Российская сельскохозяйственная наука, 2020, № 6
УДК.633.18:631.527.7:581.143.6(571.6)
DOI: 10.31857/S2500262720060034
ВНУТРИКАЛЛУСНАЯ И МЕЖКАЛЛУСНАЯ МОРФОЛОГИЧЕСКАЯ ИЗМЕНЧИВОСТЬ
УДВОЕННЫХ ГАПЛОИДОВ РИСА, ПОЛУЧЕННЫХ В АНДРОГЕНЕЗЕ in vitro
М.В. Илюшко, кандидат биологических наук, С.С. Гученко,
М.В. Ромашова, кандидат сельскохозяйственных наук
Федеральный научный центр агробиотехнологий Дальнего Востока имени А.К. Чайки,
692539, Приморский край, п. Тимирязевский, ул. Воложенина, 30
E-mail: ilyushkoiris@mail.ru
Изучена внутрикаллусная и межкаллусная морфологическая изменчивость удвоенных гаплоидов риса Oryza sativa
L., полученных в андрогенезе in vitro на каллусных линиях с множественной регенерацией. Использовали регенеран-
ты от гибридов второго поколения F2: Дон 4237×(Зарваси 70×Хейлуньдзян) - Д×З×Х, растение №8; Романика×(Дарий
122×Краснодар 9167) - Р×Д×67, растение №15; Китаец×(ВНИИР 3223×Кензо) - К×23×К, растения №26 и №28. Семенное
потомство удвоенных гаплоидов риса первого поколения R1 высевали на вегетационной площадке в сосудах размером
1,54 м2, наполненных полевой почвой. Всего проанализировано 1383 растения 144 линий удвоенных гаплоидов восьми кал-
лусных линий. В результате дисперсионнного анализа выявлены статистически значимые различия между удвоенными
гаплоидами двух каллусных агрегатов одного пыльника и разных пыльников одного гибрида. Обнаружена корреляцион-
ная зависимость (p<0,05) ряда биометрических показателей от высоты растений: длина метелки (r=0,72), масса зерна
главной метелки (r=0,80), масса зерна растения (r=0,59), масса 1000 зерен (r=0,74). Таким образом, высокорослые расте-
ния оказались болеe продуктивными, чем низко- и среднерослые. После первичной оценки линий удвоенных гаплоидов
по биометрическим показателям объединили семенное потомство наиболее продуктивных растений одной каллусной
линии с целью увеличения объема образца и его быстрого размножения. Это позволит скорее перейти от рассмотрения
продуктивности отдельных растений к полевой оценке урожайности.
INTRACALLUS AND INTERCALLUS MORPHOLOGICAL VARIABILITY
OF RICE DOUBLED HAPLOIDS, GAINED in vitro ANDROGENESIS
Ilyushko M.V., Guchenko S.S., Romashova M.V.
Federal Scientific Centre of Agrobiotechnology of the Far East named A.K. Chaika,
692539, Primorskiy kray, p. Timiryasevskiy, ul. Volozhenina, 30
Е-mail:ilyushkoiris@mail.ru
Intracalus and intercallus morphological variability of rice doubled haploids of Oryza sativa L., obtained in vitro androgenesis on
callus lines with multiple regeneration, was studied. Used regenerants gained from hybrids of the second generation F2, next hybrid
combinations: Don 4237×(Szarvasi 70×Heilunjiang) - D×S×X, plant No. 8; Romanika×(Darius 122×Krasnodar 9167) - P×D×67,
plant No. 15; Kitaets×(VNIIR 3223×Kenzo) - K×23×K, plants No. 26 and No. 28. The seed progeny of doubled haploids of the first
generation R1 were sown on the vegetation site in vessels of 1.54 m2 in size, filled with field soil. A total of 1383 plants were analyzed,
144 lines of doubled haploids of eight callus lines. The analysis of variance revealed statistically significant differences (p<0.05)
between doubled haploids of two callus aggregates of same anther and different anthers of one hybrid. A correlation dependence (p
<0.05) of biometric indicators on the plant height was found: panicle length (r = 0.72), grain mass of the main panicle (r = 0.80),
plant grain mass (r = 0.59), mass of 1000 grains (r = 0.74), i.e. tall plants were more productive than low and medium-sized plants.
After an initial assessment of doubled haploid lines by biometric indicators, the seed progeny of the most productive plants of the
same callus line were combined to increase the volume of the sample and their rapid reproduction. This will make it possible to move
more quickly from considering the productivity of individual plants to a field assessment of yield.
Ключевые слова: Oryza sativa L., удвоенные гаплоиды,
Key words: Oryza sativa L., doubled haploids, intracallus
внутрикаллусная
морфологическая
изменчивость,
morphological variability, plant breeding
селекционный процесс
Использование удвоенных гаплоидов (doubled-
[2]. Интерес к DH-технологиям наметился с внедрени-
haploids - DH) служит эффективной технологией в се-
ем маркер-ориентированной селекции (marker assistant
лекционном процессе растений [1-3], и рис - одна из
selection - MAS), позволившей получить линии удво-
первых культур, где она была успешно применена в
енных гаплоидов с целевыми признаками. Например, у
культуре пыльников in vitro [1, 4, 5]. Основные исследо-
риса - линии с генами устойчивости к грибному возбу-
вательские работы в этой области на любых сельскохо-
дителю пирикуляриоза Pyricularia grisea Sacc. [10, 11],
зяйственных культурах, как и прежде, сосредоточены в
резистентные к двум видам патогенов [12], глютиноз-
основном на оптимизации существующих методик по-
ные [13], у яровых зерновых - формы, устойчивые к
лучения удвоенных гаплоидов и связаны с изучением
грибным болезням [2].
условий произрастания растений-доноров, предобра-
DH-технология и MAS (отдельно и совместно) при-
боткой пыльников, оптимизацией питательных сред и
званы ускорить селекционный процесс [2, 14]. Однако
удвоением хромосом в клетках [1, 2, 5]. Кроме этого
можно отметить некоторые ограничения для каждой из
множество трудно культивируемых in vitro генотипов
них. Считается, что гомозиготные линии удвоенных га-
представляет большой интерес для селекционеров,
плоидов можно получить за 2-3 поколения вместо 6-9
что требует изучения факторов, детерминирующих
генераций как при привычном отборе гибридных расте-
андрогенетический успех [6-9]. Некоторые известные
ний [5, 14]. Но даже при большой отзывчивости группы
лаборатории организовали широкомасштабное произ-
генотипов (например, различные гибридные комбина-
водство удвоенных гаплоидов для селекционных целей
ции одинаковых родителей) у некоторых из них отсут-
11
Российская сельскохозяйственная наука, 2020, № 6
Средние значения морфологических признаков внутри каллусной линии,
рассчитанные по усредненным значениям линий удвоенных гаплоидов
Гибридное
Каллусная
Объем
Высота
Длина
Кущение,
Число
Масса
Масса
Масса
Соотно-
Проч-
растение
линия
выборки
растений,
метелки,
шт.
зерен
зерна
зерна
1000 шт.
шение
ность
см
см
главной
главной
расте-
зерен
зерно:со-
соломины
метелки,
метел-
ния, г
лома
шт.
ки, г
Д×З×Х(8)
88.2.1
16
87,5
18,1
3,1
85,8
2,4
5,7
27,9
1,28
0,18
97.1.1
7
85,1
18,9
2,7
89,1
2,4
5,3
28,6
1,56
0,16
125.1.1
6
74,8
16,6
3,0
93,6
2,5
5,4
25,9
1,35
0,18
р**
-
0,07
0,17
0,61
0,69
0,97
0,88
0,05
0,22
0,05
Р×Д×67(15)
54.2.1
15
77,0
14,6
1,1
60,3
1,5
3,4
24,9
1,21
0,14
55.1.1
18
80,2
15,5
0,8
67,5
1,6
4,5
24,6
1,04
0,15
62.2.2
19
90,1
15,6
0,8
65,3
1,6
4,6
24,5
0,99
0,16
62.2.4
14
82,3
15,7
1,4
61,2
1,5
4,5
23,8
0,95
0,16
р*
-
0,02
0,80
0,48
0,08
0,16
0,64
0,29
0,33
0,94
р**
-
0,000003
0,007
0,002
0,003
0,12
0,002
0,26
0,0002
0,004
К×В×К(26)
80.2.1
16
94,8
16,9
1,5
76,3
2,2
4,7
28,9
1,10
0,16
80.2.2
12
106,8
20,1
1,7
91,2
2,7
5,7
30,2
1,24
0,16
р*
-
0,00005
0,00002
0,55
0,004
0,006
0,03
0,03
0,29
0,95
К×В×К(28)
126.1.1
21
93,0
19,3
3,1
60,9
2,2
5,8
32,4
1,91
0,17
Примечание. р* - уровень достоверности различий между удвоенными гаплоидами двух каллусных агрегатов единой каллусной линии
по результатам дисперсионного анализа; р** - уровень достоверности различий между удвоенными гаплоидами разных каллусных линий
одного гибридного растения по результатам дисперсионного анализа.
ствует каллусообразование или регенерация [15, 16]. В
вительной селекции (один цикл - 7 лет), основанной
этом случае используют гибриды второго поколения,
на применении дигаплоидных линий, для сбора в гомо-
которые более отзывчивы на культуру пыльников in vitro
зиготной линии благоприятных аллелей, отвечающих
и обеспечивают больший выход зеленых регенерантов
за гетерозис [20]; пирамидирование генов гибриди-
[5, 17]. Для неподдающихся генотипов необходим под-
зацией с применением MAS [14, 21, 22]; закрепление
бор особых условий культивирования in vitro [6-9], что
гетерозиса гибридов комбинацией DH-технологии и
требует дополнительных временных и материальных
молекулярно-генетического сопровождения (6 лет) [4];
затрат. Ситуация усугубляется тем, что отдельные гены,
насыщение гермоплазмы лучших сортов целевыми ге-
ответственные за хозяйственно полезные признаки, не-
нами с молекулярно-генетическим контролем (10 лет)
гативно влияют на андрогенетический ответ растений
[23]. Эти методы работы, безусловно, результативны,
[18], а целевые гены MAS часто сцеплены с показателя-
но при этом еще больше возрастает срок создания но-
ми продуктивности растений и ведут к негативному от-
вых сортов.
бору [19], или отсутствует фенотипическое проявление
Интенсификация селекционного процесса необхо-
признаков, по которым вели молекулярно-генетический
дима не только на этапе создания исходного материала,
отбор [12]. После создания исходного материала необ-
но и на последующих этапах оценки и размножения.
ходимо оценить агрономически важные признаки полу-
Для быстрого, точного и массового описания растений
ченных линий удвоенных гаплоидов, их урожайность,
разработано автоматическое фенотипирование расте-
что занимает еще минимум шесть лет [2].
ний [14]. Ускорить размножение возможно формирова-
Потенциал увеличения урожайности за счет соз-
нием популяции растений с одинаковой комбинацией
дания новых сортов основных сельскохозяйственных
целевых генов (10-35 линий) [22], то есть проведением
культур традиционными методами в основном исчер-
массового отбора. Среди линий удвоенных гаплоидов
пан. Поэтому во всем мире стремятся вывести супер-
принято проводить индивидуальный отбор [4, 15, 24].
сорта и супергибриды, которые преодолели бы барьер
Считается, что для эффективного отбора достаточно
повышения продуктивности культур [4]. Для этого уже
100-150 линий одного образца [Zongxiu, Chengzhang,
недостаточно просто гибридизации, создания сома-
1992, по: 5]. В этом случае не обсуждается вопрос
клональных регенерантов, удвоенных гаплоидов или
принадлежности линий удвоенных гаплоидов одному
отобранных по одному признаку с помощью MAS се-
пыльнику или разным пыльникам или микроспорам.
лекционных линий. Современные селекционные схе-
Между тем на одной каллусной линии, полученной
мы усложняются различными комбинациями прежних
с одного пыльника, может образоваться более сотни
методик: предложены схема обогатительно-восстано-
удвоенных гаплоидов [25], многие из которых морфо-
12
Российская сельскохозяйственная наука, 2020, № 6
вали на вегетационной площадке в
сосудах размером 1,54 м2, напол-
ненных полевой почвой. Исполь-
зовали регенеранты, полученные
от гибридов второго поколения F2,
следующих гибридных комбина-
ций: Дон 4237×(Зарваси 70× Хей-
луньдзян) - Д×З×Х, растение №8
(три каллусных линии); Романи-
ка×(Дарий 122×Краснодар
9167)
– Р×Д×67, растение
№15 (три
каллусные линии четырех каллус-
ных агрегатов); Китаец×(ВНИИР
3223×Кензо) - К×23×К, растения
№26 (одна каллусная линия двух
каллусных агрегатов) и №28 (одна
каллусная линия). Под термином
«каллусная линия» понимали все
каллусные агрегаты, сформиро-
ванные на одном пыльнике [26].
Высеяли
144 линии удвоенных
гаплоидов, объем выборки каждой
каллусной линии представлен в
таблице.
Определяли биометрические
показатели основных элементов
продуктивности 7-10 растений с
одной линии удвоенных гаплои-
дов: высоту растения (см), дли-
ну метелки (см), кущение общее
(шт.), число зерен главной метелки
(шт.), массу зерна главной метелки
(г), массу зерна растения (г), массу
1000 зерен растения (г) и рассчи-
тывали по ним средние значения
для каждой линии, а также отно-
шение массы зерна 10 растений к
массе соломы 10 растений. Индекс
прочности соломы вычисляли по
формуле:
Ic = m c÷ t ÷ h,
где mc - масса соломы 10 растений
(г); t - среднее кущение 10 расте-
ний; h - средняя высота 10 расте-
ний.
По визуальной оценке, прове-
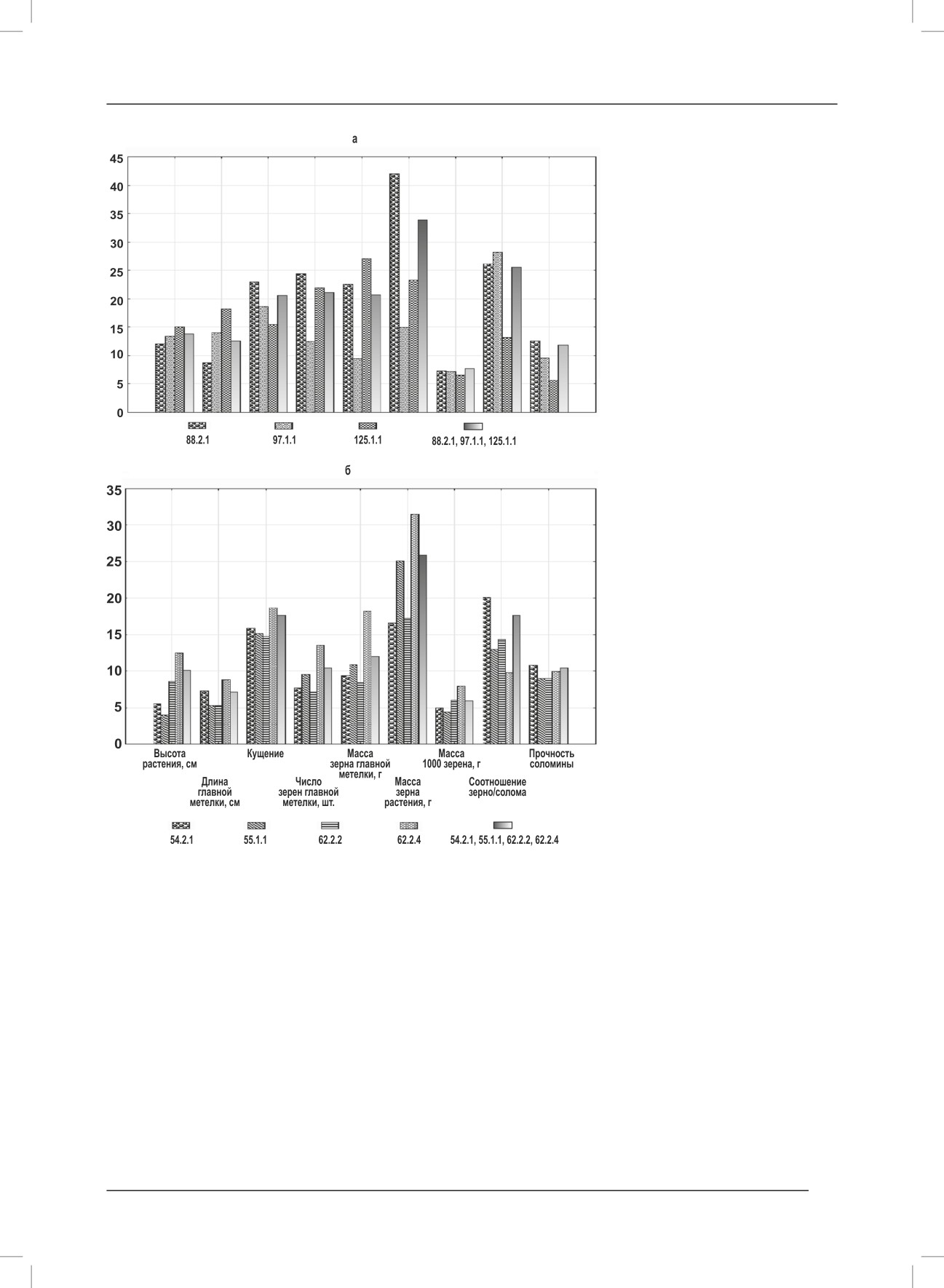
Внутрикаллусные и межкаллусные коэффициенты вариации (%)
денной на 100 растениях, значение
морфологических признаков удвоенных гаплоидов двух гибридных растений:
0,11 соответствует слабой полега-
а - Дон 4237×(Зарваси 70× Хейлуньдзян);
ющей соломине; 0,13 - соломина
б - Романика×(Дарий 122×Краснодар 9167).
средней прочности; 0,16 - соломи-
на прочная.
логически и генетически однородны, а на некоторых
Различия между значениями удвоенных гаплоидов
каллусных линиях наблюдается полная мономорф-
разных каллусных агрегатов одной каллусной линии
ность удвоенных гаплоидов по изученным признакам
и удвоенных гаплоидов двух каллусных линий одного
в условиях ex vitro [11]. Объединение морфологически
гибрида определяли в ходе дисперсионного анализа.
однородного семенного потомства удвоенных гаплоди-
Статистические параметры: среднее значение призна-
дов одной каллусной линии может обеспечить быстрое
ка (M), коэффициент вариации (Сv, %), коэффициент
размножение селекционного образца для ускорения се-
корреляции (r), критерий Фишера (F) рассчитывали в
лекционного процесса. Целью настоящего исследова-
программе Statistica. Всего в работе проанализировано
ния стало изучение внутрикаллусной и межкаллусной
1383 растения.
изменчивости удвоенных гаплоидов риса, полученных
Результаты и обсуждение. Выявлены статистиче-
в андрогенезе in vitro в условиях вегетационной пло-
ски значимые различия между удвоенными гаплоида-
щадки.
ми двух каллусных агрегатов одного пыльника: у ги-
Методика. Семенное потомство удвоенных гапло-
брида Р×Д×67(15) лишь по одному биометрическому
идов риса Oryza sativa L. первого поколения R1 высе-
показателю между каллусами 62.2.2 и 62.2.4 - высоте
13
Российская сельскохозяйственная наука, 2020, № 6
растений (при p=0,02); у гибрида К×23×К(26) по ше-
Самый большой образец из девяти растений отобран
сти признакам между калллусами 80.2.1 и 80.2.2 при
с каллусной линии 126.1.1 (531 г, Ic=0,17) - гибридное
p<0,03 (таблица). Вероятно, каллусные агрегаты од-
растение К×23×К(28). Ранее аналогичный анализ мы
ного пыльника развились из разных пыльцевых зерен,
провели с применением молекулярных маркеров [11].
что выразилось в различиях удвоенных гаплоидов од-
Семена удвоенных гаплоидов одной каллусной линии
ного пыльника. Далее разные каллусные агрегаты рас-
с идентифицированными генами устойчивости риса к
сматривали как различные пыльники одного гибридно-
пирикуляриозу Pi-ta2, после предварительной морфо-
го растения.
логической оценки были объединены и переданы для
Анализ различий между удвоенными гаплоида-
полевых испытаний.
ми разных пыльников показал достоверные различия
Таким образом, считаем возможным после первич-
по дисперсиям у двух представленных гибридов (та-
ной оценки линий удвоенных гаплоидов по биометри-
блица): по двум признакам у растений Д×З×Х(8) при
ческим показателям объединение семенного потомства
p=0,05 и по семи признакам у растений Р×Д×67(15)
наиболее продуктивных растений одной каллусной
при p<0,007. Удвоенные гаплоиды разных пыльников
линии с целью увеличения объема образца и его бы-
и различных каллусных агрегатов гетерогенны и не
строго размножения. Это позволит скорее перейти от
могут быть объединены в один образец без предвари-
рассмотрения продуктивности отдельных растений к
тельного анализа биометрических показателей.
полевой оценке урожайности.
Сравнили внутрикаллусный и межкаллусный уро-
вень изменчивости (Сv) удвоенных гаплоидов двух
Литература.
гибридных растений Д×З×Х(8) и Р×Д×67(15). Во мно-
1. Mishra R., Rao G.J.N. In-vitro androgenesis in rice:
гих случаях внутрикаллусный коэффициент вариации
advantages, constraints and future prospects // Rice
превышал межкаллусный или оставался таким же (ри-
Science. - 2016. - V. 23. - № 2. - P. 57-68.
сунок). Это означает, что существует внутрикаллусная
2. Dwivedi S., Britt A., Tripathi L., Sharma S., Upadhyaya
дифференциация удвоенных гаплоидов, поэтому целе-
H.D., Ortiz R. Haploids: constraits and opportunities
сообразно вести индивидуальный отбор регенерантов
первого поколения.
biotechadv.2015.07.001.
Выявлена корреляционная зависимость (p<0,05)
3. Germana M. Anther culture for haploid and doubled
ряда биометрических показателей от высоты растений:
haploid production // Plant Cell Tiss. Organ. Cult. -
длина метелки (r=0,72), масса зерна главной метелки
2011. - V. 104. - P. 283-300.
(r=0,80), масса зерна растения (r=0,59), масса 1000 зе-
4. Гончарова Ю.К., Харитонов Е.М. Генетические ос-
рен (r=0,74). Таким образом, высокорослые растения в
новы повышения продуктивности риса. - Красно-
нашем эксперименте оказались болеe продуктивными,
дар: ФГБНУ ВНИИ риса, Просвещение-Юг, 2015.
чем низко- и среднерослые. Все эти удвоенные гапло-
- 314 с.
иды были получены с гибридных растений одной ком-
5. Sarao N.K., Gosal S.S. In vitro androgenesis for
бинации К×23×К. В отечественной литературе среди
accelerated breeding in rice // Biotechnologies of crop
специалистов по зерновым культурам продолжается
improvement / S.S. Gosal, S.H. Wani (eds.). - Springer
обсуждение актуальности генов зеленой революции
International Publishing AG, Switzerland, 2018. - V.
(гены полукарликовости), которые позволили с середи-
1. - P. 407-435.
ны 60-х годов на очень высоком минеральном питании
6. Tripathy S.K., Swain D., Mohapatra P.M., Prusti A.P.,
значительно повысить урожайность пшеницы и риса за
Sahoo B., Panda S., Dash M., Chakma B., Behera
счет отсутствия полегаемости [27-29]. Между тем, ки-
S.K. Exploring factors affecting anther culture in rice
тайскими и японскими учеными выявлены природные
(Oryza sativa L.) // Journal of Applied Biology and
источники с генами IPM1 [30] и SCM2 [31, 32], кото-
Biotechnology. - 2019. - V. 7. - № 2. - P. 87-92.
рые ответственны за толстую прочную соломину риса.
7. Maharani A., Fanata W.I.D., Laeli F.N.L., Kim K.-M.,
Это высокорослые, устойчивые к полеганию растения,
Handoyo T. Callus induction and regeneration from
которые при значительных дозах удобрений обладают
anther cultures of Indonesian indica black rice culture //
более высокой зерновой продуктивностью, чем полу-
J. Crop. Sci. Biotech. - 2020. - V. 23. - № 1. - P. 21-28.
карликовые формы. Отдельные селекционные линии
8. Esteves P., Belzile F. TDZ in cereal gametic
рассматривают как перспективные, и некоторые их не-
embryogenesis // Thidiazuron: from urea deviation to
достатки пытаются откорректировать с помощью мето-
plant growth regulator / N. Ahmad, M. Faisal (eds.).
дов геномного редактирования [33]. Вероятно, можно
- Springer Nature Singapore Pte Ltd., 2018. - Р. 160-
говорить о наметившейся смене парадигмы в селекции
174.
риса на такие показатели как высота растений и устой-
9. Гончарова Ю.К., Харитонов Е.М., Бушман Н.Ю.,
чивость к полеганию.
Верещагина С.А. Сравнительный анализ эффек-
Для дальнейшей селекционной работы мы выдели-
тивности питательных сред для индукции каллусо-
ли линии удвоенных гаплоидов с высокими значения-
образования у гибридов риса // Доклады Российской
ми продуктивности растения, индекса прочности соло-
академии сельскохозяйственных наук. - 2013. - №
мины и массой 1000 зерен 30,0 г и более: с каллусной
6. - С. 6-9.
линии 88.2.1 - семенное потомство двух растений (75
10. Windarsih G., Utami D.W., Widyastuti U. Molecular
г, Ic=0,16), линии 97.1.1 - потомство одного растения
markers application for blast resistance selection on
(60 г, Ic=0,18) - гибридное растение Д×З×Х(8); удво-
the double haploid rice population // Makara J. Sci. -
енные гаплоиды гибридного растения Р×Д×67(15)
2014. - V. 18. - № 2. - P. 31-41.
сформировали мелкие семена - около 25 г, поэтому от-
11. Илюшко М.В., Ромашова М. В., Zhang J.-M., Deng L.-
бракованы; с каллусного агрегата 80.2.1 - семенное по-
W., Liu D.-J., Zhang R., Гученко С.С. Внутрикаллусная
томство пяти растений и с каллусного агрегата 80.2.2
изменчивость удвоенных гаплоидов риса, получен-
- четырех растений, объединенных в два отдельных
ных в андрогенезе in vitro // Сельскохозяйственная
селекционных образца (246 г, Ic=0,17 и 256 г, Ic=0,17
биология. - 2020. - Т. 55. - № 3. - С. 533-543.
соответственно) - гибридное растение К×23×К(26).
12. Yi G., Lee H.-S., Kim K.-M. Improved marker-assisted
14
Российская сельскохозяйственная наука, 2020, № 6
selection efficiency of multi-resistance in doubled
23. Satoh T., Tezuka K., Kawamoto T., Matsumoto S.,
haploid rice plants // Euphytica. - 2015. - V. 203. - P.
Satoh-Nagasawa N., Ueda K., Sakurai K., Watanabe
421-428.
A., Takahashi H., Akagi H. Identification of QTLs
13. Сартбаева И.А., Усенбеков Б.Н., Рысбекова А.Б.,
controlling low-temperature germination of the East
Мухина Ж.М., Казкеев Д.Т., Жамбакин К.Ж., Жан-
European rice (Oryza sativa L.) variety Maratteli //
бырбаев Е.А., Беркимбай Х.А., Ахметова Д.Ш.,
Euphytica. - 2016. - V. 207. - P. 245-254.
Мелдебекова А.А. Получение дигаплоидных линий
24. Гученко С.С. Оценка дигаплоидных линий риса пер-
для селекции глютинозного риса // Биотехнлогия. -
вого и второго поколений по хозяйственно ценным
2018. - Т. 34. - № 2. - С. 26-36.
признакам // Аграрная Россия. - 2018. - № 5. - С.
14. Колчанов Н.А., Кочетова А.В., Салина Е.А., Перши-
18-21.
на Л.А., Хлесткина Е.К., Шумный В.К. Состояние и
25. Илюшко М.В. Регенерационный максимум в андро-
перспективы использования маркер-ориентирован-
генных каллусных линиях риса Oryza sativa L. in vitro
ной и геномной селекции растений // Вестник Рос-
// Рисоводство. - 2019. - № 2. - С. 29-32.
сийской академии наук. - 2017. - Т. 87. - № 4. - С.
26. Ilyushko M.V., Romashova M.V. Formation of rice
348-354.
tetraploids in in vitro androgenesis // Russian Agricultural
15. Гончарова Ю.К. Использование метода культуры
Sciences. - 2020. - V. 46. - № 4. - P. 332-336.
пыльников в селекции риса. - Краснодар: ВНИИ
27. Крупрейшвили Н.Т., Вожжова Н.Н., Марченко Д.М.,
риса, 2012. - 91 с.
Ионова Е.В. Выявление гена короткостебельности
16. Илюшко М.В., Ромашова М.В. Создание регене-
Rht-b1 в образцах озимой мягкой пшеницы // Зерно-
рантных линий методом культуры пыльников in
вое хозяйство России. - 2019. - № 6 (66). - С. 55-59.
vitro для селекции риса на российском Дальнем
28. Костылев П.И., Краснова Е.В., Аксенова А.В. На-
Востоке // Дальневосточный аграрный вестник. -
следование ряда количественных признаков у ги-
2017. - № 4(44). - С. 37-45.
бридов риса Карлик1×LK // Зерновое хозяйство
17. Bishnoi U.S., Jain R.K., Gupta K.R., Chowdhury V.K.,
России. - 2018. - № 3 (57). - С. 43-47.
Chowdhury J.B. High frequency androgenesis in indica
29. Nagano H., Onishi K., Ogasawara M., Horiuchi Y.,
× Basmati rice hybrids using liquid culture media // Plant
Sano Y. Genealogy of the «Green revolution» gene in
Cell Tiss. Organ. Cult. - 2000. - Vol. 61. - P. 153-159.
rice // Genes Genet. Syst. - 2005. - V. 80. - P. 351-356.
18. Сибикеева Ю.Е., Сибикеев С.Н. Влияние комбина-
30. Jiao Y., Wang Y., Xue D., Wang J., Yan M., Liu G., Dong
ций чужеродных транслокаций на андрогенез in
G., Zeng D., Lu Z., Zhu X., Qian Q., Li J. Regulation of
vitro у почти изогенных линий яровой мягкой пшени-
OsSPL14 by OsmiR156 defines ideal plant architecture
цы // Генетика. - 2014. - Т. 50. - № 7. - С. 831-839.
in rice // Natural Genetics. - 2010. - V. 42. - № 6. - P.
19. Костылев П.И., Кудашкина Е.Б., Краснова Е.В., Во-
541-545.
жжова Н.Н. Селекция риса на солеустойчивость
31. Ookawa T., Hobo T., Yano M., Murata K., Ando T.,
// Зерновое хозяйство России. - 2019. - № 1. - С.
Miura H., Asano K., Ochiai Y., Ikeda M., Nishitani
22-27.
R., Ebitani T., Ozaki H., Angeles E., Hirasawa T.,
20. Михайлов М.Э. О новой возможности использова-
Matsuoka M. New approach for rice improvement using
ния дигаплоидных линий: схема обогатительно-вос-
a pleiotropic QTL gene for lodging resistance and yield
становительной селекции // Генетика. - 2010. - Т.
// Nature communications. - 2010. - Doi:10.1038/
46. - № 6. - С. 853-860.
ncomms1132.
21. Orasen G., Greco R., Puja E., Pozzi C. Blast resistance
32. Merugumala G.R., Satyanarayana P.V., Narne C.,
R genes pyramiding in temperate japonica rice //
Ravikumar B., Rao R., Pavani L., Deepika V. Molecular
Euphytica. - 2010. - V. 216. - P. 40-49.
breeding of «Swarna», a mega rice variety for lodging
22. Wu Y., Xiao N., Chen Y., Yu L., Pan C., Li Y., Zhang X.,
resistance // Mol. Breeding. - 2019. - V. 39. - P. 55-69.
Huang N., Ji H., Dai Z., Chen X., Li A. Comprehensive
33. Короткова А.М., Герасимова С.В., Шумный В.К.,
evaluation of resistancve effects of pyramiding lines
Хлесткина Е.К. Гены сельскохозяйственных рас-
with different broad-spectrum resistance genes against
тений, модифицированные с помощью системы
Magnaporthe oryzae in rice (Oryza sativa L.) // Rice. -
CRISPR/Cas // Вавиловский журнал генетики и се-
2019. - V. 12. - P. 11-24.
лекции. - 2017. - Т. 21. - № 2. - С. 250-258.
Поступила в редакцию 22.08.20
Принята к публикации 20.09.20
15