 69 экз.), 2 -
11-40 экз./особь (12 рыб, 316 экз.), 3 - 41-70 экз./особь (7 рыб, 404 экз.) и 4 - 71-110 экз./особь
(7 рыб, 598 экз.). Этот анализ проведен только для двух видов, т.к. Ligophorus kaohsianghsieni
был, в целом, немногочислен, и его ИИ колебался незначительно. Для анализа влияния размера
рыб на распределение моногеней выделены 3 класса: 1 - особи размерами (TL) от 13 до 30 см
(14 рыб), 2 - 31-40 см (15 рыб) и 3 - 41-66 см (20 рыб).
Для анализа влияния места локализации (половина каждой жабры, дуга, сектор, полужабра
или часть лепестка) на долю моногеней, встречающихся на них, использовались дисперсионный
анализ (ANOVA) и пост-хок тест. Оценка значимости различий между средними двух выборок
проводилась по критерию Фишера (F-критерий), достоверными считались различия при р <0.05,
при этом значения F и р выделены жирным шрифтом. Все вычисления и их графическое ото-
бражение выполнены в программе Statistica 10.
РЕЗУЛЬТАТЫ
Ligophorus pilengas встречен у 49 экз. рыб с интенсивностью инвазии от 1 до 184
экз./особь (среднее ИИ = 46 ± 6.1 экз./особь, ЭИ = 96%, ИО = 44 ± 6.0 экз./особь).
Ligophorus llewellyni инфицировал 40 экз. рыб при численности от 1 до 108 экз./особь
(среднее ИИ = 35 ± 4.9 экз./особь, ЭИ = 78%, ИО = 27 ± 4.3 экз./особь). На 10 пи-
ленгасах был встречен L. kaohsianghsieni, его численность колебалась от 1 до 18 экз./
особь (среднее значение ИИ = 5 ± 1.6 экз./особь, ЭИ = 20%, ИО = 1 ± 0.4 экз./особь).
Дисперсионный анализ долей моногеней показал, что по распределению (доле)
трех анализируемых видов правая и левая части жабр не различались достоверно
между собой. В то же время доля особей этих видов значимо зависела от других
участков жабр: полужабры (передней или задней), жаберной дуги (I-IV), сектора
(1-3) и части жаберных лепестков (дистальной или проксимальной), на котором ло-
кализовались моногенеи (табл. 1).
Попарное сравнение выборок L. pilengas и L. llewellyni, собранных c 12 локусов
жабр (3 секторов каждой из 4 дуг), с помощью пост-хок теста ANOVA (табл. 2)1 пока-
зало, что доли особей обоих видов на 2 секторе I дуги значимо (p < 0.05) отличались
от аналогичного показателя на всех секторах других дуг. Кроме того, доли особей
как L. pilengas, так и L. llewellyni на каждом из секторов на II и III дугах достоверно
различались в пределах дуг . Третий вид, L. kaohsianghsieni, паразитирующий на
пиленгасе, не встречен на I дуге, поэтому сравнивали его распределение по девяти
участкам (табл. 3), при этом существенные различия отмечены между 2 и 3 секторами
II жаберной дуги, а также между ними и другими участками жабр.
1 В этот анализ не включены полужабры и части лепестков, поскольку подавляющее
большинство моногеней обоих видов локализовались на одной из двух полужабр и одной из
двух частей лепестков.
506
Таблица 1. Анализ (ANOVA) значимости влияния участка жабр на распределение (долю)
особей видов рода Ligophorus - паразитов Planiliza haematocheilus
Table 1. Analysis (ANOVA) of significance of the gill locus effect on the specimen distribution
(proportions) of the Ligophorus species, parasitising Planiliza haematocheilus
Участок жаберного аппарата
Ligophorus
Ligophorus
Ligophorus
Planiliza haematocheilus (фактор)
pilengas
llewellyni
kaohsianghsieni
Половина жаберного аппарата
0.12 / 0.85 (40)
0.25 / 0.70 (40)
0.11 / 0.90 (10)
(df = 1)
Полужабра (df = 1)
30.5 / < 0.01 (29)
20.0 / < 0.01 (28)
-
Часть лепестка (df = 1)
96.6 / < 0.01 (22)
345.0 / < 0.01 (22)
-
Сектор (df = 2)
93.2 / < 0.01 (49)
98.7 / < 0.01 (40)
4.7 / 0.02 (10)
Дуга (df = 3)
17.4 / < 0.01 (49)
15.3 / < 0.01 (40)
89.0 / < 0.01 (10)
Дуга+сектор (df = 11)
15.5 / < 0.01 (49)
14.5 / < 0.01 (40)
7.0 / < 0.01 (10)
Дуга+сектор+полужабра
10.1 / < 0.01 (29)
7.3 / < 0.01 (28)
-
(df = 23)
Дуга+сектор+полужабра+
5.3 / < 0.01 (22)
5.2 / < 0.01 (22)
-
часть лепестка (df = 47)
Примечания. Для каждого фактора показаны F-критерий / p - уровень значимости
(размер каждой из двух сравниваемых выборок); df - степень свободы. Достоверный
уровень значимости выделен жирным шрифтом. Прочерк - нет данных.
Таблица 2. Фишер LSD тест достоверности различий в распределении Ligophorus pilengas
(правый верхний треугольник) и L. llewellyni (левый нижний треугольник) по 12 участкам
(разным секторам разных дуг) жабр Planiliza haematochelus
Table 2. Fisher LSD test of significance of the differences in the distribution of Ligophorus
pilengas (right upper triangle) and L. llewellyni (left lower triangle) across 12 gill sites
(arches and sectors) of Planiliza haematocheilus
Участки жабр
I 1
I 2
I 3
II 1
II 2
II 3
III 1
III 2
III 3
IV 1
IV 2
IV 3
Ligophorus pilengas
I
1
+
+
-
+
+
-
+
+
-
+
-
I
2
+
-
+
+
+
+
+
+
+
+
+
I
3
+
+
+
-
+
+
-
+
+
+
+
II
1
-
+
+
+
+
-
+
+
-
+
-
II
2
+
+
-
+
+
+
+
-
+
+
+
II
3
+
+
-
+
-
+
+
-
+
-
-
III 1
-
+
+
-
+
+
+
+
-
+
-
III 2
+
+
-
+
-
+
+
+
+
+
+
III 3
-
+
+
-
+
-
-
+
+
-
-
IV 1
-
+
+
-
+
+
-
+
-
+
-
IV 2
-
+
+
-
+
-
-
+
-
-
-
IV 3
-
+
+
-
+
-
-
+
-
-
-
Ligophorus llewellyni
Примечания. I-IV - маркировка жаберной дуги; 1-2 - маркировка сектора жаберной дуги.
Различия между долями особей моногеней на сравниваемых участках жабр при p ≤ 0.05:
«+» - достоверные, «-» - недостоверные. Одинаковым цветом отмечены разные сектора
одной дуги (темно-серый) или одинаковые сектора разных дуг (светло-серый).
507
Таблица 3. Фишер LSD тест достоверности различий в распределении Ligophorus
kaohsianghsieni по 9 участкам (дугам 2 и секторам) жабр Planiliza haematocheilus
Table 3. Fisher LSD тест of significance of the differences in the distribution of Ligophorus
kaohsianghsieni across 9 gill sites (arches and sectors) of Planiliza haematocheilus
Участки жабр
II 1
II 2
II 3
III 1
III 2
III 3
IV 1
IV 2
II
2
+
II
3
+
-
III 1
-
+
+
III 2
-
+
+
-
III 3
-
+
+
-
-
IV 1
-
+
+
-
-
-
IV 2
-
+
+
-
-
-
-
IV 3
-
+
+
-
-
-
-
-
Примечания. I-IV - маркировка жаберной дуги; 1-2 - маркировка сектора жаберной дуги.
Различия между долями особей моногеней на сравниваемых участках жабр при p ≤ 0.05:
«+» - достоверные, «-» - недостоверные. Одинаковым цветом отмечены разные сектора
одной дуги (темно-серый) или одинаковые сектора разных дуг (светло-серый).
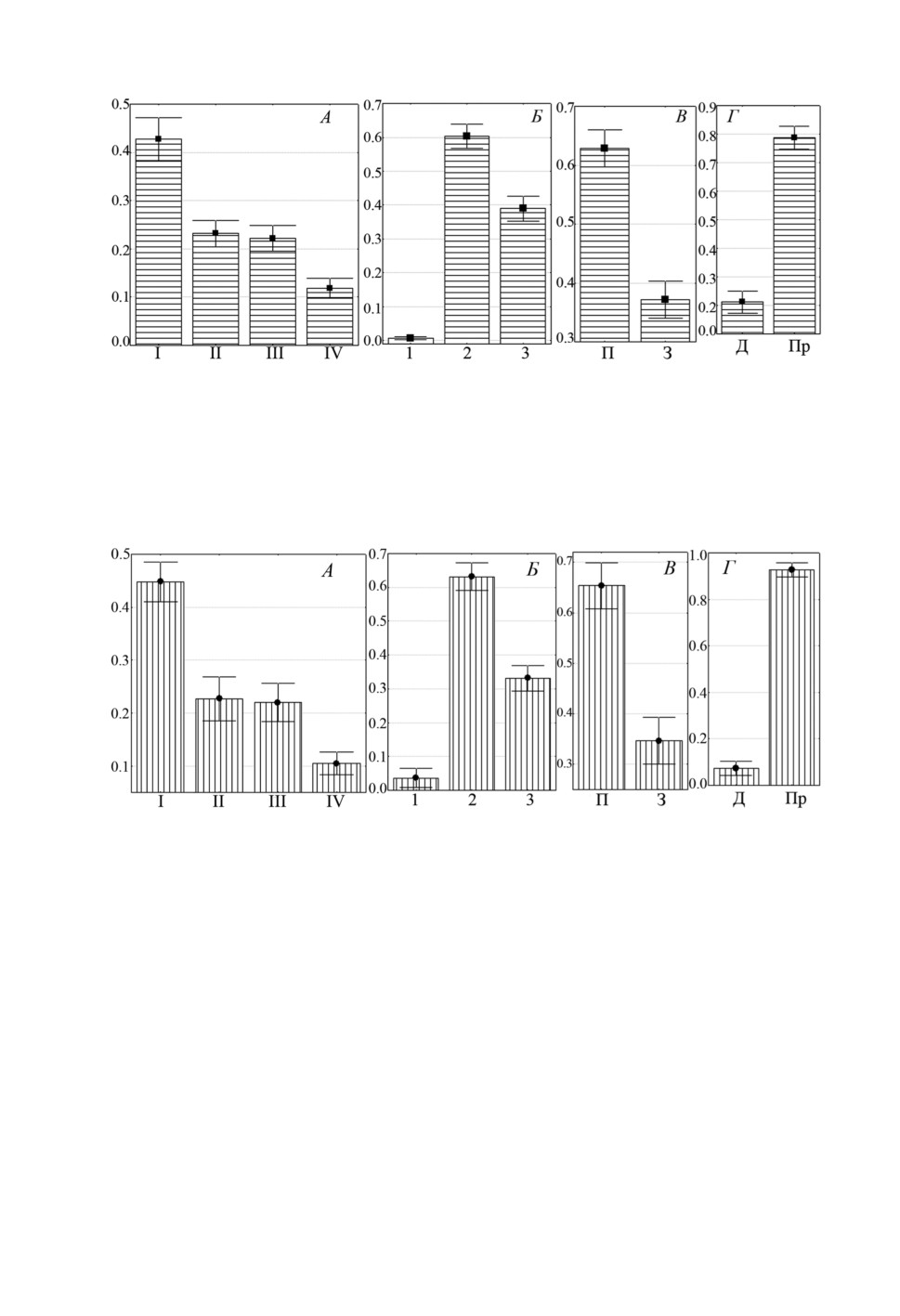
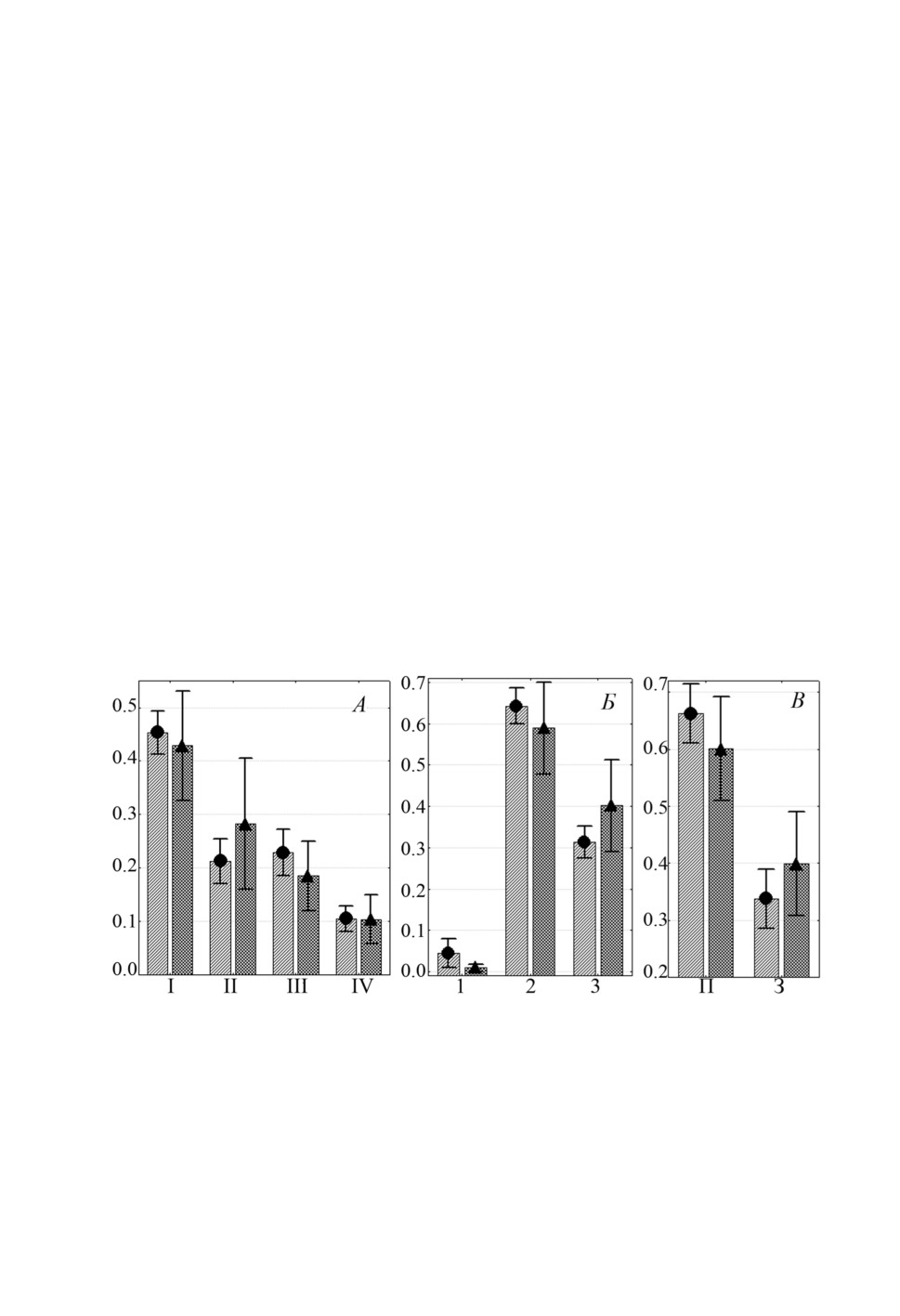
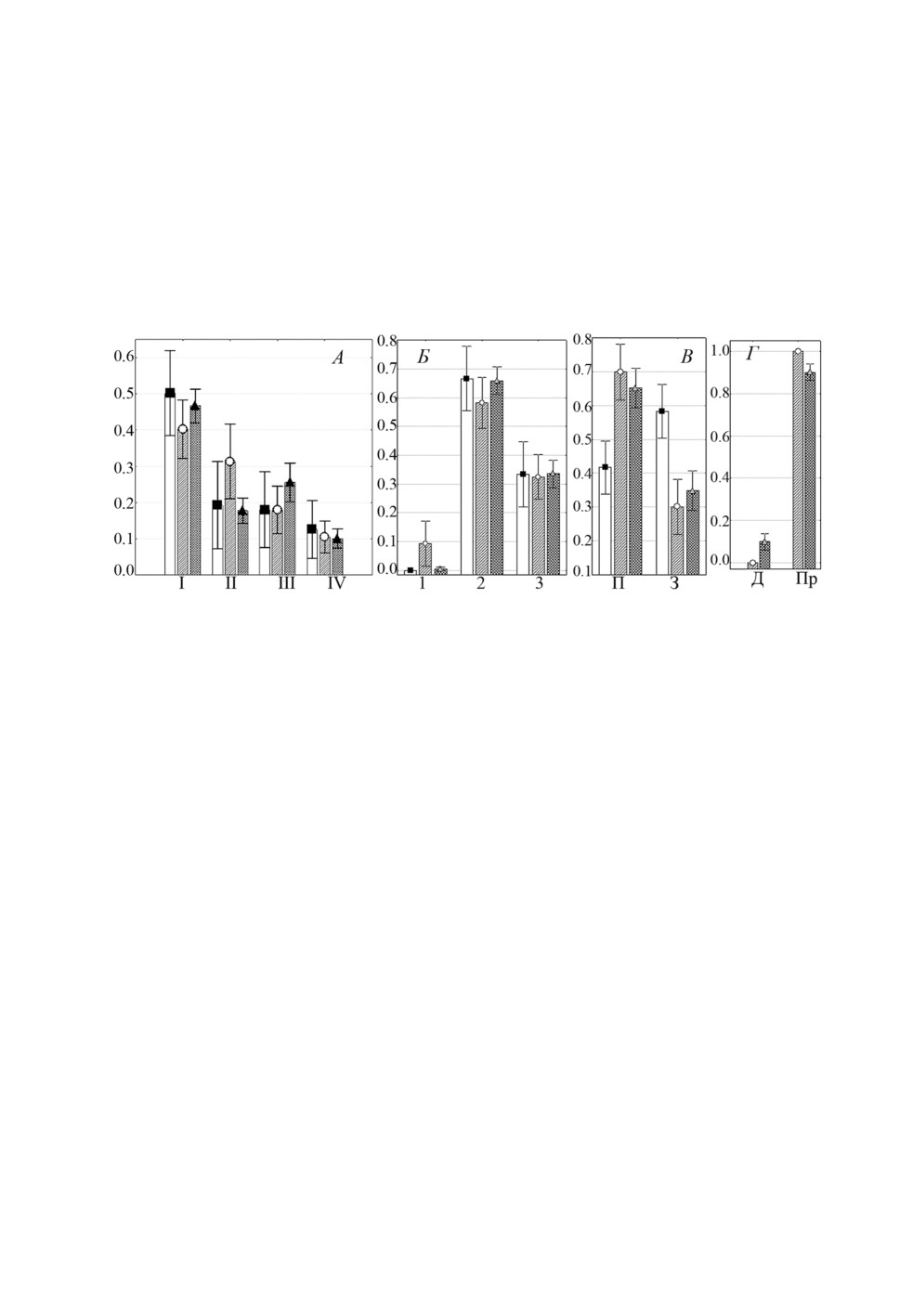
Особи L. pilengas и L. llewellyni (рис. 2А, 2Б и 3А, 3Б) предпочитали локализовать-
ся на одних и тех же участках жабр, а именно, I-III жаберных дугах, при этом более
40% особей каждого вида встречались на I дуге и на каждой жаберной пластинке наи-
большая доля лигофорусов отмечена во 2 секторе. Третий вид - L. kaohsianghsieni -
встречался на жабрах более скученно, около 80% особей локализовалось на II жабер-
ной дуге. На I жаберной дуге и в 1 секторе каждой дуги эти моногенеи не найдены,
во 2 и 3 секторах встречались примерно в равной степени (рис. 4А, 4Б).
Так как не было выявлено различий в распределении по дугам и секторам между
L. pilengas и L. llewellyni, проанализирована их встречаемость по полужабрам (перед-
няя и задняя) и частям жаберных лепестков (проксимальная и дистальная). Однако
и в распределении по этим локусам жаберного аппарата рыб различий между
L. pilengas и L. llewellyni не найдено (рис. 2В, 2Г и 3В, 3Г). Оба вида крайне неравно-
мерно встречались на этих участках: на передней полужабре было сосредоточено,
в среднем, около 65% моногеней обоих видов, при этом на проксимальной части
жаберных лепестков - около 80% особей L. pilengas и практически 100% L. llewellyni.
При этом L. kaohsianghsieni был встречен только на передней полужабре и на прок-
симальных частях жаберных лепестков.
Таким образом, моногенеи всех трех видов предпочитали локализоваться на пе-
редней полужабре и на проксимальных частях жаберных лепестков, в наименьшей
степени встречались на IV жаберной дуге и в 1 секторе. На остальных жаберных
лепестках и в остальных секторах L. kaohsianghsieni распределялся более скученно,
чем два других вида (рис. 4).
2 Данный вид не встречен на I жаберной дуге.
508
Рисунок 2. Распределение (средняя доля ± SE) Ligophorus pilengas на участках жабр
Planiliza haematocheilus: А - жаберных дугах, Б - секторах, В - полужабрах,
Г - частях жаберных лепестков. Обозначения как на рис. 1.
Figure 2. Distribution (mean proportion ± SE) of Ligophorus pilengas on gills of Planiliza
haematocheilus: A - gill arches, Б - sectors, В - hemibranchies, Г - gill filament parts.
Keys as in figure 1.
Рисунок 3. Распределение (средняя доля ± SE) Ligophorus llewellyni на участках жабр
Planiliza haematocheilus: А - жаберных дугах, Б - секторах, В - полужабрах,
Г - частях жаберных лепестков. Обозначения как на рисунке 1.
Figure 3. Distribution (mean proportion ± SE) of Ligophorus llewellyni on gills of Planiliza
haematocheilus: A - gill arches, Б - sectors, В - hemibranchies, Г - gill filament parts.
Keys as in fig. 1.
Одним из факторов, влияющих на распределение моногеней по жабрам рыб, может
быть одновременное присутствие других видов. Из 49 зараженных лигофорусами пи-
ленгасов 40 рыб было заражено более чем одним видом и на 9 рыбах был встречен
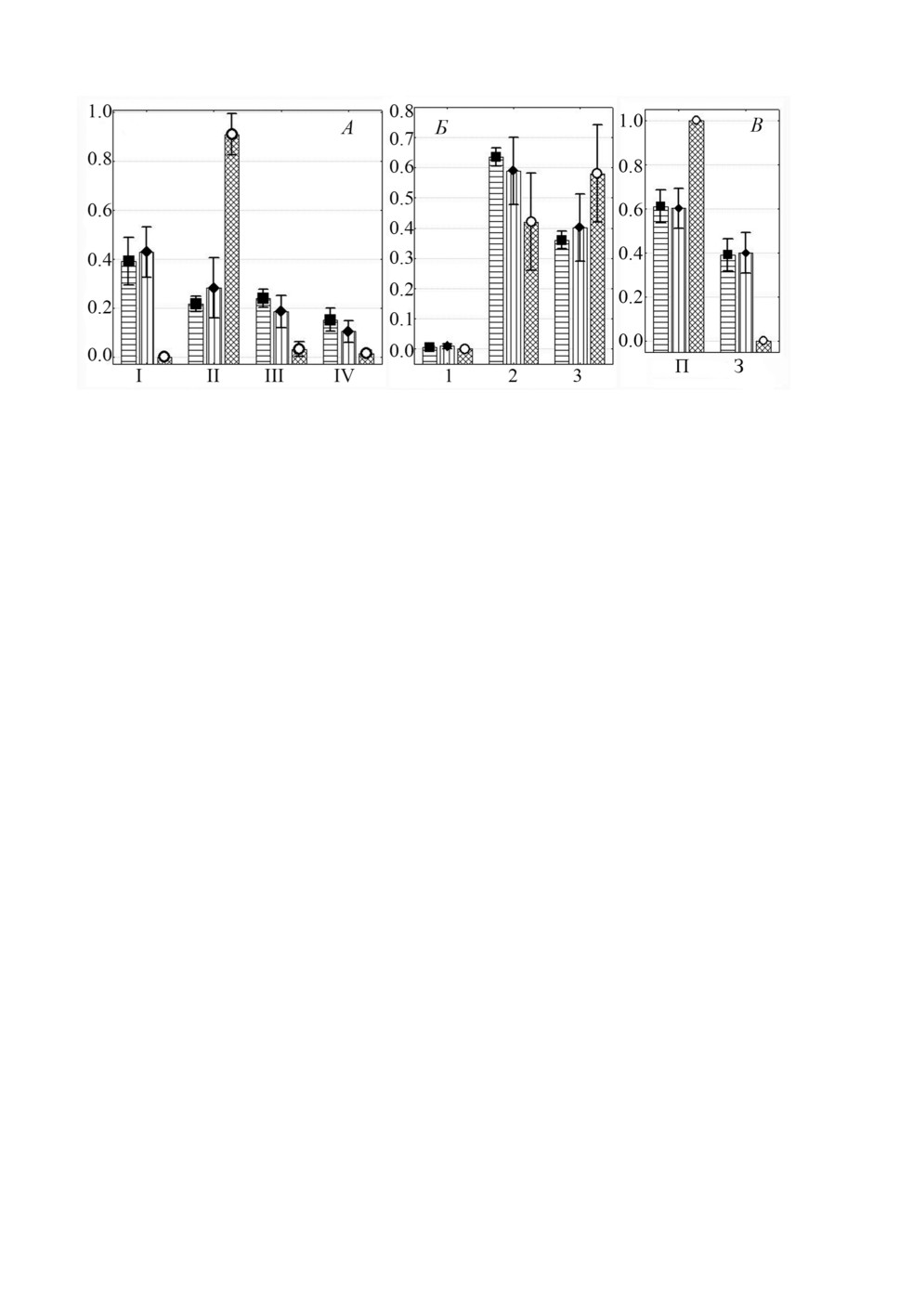
только L. pilengas. При сравнении распределения особей L. pilengas в трех вариантах -
когда только этот вид встречался на жабрах (одновидовая инвазия), когда присут-
ствовали и особи L. llewellyni (30 рыб) и когда к ним присоединялся третий вид
L. kaohsianghsieni (10 рыб) не выявлено существенных различий в общем характере
предпочтения видом L. pilengas определенных участков жабр (рис. 5).
509
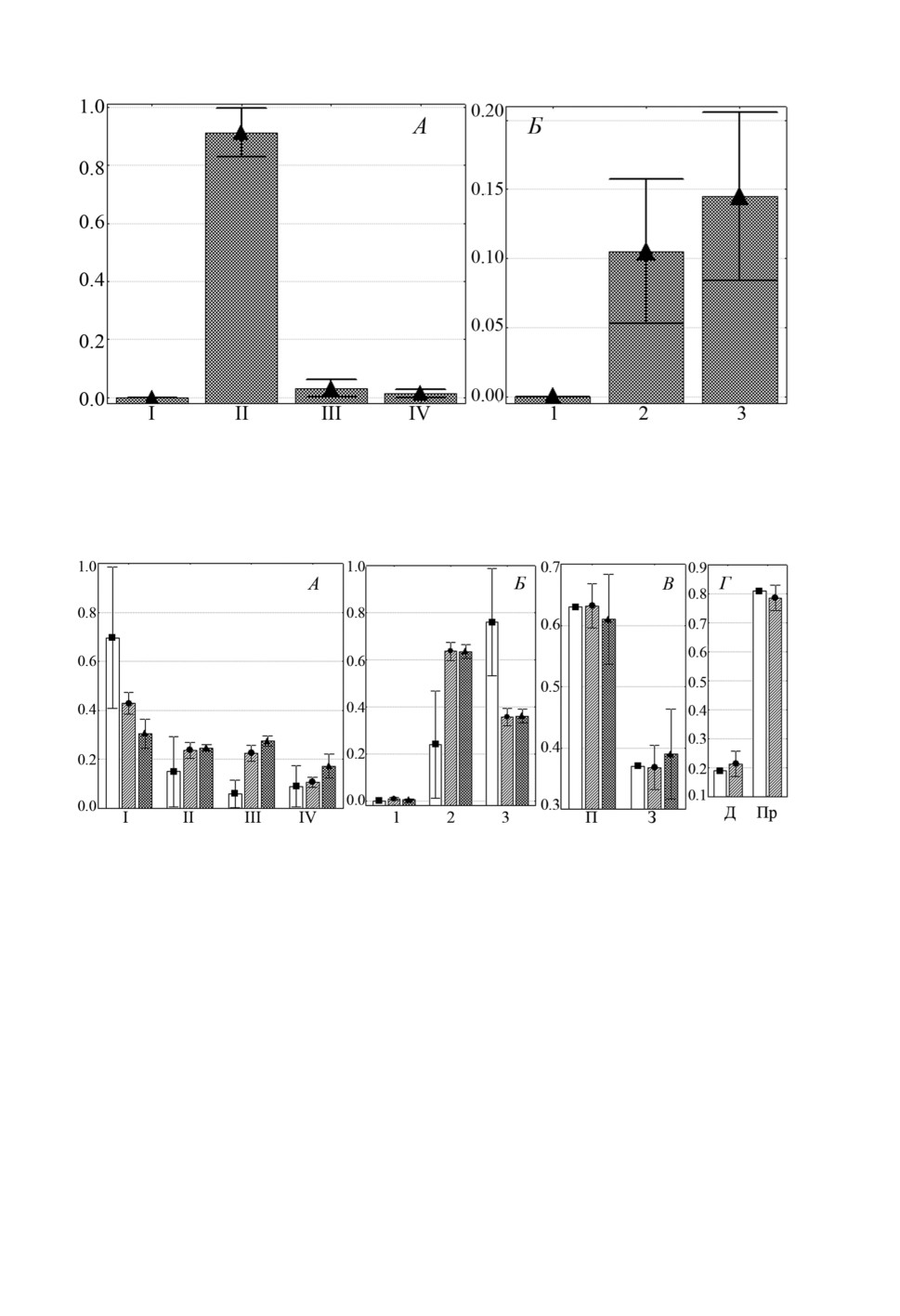
Рисунок 4. Распределение (средняя доля ± SE) Ligophorus kaohsianghsieni на участках жабр
Planiliza haematocheilus: А - жаберных дугах, Б - секторах. Обозначения как на рис. 1.
Figure 4. Distribution (mean proportion ± SE) of Ligophorus kaohsianghsieni on gills
of Planiliza haematocheilus: A - gill arches, Б - sectors. Keys as in fig. 1.
Рисунок 5. Распределение (средняя доля ± SE) Ligophorus pilengas на участках жабр
Planiliza haematocheilus при одновидовой и смешанной с другими видами Ligophorus
инвазии: А - жаберных дугах, Б - секторах, В - полужабрах, Г - частям жаберных
лепестков. Обозначения: ■ - одновидовая инвазия, ● - смешанная только с L. llewellyni,
▲ - смешанная c L. llewellyni и L. kaohsianghsieni, остальные обозначения как на рис. 1.
Figure 5. Distribution (mean proportion ± SE) of Ligophorus pilengas on gills of Planiliza
haematocheilus in infections with single-species vs mixed with other Ligophorus species:
A -gill arches, Б - sectors, В - hemibranchies, Г - gill filament parts.
Keys: ■ - single-species infection, ● - mixed with L. llewellyni infection,
▲ - mixed with L. llewellyni and L. kaohsianghsieni infection, other keys as in fig. 1.
Однако при одновидовой инвазии L. pilengas встречался более неравномерно, боль-
ше половины моногеней локализовались на I дуге (в среднем около 70%) и в 3 сек-
торе (около 75%), тогда как при совместной встречаемости с L. llewellyni и с обоими
видами (L. llewellyni и L. kaohsianghsieni) на этих участках жабр найдено не более
510
40-45% особей этого вида (сравнение долей особей L. pilengas при одновидовой vs
смешанной с другими видами инвазии на I дуге и в 3 секторе: F = 5.43, p = 0.01
и F =10.5, p < 0.01 соответственно). В целом, когда L. pilengas встречался со-
вместно с L. llewellyni, который предпочитал те же участки жабр (I жаберную дугу
и 2 сектор), что и L. pilengas, последний вид распределялся более равномерно между
I-III дугами и 2 и 3 секторами. L. llewellyni встречался только при смешанных инва-
зиях, при этом его локализация не менялась в присутствии только L. pilengas или еще
и третьего вида (рис. 6). Третий вид - L. kaohsianghsieni - найден только совместно
с двумя другими видами. При инвазии тремя видами (рис.
7) даже на II жаберной
дуге, где локализовались практически все особи L. kaohsianghsieni, распределение
(т.е. доля) двух других видов (L. pilengas и L. llewellyni) не изменялось по сравнению
с их распределением в отсутствие этого вида (рис. 5 и 6). Также не менялось распре-
деление моногеней по полужабрам (передней и задней) и частям жаберных лепестков
(дистальной и проксимальной) в зависимости от присутствия других видов (рис. 5Б,
6Б, 7Б и 5В, 6В, 7В).
В целом, каждый из исследованных видов, даже при отсутствии двух или одного
из других видов лигофорусов, паразитирующих на пиленгасе, был ограничен в своем
распределении характерными для каждого вида местами локализации на жабрах рыб,
не меняя существенно своего распределения в зависимости от присутствия других
представителей рода. Ligophorus pilengas в отсутствие других видов встречался даже
более скученно, предпочитая I жаберную дугу (рис. 5А).
Рисунок 6. Распределение (средняя доля ± SE) Ligophorus llewellyni на участках жабр
Planiliza haematocheilus при смешанной инвазии с другими видами Ligophorus:
А -жаберных дугах, Б -секторах, В - полужабрах. Обозначения: ● - только с L. pilengas,
▲ - с L. pilengas и L. kaohsianghsieni, остальные обозначения как на рис. 1.
Figure 6. Distribution (mean proportion ± SE) of Ligophorus llewellyni on gills of Planiliza
haematocheilus in infection mixed with other Ligophorus species: A - gill arches, Б - sectors,
В - hemibranchies. Keys: ● - mixed with L. pilengas infection, ▲ - with L. pilengas
and L. kaohsianghsieni infection, other keys as in fig. 1.
511
Рисунок 7. Распределение (средняя доля ± SE) Ligophorus spp. на участках жабр Planiliza
haematocheilus при смешанной инвазии тремя видами: А - жаберных дугах, Б - секторах,
В - полужабрах. Обозначения: ■ - L. pilengas, ♦ - L. llewellyni, ○ - L. kaohsianghsieni,
остальные обозначения как на рис. 1.
Figure 7. Distribution (mean proportion ± SE) of Ligophorus spp. on gills of Planiliza
haematocheilus in three species mixed infection: A - gill arches, Б -sectors, В -hemibranchies.
Keys: ■ - L. pilengas, ♦ - L. llewellyni, ○ - L. kaohsianghsieni, other keys as in fig. 1.
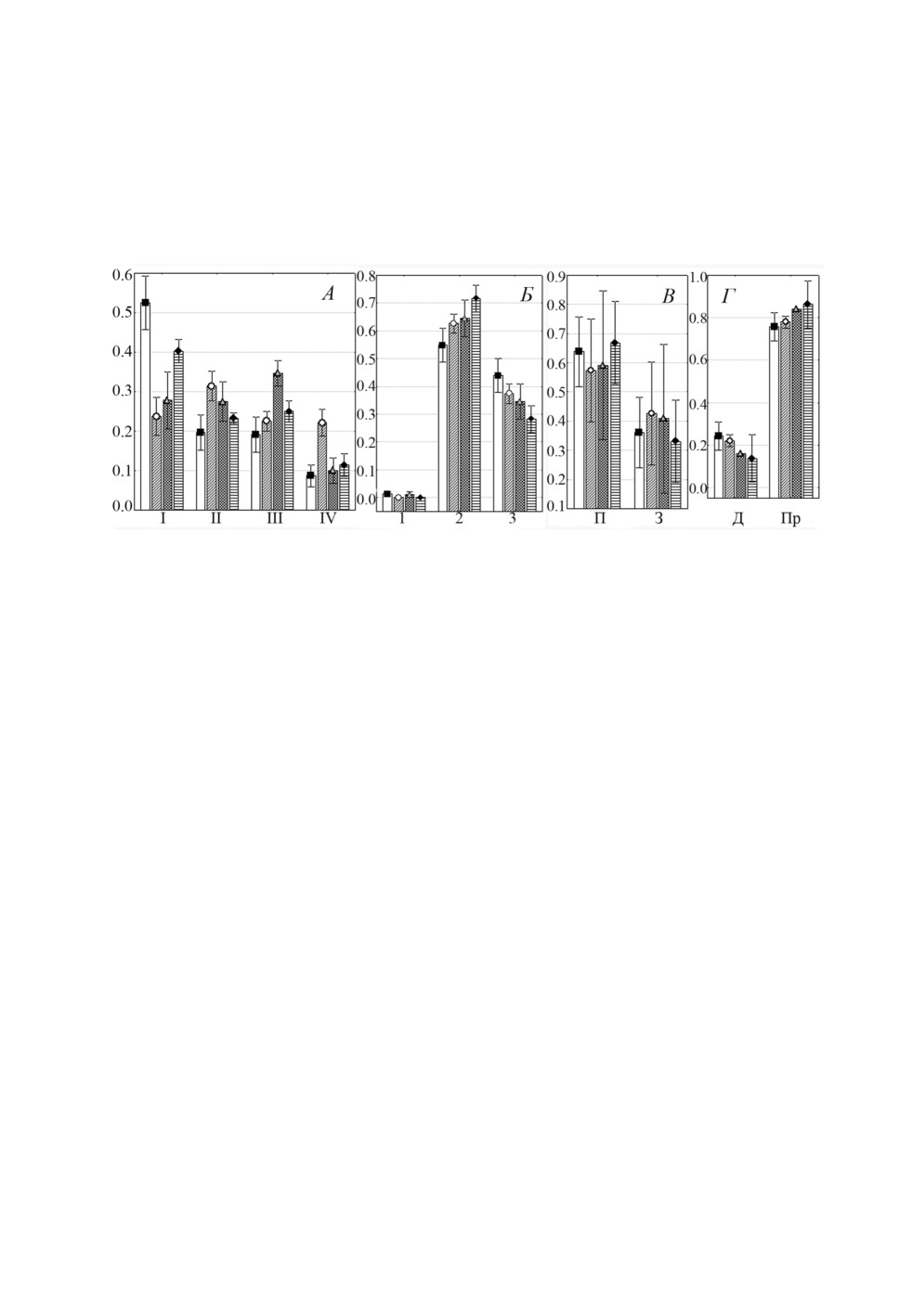
Другой фактор, который может влиять на характер локализации моногеней на
жабрах, - это их численность. При наименьшей численности (ИИ ≤ 20 экз./особь)
более половины особей L. pilengas локализовались на I жаберной дуге, по мере уве-
личения интенсивности инвазии достоверно более равномерно распределяясь между
другими дугами (сравнение долей моногеней на I жаберной дуге при ИИ ≤ vs >
20 экз./особь: F =
6.58, p = 0.015). Однако даже при высокой численности (ИИ >
100 экз./особь) наименьшая доля моногеней (в среднем около 10%) этого вида встре-
чалась на IV дуге (рис.
8А). При всех значениях численности больше половины
особей L. pilengas локализовалось во 2 секторе, и при увеличении ИИ доля моногеней
на этом секторе имела тенденцию возрастать, достигая 70% при максимальных зна-
чениях этого показателя (рис. 8Б). Однако сравнение доли особей во 2 секторе при
ИИ ≤ 20 vs >100 экз./особь не выявило достоверной разницы между ними: F = 2.14,
p = 0.16. При этом даже при максимальной численности лигофорусы не встречались
в 1 секторе. Менее 25% особей L. pilengas локализовались на дистальных частях ле-
пестков, даже когда их численность превышала 100 экз. (рис. 8В). При всех значениях
ИИ сохранялось распределение этих моногеней на передней и задней полужабрах,
примерно 40 и 60% соответственно (рис. 8Г).
Особи второго анализируемого вида - L. llewellyni - при численности менее
70 экз./особь предпочитали локализоваться на I жаберной дуге (более 40%) и толь-
ко при максимальных значениях ИИ доля моногеней, встречающихся на этой дуге,
снижалась, увеличиваясь при этом на II дуге, и распределение между этими дугами
(I и II) выравнивалось (рис. 9А). Так, доли особей на I и II жаберной дугах при
ИИ ≤ 70 экз./особь достоверно различались: F = 7.3, p < 0.01, тогда как при ИИ >
70 экз./особь были статистически равны: F = 2.3, p = 0.13. При этом распределе-
ние L. llewellyni по секторам не менялось, даже при максимальной численности во
512
2 секторе локализовалось 70% особей этого вида (рис. 9Б). При низкой численности
(ИИ < 10 экз./особь) пропорции этих моногеней на полужабрах были почти равны
(рис. 9В), тогда как при росте ИИ их доля на передней полужабре возрастала и была
достоверно выше, чем на задней полужабре (сравнение долей особей на передней vs
задней полужабре при ИИ > 10 экз./особь: F = 76.7, p < 0.01). При всех значениях
интенсивности инвазии большинство особей L. llewellyni локализовалось на прокси-
мальных частях жаберных лепестков (рис. 9Г).
Рисунок 8. Распределение (средняя доля ± SE) Ligophorus pilengas на участках жабр
Planiliza haematocheilus при разных значениях интенсивности инвазии: А - жаберных дугах,
Б - секторах, В - полужабрах, Г - частях жаберных лепестков.
Обозначения: ■ -1-20 экз./особь, ○ -21-60 экз./особь, ▲- 61-100 экз./особь,
♦ - 101-200 экз./особь, остальные обозначения как на рис. 1.
Figure 8. Distribution (mean proportion ± SE) of Ligophorus pilengas on gills of Planiliza
haematocheilus at different ranges of infection intensity: A - gill arches, Б - sectors,
В - hemibranchies, Г - gill filament parts. Keys: ■ - 1-20 spec./host, ○ - 21-60 spec./host,
▲- 61-100 spec./host, ♦ - 101-200 spec./host, other keys as in fig. 1.
Таким образом, общий характер распределения на жабрах пиленгаса как
L. pilengas, так и L. llewellyni не менялся существенно при изменении их численно-
сти. При всех уровнях интенсивности инвазии минимальная доля моногеней обоих
видов отмечена на IV жаберной дуге, а большинство лигофорусов встречалось во
2 секторе, на передней полужабре и проксимальных частях жаберных лепестков. При
значениях ИИ менее 10 экз./особь наибольшая доля особей обоих видов встречена на
I жаберной дуге, при этом при росте численности отмечена тенденция к выравнива-
нию пропорций лигофорусов, встречающихся на I и II жаберной дугах.
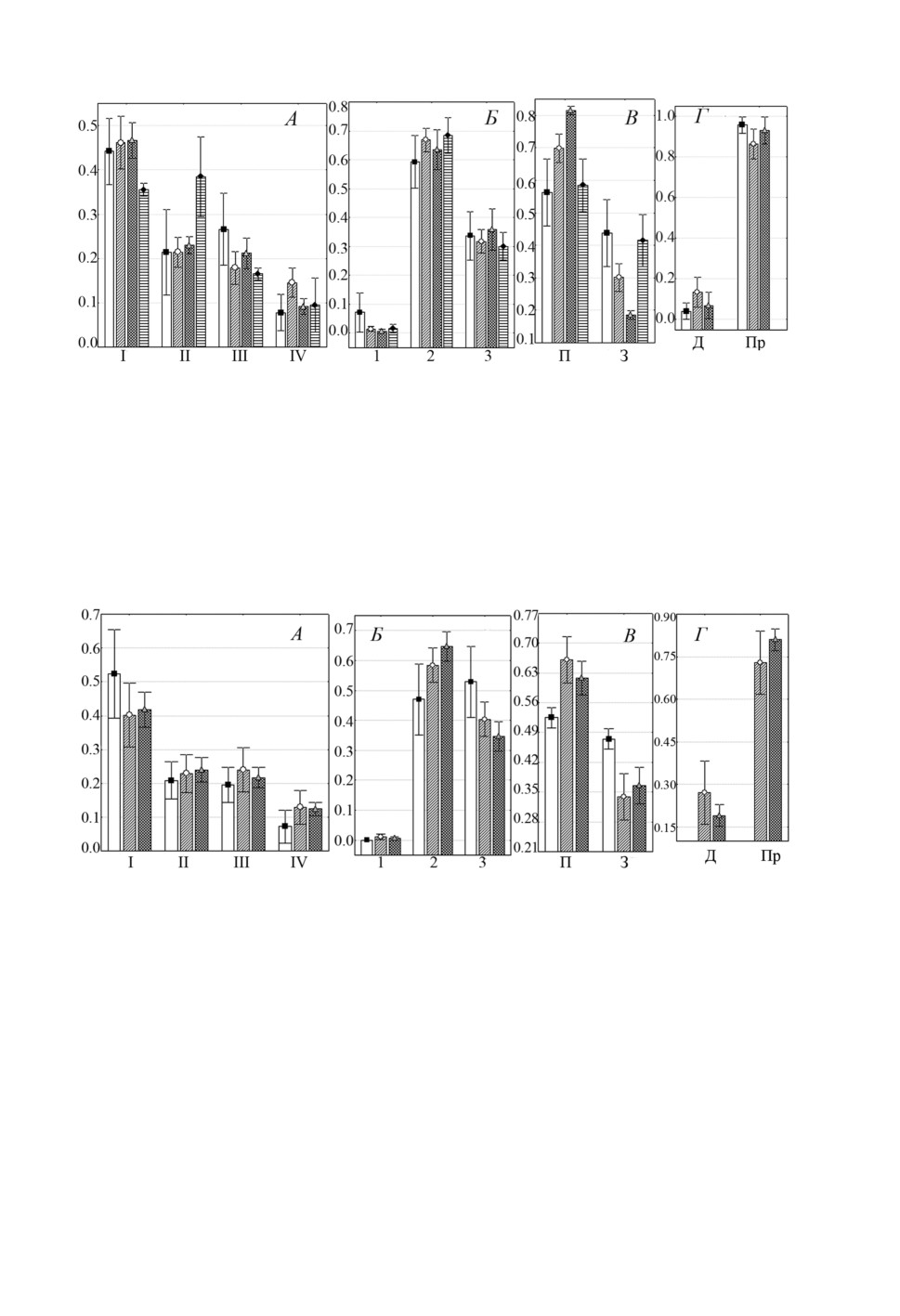
Длина рыб, в целом определяющая размер их жаберного аппарата, также может
влиять на характер встречаемости видов Ligophorus на разных участках жабр. Ана-
лиз зависимости распределения L. pilengas по участкам жабр от размера хозяина
(рис. 10) показал, что предпочтение этим видом I жаберной дуги, 2 и 3 секторов,
а также проксимальных частей лепестков сохранялось для рыб всех размерных
классов. Однако по мере роста жабр отмечено увеличение неравномерности рас-
пределения моногеней между секторами (рис. 10Б, 10В). У рыб с наименьшей дли-
ной тела доли особей L. pilengas, локализующихся на 2 и 3 секторах, практически
равны, а при увеличении этого параметра - достоверно выше во 2 секторе (срав-
нение долей особей на 2 vs 3 секторах при TL рыбы > 30 см: F=21.3, p < 0.01).
513
Рисунок 9. Распределение (средняя доля ± SE) Ligophorus llewellyni на участках жабр
Planiliza haematocheilus при разных значениях интенсивности инвазии: А - жаберных дугах,
Б - секторах, В - полужабрах, Г - частях жаберных лепестков.
Обозначения: ■ -1-10 экз./особь, ○ - 11-40 экз./особь, ▲- 41-70 экз./особь,
♦ - 71-110 экз./особь, остальные обозначения как на рис. 1.
Figure 9. Distribution (mean proportion ± SE) of Ligophorus llewellyni on gills of Planiliza
haematocheilus at different ranges of infection intensity: A - gill arches, Б - sectors,
В - hemibranchies, Г - gill filament parts. Keys: ■ - 1-10 spec./host, ○ - 11-40 spec./host,
▲- 41-70 spec./host, ♦ - 71-110 spec./host, other keys as in fig. 1.
Рисунок 10. Распределение (средняя доля ± SE) Ligophorus pilengas на участках жабр
Planiliza haematocheilus при разных значениях длины рыбы: А - жаберных дугах,
Б - секторах, В - полужабрах, Г - частях жаберных лепестков. Обозначения: ■ - 13-30 см,
○ - 31-40 см, ▲- 41-66 см, остальные обозначения как на рис. 1.
Figure 10. Distribution (mean proportion ± SE) of Ligophorus pilengas on gills of Planiliza
haematocheilus at different ranges of fish total length: A - gill arches, Б -sectors,
В - hemibranchies, Г - gill filament parts. Keys: ■ -13-30 см, ○ - 31-40 см,
▲- 41-66 см, other keys as in fig. 1.
С ростом длины тела рыб также отмечено изменение соотношения долей особей
L. pilengas, локализующихся на полужабрах (рис. 10В): они примерно равны при
TL ≤ 30 см и достоверно различаются при увеличении этого показателя (сравне-
ние долей особей на передней vs задней полужабрах при TL > 30 см: F = 30.4,
514
p < 0.01).Аналогичные закономерности отмечены и для второго вида - L. llewellyni
(рис. 11). Доли особей этого вида на жаберных дугах, в секторах и частях лепестков
практически не менялись при увеличении размера рыб (рис. 11А, 11Б, 11Г). Одна-
ко отмечено изменение распределения моногеней, локализующихся на полужабрах
(рис. 11В). Так, у рыб размерами ≤ 30 см только 40% L. llewellyni встречались на
передней полужабре, а по мере роста рыб доля моногеней на этой полужабре увели-
чивалась вдвое, при этом разница между долями особей на полужабрах становилась
достоверной(сравнение долей особей на передней vs задней полужабрах при TL рыбы
> 30 см: F = 23.3, p < 0.01).
Рисунок 11. Распределение (средняя доля ± SE) Ligophorus llewellyni на участках жабр
Planiliza haematocheilus при разных значениях длины рыбы: А - жаберных дугах,
Б - секторах, В - полужабрах, Г - частях жаберных лепестков. Обозначения: ■ -13-30 см,
○ - 31-40 см, ▲- 41-66 см, остальные обозначения как на рис. 1.
Figure 11. Distribution (mean proportion ± SE) of Ligophorus llewellyni on gills of Planiliza
haematocheilus at different ranges of fish total length: A - gill arches, Б - sectors,
В - hemibranchies, Г - gill filament parts. Keys: ■ -13-30 см, ○ - 31-40 см, ▲- 41-66 см,
other keys as in fig. 1.
Таким образом, по мере увеличения площади жабр пиленгаса существенно меня-
лось распределение обоих видов лигофорусов только на полужабрах, при этом при
росте длины тела рыб возрастала доля моногеней на передней полужабре.
ОБСУЖДЕНИЕ и ЗАКЛЮЧЕНИЕ
В результате установлено, что все три вида Ligophorus, паразитирующих на
Planiliza haematocheilus, достоверно неравномерно встречаются на разных участках
жаберного аппарата рыб (на I-IV жаберных дугах, передней/задней полужабрах,
в 1-3 секторах и на проксимальной/дистальной частях жаберных лепестков). При
этом между L. pilengas и L. llewellyni не найдено никаких существенных различий
в их распределении по выделенным локусам жабр пиленгаса. Ранее проведенный
анализ локализации на жабрах рыб L. vanbenedeni и L. szidati, паразитирующих на
Chelon auratus, а также L. acuminatus и L. szidati, встречающихся на C. saliens, выявил
существенные различия между видами как в распределении на жабрах их хозяев
(Пронькина и др., 2010), так и в морфологии прикрепительных образований (Euzet,
Suriano, 1977). Тогда как анализируемая пара видов - L. pilengas и L. llewellyni -
515
имеет сходную форму и размеры структур прикрепительного диска (Dmitrieva et al.,
2007), что может объяснить схожесть их распределения на жабрах рыб. Третий вид -
L. kaohsianghsieni, - паразитирующий на пиленгасе, имеет крючки и пластинки при-
крепительного диска, отличные от прикрепительных образований двух других видов
(Гусев, 1985). Практически все его особи найдены на медиальном (2) и вентральном
(3) секторах II жаберной дуги. Такая скученная локализация, вероятно, определяет-
ся, в первую очередь, его невысокой численностью, которая в 4-8 раз меньше, чем
у двух других видов. Однако форма крючков - массивных с широкими лезвиями
(Гусев, 1985) - также может определять выбор этими моногенеями для прикрепления
именно II жаберной дуги, которая имеет как наибольшее количество филаментов, так
и наибольшую общую площадь их поверхности (Caltran, Silan, 1996).
Многие авторы, изучавшие распределение моногеней по жаберному аппарату рыб,
связывали неравномерность этого распределения с силой водного потока через разные
участки жабр и находили то прямую зависимость между интенсивностью тока воды
и долей или численностью червей (Hanek, Fernando, 1978; Gutiérrez, Martorelli, 1994),
то обратную (Paling, 1969; Arme, Halton, 1972; Wootten, 1974; Изюмова, Жарикова,
1982; Ясюк, 1986; Герасев, Старовойтов, 1988).
Исследованные нами виды локализовались преимущественно на хорошо омывае-
мых водой II и III жаберных дугах (Paling, 1968; Hanek, Fernando, 1978; Ясюк, 1986).
При этом они предпочитали сектора (медиальный и вентральный), через которые
проходит наибольший объем воды, но в пределах этих секторов большинство особей
прикреплялось на менее омываемой проксимальной части лепестков.
Площадь участка жабр рассматривалась в ряде исследований как фактор, в зна-
чительной степени определяющий количество моногеней, прикрепляющихся к нему
(Adams, 1986; Buchmann, 1989; Доровских, 1991; Koskivaara et al., 1992). Согласно
результатам анализа количества филаментов и площади поверхности разных участ-
ков жабр Chelon ramada (Caltran, Silan, 1996), наибольшую поверхность жаберных
лепестков имеет II, затем I жаберные дуги, а наименьшую - IV. Задние полужабры
II и I дуги, а затем III дуги имеют бóльшие поверхности, чем передние полужабры
этих дуг. Исследованные нами виды Ligophorus предпочитали локализоваться на жа-
берных дугах (I-III) с наибольшей поверхностью для прикрепления, но существенно
бóльшие их доли были представлены на передних полужабрах, которые по потенци-
альной площади для прикрепления моногеней меньше, чем задние. При этом, как
отмечено выше, через эти участки жабр проходит наибольшей ток воды.
Таким образом, вероятно, оба фактора - и размер участка жабр, и сила тока воды
через него - совместно определяют выбор лигофорусами места локализации. Ранее
к таким же выводам пришли и другие авторы, изучавшие распределение жаберных
моногеней (Wootten, 1974; Gutiérrez, Martorelli, 1999; Rubio-Godoy, 2008). При этом
эти авторы отмечали, что если большинство особей какого-либо вида локализовалось
на бóльших по площади и хорошо омываемых водой дугах, то здесь они предпочи-
тали прикрепляться к участкам с меньшим током воды.
Возможно, первоначальное распределение личинок и постличиночных стадий по
жаберным дугам и полужабрам носит пассивный характер и определяется силой тока
воды через них. Однако по мере развития прикрепительных структур виды могут
менять свою локализацию, выбирая участки с менее интенсивным током воды. Так,
исследованные нами лигофорусы предпочитают локализоваться на проксимальных
516
участках жаберных лепестков I-III жаберных дуг, через которые в пределах этих дуг
проходит наименьший ток воды.
В ряде работ (Mouillot et al., 2005; Šimková, Morand, 2008) было показано, что
межвидовые взаимодействия, по-видимому, являются важным фактором, определяю-
щим структуру сообществ моногеней рода Dactylogyrus. Для четырех из семи видов
Dactylogyrus, паразитирующих на жабрах плотвы, было отмечено выравнивание их
распределения на жаберных дугах в месяцы, когда отсутствовали три других вида
(Koskivaara et al., 1992), хотя численность дактилогирусов в эти периоды также
существенно уменьшалась. Однако изученное нами распределение L. pilengas и
L. llewellyni по жабрам пиленгаса не зависело в значительной степени от присутствия
второго вида из каждой пары.
Численность (ИИ) инфрапопуляций моногеней - важный фактор, который мо-
жет оказывать влияние на их локализацию на разных участках жаберного аппарата
рыб. В целом, можно было бы ожидать, что при низкой численности паразитов их
распределение будет более неравномерно и будет иметь случайный характер, тогда
как при увеличении интенсивности инвазии моногенеи будут занимать большее про-
странство жабр и будут распределяться более равномерно. Однако, несмотря на то
что такая тенденция наблюдалась в нашем исследовании, она не была значительна и
не приводила к увеличению доли лигофорусов на тех участках жабр, на которых они
встречаются редко. Таким образом, общий характер встречаемости изученных видов
Ligophorus на жабрах кефалей не зависел от численности их инфрапопуляций, но при
увеличении интенсивности инвазии практически для всех видов отмечена тенденция
к выравниванию их распределения на тех участках жабр, на которых они предпо-
читают локализоваться. Эти результаты согласуются с данными, ранее полученными
для других моногеней (Arme, Halton, 1972; Жарикова, 1986; Доровских, Матрохина,
1987; Buchmann, 1989; Koskivaara et al., 1992).
Вероятно, размер жаберного аппарата рыб будет оказывать влияние на локализа-
цию паразитов на разных участках жабр, поэтому была проанализирована зависимость
распределения лигофорусов на жабрах от размера хозяина. Наиболее значимо при
увеличении размера пиленгаса менялись доли L. pilengas и L. llewellyni на разных
полужабрах, с ростом рыбы доли обоих видов на передней полужабре существенно
возрастали. Вероятно, такое перераспределение моногеней на участках жабр связано
с неравномерным увеличением площади филаментов жаберных пластинок по мере
увеличения размера рыб. Исследования структуры жабр Chelon ramada (Caltran, Silan,
1996) показали, что с увеличением размера рыб площадь передних полужабр I-III дуг
растет интенсивнее, чем задних. Таким образом, при увеличении длины рыб отмеча-
ется рост доли лигофорусов на тех участках жабр, площадь которых в наибольшей
степени положительно зависит от размера хозяина. Аналогичная зависимость была
отмечена в распределении Ancyrocephalus paradoxus, большинство особей которого
локализуется на истмусе жабр судака у рыб младших возрастов, а по мере роста
хозяев увеличивается доля моногеней на IV и I жаберной дугах, площадь которых
растет с возрастом (Старовойтов, 1986, 1989).
ФИНАНСИРОВАНИЕ РАБОТЫ
Исследование поддержано темой гос. задания Института биологии южных морей
имени А.О. Ковалевского, номер 121030100028-0. Никаких дополнительных грантов
на проведение или руководство данным конкретным исследованием получено не было.
517
СОБЛЮДЕНИЕ ЭТИЧЕСКИХ СТАНДАРТОВ
В данной работе отсутствуют исследования человека или животных.
КОНФЛИКТ ИНТЕРЕСОВ
Авторы данной работы заявляют, что у них нет конфликта интересов.
СПИСОК ЛИТЕРАТУРЫ
Герасев П.И., Старовойтов В.К. 1988. Распределение Ancyrocephalus paradoxus (Monogenea) по жабрам
взрослых судаков Stizostedion lucioperca Куршского залива. Труды ЗИН АН СССР 171: 89-99.
[Gerasev P.I., Starovoitov V.K. 1988. Distribution of Ancyrocephalus paradoxus (Monogenea) on the
gills of adult pikeperch Stizostedion lucioperca of the Curonian Gulf. Trudy ZIN AN SSSR 171: 89-99.
(in Russian)].
Гусев А.В. 1983. Методика сбора и обработки материалов по моногенеям, паразитирующих у рыб. Ленин-
град, Наука, 46 с. [Gusev A.V. 1983. Methods of collection and processing of materials on monogeneans
parasitising fish. Leningrad, Nauka, 46 с. (In Russian)].
Гусев А.В. 1985. Отряд Dactylogyridae. В кн.: Бауер О.Н. (ред.) Определитель паразитов пресноводных
рыб фауны СССР. Т. 2. Паразитические многоклеточные (первая часть). Ленинград, Наука, 15-251.
[Gussev A.V. 1985. Order Dactylogyridea. In: Bauer O.N. (ed.) Keys to the parasites of freshwater fishes
of the USSR fauna. V. 2. Metazoan parasites. Leningrad, Nauka, 15-251. (in Russian)].
Догель В.А. 1949. Явление «сопряженных видов» у паразитов и эволюционное значение этого явления.
Известия АН КазССР. Сер. Паразитология 7: 3-15. [Dogel V.A. 1949. The event of «co-occurring
species « in parasites and the evolutionary significance of this phenomenon. Izvestiya AN KazSSR. Ser.
Parasitologiya 7: 3-15. (in Russian)].
Доровских Г.Н., Матрохина С.Н. 1987. Распределение некоторых видов паразитов на жабрах ерша. Па-
разитология 21 (1): 64-68. [Dorovskikh G.N., Matrokhina S.N. 1987. Distribution of some species of
parasites on gills of ruff. Parazitologia 21 (1): 64-68. (in Russian)].
Доровских Г.Н. 1991. Распределение Dactylogyrus cordus (Monogenea: Dactylogyridae) на жабрах ельца
(Leuciscus leuciscus L.) в условиях нормоксии и гипоксии. Паразитология 25 (2): 163-167. [Dorovskikh
G.N. 1991. Distribution of Dactylogyrus cordus (Monogenea: Dactylogyridae) on the gills of the dace,
Leuciscus leuciscus L. under the conditions of normoxia and hypoxia. Parazitologia 25 (1): 163-167.
(in Russian)].
Жарикова Т.И. 1986. Влияние скорости течения на численность и локализацию паразитов рода Dactylogyrus
Dies., 1850 (Monogenea). Труды Института биологии внутренних вод АН СССР 53/56: 185-194.
[Zharikova T.I. 1986. Influence of flow velocity on the abundance and localisation of parasites of the
genus Dactylogyrus Dies., 1850 (Monogenea). Trudy of the Institute of Biology of Inland Waters AN
SSSR 53/56: 185-194. (in Russian)].
Изюмова Н.А., Жарикова Т.И. 1982. О некоторых особенностях распределения Dactylogyrus anchoratus и
D. сhranilowi (Monogenoidae (Beneden) Bychowsky, 1937; Dactylogyridea Bychowsky, 1937) на жа-
брах карася, карпа и синца. Труды Института биологии внутренних вод АН СССР 46: 89-100.
[Iziumova N.A., Zharikova T.I. 1982. On some peculiarities of distribution of Dactylogyrus anchoratus
and D. schranilowi (Monogenoidae (Beneden) Bychowsky, 1937; Dactylogyridea Bychowsky, 1937) on
the gills of goldfish, common carp and blue bream. Trudy Instituta biologii vnutrennikh vod AN SSSR
46: 89-100. (in Russian)].
Пронькина Н.В., Дмитриева Е.В., Герасев П.И. 2010. Распределение двух видов рода Ligophorus Euzet
et Suriano, 1977 (Plathelminthes: Monogenea) по жабрам Liza aurata (Risso, 1810) (Pisces: Mugilidae)
в Черном море. Морской экологический журнал 9 (1): 53-62. [Pronkina N.V., Dmitrieva E.V., Gerasev
P.I. Distribution of two species of Ligophorus Euzet et Suriano, 1977 (Plathelminthes: Monogenea) on
gill of Liza aurata (Risso, 1810) (Pisces: Mugilidae) from the Black Sea. Morskoj ekologicheskij zhurnal
9 (1): 53-62. (In Russian)].
Старовойтов В.К. 1986. Особенности локализации Ancyrocephalus paradoxus (Monogenea) на суда-
ке Stizostedion lucioperca. Паразитология 20 (6): 491-492. [Starovoitov V.K. 1986. Peculiarities
of localization of Ancyrocephalus paradoxus (Monogenea) on the pike perch Stizostedion lucioperca.
Parazitologia 20 (6): 491-492. (in Russian)].
Старовойтов В.К. 1989. Распределение Ancyrocephalus paradoxus (Monogene, Ancyrocephalidae) на жабрах
судака. Паразитология 23 (1): 40-47. [Starovoitov V.K. 1989. Distribution of Ancyrocephalus paradoxus
(Monogene, Ancyrocephalidae) on the pike perch gills. Parazitologia 23 (1): 40-47. (in Russian)].
518
Ясюк В.П. 1986. О некоторых особенностях распределения дактилогирусов и глохидий двухстворчатых
моллюсков на жабрах леща, плотвы и синца. В кн.: Экология и физиология рыб Куйбышевского во-
дохранилища. Ульяновск, УГПИ им. И.Н. Ульянова, 129-135. [Yasyuk V.P. 1986. On some peculiarities
of distribution of dactylogirids and glochidium of bivalve mollusks on gills of common bream, roach and
blue bream. Ekologiya i fiziologiya ryb Kuybyshevskogo vodokhranilishcha. Ulyanovsk, I.N. Ulyanov
USPI, 129-135. (in Russian)].
Abu Samak O.A., Hassan S.H. 1998. Microhabitat and distribution of three monogenean parasites on the gills of
the mugilid fish, Liza ramada. Journal of the Egyptian-German Society of Zoology. Invertebrate Zoology
& Parasitology 26 (D): 273-283.
Adams A.M. 1986. The parasite community on the gills of Fundulus kansae (Garman) from the South Platte
River, Nebraska (USA). Acta Parasitologica 31: 47-54.
Arme C., Halton D.W. 1972. Observations on the occurrence of Diclidophora merlangi (Trematoda:
Monogenea) on the gills of whiting, Gadus merlangus. Journal of Fish Biology 4 (1): 27-32.
Buchmann K. 1989. Microhabitats of monogenean gill parasites on European eel (Anguilla anguilla). Folia
Caltran H., Silan P. 1996. Gill filament of Liza ramada, a biotope for ectoparasites surface area acquisition using
image analysis and growth models. Journal of Fish Biology 49: 1269 - 1279.
Cerfontaine P. 1896. Contribution a l’etude des Octocotylides. Archives de Biologie 14 (3): 497-560.
Dmitrieva E.V., Gerasev P.I., Gibson D.I., Pronkina N.V., Galli P. 2012. Descriptions and a morphological grouping
of eight new species of Ligophorus Euzet & Suriano, 1977 (Monogenea: Ancyrocephalidae) from Red Sea
Dmitrieva E.V., Gerasev P.I., Pronkina N.V. 2007. Ligophorus llewellyni n. sp. (Monogenea: Ancyrocephalidae)
from the redlip mullet Liza haematocheilus (Temminck & Schlegel) introduced into the Black Sea from
Euzet L., Sanfilippo D. 1983. Ligophorus parvicirrus n. sp. (Monogenea, Ancyrocephalidae) parasite de Liza
ramada (Risso, 1826) (Teleostei, Mugilidae). Annales de Parasitologie Humaine et Comparée 58 (4):
Euzet L., Suriano D.M. 1977. Ligophorus n. g. (Monogenea, Ancyrocephalidae) parasite des Mugilidae
(Téléostéens) en Méditerranée. Bulletin du Muséum National d’Histoire Naturelle, Série 3. Zoologie 472:
799-821.
Hanek G., Fernando C.H. 1978. Spatial distribution of gill parasites of Lepomis gibbosus (L.) and Ambloplites
Gutiérrez P.A., Martorelli S.R. 1994. Seasonality, distribution, and preference sites of Demidospermus valenciennesi
(Monogenea: Ancyrocephalidae). Research and Reviews in Parasitologv 54 (4): 259-261
Gutiérrez P.A., Martorelli S.R. 1999. Hemibranch preference by freshwater monogeneans a function of gill area,
Koskivaara M., Valtonen E.T., Vuori K.M. 1992. Microhabitat distribution and coexistence
of Dactylogyrus (Monogenea) on the gills of roach. Parasitology
104
(2): 273-281.
Kumar R., Madhavi R., Sailaja B. 2016. Spatial distribution of ectoparasites on the gills of the
mullet, Liza macrolepis: the effects of pollution. Journal of Parasitic Diseases 41 (1): 40-47.
Mouillot D., Simkova A., Morand S., Poulin R. 2005. Parasite species coexistence and limiting similarity:
a multiscale look at phylogenetic, functional and reproductive distances. Oecologia 146: 269-278.
Paling J.E. 1968. A method of estimating the relative volumes of water flowing over the different gills a freshwater
fish. Journal of Experimental Biology 48 (3): P. 533-544.
Paling J.E. 1969. The manner of infection of trout gill by the monogenean parasites Discocotyle sagittata. Journal
Rubio-Godoy M. 2008. Microhabitat selection of Discocotyle sagittata (Monogenea: Polyopisthocotylea) in farmed
Šimková A., Morand S. 2008. Co-evolutionary patterns in congeneric monogeneans: a review of
Dactylogyrus species and their cyprinid hosts. Journal of Fish Biology 73: 2210-2227.
Wootten R. 1974. The spatial distribution of Dactylogyrus arnphibothrinrn on the gills of ruife Gyinnocephalus
cernna and its relation to the relative amounts of water passing over the parts of the gills. Journal of
519
THE DISTRIBUTION OF MONOGENEANS OF THE GENUS LIGOPHORUS
ON GILLS OF THE SOIUY MULLET PLANILIZA HAEMATOCHEILUS
N. V. Pronkina, E. V. Dmitrieva
Keywords: Monogenea, Ligophorus, congeners, distribution on gills, infrapopulation
number, fish size
SUMMARY
The distribution on the host gills of three monogenean species of Ligophorus Euzet et Suriano,
1977, parasitising soiuy mullet Planiliza haematocheilus (Temminck & Schlegel, 1845) in the Black
and Azov seas was studied. Specimens of L. pilengas and L. llewellyni prefer to localize on the same
gill sites, namely, I-III gill arches and their medial sectors. The third species, L. kaohsianghsieni,
was distributed on the gills more crowdedly, with about 80% of its specimens being present on
II gill arch, occurring on the medial and ventral sectors approximately equally. In general, most
monogeneans were found on the largest in area and well-watered gill arches and sectors, as well as
the anterior hemibranchies, however within these areas they preferred to attach to proximal half of
filaments with less amount of water current. The presence of congeners on gills had no significant
effect on the distribution of L. pilengas and L. llewellyni. Increasing infrapopulation number resulted
in a more uniform distribution of monogeneans on gills; their proportion upsurges significantly on
the faster growing anterior hemibranchs as fish size increased.
520