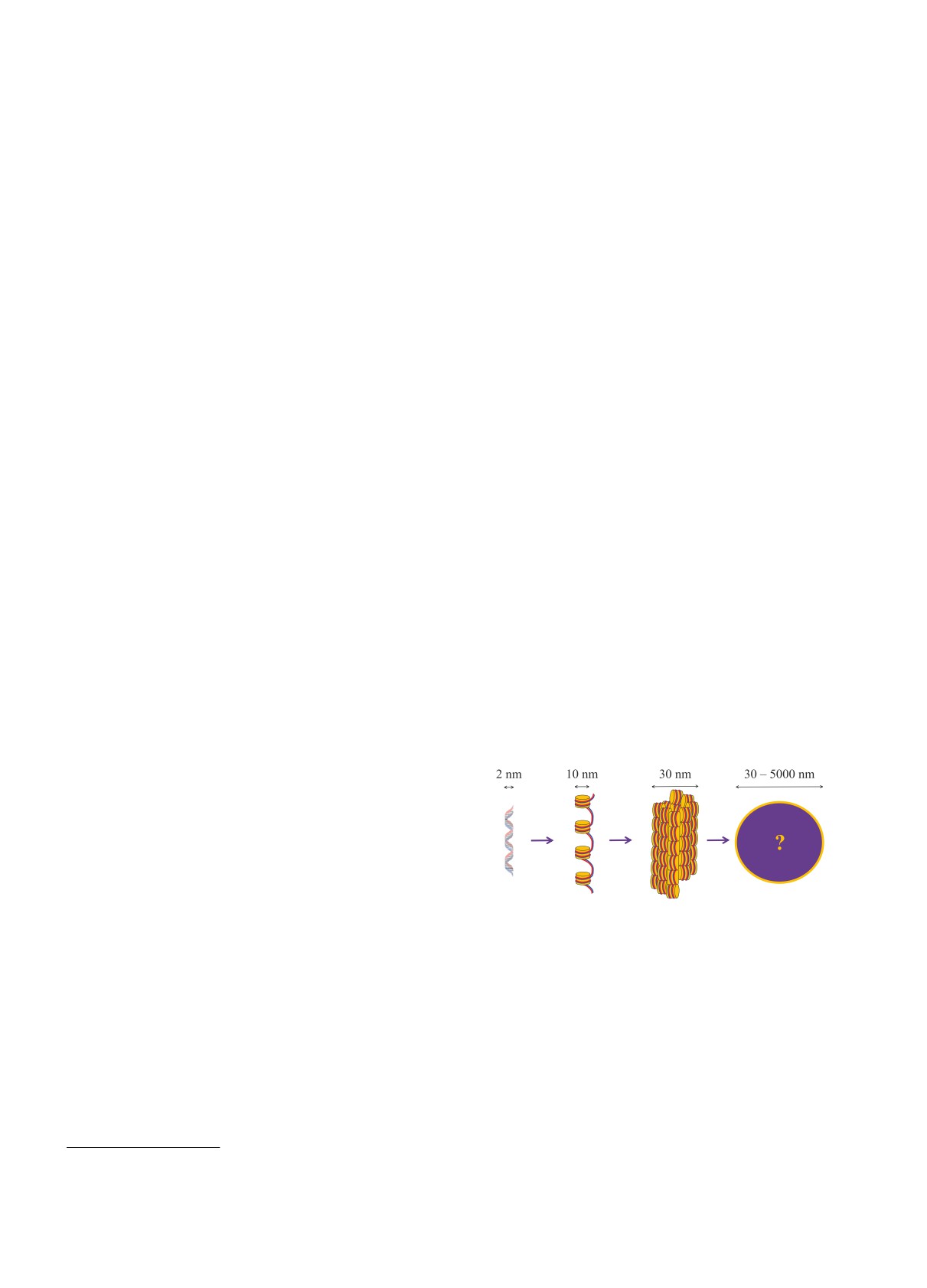Письма в ЖЭТФ, том 118, вып. 10, с. 776 - 781
© 2023 г. 25 ноября
Пространственная модель организации хроматина в ядре
биологической клетки по данным малоуглового рассеяния
Е. Г. Яшина+∗1), Е. Ю. Варфоломеева+, Р. А. Пантина+, В. Ю. Байрамуков+, Р. А. Ковалев+,
Н.Д.Федорова+, К.А.Пшеничный+, Ю.Е.Горшкова×, С.В.Григорьев+∗
+Петербургский институт ядерной физики,
Национальный исследовательский центр “Курчатовский институт”, 188300 Гатчина, Россия
∗Санкт-Петербургский государственный университет, 198504 С.-Петербург, Россия
×Объединенный институт ядерных исследований, 141980 Дубна, Россия
Поступила в редакцию 15 мая 2023 г.
После переработки 17 октября 2023 г.
Принята к публикации 17 октября 2023 г.
Представлены результаты малоуглового рассеяния нейтронов и рентгеновского излучения на ядрах
HeLa с нормальной и подавленной транскрипционной активностью. Данные малоуглового рассеяния ней-
тронов демонстрируют наличие бифрактальной структуры внутри ядра. На масштабах от размера ядра
до нескольких сотен нанометров наблюдается структура логарифмического фрактала, а на меньших
масштабах, вплоть до нуклеосомной организации - структура объемного фрактала. Данные малоуг-
лового рассеяния рентгеновского излучения показывают, что наличие структуры объемного фрактала
коррелирует с транскрипционной активностью клетки. Принимая во внимание успех описания хрома-
тина моделью складчатой глобулы (результаты данных метода Hi-C), мы предлагаем сценарий фор-
мирования бифрактальной структуры внутри ядра. Внутри плотнозапакованного закрытого хроматина
находится система транспортных каналов (логарифмический фрактал), а активный хроматин локализу-
ется вблизи или внутри транспортных каналов и, в результате транскрипционной активности, образует
объемнофрактальные структуры. Таким образом, структура логарифмического фрактала обеспечивает
равномерное рассредоточение пустот на разных масштабах, потенциально необходимое для осуществле-
ния транскрипции и транспорта веществ внутри ядра, в то время как структура объемного фрактала
является следствием транскрипционной активности клетки.
DOI: 10.31857/S1234567823220123, EDN: pftlyr
Вопрос о том, каким образом невероятно длин-
ная макромолекула ДНК уложена в относительно
компактном ядре биологической клетки является од-
ной из фундаментальных проблем биологии. На сего-
дняшний момент хорошо известны первые два уров-
ня компактизации хроматина. Первый из них - нук-
леосомный - на этом уровне двуспиральная нить
ДНК, толщиной 2 нм, а длиной несколько метров,
Рис. 1. (Цветной онлайн) Первые два этапа организа-
наматывается на 8 гистоновых белков, образуя 10 нм
ции хроматина: молекула ДНК наматывается на ги-
нуклеосому. Этот уровень компактизации иногда на-
стоновые белки, образуя нуклеосомы, которые форми-
зывают “бусины на нитке”, где бусины - это нук-
руют 30 нм фибриллу
леосомы, а нитка - это ДНК. Благодаря первому,
нуклеосомному, уровню упаковки, линейные разме-
дований показывают, что в клетке в основном на-
ры ДНК компактизуются в 6-7 раз. Считается, что
блюдаются неравномерно свернутые 10 нм, а не 30 нм
второй уровень организации хроматина - 30 нм фиб-
фибриллы [1-3].
рилла: нуклеосомы плотно прилегая друг к другу,
Таким образом, на первых двух уровнях орга-
образуют спираль диаметром 30 нанометров (рис.1).
низации хроматина ДНК компактизована в десят-
Однако, ряд структурных и визуализирующих иссле-
ки раз, и вопрос укладки ДНК сводится к вопросу
укладки хроматиновой фибриллы, являющейся ком-
1)e-mail: yashina_91@inbox.ru
плексом ДНК и связанных с ней гистоновых бел-
776
Письма в ЖЭТФ том 118 вып. 9 - 10
2023
Пространственная модель организации хроматина в ядре биологической клетки. . .
777
ков. Хотя длина фибриллы получается меньше дли-
та, воспроизводящего его форму. Следовательно,
ны ДНК в десятки раз, т.е. порядка сантиметра, она
малоугловое рассеяние на складчатой глобуле долж-
все же достаточно велика по сравнению с диамет-
но представлять классический случай рассеяния
ром ядра, который составяет всего несколько мик-
на нефрактальных трехмерных неоднородностях
рометров. Для того, чтобы представить себе пробле-
[12, 13]. В действительности эксперименты по ма-
му укладки хроматиновой фибриллы, полезно увели-
лоугловому рассеянию нейтронов на хроматине
чить все масштабы в миллион раз, тогда ситуация бу-
уверенно демонстрируют наличие бифрактальной
дет аналогична необходимости упаковать около ста
структуры на всех масштабах от нанометров до
километров обычной веревки сантиметровой толщи-
микрометров [13-19], что не укладывается в модель
ны в грузовик и распаковать веревку из грузовика,
складчатой глобулы и требует ее доработки. Для
не запутав ее [4]. Кроме того, физическая топология
описания экспериментальных данных малоуглового
хроматина и экспрессия генов тесно связаны: измене-
рассеяния нейтронов мы предположили, что струк-
ние фрактальной размерности структуры хромати-
тура хроматина в компартментах А и В различна, и
на на нанометровом масштабе влияет на активацию
если компартмент B описывается кривой полностью
транскрипционных генов [5]. Таким образом, фрак-
заполняющей пространство (плотный, неактивный,
тальная организация хроматина играет важную роль
закрытый хроматин), то компартмент А представ-
в экспрессии генов и эпигенетической репрессии.
ляет собой фрактальную систему разномасштабных
На сегодняшний день наиболее популярным ме-
полостей, связанных сетью транспортной инфра-
тодом исследования генома является метод захвата
структутуры, в которой функционирует открытый
конформации хромосом Hi-C [4, 6, 7]. С помощью
активный хроматин. На малых масштабах систе-
этого метода было показано, что хроматин делит-
ма характеризуется объемным фракталом, а на
ся на два полногеномных компартмента: “открытый”
больших - логарифмическим фракталом.
хроматин образует компартмент А, “закрытый” хро-
Эксперименты по МУРН (малоугловое рассеяние
матин - компартмент В. Компартмент В плотно упа-
нейтронов), Ультра МУРН и Спин-Эхо МУРН на
кован, а взаимодействующие гены проявляют боль-
разных типах ядер: куриных эритроцитов [13-16],
шую тенденцию к близкой пространственной лока-
HeLa [13, 17, 18] и крысиных лимфоцитов [19], по-
лизации, иными словами, гены чаще всего взаимо-
казали, что наднуклеосомная организация хромати-
действуют с соседними генами. В то время как ком-
на представляет собой двухуровневую фрактальную
партмент А упакован менее плотно, а находящиеся
структуру. Во всех исследованных ядрах организа-
в нем гены могут взаимодействовать друг с другом
ция хроматина на масштабах от размера ядра до
вне зависимости от их близости на нуклеотидной по-
нескольких сотен нанометров имеет структуру ло-
следовательности ДНК. Кроме того, в отличие от
гарифмического фрактала, а на меньших масшта-
компартмента В, в котором упакованы редко счи-
бах, вплоть до нуклеосомной организации (∼ 10 нм) -
тываемые гены, в компартменте А находятся актив-
структуру объемного фрактала.
но транскрибируемые гены, для которых необходима
В отличие от крысиных лимфоцитов и куриных
большая доступность для осуществления экспрессии
эритроцитов, клеточная линия HeLa обладает вы-
генов и сборок молекулярных машин [6].
сокой транскрипционной активностью. На рисунке 2
Наиболее популярной моделью для описания
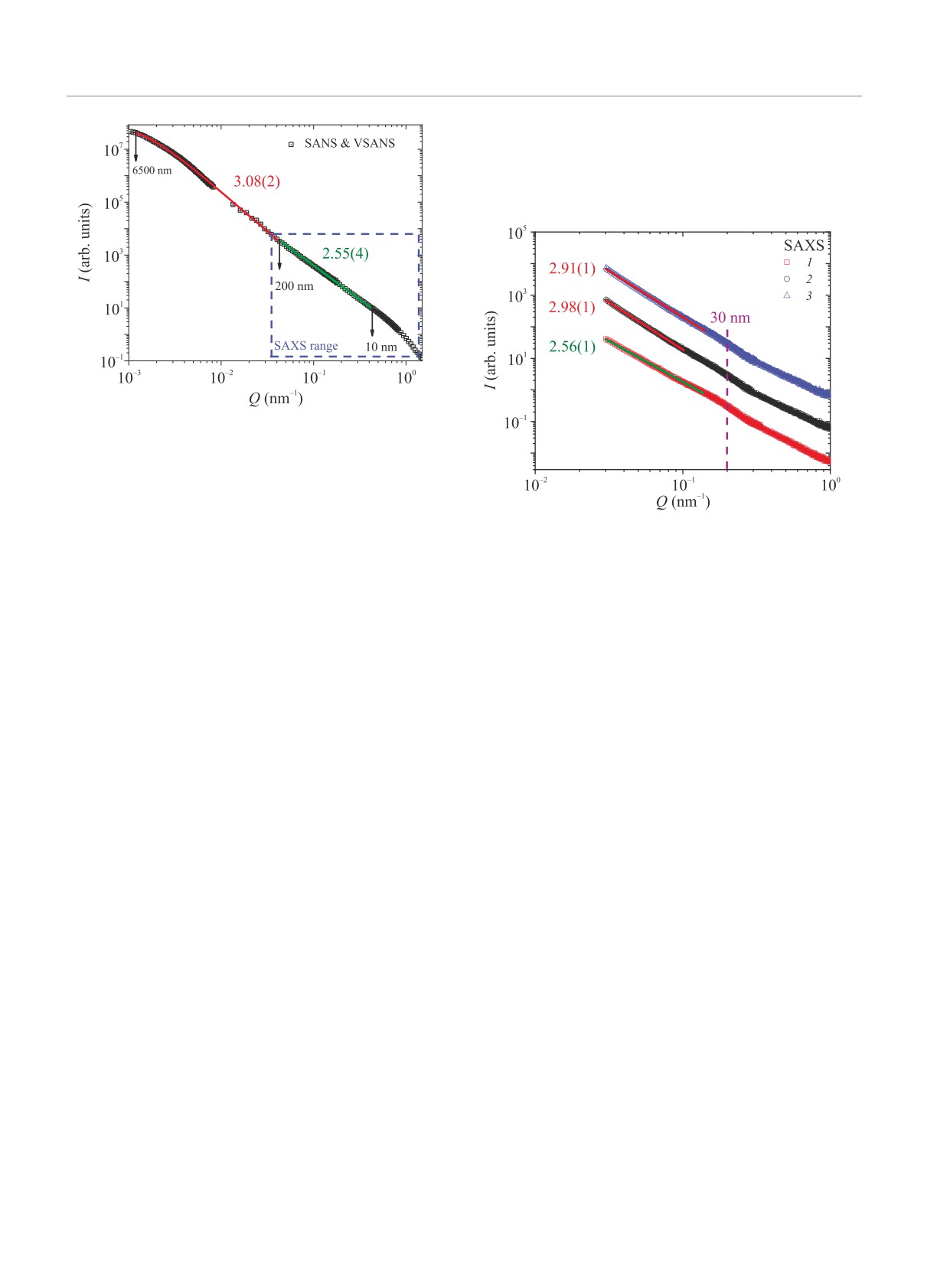
представлены результаты экспериментов по малоуг-
крупномасштабной упаковки ДНК является модель
ловому и ультрамалоугловому рассеянию нейтронов,
складчатой (фрактальной) глобулы, предложенная
полученных на ядрах HeLa, проведенных на установ-
Гросбергом в конце ХХ в. [8, 9]. Согласно этой моде-
ках KWS-2 и KWS-3 (MLZ, Германия Гархинг) в диа-
ли нить ДНК, подобно кривой Гильберта (Пеано),
пазоне переданных импульсов [10-3 -1] нм-1. Интен-
полностью заполняет трехмерное пространство без
сивность рассеяния нейтронов в диапазоне передан-
самопересечений, а ее размерность равна DF
= 3
ных импульсов [10-3 -4·10-2] нм-1 описывается сте-
[4, 6-11]. Модель складчатой глобулы является
пенной функцией с учетом конечного размера рассе-
хорошим приближением для описания
“закрыто-
ивателя I(Q) ∼ (1+(Qξ)2)-ν/2 со степенью ν = 3.08±
го” хроматина (компартмента В), массовая доля
± 0.002 и корреляционной длиной ξ = 6500 ± 100 нм.
которого превалирует в ядре. Однако, эта модель
Интенсивность малоуглового рассеяния нейтронов,
не описывает структуру
“открытого” хроматина
соответствующая хроматину в диапазоне передан-
(компартмента А) [6]. C точки зрения малоугло-
ных импульсов [4 · 10-2 - 7 · 10-1] нм-1, описыва-
вого рассеяния складчатая глобула неотличима
ется степенной функцией I(Q) ∼ Q-ν со степенью
от любого другого однородно заполненного объек-
ν = 2.55 ± 0.01. На кривой рассеяния наблюдается
Письма в ЖЭТФ том 118 вып. 9 - 10
2023
778
Е. Г. Яшина, Е. Ю. Варфоломеева, Р. А. Пантина и др.
Измеренная прежде кривая малоуглового рассея-
ния нейтронов на контрольном образце в тяжелой во-
де (рис. 2) совпадает с кривой рассеяния рентгенов-
ского излучения (рис. 3) в диапазоне переданных им-
Рис. 2. (Цветной онлайн) Интенсивность малоуглового
рассеяния нейтронов на ядрах HeLa, помещенных в тя-
желую воду, измеренная на установках KWS-2 и KWS-
3 (MLZ, Германия, Гархинг). Выделенная пунктиром
область соответствует диапазону переданных импуль-
сов, доступному в эксперименте по малоугловому рас-
Рис. 3. (Цветной онлайн) 1 - (красные открытые квад-
сеянию рентгеновского излучения
раты) - Интенсивность малоуглового рассеяния рент-
геновского излучения для референсных ядер HeLa, по-
мещенных в тяжелую воду. 2 - (черные открытые кру-
четкая точка перегиба между двумя фрактальными
ги) - Интенсивность малоуглового рассеяния рентге-
уровнями, соответствующая Q = 4 · 10-2 нм-1. Мел-
новского излучения для ядер HeLa, культивированных
комасштабный фрактальный уровень организации
в условиях дефицита питательной среды, помещенных
хроматина описывается моделью объемного фрак-
в тяжелую воду. 3 (cиние открытые треугольники) -
тала, в то время как крупномасштабный фракталь-
Интенсивность малоуглового рассеяния рентгеновско-
ный уровень описывается моделью логарифмическо-
го излучения для референсных ядер HeLa, культиви-
го фрактала.
рованных с добавления ингибитора транскрипции Ак-
Для того, чтобы определить связь транскрип-
тиномицином Д, помещенных в тяжелую воду
ционной активности клетки с изменением структу-
ры хроматина были исследованы три образца ядер
пульсов [3·10-2-1] нм-1. В этой области переданных
HeLa на установке малоуглового рассеяния рент-
импульсов наблюдается структура объемного фрак-
геновского (МУРР) излучения XEUSS 3.0 (ОИЯИ,
тала ν = 2.56 ± 0.01. Точка перегиба и перехода в
Россия, Дубна). Контрольный образец ядер, на кото-
крупномасштабный фрактальный уровень из рентге-
ром проводились эксперименты по МУРН, был полу-
новских данных невозможно определить, поскольку
чен при выделении ядер из клеток, культивирован-
она находится на границе разрешающей способности
ных в нормальных условиях, которые проявляли ти-
прибора. Однако, из нейтронных данных она опреде-
пичную для этих клеток транскрипционную актив-
лена и равна Qc = 0.04 нм-1. С другой стороны, кри-
ность. Второй образец был получен при выделении
вые рассеяния на образцах с подавленной транскрип-
ядер из клеток, культивированных в условиях дефи-
ционной активностью демонстрируют закон рассея-
цита питательной среды, за счет чего транскрипци-
ния близкий к кубическому, что соответствует струк-
онная активность клеток была занижена из-за от-
туре логарифмического фрактала в трехмерном про-
сутствия “строительного материала” для осуществ-
странстве. Как показано на рис.3, для образца ядер
ления транскрипции. Третий образец ядер был полу-
клеток, культивированных в условиях дефицита пи-
чен при выделении ядер из клеток, культивирован-
тательной среды наблюдается степенной закон рас-
ных с добавлением ингибитора транскрипции Акти-
сеяния с показателем ν = 2.98 ± 0.01 в диапазоне
номицина Д, за счет чего транскрипционная актив-
переданных импульсов [3·10-2-10-1] нм-1, а для об-
ность клеток была подавлена. Получение всех трех
разца ядер клеток, культивированных с добавлением
образцов проводили по одной схеме, подробно опи-
ингибитора транскрипции Актиномицина Д (рис. 3)
санной в работах [17, 18].
наблюдается степенной закон рассеяния с показате-
Письма в ЖЭТФ том 118 вып. 9 - 10
2023
Пространственная модель организации хроматина в ядре биологической клетки. . .
779
лем ν = 2.91 ± 0.01 в диапазоне переданных импуль-
сов [3· 10-2 -1.5· 10-1] нм-1. Таким образом, можно
сделать вывод о том, что наличие структуры объ-
емного фрактала на масштабе [3 · 10-2 - 10-1] нм-1
связано с транскрипционной активностью клетки, а
структура логарифмического фрактала связана с ее
отсутствием.
В классификации малоуглового рассеяния на
фрактальных объектах [12] модель логарифмическо-
го фрактала описывает иерархическую структуру,
которая характеризуется логарифмической мерой и
Рис. 5. (Цветной онлайн) Построение первых 3 поко-
формируется согласно принципу сохранения объема
лений фрактала Снежинка Коха: (а) - геометрически
при изменении масштаба. Экспериментально струк-
регулярного фрактала Снежинка Коха; (b) - стохасти-
тура логарифмического фрактала наблюдается как
ческого фрактала Снежинка Коха со случайной ори-
кубический закон в интенсивности малоуглового рас-
ентацией сторон треугольника, на которые заменяется
сеяния в зависимости от модуля переданного импуль-
средний сегмент отрезка
са. На рисунке 4 показан принцип построения этого
разделяется на три равные части и средний сегмент
заменяется двумя сторонами равностороннего тре-
угольника длины 1/3. В результате образуется ло-
маная, состоящая из четырех звеньев длины 1/3.
На следующем шаге повторяем операцию для каж-
дого из четырех получившихся звеньев (рис. 5a) и
т.д
Если в процесс замены среднего сегмента на
две стороны треугольника добавить фактор случай-
Рис. 4. (Цветной онлайн) Построение первых
4
по-
ности, то кривая Коха приобретет менее регулярный
колений логарифмического фрактала, основанного на
стохастический вид, однако, сохранит свои фрак-
принципе сохранения объема при масштабировании
тальные свойства [22] (рис. 5b).
фрактала до четвертой итерации: к вершинам куба
Гипотеза о механизме формирования бифрак-
объемом V добавляются 8 одинаковых кубов помень-
тальной структуры активного хроматина заключает-
ше объемом V1, а сумма их объемов равна объему
ся в следующем: структура логарифмического фрак-
исходного куба
∑V1 = V ; на следуюущей итерации
тала образуется из пустот, которые формируют неза-
к вершинам каждого из добавленных в предыдущей
полненное хроматином пространство, потенциаль-
итерации куба добавляются еще по 8 кубов поменьше
но необходимое для транспорта веществ внутри яд-
объемом V2, а сумма их объемов равна сумме объе-
ра и транскрипционной активности клетки. Далее,
мов кубов, добавленных на предыдущей итерации, а
когда начинается транскрипция, ДНК выпетлива-
также объему исходного куба
∑V2 = ∑V1 = V и т.д.
ется в пустоты логарифмического фрактала или
В ядрах некоторых клеток число итераций доходит
вблизи его границ, образуя объемнофрактальную
до 4 или 5.
структуру вследствие суперскрученности ДНК, что
В отличие от модели логарифмического фракта-
в свою очередь облегчает диффузию веществ и, как
ла, модель объемного фрактала соответствует од-
следствие, сборку молекулярных машин и белково-
нородной самоподобной классической фрактальной
нуклеиновых комплексов. Таким образом, структу-
структуре, характеризующейся хаусдорфовой раз-
ра логарифмического фрактала обеспечивает равно-
мерностью DF [20, 21] и наблюдается в эксперименте,
мерное рассредоточение пустот на разных масшта-
как убывающая степенная зависимость интенсивно-
бах, потенциально необходимое для осуществлении
сти малоуглового рассеяния от вектора рассеяния с
транскрипции и транспорта веществ внутри ядра, в
показателем 2 < ν < 3 (ν = DF ).
то время как структура объемного фрактала явля-
В качестве модели для описания объемнофрак-
ется следствием транскрипционной активности клет-
тальной структуры, наблюдаемой в малоугловом
ки. Сосуществование в ядре вышеописаных структур
рассеянии, можно выбрать кривую Коха (рис. 5a).
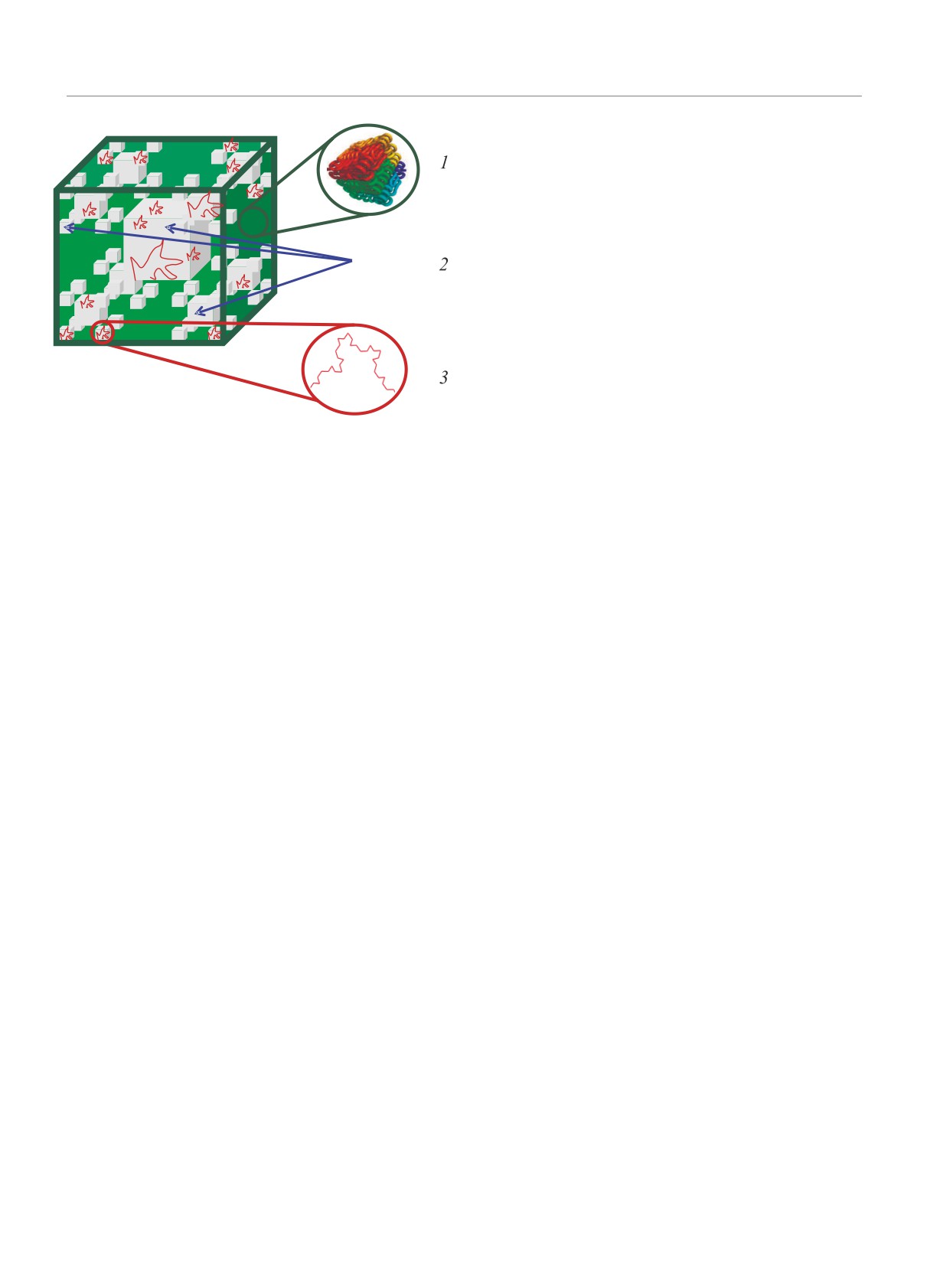
схематично изображено на рис. 6.
Процесс построения классической кривой Коха за-
Построенная таким образом бифрактальная кон-
ключается следующем: берется единичный отрезок,
струкция является еще одним шагом на пути созда-
Письма в ЖЭТФ том 118 вып. 9 - 10
2023
780
Е. Г. Яшина, Е. Ю. Варфоломеева, Р. А. Пантина и др.
Таким образом, наблюдаемая в экспериментах по
малоугловому рассеянию нейтронов и рентгеновско-
го излучения, структура логарифмического фрак-
тала обеспечивает равномерное по всему простран-
ству ядра рассредоточение полостей на разных мас-
штабах, потенциально необходимых для обеспечения
быстрого и эффективного транспорта и возможно-
сти осуществления биологической функции в любом
месте ядра. Как показано выше, транскрипционная
активность хроматина скоррелирована и, возможно,
определяет появление, размер и размерность объем-
ного фрактала.
В заключение, стоит отметить, что структура ло-
гарифмического фрактала представляет собой неза-
полненную хроматином систему полостей, необходи-
Рис. 6. (Цветной онлайн) Пространственная модель ор-
мую для метаболизма и инфраструктуры внутри яд-
ганизации хроматина, согласно которой внутри плот-
ра, а также формирует собой пространство, потен-
нозапакованного транскрипционно-неактивного
(“за-
циально необходимое для осуществления биологиче-
крытого”) хроматина - 1, упакованного по принци-
ских функций ДНК, таких как транскрипция. Пред-
пу трехмерной кривой Пеано, находится система по-
положительно, такая транспортная система каналов
лостей - 2, формирующая структуру транспортных
заполена нуклеоплазмой и сформирована гидроди-
каналов по принципу логарифмического фрактала, а
транскрипционно-активный (“открытый”) хроматин -
намическими силами, возникающими из-за разности
3
образует объемнофрактальные структуры внутри
токов на границе раздела фаз.
транспортных каналов или вблизи их границ
Далее, когда и если начинается транскрипцион-
ная активность, ДНК выпетливается в пустоты ло-
ния модели организации хроматина в ядре биоло-
гарифмического фрактала и там начинается сборка
гической клетки. Отметим, что структура логариф-
молекулярных машин и белково-нуклеиновых ком-
мического фрактала является универсальной для
плексов, и тем самым, в пустотах логарифмическо-
большинства типов ядер и образуется из системы по-
го фрактала активный хроматин образует струк-
лостей, которые формируют систему транспортных
туру объемного фрактала, в котором происходит
каналов, необходимую для метаболизма и инфра-
облегченная диффузия для осуществления тран-
структуры внутри ядра, а также формирует собой
скрипции. Предположительно, возникновение объ-
пространство, потенциально необходимое для осу-
емнофрактальных структур является следствием ме-
ществления биологических функций ДНК, таких как
ханизмов суперскрученности ДНК, которая появля-
транскрипция. Эта система полостей-каналов лога-
ется в следствии транскрипции [28]. Транскрипци-
рифмического фрактала находится внутри плотно-
онно активный хроматин демонстрирует большую
запакованной ДНК по принципу фрактальной гло-
физическую жесткость, по сравнению с неактивным
булы и заполнена активным хроматином. Необходи-
хроматином, являясь при этом менее плотным [29].
мость существования в ядре системы каналов была
Также, принимая во внимание, что хроматин не яв-
высказана в работе [23]. Предполагалось, что ядер-
ляется статичным, т. е. в нем постоянно происхо-
ный матрикс представляет собой систему каналов,
дит та или иная динамика, логично предположить,
по которым осуществляется транспорт мРНК из яд-
что наиболее часто считываемые гены локализуют-
ра в цитоплазму и транспорт различных цитоплаз-
ся либо внутри полостей-каналов логарифмического
матических продуктов к местам их утилизации в яд-
фрактала, либо вблизи его границы в запакованном
ре. Наличие транспортных каналов в хроматине для
состоянии внутри фрактальной глобулы. Структура
облегченной диффузии РНК, белков и других кле-
каналов-полостей логарифмического фрактала явля-
точных компонентов была теоретически обоснована
ется местом локализации активного хроматина. В ра-
в работе [24]. В работах [25, 26, 27] были получены
боте [13, 17] было показано, что структура логариф-
свидетельства того, что новосинтезированная РНК
мического фрактала обладает очень разветвленной
не просто диффундирует от места ее синтеза во всех
поверхностью и, как следствие, большой площадью
возможных направлениях, но следует по определен-
поверхности. В связи с этим, в плотнозапакованном
ным путям (каналам).
хроматине вблизи поверхности полостей-каналов мо-
Письма в ЖЭТФ том 118 вып. 9 - 10
2023
Пространственная модель организации хроматина в ядре биологической клетки. . .
781
жет локализоваться большое количество генов, кото-
12.
Е. Г. Яшина, С. В. Григорьев, Поверхность. Рентге-
рые способны быстро распаковаться в пространство
новские, синхротронные и нейтронные исследования
полости логарифмического фрактала. Таким обра-
9, 5 (2017).
зом, структура логарифмического фрактала обеспе-
13.
Е. Г. Яшина, С. В. Григорьев, ЖЭТФ 156(3), 540
чивает равномерное рассредоточение пустот-каналов
(2019).
на разных масштабах, потенциально необходимых
14.
D. V. Lebedev, M. V. Filatov, A. I. Kuklin,
для обеспечения транспортной инфраструктуры яд-
A. K. Islamov, E. Kentzinger, R. Pantina,
ра и пространства для биологической активности
B. P. Toperverg, and V. V. Isaev-Ivanov, FEBS
Lett. 579(6), 1465 (2005).
ДНК, в то время как структура объемного фрактала
15.
E. G. Iashina, E. V. Velichko, M. V. Filatov,
является следствием транскрипционной активности
W. G. Bouwman, C. P. Duif, A. Brulet, and
клетки.
S. V. Grigoriev, Phys. Rev. E 96(1), 012411 (2017).
Работа выполнена при поддержке Российского
16.
S. V. Grigoriev, E. G. Iashina, V.Yu. Bairamukov,
научного фонда (грант # 20-12-00188).
V. Pipich, A. Radulescu, M. V. Filatov, R. A. Pantina,
Авторы выражают благодарность нейтронному
and E. Yu. Varfolomeeva, Phys. Rev. E 102, 032415
центру MLZ за предоставление пучкового време-
(2020).
ни на установках KWS-2 и KWS-3, и А. Радулеску
17.
E. G. Iashina, M. V. Filatov, R. A. Pantina,
(A. Radulescu) и В. Пипича (V. Pipich) за помощь в
E. Yu. Varfolomeeva, W. G. Bouwman, Ch. P. Duif,
проведении эксперимента. Также авторы благодар-
D. Honecker, V. Pipich, and S.V. Grigoriev, J. Appl.
ны Объединенному институту ядерных исследова-
Cryst. 52, 844 (2019).
ний за предоставление пучкового времени на мало-
18.
S. V. Grigoriev, E. G. Iashina, B. Wu, V. Pipich,
угловой установке рентгеновского излучения XEUSS
Ch. Lang, A. Radulescu, V. Yu. Bairamukov,
3.0, и Ю. Горшковой и Г. Копице за помощь в прове-
M. V. Filatov, R. A. Pantina, and E. Yu. Varfolomeeva,
дении эксперимента.
Phys. Rev. E 104, 044404 (2021).
19.
E. G. Iashina, E. Yu. Varfolomeeva, R. A. Pantina,
V. Yu. Bairamukov, R.A. Kovalev, N. D. Fedorova,
1. Ya. Joti, T. Hikima, Y. Nishino, F. Kamada, S. Hihara,
V. Pipich, A. Radulescu, and S.V. Grigoriev, Phys. Rev.
H. Takata, T. Ishikawa, and K. Maeshima, Nucleus 3,
E 104, 064409 (2021).
404 (2012).
20.
B. Mandelbrot, The Fractal Geometry of Nature,
2. Y. Takizawa and H. Kurumizaka, Biochimica et
Freeman, N.Y. (1983).
Biophysica Acta (BBA)-Gene Regulatory Mechanisms
21.
J. Feder, Fractals, Plenum, N.Y. (1998).
1865, 194851 (2022).
22.
A. V. Ilatovskiy, D.V. Lebedev, M. V. Filatov,
3. K. Maeshima, S. Ide, and M. Babokhov, Curr. Opin.
M. G. Petukhov, and V. V. Isaev-Ivanov, J. Phys.:
Cell Biol. 58, 95 (2019).
Conf. Ser. 351(1), 012007 (2012).
4. J. D. Halverson, J. Smrek, K. Kremer, and
23.
S. V. Razin and I. I. Gromova, BioEssays 17(5), 443
A.Y. Grosberg, Rep. Prog. Phys.
77(2),
022601
(1995).
(2014).
24.
T. Cremer, M. Cremer, B. Hubner, A. Silahtaroglu,
5. L. M. Almassalha, A. Tiwari, P. T. Ruhoff, Y. Stypula-
M. Hendzel, C. Lanctôt, and H. Strickfaden, BioEssays
Cyrus, L. Cherkezyan, H. Matsuda, M. A. Dela Cruz,
42(2), 1900132 (2020).
J. E. Chandler, C. White, C. Maneval, H. Subramanian,
25.
Y. Xing, C. V. Johnson, P. R. Dobner, and
I. Szleifer, H.K. Roy, and V. Backman, Sci. Rep. 7,
J. B. Lawrence, Science 259(5099), 1326 (1993).
41061 (2017).
26.
M. Mazzocca, T. Fillot, A. Loffreda, D. Gnani, and
6. E. Lieberman-Aiden, N. L. Van Berkum, L. Williams
D. Mazza, Biochem. Soc. Trans. 49(3), 1121 (2021).
et al. (Collaboration), Science 326(5950), 289 (2009).
27.
M. Mazzocca, E. Colombo, A. Callegari, and D. Mazza,
7. L. A. Mirny, Chromosome research 19, 37 (2011).
Current Opinion in Structural Biology 71, 239 (2021).
8. A. Y. Grosberg, S. K. Nechaev, and E. I. Shakhnovich,
28.
P. Chatterjee, N. Goldenfeld, and S. Kim, Phys. Rev.
Journal de physique 49(12), 2095 (1988).
Lett. 127(2)1, 218101 (2021).
9. A. Grosberg, Y. Rabin, S. Havlin, and A. Neer,
29.
V. Yu. Bairamukov, M. V. Filatov, R.A. Kovalev,
Europhysics Letters 23(5), 373 (1993).
N. D. Fedorova, R. A. Pantina, A.V. Ankudinov,
E. G. Iashina, S. V. Grigoriev, and E. Yu. Varfolomeeva,
10. J. D. Halverson, W. B. Lee, G. S. Grest, A. Y. Grosberg,
Biochim. Biophys. Acta Gen. Subj. 1866(12), 130234
and K. Kremer, J. Chem. Phys. 134(20), 204904 (2011).
(2022).
11. M. V. Imakaev, K. M. Tchourine, S. K. Nechaev, and
L. A. Mirny, Soft matter 11(4), 665 (2015).
Письма в ЖЭТФ том 118 вып. 9 - 10
2023