Российская сельскохозяйственная наука, 2021, № 6
УДК 631.527.34
DOI: 10.31857/S2500262721060065
ТРАНСГРЕССИВНАЯ СЕЛЕКЦИЯ: МЕТОДОЛОГИЯ УСКОРЕННОГО
ПОЛУЧЕНИЯ НОВЫХ ФОРМ РАСТЕНИЙ С ПРОГНОЗИРУЕМЫМ
КОМПЛЕКСОМ ХОЗЯЙСТВЕННО ЦЕННЫХ ПРИЗНАКОВ
А.А. Кочетов, кандидат биологических наук,
Г.В. Мирская, кандидат биологических наук,
Н.Г. Синявина, кандидат биологических наук, К.В. Егорова, аспирант
Агрофизический научно-исследовательский институт,
195220, Санкт-Петербург, Гражданский проспект, 14
E-mail: kochetoval@yandex.ru
Трансгрессивная селекция - одно из наиболее перспективных направлений развития современной сельскохозяйственной
науки. Феномен трансгрессии, несмотря на его широкое распространение, изучен недостаточно, что препятствует
эффективному использованию его в селекционном процессе. При прогнозируемом получении трансгрессий возможно
ускоренное создание новых сортов с заданным комплексом хозяйственно ценных признаков, адаптированных к различ-
ным условиям выращивания и технологиям производства. Цель исследований - разработка и апробирование методо-
логии, повышающей эффективность трансгрессивной селекции. Ключевой аспект методологии - изучение биоразноо-
бразия селектируемой культуры в регулируемой агроэкосистеме при специально создаваемых режимах выращивания,
позволяющих оценить потенциальные возможности генотипа и подобрать родительские пары для гарантированного
получения трансгрессий с прогнозируемым комплексом хозяйственно ценных признаков. Применение разработанной
методологии позволило за 5...7 лет создать ультраскороспелые линии яровой мягкой пшеницы; высокоустойчивые к
действию светотемпературных стрессоров Северо-Западного региона России линии дайкона и сорт Петербургский, а
также предназначенные для светокультуры высокопродуктивные образцы и сорт редиса Петербургский фиолетовый.
Предлагаемый подход способствует значительному уменьшению объема прорабатываемого материала и сокращает
сроки создания новых сортов различных культур.
TRANSGRESSIVE BREEDING: A METHODOLOGY FOR ACCELERATED
CREATING OF NEW FORMS OF PLANTS WITH A PREDICTABLE COMPLEX
OF ECONOMICALLY VALUABLE TRAITS
Kochetov A.A., Mirskaya G.V., Sinyavina N.G., Egorova K.V.
Agrophysical Research Institute
195220, St. Petersburg, Grazhdanskiy prosp., 14
E-mail: kochetoval@yandex.ru
Transgressive breeding is one of the most promising trends of development of modern agricultural science. The phenomenon of
transgression, despite its widespread distribution, has not been sufficiently studied, which prevents its effective use in the breeding
process. With the predicted getting of transgressions, it is possible to accelerate the creation of new accessions of plants with a given
complex of economically valuable traits adapted to different growing conditions and technologies. The purpose of this article is to
present the author’s view of the essence of the phenomenon of transgression and its role in breeding, as well as the methodology
of accelerated breeding, developed and tested at the Agrophysical Research Institute, based on the use of this phenomenon. The
key aspect of the methodology is the study of the biodiversity of the selected crop in a regulated agroecosystem with specially
created growing modes that allow assessing the potential of the genotype and selecting parent pairs for guaranteed obtaining
transgressions with a predicted complex of economically valuable traits. Application of the developed methodology made it possible
to breed ultra-early ripening lines of spring soft wheat in 5-7 years as well as highly resistant to the action of light-temperature
stressors of the North-West region of Russia daikon lines and daikon Petersburgsky variety. Besides that, highly productive lines
and the Petersburg violet small radish variety for intensive light culture were created. It is shown that this approach contributes to a
significant reduction in the volume of the material being worked out and reduces the time required for the creation of new cultivars
of different crops.
Ключевые слова: трансгрессивная селекция, светокультура,
Key words: transgressive breeding, light culture, breeding-
селекционно-ценные признаки, пшеница (Triticum aestivum L.),
valuable traits, wheat (Triticum aestivum L.), daikon, small
дайкон, редис (Raphanus sativus L.)
radish (Raphanus sativus L.)
В современном мире, столкнувшемся со значитель-
тивных, специфично адаптированных сортов, дающих
ным ростом населения, возрастающей техногенной на-
стабильно высокий урожай благодаря эффективному
грузкой на окружающую среду и глобальным измене-
использованию питательных веществ и почвенной вла-
нием климата, необходима смена парадигмы стратегии
ги, устойчивых к действию стрессовых факторов среды,
развития сельскохозяйственного производства. Одним
характерных для регионов выращивания, резистентных
из базовых элементов новой стратегии, ориентирован-
к возбудителям различных заболеваний. Немаловажным
ной на биологизацию и экологизацию интенсификаци-
требованием к новым сортам должны также стать их пи-
онных процессов, должна стать адаптивная селекция,
тательная ценность и биохимический состав, обеспечи-
предусматривающая усиление генетического контроля
вающие полноценное здоровое питание населения. Все
за устойчивостью растений к абиотическим стрессо-
это диктует необходимость кардинального обновления
рам и эффективностью использования благоприятных
и расширения ассортимента существующих сортов в
факторов внешней среды. Приоритетной задачей совре-
короткие сроки. С такими задачами селекция, ориенти-
менной селекции становится создание высокопродук-
рованная на использование традиционных подходов и
29
Российская сельскохозяйственная наука, 2021, № 6
методов, успешно справиться не может. Наиболее акту-
ства исследований, посвященных картированию локу-
альными требованиями к селекции на современном эта-
сов количественных признаков, в которых наблюдали
пе становятся повышение эффективности, уменьшение
трансгрессивное расщепление, основной причиной об-
объемов работ, ускорение селекционного процесса, а
разования трансгрессий выступает действие комплемен-
также создание новых сортов с прогнозируемым ком-
тарных генов. В качестве примера авторы ссылаются на
плексом хозяйственно ценных признаков. Необходима
работу L.M. Mansur et al. (1993), проанализировавших
разработка принципиально новых подходов и методов,
гибридное потомство от скрещивания двух фенотипиче-
обеспечивающих возможность реализации этих задач.
ски сходных сортов сои. Трансгрессивную сегрегацию
Перспективными направлениями современной селек-
благодаря комплементарным аллелям в этих опытах от-
ции выступают геномный отбор и редактирование ге-
мечали для всех 15 изученных признаков.
нома [1]. Однако традиционные подходы, включающие
Современные представления о механизмах образо-
трансгрессивную селекцию, имеют ряд преимуществ,
вания трансгрессий обсуждаются в статье de los Reyes
которые могут по-новому раскрыться при дополнении
B.G. [1]. Они включают в себя классическое объяснение
их современными методами.
образования трансгрессивных фенотипов через омни-
Одним из наиболее перспективных направлений,
генную теорию количественных признаков, дополнен-
решающих большинство существующих задач, может
ную современными парадигмами эпигенетики. Автор
стать развитие трансгрессивной селекции, ориентиро-
считает, что для феномена трансгрессивной сегрегации
ванной на целенаправленное получение трансгрессий
характерна физиологическая реконфигурация родитель-
по хозяйственно ценным признакам. В современной
ских геномов у гибридного потомства посредством пе-
биологии под термином «трансгрессия» понимается
ренастройки геномной сети, включающей в себя слож-
образование при гибридизации наследственно закре-
ные молекулярные синергии, контролируемые как на
пленных генотипов, превышающих амплитуды измен-
генетическом, так и на эпигенетическом уровнях. Од-
чивости обоих родителей по изучаемому признаку. В
ним из возможных механизмов появления трансгрес-
определении, приведенном в современном словаре Глаз-
сивных фенотипов, по его мнению, выступает измене-
ко В.И., Глазко Г.В. [2], повторяющем понятие, сформу-
ние областей метилирования ДНК, которое происходит
лированное в более ранней публикации (Картель Н.А. и
при геномном шоке, вследствие рекомбинации сильно
др., 1999), трансгрессия - это «эффект суммирующего
различающихся между собой родительских геномов,
действия полимерных генов, выражающийся в устой-
что влечет изменения в метаболомном профиле у ги-
чивом увеличении (положительная Т.) или уменьшении
бридных растений. В результате у отдельных особей от-
(отрицательная Т.) значения (величины) какого-либо по-
мечается появление новых, отличных от родительских,
лимерно наследующегося признака у отдельных особей
продуктов - белков, регуляторных факторов и др., что
F2, по сравнению с крайними (+ или -) значениями этого
может приводить к возникновению трансгрессий. Ав-
признака у родительских форм». В определении, дан-
тор предлагает пересмотреть стратегию селекции в на-
ном Р. Ригером и А. Михаэлисом [3], указывалось также,
правлении максимального использования генетического
что трансгрессия должна превышать спектр изменчи-
разнообразия культур для создания новых геномных и
вости не только родительских форм, но и F1. Ценность
эпигеномных комбинаций, путем взаимодействия со-
феномена трансгрессии для селекции очевидна. Еще в
временной геномной биологии с классическими подхо-
1935 г. Н. И. Вавилов писал: «Серьезным дефектом в
дами к созданию рекомбинантов.
развитии генетической теории селекции является само-
По сравнению с традиционными представлениями,
отстранение генетики от изучения сложных хозяйствен-
сводящими механизмы образования трансгрессий к ад-
ных признаков. Трансгрессии, типы трансгрессий, их
дитивным взаимодействиям, современные исследовате-
механизмы - это первоочередная задача, учитывая всю
ли рассматривают разные типы взаимодействий между
значимость трансгрессий» [4]. Однако практическое ис-
родительскими геномами при рекомбинациях, что спо-
пользование трансгрессий далеко не соответствует их
собствует развитию трансгрессивной селекции. Важным
биологической и хозяйственной важности. Сложивша-
обстоятельством, позволяющим эффективно использо-
яся ситуация связана со сложностью прогнозирования
вать феномен трансгрессии в селекционном процессе,
трансгрессий из-за недостаточной изученности генети-
выступает гомозиготность трансгрессивных генотипов
ческих механизмов, обусловливающих трансгрессивное
по селектируемым признакам, о которой в определени-
расщепление, различного понимания аспектов феноме-
ях говорится косвенно, как об устойчивом увеличении
на трансгрессии, а также терминологической неопреде-
или уменьшении признака у некоторых особей второго
ленности.
поколения, по сравнению с родителями. L.H. Rieseberg с
Согласно современной концепции образования
соавторами [5] считают трансгрессию проявлением фе-
трансгрессий, они возникают в результате взаимодо-
нотипической изменчивости, и степень гомозиготности
полнения у гибридных особей родительских генов, ко-
трансгрессивных форм не обсуждают. De los Reyes B.G.
торые образуют полиген и определяют селектируемый
[1] отмечает, что в отличие от гетерозиса, трансгрессив-
признак. Принято считать, что вероятность выявления
ные фенотипы, встречающиеся среди рекомбинантных
и целенаправленного подбора в одном генотипе значи-
форм, стабильны в транс-поколении (то есть гомозигот-
тельного числа генов (компонентов полигена), находя-
ны). Вероятность появления генотипов, гомозиготных
щихся в разных участках генома и обеспечивающих
по селекционно значимым количественным признакам,
образование трансгрессий вследствие аддитивного вза-
детерминирующимся значительным числом генов, во
имодействия, очень низка. В общепринятых определе-
втором поколении обычно крайне невысокая. Некото-
ниях трансгрессии отсутствует один из возможных ме-
рые исследователи считают, что выявление истинных
ханизмов ее образования, который отмечал Г. Нильсон
трансгрессий следует проводить не во втором, а в по-
Эле (1909, 1911) - комплементация двух разных систем
следующих поколениях гибридов: «…отбор трансгрес-
генов. Об этом механизме образования трансгрессий,
сивных форм из второго поколения может быть случай-
как об основном, сообщают L.H. Rieseberg с соавторами
ным… Более целесообразно его осуществлять, начиная
[5]. Они отмечают, что согласно результатам большин-
с третьего поколения, по скороспелости, морфологиче-
30
Российская сельскохозяйственная наука, 2021, № 6
ским признакам, качественным показателям, а по про-
тем, что в механизме проявления гетерозиса наблюда-
дуктивности - с четвертого» [6]. А.А. Трушко и С.П. Ха-
ются все формы межгенного неаллельного и алллельно-
лецкий [7] отмечают: «Общеизвестно, что большинство
го взаимодействия генов, природу этого явления до сих
отбираемых в F2 так называемых трансгрессий является
пор было трудно объяснить» [10]. То же можно конста-
гетерозиготами или положительными модификациями.
тировать и в отношении трансгрессии. Скорее всего, ме-
Поэтому вопрос времени проведения отбора ценных
ханизмы гетерозиса и положительной трансгрессии во
трансгрессивных форм в гибридных популяциях оста-
многом сходны, а их вклад в наблюдаемый эффект труд-
ется дискуссионным. Ряд авторов считает, что отбор по-
но оценить и разделить. В то время, как при гетерозисе
ложительных трансгрессий из второго поколения носит
эффект гетерозиготности гибридов первого поколения
случайный характер и его целесообразно проводить, на-
уменьшается в последующих поколениях, ослабляя его
чиная с третьего поколения».
последствия, при формировании трансгрессий аддитив-
Значительная часть исследователей относит к транс-
ное и комплементарное взаимодействие генов выходит
грессивным все особи гибридной популяции, фено-
на первый план, стабилизируется и не исчезает в потом-
типически превосходящие родительские формы по
стве.
селектируемым признакам, включая как гомо-, так и
Важным аспектом в понимании феномена транс-
гетерозиготные генотипы. Высокую частоту появления
грессии выступает ее экологическая зависимость. Эко-
трансгрессий, отмечаемую во втором поколении гибри-
лого-генетический подход к изучению трансгрессий
дов в ряде исследований, объясняют как раз этим обсто-
предусматривает приоритетное внимание к исследова-
ятельством, а также учетом как положительных, так и
нию механизмов взаимодействия генотип-среда, мето-
отрицательных трансгрессий. Сообщение L.H. Rieseberg
дов прогнозирования и управления ими. Как отмечает
с соавторами о том, что фактически в 155 из 171 проана-
академик РАН В.А. Драгавцев, гипотеза формирования
лизированных ими исследований (91 %) описывается по
трансгрессий, основанная на геноцентрической пара-
крайней мере один трансгрессивный признак, а 44 % из
дигме наследования, «опровергается фактами, когда
1229 исследованных признаков были трансгрессивны-
сорт-трансгрессия в зоне районирования дает макси-
ми [5], вероятно, соответствует действительности, если
мальный урожай, а при переносе в другую зону уходит
относить к трансгрессиям особи, которые отобраны по
на 5-е и даже 10-е место по урожаю среди других со-
фенотипу без учета их гомозиготности. О большом про-
ртов, то есть трансгрессии оказались экологически за-
центе выявления трансгрессий во втором гибридном
висимыми» [11].
поколении сообщают многие авторы. Так, при селекции
Теоретической базой эколого-генетического под-
овса по признаку «масса зерна с растения» установлено
хода к селекции растений, аккумулирующей основной
«60 % гибридных комбинаций от общего числа с отри-
спектр накопленных знаний о природе количественных
цательной и невысокой положительной степенью транс-
признаков и механизмах взаимодействия генотип-сре-
грессии» [7]. В другой работе частота появления поло-
да, служит созданная академиком В.А. Драгавцевым с
жительных трансгрессий по числу зерен с растения у
соавторами теория эколого-генетической организации
мягкой пшеницы варьировала от 51 до 55 % [8]. N. Putri
количественных признаков (ТЭГОКП) [12]. Согласно
с соавторами отмечали трансгрессивное расщепление
ее положениям, при смене лимитирующих факторов
(по массе зерна с растения у мягкой пшеницы) в обоих
внешней среды меняется спектр и число генов, опре-
направлениях у более чем 50 % гибридных популяций
деляющих уровень и генетическую дисперсию количе-
[9]. Наш опыт работы с редисом и дайконом показал, что
ственного признака. Важным аспектом также выступает
при селекции по сложным количественным признакам
то, что компоненты сложного признака закладываются в
(масса корнеплода, устойчивость к стрессорам) число
разные фазы онтогенеза и мультипликативно образуют
трансгрессивных форм (по фенотипу) часто превышало
результирующий признак. Поэтому при изучении слож-
50 %. В потомстве большинства выделенных форм от-
ного признака необходимо учитывать условия формиро-
мечали расщепление по селектируемым признакам при
вания его компонентов. Следствием теории эколого-ге-
самоопылении до 4…5 поколения.
нетической организации количественных признаков
Выделение гомозиготных трансгрессивных форм
служат представления о модульной организации слож-
среди других, превышающих родительские по феноти-
ного количественного признака и наличии у растений
пическим признакам, особенно важно для сокращения
семи генетико-физиологических систем, управляя ко-
сроков получения стабильных линий при селекции.
торыми селекционеры могут осуществлять повышение
Несомненно, превышение по фенотипу у части него-
продуктивности и урожайности, ориентируясь на эти
мозиготных трансгрессивных форм над родительски-
системы необходимо вести селекцию в целом и транс-
ми по селектируемым признакам возникает вследствие
грессивную селекцию в частности.
сверхдоминирования и кодоминирования, а также взаи-
На наш взгляд, с учетом современных представле-
модействия генотип-среда. L.H. Rieseberg с соавторами
ний о механизмах образования трансгрессий, ее опре-
на основании анализа данных различных исследований
деление должно быть следующим: «Трансгрессия - это
сообщают, что количественные генетические исследо-
устойчивое, наследственно закрепленное увеличение
вания гибридных популяций неизменно указывают на
(положительная Т.) или уменьшение (отрицательная Т.)
действие комплементарных генов как на первичную
значения (величины) какого-либо признака у отдельных
причину трансгрессий, хотя сверхдоминирование и
особей F2 или последующих поколений по сравнению с
эпистаз также вносят свой вклад [5]. Таким образом,
крайними (+ или -) значениями этого признака у роди-
фенотипически выраженная положительная трансгрес-
тельских форм, формирующееся благодаря всем типам
сия у гибридов второго или последующих поколений по
межаллельного и межгенного взаимодействия, а также
селектируемым признакам может обеспечиваться как
взаимодействия генотип-среда (эпигенетический эф-
межаллельным, так и межгенным взаимодействием. В
фект)».
образовании трансгрессий могут быть задействованы
Существует значительное число публикаций, посвя-
механизмы, аналогичные тем, что отмечены при про-
щенных получению трансгрессий: по устойчивости к
явлении гетерозиса. По мнению С. Хаблака, «в связи с
засолению у риса [1], созданию соле- и засухоустойчи-
31
Российская сельскохозяйственная наука, 2021, № 6
вых трансгрессивных генотипов у подсолнечника [13], у
оценка степени выраженности компонентов селек-
мягкой пшеницы по длине колоса и числу зерен в коло-
тируемых признаков при моделировании в РАЭС основ-
се [8], у пшеницы по высоте растений и числу стеблей
ных лимитирующих факторов среды, действующих в
[14], по показателям хлебопекарных свойств (общая
регионе селекции;
стекловидность, содержание белка, седиментация и
подбор родительских пар по принципу благоприят-
ряд других параметров) у озимой пшеницы [15], по
ного взаимодополнения разных компонентов селектиру-
содержанию белка у озимой мягкой пшеницы [16],
емых признаков, детерминируемых генами или блоками
по морозостойкости и продуктивности пшеницы
генов, обеспечивающих получение трансгрессий благо-
[17], по способности мягкой пшеницы накапливать в
даря комплементарным взаимодействиям;
зерне Zn и Fe [18]. Показана возможность использо-
проведение в регулируемых условиях скрещиваний
вания маркер-опосредованного отбора для селекции (в
и изучение гибридов F1 и F2, в сравнении с родитель-
том числе трансгрессивной) картофеля на устойчивость
скими формами, с целью получения и выявления транс-
к болезням и вредителям [19].
грессивных генотипов с прогнозируемым комплексом
Цель исследований - разработка и апробирование
хозяйственно ценных признаков;
методологии, повышающей эффективность трансгрес-
сравнение селектируемых признаков у гетерозисных
сивной селекции.
генотипов первого и трансгрессивных генотипов второ-
Для ее достижения необходимо решить две наиболее
го поколений с родительскими генотипами посредством
важные задачи:
модифицированного гибридологического анализа;
разработать эффективные методы изучения и оцен-
получение стабильных по исследуемым признакам
ки биоразнообразия селектируемой культуры и, на их
линий из отобранных трансгрессивных образцов путем
основе, принципы подбора родительских пар, обеспе-
инбридинга и стабилизирующего отбора с использова-
чивающие максимальное проявление трансгрессивного
нием режимов выращивания, ускоряющих генератив-
эффекта по селектируемым признакам у гибридного по-
ное развитие.
томства в конкретных условиях выращивания;
При работе по созданию ультраскороспелых линий
разработать экспресс-методы выявления
«истин-
яровой мягкой пшеницы было изучено 14 образцов из
ных» трансгрессий (гомозиготных по селектируемым
коллекции ФИЦ ВИР им. Н. И. Вавилова. Выращивание
признакам) в гибридном потомстве.
и оценку образцов, а также гибридов первого и после-
Это позволит целенаправленно получать формы с
дующих поколений проводили в РАЭС на биополигоне
прогнозируемым комплексом селектируемых призна-
АФИ в вегетационно-облучательных установках, осна-
ков, значительно уменьшить число прорабатываемых
щенных лампами ДНаТ-400, при облученности 50±5
комбинаций скрещивания и число генераций, необхо-
Вт/м2 ФАР. Режимы выращивания исходных образцов
димых для формирования стабильных линий. Возмож-
яровой мягкой пшеницы включали четыре варианта:
ность решения поставленных задач существует как на
12- и 20-часовое ежесуточное освещение, без яровиза-
молекулярном (анализ генома), так и на организменном
ции и с предварительной яровизацией наклюнувшихся
(оценка и подбор по фенотипу) уровне.
проростков. Гибридные растения культивировали в ус-
Методика. В результате многолетних исследований
ловиях 12-часового фотопериода без предварительной
феномена трансгрессии у разных культур и опыта ис-
яровизации. Этот режим использовали для обеспечения
пользования регулируемой агроэкосистемы (РАЭС) в
наиболее четкого фенотипического проявления разли-
селекционном процессе была разработана и апробиро-
чий исследуемого материала по генотипам систем Vrn
вана оригинальная методология ускоренной селекции
и Ppd, ответственных за реакцию на яровизирующие
[20]. Основоположником этих работ в АФИ была Г.А.
температуры и фотопериод, контролирующие длитель-
Макарова, пионерские работы которой по селекции
ность онтогенеза. Температурный режим поддерживали
редиса и яровой мягкой пшеницы заложили основу
на уровне 22...26 °C днем и 18...20 °C ночью. Растения
методологии получения новых трансгрессивных форм
выращивали в вегетационных сосудах объемом 2 л. В
растений с прогнозируемыми комплексом селекцион-
качестве корнеобитаемой среды использовали субстрат
но ценных признаков [21, 22]. Разработку и совершен-
на основе верхового торфа низкой степени разложения с
ствование методологии в дальнейшем проводили путем
минеральными добавками. Полив осуществляли водой,
генетико-селекционных исследований в регулируемой
подкормку - раствором Кнопа 2 раза в неделю. Повтор-
агроэкосистеме яровой мягкой и твердой пшеницы,
ность для каждого сорта и линии - 25-кратная, для ги-
редьки, дайкона и редиса.
бридов F1 - 15-кратная. В F2 высевали потомство 1…3
Среди основных особенностей методологии уско-
растений первого поколения в количестве 100…110
ренной трансгрессивной селекции новых линий и сортов
зерновок. Индивидуально для каждого растения фик-
растений с прогнозируемым комплексом хозяйственно
сировали сроки начала кущения, выхода в трубку, ко-
ценных признаков и высокой степенью адаптации к
лошения и созревания, проводили изучение элементов
агроэкологическим условиям их возделывания при ис-
продуктивности главного колоса (длина колоса, число
пользовании регулируемой агроэкосистемы можно вы-
колосков, число и масса зерен) и растения (высота сте-
делить следующие:
бля, число колосьев, число и масса зерен).
учет эколого-генетической организации количе-
Японская редька (дайкон) служила одним из важных
ственных признаков;
объектов проводимой селекционной работы для созда-
учет модульной структуры селектируемых призна-
ния линий и сортов, реализующих потенциал продук-
ков, представленной комплексом результирующего при-
тивности культуры в климатических условиях Ленин-
знака и формирующих его компонентов;
градской области [23]. Для достижения такой цели было
учет независимости наследуемости компонентов се-
необходимо объединить в генотипе высокую устойчи-
лектируемых признаков;
вость к длинному дню и пониженным температурам, а
использование режимов выращивания в РАЭС, обе-
также обеспечить формирование товарных корнеплодов
спечивающих уменьшение модификационной и повы-
при майской посадке в открытый грунт. В ходе работы в
шение четкости проявления генотипической изменчиво-
условиях РАЭС выявляли формы восточных редек, наи-
сти, а также режимов, ускоряющих развитие растений;
более устойчивые к действию каждого из стрессоров,
32
Российская сельскохозяйственная наука, 2021, № 6
вызывающих преждевременное стеблевание, подбира-
тической статистики с использованием программного
ли родительские пары по принципу взаимодополнения
обеспечения MS Exсel 2016.
по компонентам устойчивости, в потомстве которых
Результаты и обсуждение. Основным подходом к
были получены трансгрессивные формы с комплексной
получению трансгрессий традиционно выступает скре-
устойчивостью. Особенность наших экспериментов со-
щивание географически и генетически отдаленных
стояла в том, что существовала необходимость совме-
форм. Однако в большинстве работ отсутствует обсуж-
щения трансгрессии по размерам и массе корнеплода
дение механизмов образования трансгрессий и методов
с трансгрессией по устойчивости к действию соответ-
их целенаправленного получения.
ствующих стрессоров, провоцирующих преждевремен-
Наши представления о работе с феноменом транс-
ное цветение. Для этого в РАЭС изучали реакцию на
грессии заключаются, с одной стороны, в понимании
длинный день и пониженную температуру более чем 40
необходимости изучения наличия и локализации генов
сортов восточных редек (дайкон и лоба) из коллекции
или коадаптированных блоков генов, детерминирующих
ФИЦ ВИР им. Н. И. Вавилова. В качестве источников
селектируемые признаки и их компоненты, с примене-
света использовали лампы ДНаТ-400, облученность со-
нием современных молекулярно-генетических методов
ставляла 60…70 Вт/м2 ФАР. При изучении воздействия
в имеющейся коллекции сортов, с другой - в понимании
низкой температуры проклюнувшиеся семена выдержи-
невозможности прогнозирования реализации этих генов
вали в холодильнике при температуре 0…5 °C в течение
на уровне фенотипа без изучения влияния на них кон-
14 дней, затем высевали в субстрат (торф с керамзитом
кретных факторов среды. Один из основных принципов
1:1 по объему) и выращивали до начала стеблевания при
нашего подхода - гармонизация взаимодействия гено-
коротком 12-часовом фотопериоде. Для определения
тип-среда для получения и максимальной выраженности
влияния длинного дня сухие семена высевали в тот же
трансгрессий. Сложность межгенных взаимодействий и
субстрат и выращивали при 18-часовом фотопериоде до
взаимодействий генотип-среда заставляет выдвигать на
начала стеблевания. Температуру в помещении поддер-
первое место изучение не молекулярно-генетической
живали на уровне 22...26 °C днем и 18...20 °C ночью. По-
структуры объектов селекции, а фенотипических разли-
лив осуществляли водой, подкормку - раствором Кнопа
чий у имеющихся образцов в контролируемых условиях
2 раза в неделю. При создании гибридов F1 родитель-
выращивания. Работа на уровне фенотипа, подбор ро-
ские пары подбирали таким образом, чтобы длиннокор-
дительских пар с учетом взаимодополнения по компо-
неплодная форма обладала устойчивостью к одному из
нентам сложного признака, фактически подразумевает
неблагоприятных факторов среды, а круглокорнеплод-
использование не аддитивных эффектов полигенов, а
ная - к другому. Выявление трансгрессивных форм в F2
комплиментарных взаимодействий между генами или
и сравнение их с родительскими сортами и гибридами
блоками генов, детерминирующими как хозяйственно
F1 проводили в открытом грунте в Ленинградской обла-
ценные признаки, так и их компоненты.
сти при майском посеве на фоне обычно действующих
Предлагаемый подход связан с прогнозируемым по-
в регионе стрессоров. Повторность для каждого сорта
лучением трансгрессий по селектируемым признакам
и линии - 12-кратная, для гибридов F1 - 6-кратная.
(компонентам продуктивности и устойчивости) путем
При выращивании гибридов второго поколения изуча-
целенаправленного подбора родительских пар, исходя
ли 150...200 образцов в каждом варианте скрещивания.
из результатов проведенного в условиях регулируемой
Учитывали сроки начала цветения, надземную массу и
агроэкосистемы изучения биоразнообразия культуры.
массу корнеплода, число листьев в розетке, размеры и
Причем оценку первичного селекционного материала и
число долей у развитого листа, диаметр и длину корне-
выявление источников хозяйственно ценных признаков
плода, его окраску и форму.
возможно проводить, моделируя стрессовые воздей-
При селекции редиса исследовали особенности про-
ствия или комплекс факторов среды, типичных для зоны
дукционного процесса и проявления хозяйственно цен-
селекции. Использование регулируемой агроэкосистемы
ных признаков более чем 30 сортов из коллекции ФИЦ
позволяет более четко оценивать реакцию генотипа на
ВИР им. Н.И. Вавилова, а также российских селекци-
действие различных факторов среды и их комбинаций,
онно-семеноводческих фирм в условиях интенсивной
значительно снижая влияние экологических шумов, не-
светокультуры. Растения культивировали при 12-часо-
избежно возникающих при проведении селекции в от-
вом фотопериоде в вегетационно-облучательной ярус-
крытом грунте. Наиболее важной характеристикой при
ной установке, оборудованной лампами ДНаЗ-400 при
оценке первичного селекционного материала, помимо
облученности 60…70 Вт/м2 ФАР. Температуру поддер-
общей продуктивности, служит степень выраженности
живали на уровне 22...26 °C днем и 18...20 °C ночью.
компонентов селектируемых признаков. Полученные
Субстратом служил верховой торф с минеральными
данные служат основой для подбора родительских пар
добавками, толщина корнеобитаемого слоя составляла
и прогноза возможных трансгрессий, а также эталона-
4…5 см, плотность посева - 100…200 растений на 1 м2.
ми для идентификации в расщепляющихся гибридных
Полив и подкормку проводили по схеме, аналогичной
популяциях трансгрессивных генотипов. Применяемый
дайкону. Повторность для каждого сорта и линии -
метод подбора родительских пар (взаимодополнение по
40-кратная, для гибридов F1 - 20-кратная, в F2 высевали
разным независимо наследуемым компонентам селекти-
150…200 растений, в F3 - 40…60 растений. Для полу-
руемых признаков) позволяет на порядок повысить эф-
чения семян маточные корнеплоды после 2-недельного
фективность селекции, по сравнению с традиционными
воздействия низкой температуры (0…5 °C) высаживали
методами, путем сокращения числа прорабатываемых
в сосуды объемом 2 л (торф с минеральными добавка-
гибридных комбинаций и уменьшения числа растений
ми), выращивая их при 16-часовом фотопериоде. Учи-
в изучаемых гибридных популяциях при более четкой
тывали высоту и диаметр розетки, массу растения и
идентификации трансгрессий по фенотипу в регулиру-
корнеплода, число и размеры листьев, длину и диаметр
емой агроэкосистеме. Все эти элементы разработанной
корнеплода, степень опушенности листа, устойчивость
методологии ускоренной трансгрессивной селекции
к стеблеванию.
могут быть наглядно продемонстрированы на примере
Обработку данных проводили методами матема-
наших работ по селекции различных культур.
33
Российская сельскохозяйственная наука, 2021, № 6
Реализация разработанной методологии транс-
ствующих четкости фенотипического проявления ге-
грессивной селекции при создании ультраскороспелых
нетических систем Vrn и Ppd. Благоприятным фоном
линий яровой мягкой пшеницы (Triticum aestivum L.). В
для оценки их вкладов в фенотипы компонентных (про-
работе с мягкой пшеницей главой задачей исследова-
должительность отдельных периодов онтогенеза) и ре-
ний было изучение в РАЭС механизмов, определяю-
зультирующего (срок колошения) признаков выступал
щих скороспелость и обеспечивающих получение уль-
короткодневный 12-часовой режим освещения без пред-
траскороспелых трансгрессивных форм. В ее основу
варительной яровизации, при выращивании на котором
была положена концепция независимого наследования
фенотипическая дисперсия по признакам длительности
длительности периодов онтогенеза пшеницы «всхо-
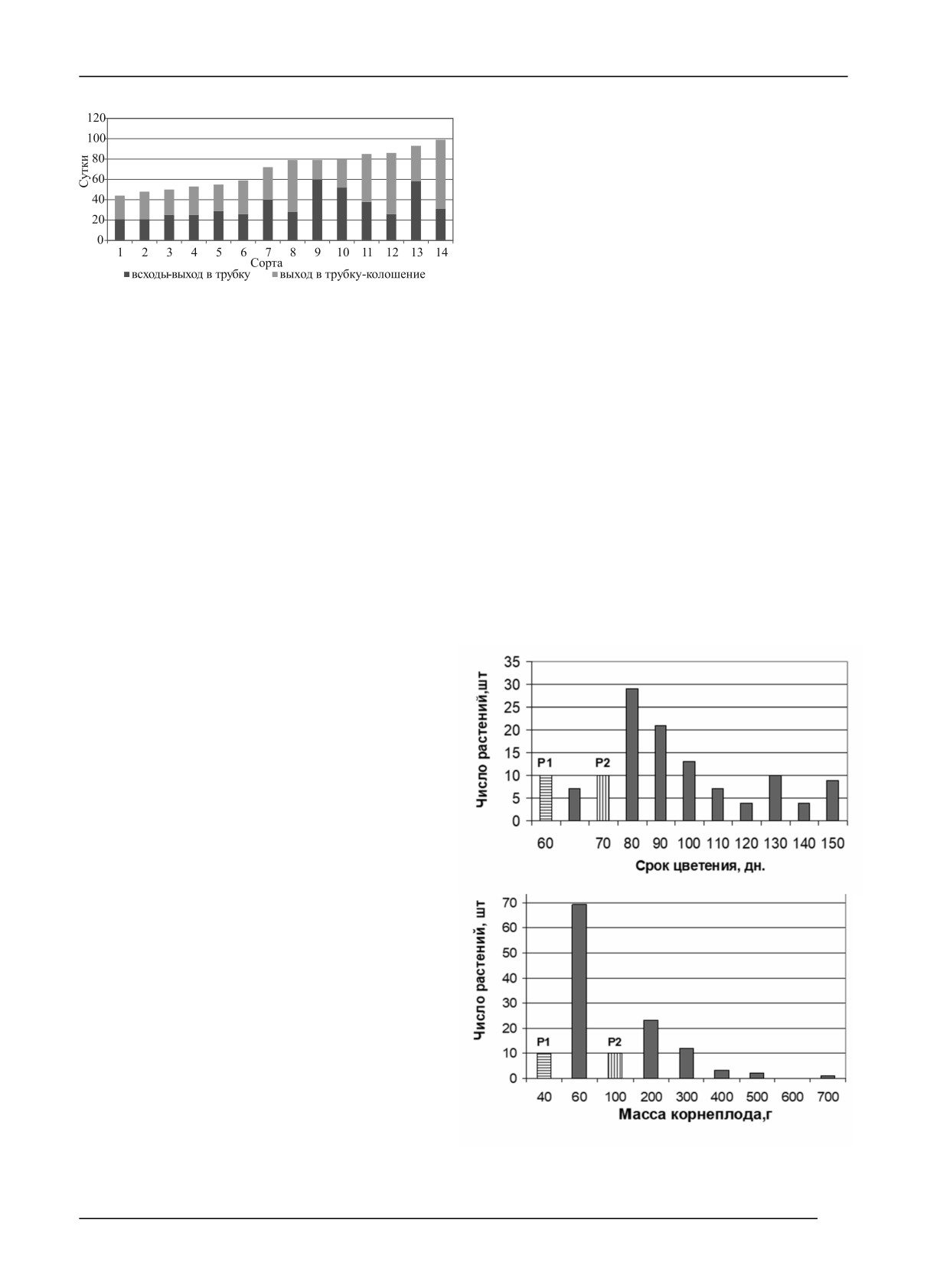
периодов онтогенеза выражена очень отчетливо (рис. 1).
ды - выход в трубку», «выход в трубку - колошение»
Это иллюстрирует преимущество РАЭС, по сравнению
[20, 22]. Эта концепция открывает возможности для
с природными условиями, состоящее в возможности
целенаправленного создания трансгрессивных по сро-
создания режимов выращивания, обеспечивающих сни-
ку колошения генотипов с использованием взаимодо-
жение модификационной изменчивости и более четкое
полняющего подбора компонентов родительских форм
выявление генотипической изменчивости.
(длительность периодов), определяющих степень про-
явления селектируемого признака.
Продолжительность вегетационного периода яровой
мягкой пшеницы выступает важным фактором, опреде-
ляющим адаптацию растений к различным условиям
окружающей среды и, в конечном счете, их продуктив-
ность [24]. До фазы выхода в трубку пшеница находится
в вегетативном периоде развития, от начала колошения
до конца цветения - в генеративном, от первой стадии
созревания до полной спелости - в репродуктивном
периоде. Различные фазы определяют формирование
определенных количественных показателей продук-
тивности. Увеличение длительности периода всходы -
Рис. 1. Длительность периода от всходов до колошения
кущение сопровождается наращиванием вегетативной
у сортов яровой мягкой пшеницы при 12- и 20-часовом
массы. Многие авторы отмечают, что продолжитель-
фотопериодах: 1 - СКФ 2; 2 - Gecora 70; 3 - Santa Elena;
ность фазы выход в трубку - колошение (IV…VII эта-
4 - Белорусская 80; 5 - Triple Dirk D; 6 - Ленинградка;
пы органогенеза) важна, поскольку в этот период фор-
7 - Родина; 8 - Triple Dirk E; 9 - Московская 35;
мируются фертильные цветки, определяющие будущее
10 - АФИ-М-1; 11 - Ленинградская ранняя; 12 - Sonora 64;
число зерен в колосе [25]. Во время периода колошение
13 - Triple Dirk B; 14 - Asakaze komugi.
- созревание (IX…XII этапы органогенеза) происходят
налив и увеличение массы зерна [26].
В отличие от широко распространенного способа
Эффективную адаптацию пшеницы в процессе онто-
оценки особенностей генетических систем Vrn и Ppd,
генеза к разным условиям среды обеспечивают генети-
период всходы - колошение был рассмотрен не как еди-
ческие системы, контролирующие реакцию растений на
ный этап онтогенеза, а как два подэтапа: «всходы - вы-
яровизирующие температуры (гены Vrn-vernalisation) и
ход в трубку», «выход в трубку - колошение». Прогноз
фотопериод (гены Ppd-photoperiod response) [27, 28, 29].
количественных характеристик трансгрессий по при-
Следует отметить, что основное количество работ по
знаку срок колошения пшеницы осуществляли по фор-
генетике вегетационного периода у пшеницы связано с
муле, изложенной в методике [20].
изучением продолжительности периода от всходов до
Данные, полученные при оценке срока колошения
колошения. В литературе имеются немногочисленные
по его компонентной структуре в наборе сортов яровой
исследования по изучению длительности отдельных
мягкой пшеницы на провокационном фоне (короткий
фаз развития этой культуры. Выявлены различия по вре-
день), позволили выявить ряд комбинаций скрещива-
мени наступления и скорости прохождения отдельных
ния, перспективных для получения трансгрессий по
этапов органогенеза у различающихся по локусам Vrn
скороспелости пшеницы (рис. 2).
и Ppd генотипов [30, 31, 32]. Установлено, что действие
Подбор комбинаций скрещивания проводили по
генов Ppd длится с середины IV до конца VI этапов ор-
принципу благоприятного взаимодополнения компо-
ганогенеза и оказывает влияние на скорость роста и раз-
нентов селектируемого признака (продолжительность
вития апекса побега [32, 33, 34]. Показано, что различия
периода всходы - колошение). Очевидно, что трансгрес-
в проявлении локусов Vrn A1, Vrn B1 и Vrn D1 связаны
сивный по скороспелости эффект возможен в тех комби-
с разной скоростью прохождения II и III этапов (куще-
нациях скрещивания, где один из родительских сортов
ние-выход в трубку). Отмечено, что период «кущение-
характеризуется минимальной длительностью периода
выход в трубку» выступает критическим и определяет
всходы - выход в трубку, а второй сорт - коротким пе-
продолжительность всего вегетационного периода от
риодом выход в трубку - колошение. В анализируемой
всходов до колошения и созревания растений [35, 36,
выборке сортов наиболее перспективными оказались
37]. Адаптивность пшеницы к широкому диапазону ус-
комбинации скрещивания Asakaze komugi × Ленинград-
ловий выращивания во многом определяет взаимодей-
ская ранняя, Asakaze komugi × Sonora 64. У гибридов F2
ствие систем генов Vrn и Ppd, представленных разной
в этих комбинациях скрещивания, согласно сделанному
долей доминантных и рецессивных аллелей. Регуляция
прогнозу, отмечалось трансгрессивное расщепление по
времени цветения и созревания позволяет растениям из-
сроку колошения. В ходе дальнейшей работы из выде-
бегать стрессовых климатических условий [28].
ленных трансгрессивных гибридных форм F2, путем
Ключевую роль при получении ультраскороспелых
стабилизирующего отбора на провокационном фоне,
форм мягкой пшеницы сыграло проведение исследова-
был получен ряд константных ультраскороспелых ли-
ний в регулируемой агроэкосистеме, обеспечивающее
ний AFI 39, AFI 91, AFI 68, AFI 177, опережающих по
возможность создания анализирующих фонов, способ-
сроку колошения и родительские скороспелые сорта, и
34
Российская сельскохозяйственная наука, 2021, № 6
шей селекции оказались комбинации лобы Красная
мелкая с сортами дайкона Акицумари и Химани. При
майской посадке в открытый грунт оба родительских
сорта и гибриды F1 зацвели очень рано, практически
не образовав товарных корнеплодов, в то время как у
гибридов в F2 отмечали трансгрессивное расщепление
как по сроку перехода к цветению, так и по массе кор-
неплода (рис. 3). В гибридном потомстве была выявле-
на высокая степень корреляции между сроком перехода
растений к цветению и массой корнеплода (r=0,76).
Отсутствие гетерозиса в первом поколении в изучен-
ных комбинациях скрещивания было, вероятно, связано
Рис. 2. Структура срока колошения по длительности
с эпистатическим действием генов, детерминирующих
составляющих его периодов онтогенеза сортов яровой
устойчивость (отсутствие рецессивных гомозигот), по
мягкой пшеницы при 12-часовом фотопериоде: 1 - СКФ
2; 2 - Родина; 3 - АФИ-М-1; 4 - Gecora 70; 5 - Sonora 64;
отношению к генам, определяющим форму и массу
6 - Santa Elena; 7 - Triple Dirk D; 8 - Белорусская 80;
корнеплода. В ходе дальнейшей работы из выделенных
9 - Asakaze komugi; 10 - Triple Dirk E; 11 - Ленинградка;
гибридных форм, обладающих значительной массой
12 - Ленинградская ранняя; 13 - Triple Dirk B;
корнеплода и не зацветающих до конца сентября при
14 - Московская 35.
весенней посадке в открытый грунт, путем стабилизи-
рующего отбора на провокационном фоне и инбридинга
районированные сорта Ленинградка, Ленинградская 6,
были созданы стабильные высокопродуктивные линии
как при выращивании в регулируемых условиях, так и
дайкона, устойчивые к комплексу светотемпературных
в условиях естественного фотопериода Северо-Западной
стрессоров. Масса корнеплода у отдельных экземпляров
зоны РФ [38, 39].
этих линий достигала к концу сентября 2...3 кг при со-
Предлагаемый подход не ограничен вариантом полу-
хранении сочной мякоти и отсутствии стеблевания. Ста-
чения ультраскороспелых линий, а позволяет создавать
билизированную по форме и цвету корнеплода, устой-
формы с различными сроками колошения, в перспекти-
чивую к действующим в регионе стрессорам линию,
ве - адресно ориентированные сорта, адаптированные
выделенную в комбинации скрещивания дайкон Акицу-
к конкретным условиям региона выращивания и совре-
мари × лоба Красная мелкая, зарегистрировали как сорт
менным агротехнологиям.
дайкона Петербургский [40]. Другая линия в комбина-
Реализация методологии ускоренной трансгрес-
ции скрещивания дайкон Химани × лоба Красная мел-
сивной селекции при создании новых форм Raphanus
кая не была зарегистрирована из-за сходства с дайконом
sativus L. Разработанная нами стратегия интродукции
Петербургский по устойчивости и продуктивности.
дайкона в Северо-Западный регион России была реа-
лизована при проведении большой части исследований
в регулируемой агроэкосистеме. Особенно это касает-
ся изучения влияния светового дня и низких положи-
тельных температур на переход растений к цветению.
Показано, что при 18-часовом фотопериоде значитель-
ная часть восточных редек (дайкон и лоба) не пере-
ходит к цветению и способна образовывать товарные
корнеплоды, если к этому не добавляется действие
пониженных температур и других стрессоров [23]. Это
опровергает широко распространенное представление
о высокой цветушности большинства сортов дайкона
в условиях длинного дня, которая приводит к раннему
стеблеванию и отсутствию товарных корнеплодов. Та-
кой результат был получен благодаря моделированию
действия в РАЭС только одного из факторов, провоци-
рующих преждевременное стеблевание, при поддер-
жании других условий выращивания в зоне оптимума.
Значительно меньшее количество образцов восточных
редек оказалось слабо чувствительными к действию
пониженных температур. При низкотемпературной
индукции проклюнувшихся семян большинство ис-
следованных сортов быстро переходило к стеблеванию
и образовывало небольшие корнеплоды. В результате
проведения комплексных исследований в РАЭС были
отобраны сорта, наиболее устойчивые к действию каж-
дого из имеющихся стрессоров в отдельности. Подбор
родительских пар по принципу благопрятного взаимо-
дополнения по компонентам устойчивости и продук-
тивности позволил получить в их гибридном потом-
стве высокопродуктивные трансгрессивные генотипы
с комплексной устойчивостью.
Среди комбинаций скрещивания, подобранных на
Рис. 3. Расщепление по сроку цветения и массе корнеплода
основании проведенных исследований, наиболее пер-
у гибридных растений F2 в комбинации скрещивания
спективными для получения трансгрессий и дальней-
дайкон Химани (Р1 ) × лоба Красная мелкая (Р2 ).
35
Российская сельскохозяйственная наука, 2021, № 6
Анализ результатов, полученных при селекции
гарантированно создавать новые сорта различных куль-
восточных редек, свидетельствуют о том, что их неу-
тур, адаптированные к конкретным условиям и техноло-
стойчивость к изучаемым неблагоприятным факторам
гиям выращивания.
(длинный день, низкие положительные температуры
Развитие разработанной методологии трансгрессив-
в начале вегетации) доминирует над устойчивостью;
ной селекции с использованием современных методов
большие размеры корнеплода доминируют над меньши-
молекулярной генетики позволит усовершенствовать
ми; гены, контролирующие реакцию на факторы среды,
предлагаемый подход благодаря возможностям моле-
эпистатичны по отношению к генам, обусловливающим
кулярно-генетического скрининга исходного материала
размеры и массу корнеплода. Благодаря использованию
и гибридных популяций, более четкому картированию
разработанной методологии удалось за короткий срок
селекционно значимых количественных признаков с
(4…5 лет) создать линии дайкона, адаптированные к
учетом влияния факторов среды, а также осуществлять
комплексу стрессоров, характерных для Северо-За-
разработку молекулярно-генетических маркеров для по-
падного региона России, ограничившись изучением
следующей оценки и характеристики объектов селекции.
небольшого числа комбинаций скрещивания и относи-
тельно небольших выборок гибридных популяций.
Литература.
Еще один пример эффективности использования
1. De los Reyes B. G. Genomic and epigenomic bases of
разработанной селекционной методологии - создание
transgressive segregation - New breeding paradigm for
новых сортов редиса, адаптированных для выращива-
novel plant phenotypes // Plant Science. 2019. Vol. 288.
ния в светокультуре. Потребность в таких сортах связа-
на с широким распространением технологии сити-фар-
article/pii/S016894521930528X. (дата обращения:
минга, позволяющей массово производить свежую
14.09.2021).
овощную продукцию в мегаполисах, в сооружениях
2. Глазко В. И., Глазко Г. В. Толковый словарь терминов
искусственного климата при искусственном освещении.
по общей и молекулярной биометрии и общей и при-
При селекции сортов для светокультуры основными
кладной генетике. М.: Академкнига, 2008. Т. 2. 529 с.
признаками выступают адаптация к искусственному ос-
3. Ригер Р., Михаэлис А. Генетический и цитогенетиче-
вещению, малообъемным технологиям промышленного
ский словарь / Под ред. д-ра биол. наук Я. Л. Глембоц-
выращивания и повышенным температурам. В результа-
кого. М.: Колос, 1967. 607 с.
те проведенных исследований было создано несколько
4. Вавилов Н. И. Пути советской селекции // Теоретиче-
новых трансгрессивных линий редиса, превышающих
ские основы селекции. М.: Наука, 1987. С. 40-68.
при выращивании в интенсивной светокультуре по тем-
5. Rieseberg L.H., Archer M. A., Wayne R. K. Transgressive
segregation, adaptation and speciation // Heredity. 1999.
пам роста, товарности и массе корнеплода все изучен-
Vol. 83(4). P. 363 -372. doi:10.1038/sj.hdy.6886170.
ные сорта. Одна из линий с корнеплодом фиолетового
6. Минькач Т. В., Селихова О. А. Селекционно-генети-
цвета была зарегистрирована в 2021 г. как сорт редиса
ческая оценка межвидовых гибридов сои третьего
Петербургский фиолетовый для светокультуры [41].
поколения // Вестник Алтайского государственного
Он характеризуется комплексом хозяйственно ценных
аграрного университета. 2012. Т. 94 (8). С. 26-28.
признаков: при урожайности 3,5...4,0 кг/м2 за 25 суток
7. Трушко А. А., Халецкий С. П. Трансгрессия признаков
вегетации устойчив к стеблеванию в условиях свето-
у гибридов овса посевного в селекции на продуктив-
культуры, обладает компактной листовой розеткой и ма-
ность // Земледелие и селекция в Беларуси. 2019. №
лоопушенным листом салатного типа (рис. 4).
55. С. 325-332.
Таким образом, разработанная методология уско-
8. Studies on transgressive segregation in wheat (Triticum
ренной трансгрессивной селекции позволяет наиболее
aestivum Linn.) / D. Dahat, H. Rajput, B. Chavan, et al. //
эффективно использовать феномен трансгрессии для
Trends in Biosciences. 2017. Vol. 10 (1). P. 104-107.
получения новых форм различных культур с прогнози-
9. Wheat transgressive segregants and their adaptation in
руемым комплексом хозяйственно ценных признаков,
the tropical region / N. Putri, S. Sutjahjo, A. Nur, et al.
что подтверждают результаты селекции пшеницы, дай-
// Journal of Breeding & Genetics. 2020. Vol. 52 (4). P.
кона и редиса. Целенаправленное использование мето-
506-522.
дологических возможностей регулируемой агроэкоси-
10. Хаблак С. Новая теория гетерозиса. Концепция ал-
стемы значительно ускоряет селекционный процесс,
лельного и неаллельного механизма возникновения ге-
сокращает объем прорабатываемого материала при про-
терозиса / М.: ЛитРес, 2019. 70 с.
гнозируемом получении трансгрессий по селектируе-
11. Драгавцев В. А. К выяснению эпигенетической (эко-
мым признакам. Моделирование действия стрессоров и
лого-генетической) природы трансгрессий признаков
различных сочетаний экологических условий выращи-
продуктивности растений // 50 лет ВОГИС: успехи
вания в регулируемой агроэкосистеме дает возможность
и перспективы: сб. тезисов Всерос. конф. М.: Изд-во
Вавиловского общества генетиков и селекционеров,
2016. С. 63-66.
12. Модель эколого-генетического контроля количе-
ственных признаков растений / В. А. Драгавцев, П.
П. Литун, Н. М. Шкель и др. // Доклады АН СССР.
1984. Т. 274 (3). С. 720-723.
13. Major ecological transitions in wild sunflowers facilitated
by hybridization / L. H. Rieseberg, O. Raymond, D. M.
Rosenthal, et al. // Science. 2003. Vol. 301 (5637). P.
1211-1216. doi:10.1126/science.1086949.
14. Inheritance and genetic mapping of the reduced height
(Rht18) gene in Wheat / N. P. Grant, A. Mohan, D.
Sandhu, et al. // Plants. 2018. Vol. 7(3). P. 58-66. URL:
Рис. 4. Сорт редиса Петербургский фиолетовый
для светокультуры.
ращения: 14.09.2021).
36
Российская сельскохозяйственная наука, 2021, № 6
15. Analysis of genetic control and QTL mapping of essential
29. Kamran A., Iqbal M., Spaner D. Flowering time in
wheat grain quality traits in a recombinant inbred
wheat (Triticum aestivum L.): a key factor for global
population / S. Goel, K. Singh, B. Singh, et al. // PLoS
adaptability // Euphytica. 2014. Vol. 197. P. 1-26. URL:
semanticscholar.org/c0e0/9121e897fff7e46bcebb89f007
1075-7 (дата обращения: 14.09.2021).
3601303f87.pdf (дата обращения: 14.09.2021).
30. Стельмах А.Ф. Роль генетических систем в онтоге-
16. Фоменко М. А., Грабовец А. И. Селекция озимой мяг-
нетической адаптации мягкой пшеницы // Экологи-
кой пшеницы на качество зерна в степной зоне // До-
ческая генетика и эволюция. Кишинев: Штиинца,
стижения науки и техники АПК. 2016. Т. 30. № 2. С.
1987. С. 146-161.
78-81.
31. Kosner J., Pankova K. Chromosome substitutions with
17. Грабовец А. И., Фоменко М. А. Плюстрансгрессии
dominant loci Vrn_1 and their effect on developmental
при селекции озимой пшеницы на морозостойкость
stages of wheat // Czech journal of genetics and
и продуктивность // Российская сельскохозяйствен-
plant breeding. 2004. Vol.
40
(2). P. 37-44. doi:
ная наука. 2019. № 4. С. 3-7. doi:10.31857/S2500-
10.17221/3698-CJGPB.
2627201943-7.
32. Емцева М. В., Ефремова Т. Т., Арбузова В. С. Влияние
18. QTL mapping of grain Zn and Fe concentrations in two
аллелей Vrn-B1a и Vrn-B1c на продолжительность
hexaploid wheat RIL populations with ample transgressive
фаз развития замещенных и изогенных линий мягкой
segregation / L. A. Crespo-Herrera, V. Govindan, J.
пшеницы // Генетика. 2013. Т. 49 (5). С. 632-632. doi:
Stangoulis, et al. // Frontiers in Plant Science. 2017.
10.7868/S0016675813050056.
Vol.
33. Стельмах А. Ф., Мартынюк В. Р. Эффекты доми-
articles/10.3389/fpls.2017.01800/full?report=reader.
нантных генов Ppd по особенностям органогенеза
(дата обращения: 14.09.2021).
у озимой мягкой пшеницы // Цитология и генетика.
19. Использование генетических ресурсов картофеля для
1998. Т. 32 (6). С. 27-34.
повышения эффективности селекции / Е.А. Симаков,
34. Файт В.И., Стельмах А.Ф., Федорова В.Р. Начало
В.А. Жарова, А.В. Митюшкин и др. // Труды по приклад-
включения и продолжительность экспрессии генов
ной ботанике, генетике и селекции. 2017. Т. 178 (2). С.
фотопериодической реакции у озимой мягкой пшени-
113-121. doi:10.30901/2227-8834-2017-2-113-121.
цы // Цитология и генетика. 2006. T. 40 (2). С. 12-19.
20. Методология прогнозирования трансгрессий по хо-
35. Зырянова А. Ф., Лихенко Н. И., Салина Е. А. Изучение
зяйственно-ценным признакам растений: Методиче-
аллельного состава генов Vrn-1 и Ppd-1 у раннеспе-
ские рекомендации / Г.А. Макарова, Г.В. Мирская, А.А.
лых и среднеранних сортов яровой мягкой пшеницы
Кочетов и др. СПб.: Россельхозакадемия, 2009. 48 с.
Сибири // Вавиловский журнал генетики и селекции.
21. Макарова Г. А., Иванова Т. И. Наследование призна-
2015. Т. 18 (4/1). С. 691-703.
ков корнеплода и листа у редиса // Генетика. 1983. Т.
36. Environmental and Genetic Variation for Yield-Related
10 (2). С. 304-311.
Traits of Durum Wheat as Affected by Development / F.
22. Макарова Г. А., Ермаков. Е. И. Прогнозированный
Giunta, P. De Vita, A. M. Mastrangelo, et al. // Frontiers
трансгрессивный по сроку колошения рекомбиноге-
in Plant Science. 2018. Vol. 9. P. 8. URL: ttps://www.
нез пшеницы // Доклады Российской академии сель-
frontiersin.org/articles/10.3389/fpls.2018.00008/full
скохозяйственных наук. 1993. № 5. С. 3-5.
(дата обращения: 14.09.2021).
23. Кочетов А. А. Генотипическая адаптация восточно-
37. Чуманова Е. В., Ефремова Т. Т., Кручинина Ю. В. Вли-
азиатских подвидов Raphanus sativus при интродук-
яние различных доминантных аллелей локусов VRN
ции в Северо-Западный регион России // Сельскохо-
и их комбинаций на продолжительность фаз раз-
зяйственная биология. 2004. Т. 39 (1). С. 83-91.
вития и продуктивность у линий мягкой пшеницы //
24. Worland A. The influence of flowering time genes
Генетика. 2020. Т. 56 (7). С. 805-818. doi: 10.31857/
on environmental adaptability in European wheats
S0016675820070024.
// Euphytica. 1996. Vol. 89. P. 49-57. doi:10.1007/
38. Макарова Г. А., Нерушева Г. В. Прогноз длительно-
BF00015718.
сти периодов онтогенеза у рекомбинантных по сро-
25. González F. G., Slafer G. A., Miralles D. J. Grain and
ку колошения линий пшеницы // Доклады Российской
floret number in response to photoperiod during stem
академии сельскохозяйственных наук. 1995. № 2. С.
elongation in fully and slightly vernalized wheats // Field
10-12.
Crops Research. 2003. Vol. 81. P. 17-27. doi:10.1016/
39. Ермаков Е. И., Макарова Г. А., Нерушева Г. В. Про-
S0378-4290(02)00195-8.
граммированное получение в регулируемой агроэко-
26. Slafer G. A., Rawson H. M. Sensitivity of wheat phasic
системе трансгрессивных по сроку колошения линий
development to major environmental factors: a re-
пшеницы: Методические рекомендации. М.: РАСХН,
examination of some assumptions made by physiologists
2002. 32 с.
and modelers // Austral. J. Plant Physiology. 1994. Vol.
40. Патент на селекционное достижение 6392 (8953530)
21. P. 393-426. doi:10.1071/PP9940393.
РФ дайкон Петербургский / А. А. Кочетов, А. М. Ар-
27. Waiting for fine times: genetics of flowering time in
темьева; заявитель и патентообладатель: ГНУ
wheat / J. W. Snape, K. Butterworth, E. Whitechurch,
Агрофизический НИИ. N 8953530; заявл. 27.10.2010;
Worland A. J. // Euphytica. 2001. Vol. 119. P. 185-190.
выдан 22.03.2012.
doi:10.1007/978-94-017-3674-9_7.
41. Патент на селекционное достижение 11518 РФ, ре-
28. Distelfeld A., Li C., Dubcovsky J. Regulation of
дис Raphanus sativus var. sativus Петербургский фио-
flowering in temperate cereals // Current opinion in plant
летовый / А. А. Кочетов, Н. Г. Синявина; заявитель и
biology. 2009. Vol. 12 (2). P. 178-184. doi:10.1016/j.
патентообладатель: ФГБНУ Агрофизический НИИ.
pbi.2008.12.010.
N 8058521; заявл. 28.112019; выдан 25.03.2021.
Поступила в редакцию 30.06.2021
После доработки 03.09.2021
Принята к публикации 24.09.2021
37