Российская сельскохозяйственная наука, 2022, № 6
УДК 636.52/.58:611.013
DOI: 10.31857/S2500262722060102, EDN: MKMPDF
СИНЕРГЕТИЧЕСКОЕ ВОЗДЕЙСТВИЕ ПЕРЕМЕННОЙ ТЕМПЕРАТУРЫ И КРАСНОГО
СВЕТОДИОДНОГО ОСВЕЩЕНИЯ ВО ВРЕМЯ ИНКУБАЦИИ НА РОСТ, МЕТАБОЛИЗМ КУРИНЫХ
ЭМБРИОНОВ И КАЧЕСТВО СУТОЧНЫХ ЦЫПЛЯТ ЯИЧНОГО КРОССА
М.И. Челнокова, кандидат биологических наук, Ф.И. Сулейманов, доктор ветеринарных наук,
А.А. Челноков, доктор биологических наук
1Великолукская государственная сельскохозяйственная академия,
182112, Псковская обл., Великие Луки, просп. Ленина, 2
Е-mail: marinachelnokova@yandex.ru
Исследование проводили с целью оценки влияния переменной температуры и красного светодиодного освещения отдельно
и в сочетании во время инкубации яиц на рост куриных эмбрионов, развитие их висцеральных органов и метаболизм, а
также качество суточных цыплят. Инкубацию яиц кросса Ломанн Браун (n=600) с 1 по 21 день проводили в инкубаторе
ИЛБ-0,5 с использованием 3 режимов: ПТ - инкубация при переменных температурах в темноте (37,8 °С - 1…14 сутки;
39,5 °С в течение 2 ч ежедневно - 15…17 сутки; 37,5 °С - 18 сутки; 37,0 °С - 19…21 сутки; относительная влажность
воздуха - 57,0 %); КС - круглосуточное красное освещение при стандартной температуре инкубации 37,6±0,1 °С с от-
носительной влажностью воздуха 55,0 %; ПТ+КС - инкубация при переменных температурах с круглосуточным крас-
ным освещением. С использованием морфометрических методов проведена оценка росто-весовых показателей куриных
эмбрионов, их висцеральных органов (сердце, мышечный желудок, печень, селезёнка), длины и массы тела суточных цы-
плят. Для оценки частоты сердечных сокращений (ЧСС), интенсивности дыхания по СО2 и теплопродукции у куриных
эмбрионов использовали метод скейлинга. Режим ПТ+КС к периоду вылупления (19 сутки) приводил к снижению массы
тела эмбрионов, по сравнению с КС, на 3,464 г, массы селезёнки, в сравнении с ПТ, - на 0,005 г, но оптимизировал уровень
метаболизма (по выделению СО2), по сравнению с указанными вариантами, соответственно на 47,20 и 21,42 мл/ч. ЧСС
и уровень теплопродукции эмбрионов кур на 19 сутки при разных режимах инкубации варьировали в незначительных
пределах без чётких различий. Эффект режима ПТ+КС проявлялся повышение массы тела цыплят, в сравнении с ПТ, на
1,93 г. Предполагаемые нейроэндокринные механизмы регуляции роста, метаболизма, иммунных реакций, терморегуляции
и реакций на стресс куриных эмбрионов на основе синергетического эффекта ПТ+КС представлены в виде блок-схемы.
THE SYNERGISTIC EFFECT OF VARIABLE TEMPERATURE AND RED LED LIGHTING
DURING INCUBATION ON THE GROWTH, METABOLISM OF CHICKEN EMBRYOS
AND THE QUALITY OF DAY-OLD EGG-CROSS CHICKENS
Chelnokova M.I., Suleymanov F.I., Chelnokov А.А.
1State Agricultural Academy of Velikie Luki,
182112, Pskovskaya obl., Velikie Luki, prosp. Lenina, 2
Е-mail: marinachelnokova@yandex.ru
The effect of variable temperature, red LED lighting and variable temperature with red LED lighting during egg incubation on the
growth of chicken embryos, the development of their visceral organs and metabolism, as well as the quality of day-old chickens were
studied. Incubation of Lohmann Brown cross eggs (n=600) from 1 to 21 days was carried out in an ILB-0.5 incubator using 3 modes:
VT - incubation at variable temperatures in the dark (37.8 °C - 1...14 days; 39.5 °C for 2 hours daily - 15...17 days; 37.5 °C - 18 days;
37.0 °C - 19...21 days; relative humidity - 57.0%); RL - round-the-clock red LED lighting at a standard incubation temperature of
37.6±0.1 °C with a relative humidity of 55.0%; VT+RL - incubation at variable temperatures with round-the-clock red LED lighting.
Morphometric methods were used to evaluate the height and weight parameters of chicken embryos, their visceral organs (heart,
muscular stomach, liver, spleen), and the length and body weight of day-old chickens. To assess the heart rate, respiration rate by CO2,
heat production in chicken embryos, the scaling method was used. The results of the studies showed that the VT+RL regime by the
hatching period (day 19) suppressed the growth of embryo body weight by 3.464 g, compared with RL and spleen weight by 0.005 g
compared with VT, but optimized the metabolic rate (by CO2 release) by 21.42 and 47.20 ml/h compared to with VT and RL options.
The heart rate and the level of heat production of chicken embryos on day 19 varied slightly under different incubation regimes and
differences were no revealed. The effect of the VT+RL regime was manifested by an increase in the body weight of chickens by 1.93 g
compared to VT. The proposed neuroendocrine mechanisms of regulation of growth, metabolism, immune responses, thermoregulation
and stress responses of chicken embryos based on the synergistic effect VT+RL are discussed in the form of a block diagram.
Ключевые слова: куриные эмбрионы, переменная температура,
Key words: chicken embryos, variable temperature, red LED
красный светодиодный свет, висцеральные органы, частота сер-
light, visceral organs, heart rate, metabolism, heat production,
дечных сокращений, метаболизм, теплопродукция, гипоталамо-
hypothalamic-pituitary system, epigenetic adaptation.
гипофизарная система, эпигенетическая адаптация.
Рост и метаболизм суточного цыплёнка в основном
развития куриных эмбрионов и их выживания в процессе
определяют процессы, происходящие во время эм-
инкубации. Изменять эти физиологические параметры и
брионального развития. Основная цель коммерческих
влиять на эмбриональное развитие птицы могут такие
инкубаториев - достижение максимального вывода вы-
условия инкубации, как температура, свет, гипоксия,
сококачественных суточных цыплят. Для решения этой
гипероксия и гиперкапния [1, 2].
задачи жизненно важно определить факторы, влияющие
В естественных условиях инкубации происходят
на оптимальное эмбриональное развитие кур [1]. Такие
изменения температуры и освещения, когда наседка по-
физиологические параметры, как теплопродукция, метабо-
кидает гнездо для кормления и поения [3] или поднима-
лизм, газообмен (O2, CO2), имеют решающее значение для
ется, чтобы перевернуть яйца в ответ на зов эмбриона [4].
51
Российская сельскохозяйственная наука, 2022, № 6
Переменные температуры инкубации, а также освещение
Для оценки физиологических показателей (частота
яиц светодиодными лампами положительно влияет на
сердечных сокращений (ЧСС), интенсивность дыхания
эмбриональный рост, выводимость, качество цыплят и их
по СО2, базальный метаболизм (теплопродукция)) ис-
рост после вылупления [5, 2]. Варьирование температуры и
пользовали метод скейлинга [8-10]. ЧСС и интенсивность
освещения (фотопериод, цвет, интенсивность освещения)
дыхания по СО2 рассчитывали на 14-е, 17-е, 19-е сутки,
яиц во время инкубации способствует адаптации цыплят
уровень базального метаболизма у эмбрионов кур - на
к окружающей среде после вылупления посредством
17-е и 19-е сутки инкубации.
адаптивной реакции. Это обусловлено эпигенетическими
ЧСС у куриных эмбрионов определяли по формуле
изменениями в ходе эмбриогенеза, которые служат основ-
аллометрической зависимости средней ЧСС в течении 80
ными механизмами адаптации к постинкубационной среде
% инкубации (СрЧСС, уд/мин-1) от массы яйца (Мяйца, г),
продуктивной птицы [2]. Однако до сих пор нет сведений
предложенной H. Tazawa с соавторами [8] (1):
о синергетическом воздействии переменной температуры
СрЧСС = 371×(Мяйца) - 0,121.
(1)
и красного светодиодного освещения во время инкубации
Для определения интенсивности дыхания использова-
яиц на рост, метаболизм куриных эмбрионов яичного
ли формулу аллометрической зависимости уровня выде-
кросса и качество суточных цыплят.
ляемого углекислого газа (QСО2, мл/ч) от массы тела (Мтела,
Цель исследования - оценка влияния переменной
г) эмбрионов кур в гомойотермной фазе, предложенную
температуры, красного светодиодного освещения от-
А.М. Болотниковым с соавторами [9] (2):
дельно и в сочетании во время инкубации яиц на рост
(2)
Q СО2 = 1,84×Мтела 0,739.
куриных эмбрионов, развитие их висцеральных органов
Для описания уровня базального метаболизма
и метаболизм, а также качество суточных цыплят яичного
(теплопродукции) в зависимости от скорости роста и
кросса Ломанн Браун.
массы тела куриных эмбрионов применяли уравнение
Методика. Объектом для исследований служили
наименьших квадратов, предложенное C.M. Vleck и
инкубационные яйца кур кросса Ломанн Браун (ОАО
D.F. Hoyt [10] (3):
«Волжанин» Ярославская область, пос. Ермаково), кото-
P = 12,17×GR + 1,66×M + 1,81,
(3)
рые по комплексу основных показателей соответствовали
где P - уровень базального метаболизма, ккал/ч; GR
нормативам, подтверждающим их качество. Возраст
- скорость роста, г/сутки; М - масса эмбриона, г; 1,66
поголовья родительского стада, от которого получено
ккал/г·ч - коэффициент массы, характеризующий затраты
инкубационное яйцо, - 30 недель. Инкубацию яиц с 1-го
на поддерживающий метаболизм; 1,81 - константа, про-
по 21-й день проводили в инкубаторе ИЛБ-0,5 (Волгасель-
гнозируемая скорость метаболизма эмбриона с нулевой
маш, Россия) с использованием 3 режимов:
массой (М) и скоростью роста (GR).
ПТ - инкубация при переменных температурах в
Статистический анализ данных проводили в програм-
темноте (37,8 °С - 1…14 сутки; 39,5 °С в течение 2 ч еже-
ме Statistica 10.0.1011.0 (Statsoft Inc, USA, 2010). Нормаль-
дневно - 15…17 сутки; 37,5 °С - 18 сутки; 37,0 °С - 19…21
ность распределения выборок определяли с использовани-
сутки; относительная влажность воздуха - 57,0 %);
ем Shapiro-Wilk’s W-test и Levene’s test. При нормальном
КС - круглосуточное красное освещение при стандарт-
распределении выборок применяли параметрический
ной температуре инкубации 37,6±0,1 °С с относительной
дисперсионный анализ One-way Anova с апостериорным
влажностью воздуха 55,0 %;
анализом Bonferroni test, при ненормальном - непараме-
ПТ+КС - инкубация при переменных температурах с
трический критерий Mann-Whitney U-test.
круглосуточным красным освещением.
Результаты и обсуждение. Под воздействием ПТ+КС
В предварительных исследованиях мы установили,
на 14-е сутки эмбриогенеза масса тела куриных эмбрио-
что максимальная скорость роста эмбрионов кросса кур
нов была меньше, чем в варианте ПТ, на 1,245 г (табл. 1,
Ломанн Браун наблюдается при красном светодиодном
р=0,013а), КС - на 0,611 г (р=0,045b), массы сердца - на
спектре с длиной волны 632 нм, чем при зелёном [6]. A.
0,010 г (р=0,030a), в сравнении с ПТ. Масса мышечного
Drozdova с соавторами [7] обнаружили, что инкубация
желудка в этот же срок развития при ПТ была больше, по
яиц при красном (длина волны 632 нм) освещении яиц,
сравнению с КС, на 0,057 (р=0,005c), с ПТ+КС - на 0,045
по сравнению с синим (463 нм), приводила к увеличению
г (р=0,005a), селезёнки - на 0,002 г (р=0,005c), только
массы тела цыплят в постнатальный период (с 18 по 21
по сравнению с вариантом КС. На 17-е сутки развития
дни). Поэтому для освещения яиц во время инкубации ис-
воздействие ПТ+КС подавляло рост длины и массы тела
пользовали красные светодиодные неоны (Elektrostandard
на 0,784 см и 5,049 г, сердца - на 0,023 г, мышечного
LS001, Россия-Китай), которые устанавливали на верхней
желудка - на 0,354 г, печени - на 0,099 г, по сравнению с
части инкубатора. Они излучали монохроматический крас-
ПТ (р=0,005a), селезёнки - на 0,004 и 0,003 г, в сравнении
ный свет с длиной волны 632 нм. Предварительно перед
с ПТ и КС (р=0,005a,c). При КС на 17-е эмбриогенеза на-
инкубацией яйца взвешивали, отбирали по приблизитель-
блюдали увеличение массы тела на 4,790 г, сердца - на
но одинаковой массе методом пар-аналогов и закладывали
0,041 г, мышечного желудка - на 0,344 г, печени - на
в инкубатор по 200 шт. при каждом экспериментальном
0,098 г, в сравнении с ПТ+КС (р=0,005b). На 19-е сутки
режиме. В общей сложности в ходе эксперимента при трёх
влияние ПТ+КС, а также ПТ проявлялось достоверно
режимах было заложено 600 инкубационных яиц.
значимым снижением массы тела куриных эмбрионов на
Для анализа морфологических росто-весовых по-
3,464 г (р=0,030b) и 1,916 г (р=0,045c), в сравнении с КС,
казателей куриных эмбрионов и их висцеральных ор-
соответственно. К 19-м суткам инкубации воздействие
ганов (сердце, мышечный желудок, печень, селезёнка)
ПТ стимулировало увеличение длины тела на 0,379 см, по
инкубируемые яйца (n=5) вскрывали на 14-е, 17-е, 19-е
сравнению с КС (р=0,016c), и массы селезёнки на 0,005 г,
сутки. Длину тела эмбрионов измеряли от верхушки
в сравнении с КС и ПТ+КС (р=0,016a,c).
черепа до конца хвоста электронным штангенциркулем
Вне зависимости от режима инкубации прослежи-
Finch Industrial Tools 19856 (Canada Inc.), длину тела цы-
вается общая тенденция снижения ЧСС, повышение
плят - от гребня до среднего пальца стопы. Массу тела
выделения СО2 и теплопродукции у куриных эмбрионов
эмбрионов и цыплят, а также отдельных висцеральных
с увеличением срока инкубации (рис. 1). Достоверно
органов определяли на аналитических весах Сартогосм
значимые различия в ЧСС наблюдали только на 17-е
ЛВ 210-А (Россия).
сутки при использовании режима инкубации с ПТ+КС.
52
Российская сельскохозяйственная наука, 2022, № 6
Табл. 1. Влияние режима инкубации на абсолютные размеры тела эмбрионов кур и их висцеральных органов*
Режим инкубации
Показатель
ПТ
КС
ПТ + КС
14-е сутки развития
ДТ, см
6,867 ±0,100
6,702±0,139
6,713±0,057
МТ, г
11,666±0,255a
11,032±0,147b
10,421±0,227
МС, г
0,105±0,002a
0,096±0,002
0,095±0,002
ММЖ, г
0,326±0,007a,c
0,269±0,003
0,281±0,005
МП, г
0,235±0,007
0,224±0,008
0,222±0,007
МСел, г
0,009±0,001c
0,007±0,001
0,008±0,001
17-е сутки развития
ДТ, см
8,324±0,012a
8,135±0,375
7,540±0,198
МТ, г
21,100±0,441a
20,841±0,731b
16,051±0,564
МС, г
0,161±0,005a
0,179±0,011b
0,138±0,004
ММЖ, г
0,825±0,018a
0,815±0,013b
0,471±0,029
МП, г
0,468±0,003a
0,467±0,015b
0,369±0,018
МСел, г
0,014±0,001a,c
0,011±0,001
0,010±0,001
19-е сутки развития
ДТ, см
9,064±0,108c
8,685±0,0,061
8,806±0,148
МТ, г
28,617 ±0,714
30,533±0,869b,c
27,069±0,680
МС, г
0,205 ±0,006
0,235±0,018
0,216±0,003
ММЖ, г
1,220 ±0,077
1,384±0,152
1,167±0,021
МП, г
0,606 ±0,028
0,541±0,049
0,526±0,022
МСел, г
0,019 ±0,001a,c
0,014±0,001
0,014±0,001
*здесь и в табл. 2 ПТ - переменные температуры; КС - красный светодиодное освещение; ПТ+КС - переменные температуры в сочетании с
красным светодиодным освещением; a - статистически значимое различие между показателями при переменной температуре в сочетании с
красным освещением (ПТ+КС), по отношению к переменной температуре (ПТ); b - статистически значимое различие между показателями
при переменной температуре в сочетании с красным светодиодным освещением (ПТ+КС), по отношению к красному свету (КС); c - стати-
стически значимое различие между показателями при переменной температуре (ПТ) и красным светодиодным освещением (КС). ДТ - длина
тела, МТ - масса тела, МС - масса сердца, ММЖ - масса мышечного желудка, МП - масса печени, МСел - масса селезёнки.
Величина этого показателя возросла, по сравнению с
(р=0,004), чем при режиме с ПТ, и на 73,09 мл/ч и
режимом с ПТ, на 8,71 уд/мин-1 (р=0,000), КС - на и
246,17 ккал/сут (р=0,000), в сравнении с режимом с КС. На
8,26 уд/мин-1, что вероятно обусловлено стресс-реакцией
19-е сутки развития при совместном воздействии ПТ+КС
куриных эмбрионов на совместное воздействие пере-
уровень выделяемого СО2 был ниже, чем при режиме с
менной температуры и светодиодного освещения во
КС на 47,20 мл/ч (р=0,030), ПТ - на 21,42 мл/ч (р=0,077).
время инкубации.
Уровень теплопродукции эмбрионов кур на 19-е сутки
При ПТ+КС выделение СО2 куриными эмбрионами на
развития варьировал в незначительных пределах и чёткой
14-е сутки развития было ниже, по сравнению с эмбриона-
зависимости выявить не удалось (р>0,05).
ми, развивающимися при режимах с ПТ и КС, на 21,71 мл/ч
Режимы ПТ+КС и КС во время инкубации положи-
(р=0,013) и 10,76 мл/ч (р=0,045), соответственно. Ана-
тельно влияли на развитие суточных цыплят (табл. 2).
логичную тенденцию наблюдали на 17-е сутки развития,
Выявлено достоверно значимое увеличение массы их тела
когда уровень выделения СО2 и теплопродукции в вари-
на 2,35 г (р=0,000а) при ПТ+КС и на 1,93 г (р=0,004с) при
анте ПТ+КС был меньше на 77,06 мл/ч и 245,73 ккал/сут
КС, по сравнению с ПТ. Достоверных изменений в длине
Рис. 1. Влияние ПТ, КС и ПТ+КС на ЧСС (а), интенсивность дыхания (б) и уровень базального метаболизма (в)
у эмбрионов кур на 14-е, 17-е, 19-е сутки развития;
- ПТ,
- КС,
- ПТ + КС.
53
Российская сельскохозяйственная наука, 2022, № 6
Табл. 2. Влияние режимов инкубации
на основе синергетического эффекта переменной тем-
на качество суточных цыплят
пературы и красного светодиодного освещения (рис. 2).
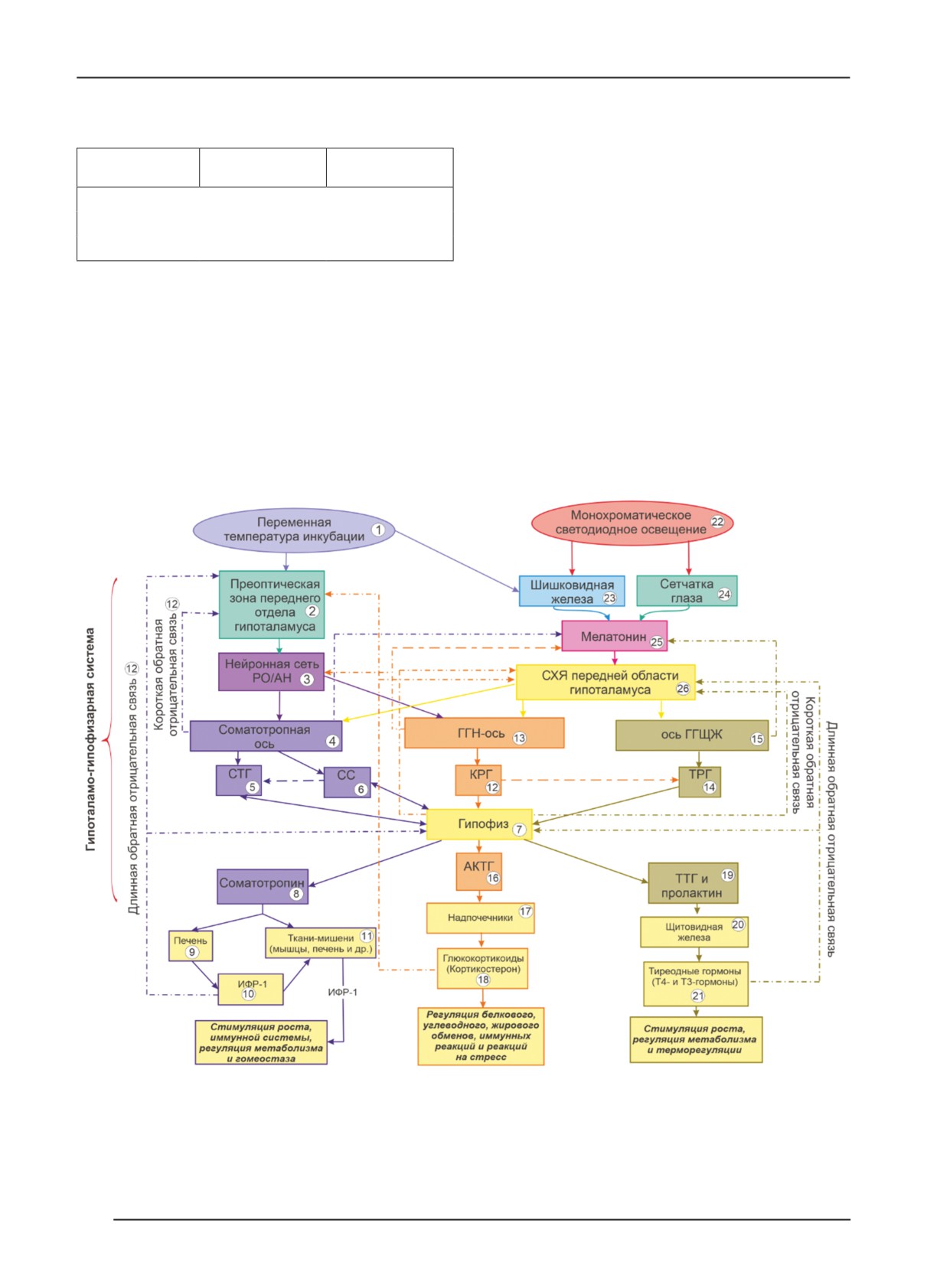
Согласно этой схеме, соматотропная ось (4) служит
Режим
основным нейроэндокринным путём регуляции роста и
Длина тела, см
Масса тела, г
инкубации
метаболизма птиц [12]. С её участием контролируется
синтез гормона инсулиноподобного фактора роста-1
ПТ (n=10)
18,21±0,15
43,11±0,30
(ИФР-1) (10) [13]. Соматотропин-рилизинг гормон (СТГ)
КС (n=10)
18,27±0,20
45,04±0,39с
(5), секретируемый преотической зоной переднего от-
дела гипоталамуса (2), воздействует на переднюю долю
ПТ+КС (n=10)
18,32±0,21
45,46±0,29а
гипофиза (7), вызывая секрецию соматотропина (гормона
роста) (8). Соматостатин (СС) (6) снижает продукцию
тела суточных цыплят в зависимости от режима инкубации
СТГ (5) гипофизом. Печень (9) вносит значительный
не наблюдали.
вклад в концентрацию гормона инсулиноподобного фак-
Основную роль в регуляции роста, метаболизма,
тора роста-1 (ИФР-1) (10) в плазме крови, и его секреция
терморегуляции и реакции на стресс куриных эмбрионов
запускается соматотропином (8). Остальные ИФР-1
отводят нейроэндокринным механизмам. Кроме того, есть
вырабатываются в различных тканях (11) с инициацией
сведения, описывающие развитие стрессовой реакции у
соматотропина и без него [13]. Регуляция секреции как
куриных эмбрионов под действием исследуемых режимов
соматотропина, так и ИФР-1 опосредована механизмами
инкубации [1, 2, 11]. На основе собственных результатов
короткой и длинной отрицательной обратной связи (12)
исследования и литературных данных разработана веро-
с участием гипоталамуса (2) [14]. Известно, что высо-
ятная блок-схема центральных нейроэндокринных меха-
кая температура инкубации (39,5 °C в течение 3 или 6 ч
низмов регуляции роста, метаболизма, иммунных реакций,
ежедневно) на поздних этапах эмбриогенеза (16…18-е
терморегуляции и реакций на стресс куриных эмбрионов
сутки) увеличивает ЧСС и потребление О2 эмбрионами
Рис. 2. Блок-схема центральных нейроэндокринных механизмов регуляции роста, метаболизма, иммунных реакций,
терморегуляции и реакций на стресс куриных эмбрионов на основе синергетического воздействия переменных
температур и монохроматического красного освещения: ГГН-ось - гипоталамо-гипофизарно-надпочечниковая ось; ось
ГГЩЖ - ось гипоталамус-гипофиз-щитовидная железа; СТГ - соматотропин-рилизинг гормон; СС - соматостатин;
КРГ - кортикотропин-рилизинг гормон; ТРГ - тиреотропин-рилизинг гормон; АКТГ - адренокортикотропный
гормон; ТТГ - тиреотропный гормон; ИФР-1 - инсулиноподобный фактор роста-1; СХЯ - супрахиазматическое ядро
передней области гипоталамуса; Т4 - тироксин, Т3 - трийодтиронин.
54
Российская сельскохозяйственная наука, 2022, № 6
кросса Cobb, повышает ИФР-1 скелетных мышц, усиливая
ГГЩЖ (15) и ГГН-ось (13) ответственны за ритмичную
пролиферацию и дифференцировку мышечных клеток, а
выработку мелатонина (25) [25, 26, 2].
также диаметр миофибрилл [15]. S. Yalcin с соавторами
Ранние исследования показали, что температура 39 °C
[16] установили, что воздействие температуры 38,8 °C с
в течение 3 ч/сут на 16…18-е сутки в сочетании с зелёным
10-х по 14-е сутки эмбриогенеза привело к увеличению
светодиодным освещением с 6-х суток до вывода стимули-
массы тела, повышению экспрессии ИФР-1 и увеличению
рует пролиферацию миобластов куриных эмбрионов Ross
площади волокон в грудной мышце 42-дневных цыплят-
308 [27, 28]. В работе T. Shah и S. Özkan [29] показано,
бройлеров кроссов Ross308 и Cobb.
что повышение температуры яичной скорлупы до 38,5 °C
Существует гипотеза о том, что воздействие кратков-
в течение 6 ч в сутки на 11…16-е сутки в комбинации с
ременных переменных или постоянных температур ниже
фотопериодом (16 ч освещение, 8 ч темнота) увеличивает
или выше оптимальных приводит к развитию эпигенети-
длину тела суточных цыплят кросса Ross 308 и массу их
ческой памяти у куриных эмбрионов, которая усиливает
печени.
устойчивость цыплят к низким или высоким температурам
В нашем исследовании показано, что эффекты синер-
окружающей среды в постнатальный период. Эпигене-
гетического воздействия сопровождались подавлением
тическая память связана с изменениями гормонального
роста длины и массы тела, массы селезёнки и уровня
профиля, активности и экспрессии генов, которые кон-
метаболизма (по выделению СО2) к периоду вылупления
тролируют систему терморегуляции [17, 18]. Ранее было
(19-е сутки), но повышением массы тела суточных цыплят.
показано, что эпигенетическая адаптация к переменным
Известно, что кортикостерон влияет на функциональную
температурам, формирующаяся в течение эмбриогенеза,
активность соматотропной оси посредством ингиби-
повышает вывод молодняка кур и его качество, по срав-
рующего действия на СТГ, который уменьшает рост тела
нению со стабильным режимом инкубации [19].
путем подавления синтеза ИФР-1 [30]. Следовательно,
Переменные температуры инкубации (1) в эм-
можно предположить, что на фоне воздействия стресс-
бриональном развитии курицы (см. рис. 2) оказывают
факторов (температура и свет) во время искусственной
тренировочный эффект и приводят к изменениям в
инкубации снижается функциональная активность со-
преоптической области передних гипоталамических
матотропной и ГГЩЖ осей посредством ингибирующего
нейронов (PO/AH) (3), что влияет на температурную
действия на СТГ и ТРГ, которые задерживают процессы
чувствительность куриных эмбрионов [18]. Преоптиче-
роста и метаболизма у куриных эмбрионов в антенаталь-
ская зона переднего отдела гипоталамуса (2) участвует
ном онтогенезе.
в регуляции синтеза кортикотропин-рилизинг-гормона
Таким образом, ориентируясь на представленные
(КРГ) (12) от гипоталамо-гипофизарно-надпочечниковой
данные, можно полагать, что механизмы, опосредую-
оси (ГГН-ось) (13) и тиреотропин-рилизинг-гормона (ТРГ)
щие возможные стимулирующие эффекты переменной
(14) от оси гипоталамус-гипофиз-щитовидная железа (ось
температуры и красного светодиодного освещения яиц
ГГЩЖ) (15) гипоталамо-гипофизарной системы. КРГ (12)
во время инкубации на развитие куриных эмбрионов и
стимулирует переднюю долю гипофиза (7), что приводит
их органов, а также морфофизиологические процессы,
к синтезу и секреции адренокортикотропного гормона
связаны с пластическими изменениями в функциональной
(АКТГ) (16), который, в свою очередь, вызывает секрецию
активности гипоталамо-гипофизарной системы в процессе
надпочечниками (17) кортикостерона (18), регулирующего
эмбриогенеза и раннего постнатального онтогенеза. Так,
у эмбрионов кур белковый, углеводный, жировой обмены,
при режиме ПТ+КС развитие эмбрионов кур и их селе-
иммунные реакции и реакции на стресс. КРГ (12) также
зёнки к 19-м суткам проходит медленнее, чем при режиме
играет роль в активации ТРГ (14), который стимулирует
КС и ПТ, соответственно. Кроме того, воздействие ПТ+КС
высвобождение ТТГ (19) гипофизом (7). ТТГ (19), в свою
вызывает оптимизацию метаболизма у эмбрионов на 19-е
очередь, повышает уровень гормонов щитовидной железы
стуки эмбриогенеза и вместе с тем повышает качество
(20), главным образом, гормона Т4 (тироксина) (21) [20].
суточных цыплят, выражающееся в увеличение массы их
Перед тем, как оказать воздействие на клетки органов-
тела на 1,93 г, в сравнении с ПТ.
мишеней (все кроме селезёнки и семенников), большая
часть тироксина непосредственно в клетках конвертиру-
Литература
ется в биологически активную форму - трийодтиронин
1. Chicken Incubation Conditions: Role in Embryo
(Т3) (21). Пролактин (19) регулирует функции щитовидной
Development, Physiology and Adaptation to the Post-
железы (20), обеспечивающие стимуляцию роста куриных
Hatch Environment / K. Tona, K. Voemesse, O. N’nanlé,
эмбрионов, регуляцию у них метаболизма и терморегуля-
цию. Поскольку оси ГГЩЖ и ГГН играют важную роль в
frontiersin.org/articles/10.3389/fphys.2022.895854/
адаптации терморегуляции организма [21, 22], модуляция
full (дата обращения: 04.07.2022). doi: 10.3389/
температуры инкубации во время формирования этих осей
fphys.2022.895854.
в эмбриогенезе может улучшить термотолерантность птиц
2. Yalcin S., Özkan S., Shah T. Incubation Temperature and
и вызывать долгосрочную адаптацию осей гипоталамо-
Lighting: Effect on Embryonic Development, Post-Hatch
гипофизарной системы в постнатальный период развития
Growth, and Adaptive Response // Front. Physiol. 2022.
[23]. Имеющиеся литературные сведения ясно указывают,
что на эмбриональное развитие осей ГГЩЖ и ГГН влияют
fphys.2022.899977/full (дата обращения: 04.07.2022).
изменения температуры во время инкубации, которые
doi: 10.3389/fphys.2022.899977.
формируются между 10,5…11,5 и 14…15 сутками эмбрио-
3. Archer G.S., Mench J.A. The Effects of the Duration
генеза продуктивных птиц соответственно [24, 2].
and Onset of Light Stimulation during Incubation on the
Светодиодное освещение во время инкубации (22)
Behavior, Plasma Melatonin Levels, and Productivity of
активирует фоторецепторы сетчатки глаз (23) и шиш-
Broiler Chickens // J. Anim. Sci. 2014. Vol. 92. Р. 1753-
ковидную железу (эпифиз) (24) - посредников гормона
1758. doi: 10.2527/jas.2013-7129.
мелатонина (25), поступающего в кровь и спинномозговую
4. Rogers L.J. «Environmental Influences on Development of
жидкость (ликвор), пройдя через которую, накапливается
the Embryo» in The Development of Brain and Behaviour
в супрахиазматическом ядре передней области гипотала-
in the Chicken. Wallingford, Oxon: CABI Publishing,
муса (СХЯ) (26) гипоталамо-гипофизарной системы. Ось
1995. Р. 41-70.
55
Российская сельскохозяйственная наука, 2022, № 6
5. Chelnokova M.I. Differential Incubation Temperature
Epigenetic Temperature Adaptation // Comp. Biochem.
Effects on Growth of Hisex Brown Chick Embryos
Physiol. Part A: Mol. Integr. Physiol. 2002. Vol. 131. Р.
and Development of Their Visceral Organs // Russian
825-832. doi: 10.1016/s1095-6433(02)00020-x.
Agricultural Sciences. 2021. Vol. 47(4). Р. 418-424. doi:
19. Челнокова М.И., Сулейманов Ф.И., Челноков А.А.
10.3103/S1068367421040042
Эпигенетическая адаптация эмбрионов кур крос-
6. Челнокова М.И., Сулейманов Ф.И., Челноков А.А. Раз-
са «Ломанн Браун» к переменным температурам
витие и метаболизм эмбрионов курицы в эмбриогенезе
инкубации // Ветеринария. 2022. № 3. С. 65-70. doi:
при разном светодиодном освещении яиц во время
10.30896/0042-4846.2022.25.3.65-69.
инкубации // Иппология и ветеринария. 2021. № 4(42).
20. Decuypere E., Kühn E. R. Thyroid Hormone Physiology
С. 219-224.
in Galliformes: Age and Strain Related Changes in
7. Prenatal Effects of Red and Blue Light on Physiological
Physiological Control // Am. Zool. 1988. Vol. 28. Р. 401-
and Behavioural Parameters of Broiler Chickens / A.
415. doi: 10.1093/icb/28.2.401.
Drozdová, Z. Kaňková, B. Bilčík, et al. // Czech J. Anim. Sci.
21. Heat Stress Responses in Birds: A Review of the Neural
2021. Vol. 66. P. 412-419. doi:10.17221/80/2021- cjas.
Components / M.W. Bohler, V.S. Chowdhury, M.A. Cline,
8. Allometric relationships between embryonic heart rate and
fresh egg mass in birds / Tazawa H., Pearson J.T., Komoro
mdpi.com/2079-7737/10/11/1095 (дата обращения:
T., et al. // J. Exp. Biol. 2001. Vol. 204. Р. 165-174. doi:
04.07.2022). doi: 10.3390/biology10111095
10.1242/jeb.204.1.165.
22. Ruuskanen S., Hsu B.-Y., Nord A. Endocrinology of
9. Болотников А.М. Каменский Ю.Н., Добринский Л.Н.
Thermoregulation in Birds in a Changing Climate // Mol.
Экология раннего онтогенеза птиц. Свердловск: УНЦ
АН СССР, 1985. 228 с.
hub.ru/10.1016/j.mce.2020.111088 (дата обращения:
10. Vleck C.M., Hoyt D.F. Metabolism and energetics of
04.07.2022). doi: 10.1016/j.mce.2020.111088.
reptilian and avian embryos. Deeming C.D., Ferguson
23. Halevy O., Yahav S. The Effect of thermal Manipulations
M.W. (red) Egg incubation: its effects on embryonic
during the Development of the Thyroid and Adrenal
development in birds and reptiles. Cambridge: Cambridge
Axes on in-hatch and post-hatch Thermoregulation /
University Press, 2004. P. 285-304.
Y. Piestun, D. Shinder, M. Ruzal, et al. // J. Therm.
11. Scanes C.G. Perspectives on the Endocrinology of Poultry
Biol. 2008. Vol. 33. Р. 413-418. doi: 10.1016/j.
Growth and Metabolism // General and Comparative
jtherbio.2008.06.007.
Endocrinology. 2009. Vol. 163. P. 24-32. doi: 10.1016/j.
24. de Groef B., Grommen S.V.H., Darras V.M. The Chicken
ygcen.2009.04.013.
Embryo as a Model for Developmental Endocrinology:
12. Effects of Monochromatic Blue Light on Reducing the
Development of the Thyrotropic, Corticotropic, and
Adverse Impact of Induced Cyclic Chronic Heat Stress
Somatotropic Axes // Mol. Cell Endocrinol. 2008. Vol. 293.
during the Thermal Manipulation of Broiler Embryos /
Р.17-24. doi: 10.1016/j.mce.2008.06.002.
L. Zeng, Q. Liu, T. Wang, et al. // Oxid Med Cell Longev.
25. Various LED Wavelengths Affected Myofiber Development
and Satellite Cell Proliferation of Chick Embryos via the
omcl/2022/9898311/ (дата обращения: 04.07.2022). doi:
IGF-1 Signaling Pathway / Y. Wang, X. Bai, Z. Wang, et
10.1155/2022/9898311.
al. // Photochem. Photobiol. 2017. Vol. 93. Р. 1492-1501.
13. Mcmurtry J.P., Francis G.L., Upton Z. Insulin-like Growth
doi: 10.1111/php.12806.
Factors in Poultry // Domestic Animal Endocrinology.
26. Effect of a Photoperiodic green Light Programme
1997. Vol. 14. P. 199-229. doi: 10.1016/S0739-
during Incubation on Embryo Development and hatch
7240(97)00019-2.
Process / Q. Tong, I.M. McGonnell, T.G.M. Demmers,
14. Yakar S., Isaksson O. Regulation of Skeletal Growth and
et al. // Animal. 2018. Vol. 12. Р.765-773. doi: 10.1017/
Mineral Acquisition by the Gh/igf-1 Axis: Lessons from
s1751731117002117.
Mouse Models // Growth Hormone & IGF Research. 2016.
27. The Effects of thermal and Light Exposure on the
Vol. 28. Р. 26-42. doi: 10.1016/j.ghir.2015.09.004.
Development of Broiler Chicken Leg Musculature / S.
15. Piestun Y., Halevy O., Yahav S. Thermal Manipulations
Stojanovic, D. Zikic, Z. Kanacki, et al. // Arch. Biol.
of Broiler Embryos-The Effect on Thermoregulation and
Sci. Belgra. 2014. Vol. 66. Р. 1547-1557. doi: 10.2298/
Development during Embryogenesis // Poult. Sci. 2009.
abs1404547s.
Vol. 88. Р. 2677-2688. doi: 10.3382/ps.2009-00231.
28. Influence of Modified Incubation Factors on Meat
16. Yalcin S., Aksit M., Ozkan S., Hassanzadeh M., Bilgen
Characteristics of Broiler Chickens / Z. Kanački, S.
G., Helva I. B., et al. Effect of Temperature Manipulation
Stojanović, D. Žikić, et al. // Anim. Sci. Pap. Rep. 2017.
during Incubation on Body Weight, Plasma Parameters,
Vol. 1. Р. 87-96. doi: 10.2298/BAH1104605K.
Muscle Histology, and Expression of Myogenic Genes
29. Shah T., Özkan S. Effect of thermal Manipulation and
in Breast Muscle of Embryos and Broiler Chickens from
Photoperiodic Lighting during Incubation on Hatching
Two Commercial Strains / S. Yalcin, M. Aksit, S. Ozkan,
Performance, Hatching Time, Chick Quality and Organ
et al. // Br. Poult. Sci. 2021. Vol. 63. Р. 21-30. doi:
Growth // Ege Univ. Ziraat Fak. Derg. 2022. Vol. 59. Р.
10.1080/00071668.2021.1958297.
17-31. doi: 10.20289/zfdergi.
17. Nichelmann M., Janke O., Tzschentke B. Efficiency of
30. Tolon R., Lopez J., Sanchez-Franco F. Corticosterone
Thermoregulation in Precocial Avian Species during the
Modulates Growth Hormone-releasing Factor and
Prenatal Period // J. Therm. Biol. 2001. Vol. 26. Р. 273-
Somatostatin in Fetal Rat Hypothalamic Cultures /
280. doi: 10.1016/s0306-4565(01)00030-4.
G. Fernandez-Vazquez, L. Cacicedo, M.J. Lorenzo, et
18. Tzschentke B., Basta D. Early Development of Neuronal
al.// Neuroendocrinology. 1995. Vol. 61. Р. 31-35. doi:
Hypothalamic Thermosensitivity in Birds: Influence of
10.1159/000126824.
Поступила в редакцию 19.07.2022
После доработки 29.09.2022
Принята к публикации 08.11.2022
56