Изучение закономерностей распределения особей в пространстве – это один из вопросов фундаментальной экологии (Pianka, 1973; Шилов, 1998). Предполагается, что птицы неслучайно выбирают место для гнездования (Smith et al., 1999; Henrioux, 2002; Forstmeier, Weiss, 2004; Blakey et al., 2019). Выбор подходящего гнездового местообитания является ключевым фактором будущего успешного гнездования птицы. Важную роль на ландшафтном уровне играют тип растительности и гетерогенность местообитаний. Гнездо и его ближайшее окружение обеспечивают скрытость и защиту гнезда от хищников (Lack, Newton, 1981; Frommhold et al., 2019).
Неравномерное распределение условий на территории обитания вида определяет агрегированное, равномерное или случайное распределение особей локальной популяции (Pianka, 1973). По значимости и силе влияния факторы разделяют на основные и уточняющие. Основные факторы определяют вероятность выживания особей (защита от хищников и непогоды, наличие корма), а уточняющие влияют на выбор гнездового участка (например, ландшафт, рельеф, структура биотопов). Различия в соотношении характеристик местообитаний определяют их качество для птиц и, соответственно, порядок занятия гнездовых участков особями. Предполагается, что сначала особи занимают оптимальные местообитания с подходящими факторами среды, потом – второстепенные местообитания. Этот процесс описан как поэтапное заселение мест обитания (Brown, 1969; Tome, 2003). Структура местообитаний при выборе места гнездования будет положительно влиять на параметры размножения (Green, Anthony, 1989; Rodríguez et al., 2006; Sumasgutner et al., 2019).
Изучение влияния качества местообитаний на выбор гнездовых участков хищных птиц, прежде всего, связано c описанием структуры растительности на гнездовом участке. Эти параметры часто описывают качественно, но это затрудняет обоснованное выделение наиболее значимых факторов, влияющих на заселение, а также редко оценивается изменение структуры местообитаний во времени (Lack, Newton, 1981; LaHaye et al., 1997; Penteriani et al., 2001). Современный метод анализа для описания местообитаний использует геоинформационные системы и дистанционное зондирование (Skov, 2000; Foody, 2008). Такой метод предоставляет разнообразную пространственную информацию для анализа растительности и ее изменение с течением времени, поэтому дает широкие возможности для оценки динамики пространственного размещения гнездящихся особей, структуры гнездовых местообитаний и ее долговременной динамики (Murali et al., 1998; Efe et al., 2012).
Долговременные исследования дают возможность изучать динамику местообитаний и ее влияние на репродуктивные параметры, биотопическое и пространственное распределение птиц, однако до сих пор эта область сравнительно малоизучена. Большинство работ по близким темам посвящены, как правило, исследованию влияния структуры местообитаний на биоразнообразие в целом или оценке зависимости репродуктивного успеха отдельных видов от структуры местообитаний на коротком промежутке наблюдений – до 5 лет (Henrioux, 2002; Benton et al., 2003; Moreno-Mateos et al., 2011).
Ушастая сова (Asio otus (Linnaeus 1758)) – обычный вид, широко распространенный в Евразии и Северной Америке. В южных частях ареала птица оседлая, а на большей части области распространения является мигрантом или кочует (Cramp, 1983; Приклонский, 1993). В центральных районах России, в том числе на изучаемой площадке, ушастые совы возвращаются с зимовок на места размножения в конце марта-начале апреля и занимают гнездовые участки не случайно, а ориентируясь на определенные параметры местообитаний (Marks, 1986; Henrioux, 2000). Оптимальность местообитаний влияет на успех размножения и гнездования сов (Green, Anthony, 1989; Rodríguez et al., 2006). Низкий успех гнездования кроличьего сыча (Athene cunicularia) автор связывает с выбором неподходящего местообитания, из-за чего птицы селились близко друг к другу и конкурировали за корм (Green, Anthony, 1989). А наличие плотного кустарника положительно коррелировало с количеством слетков ушастой совы в Испании, предположительно из-за уменьшения риска хищничества (Rodríguez et al., 2006).
Цель данной работы – выявление оптимальных местообитаний (структура, изменения во времени) для успешного гнездования ушастой совы.
МАТЕРИАЛ И МЕТОДЫ
Исследование было проведено в сети заказников “Журавлиная родина” на севере Московской обл. (56.7553 N, 37.7974 E) в 2001–2020 гг. Площадь модельной территории 48 км2. Она представляет собой использующиеся и заброшенные сельскохозяйственные угодья, с закустаренными в различной степени местообитаниями, окружена лесами и включает несколько деревень с садово-огородными участками, в которых люди проживают в летний период. За время исследования на модельной территории произошли существенные изменения структуры растительного покрова из-за прекращения сельскохозяйственной деятельности на обширных территориях и зарастания их кустарником и лесом (Свиридова и др., 2006, 2016; Volkov, Sviridova, 2013).
Картирование территориальных пар ушастой совы, поиск гнезд и выводков проводили с марта по июль. Для найденных гнезд были определены различные параметры: тип гнезда (вид птицы, чье гнездо занято совой), наличие крыши (для сорочьих гнезд), вид дерева и высота расположения гнезда над землей. За 20 лет было найдено 145 гнезд. За гнездовой участок принималась окружность радиусом 500 м с гнездом в центре. Успешными считали гнезда, которые покинул хотя бы один слеток.
Описание структуры гнездового местообитания анализировали по многоканальным спутниковым снимкам Landsat 4–5, 7 и 8. Снимки получены с доступного сайта Геологической службы США “USGS” (USGS: U.S. Geological Survey, 2019). Для каждого года был выбран один снимок, относящийся к началу гнездового периода ушастой совы (апрель) – времени прилета и окончательного формирования территориальной структуры на исследовательской площадке. Все пространственные данные обрабатывали в программе “QGIS” v. 3.14 (QGIS: Свободная географическая информационная система с открытым кодом, 2020) в модуле “Semi-Automatic Classiffication Plugin” v. 7 (Congedo, 2014).
Для каждого типа спутника, в зависимости от его характеристик, выбирали определенный набор каналов и длину волны для распознавания программой правильных спектральных характеристик. Каналы снимка обрабатывали функцией “Atmospheric correction DOS1” для преобразования исходных значений яркости (DN) в значения коэффициента отражения. Затем использовали функцию повышения качества снимка “Pansharpening” для Landsat 7 и 8. Каналы космоснимка обрезали по площади исследовательской площадки и соединяли в одно растровое изображение. В качестве цветного композита изображения выбран RGB = 432 (зеленый, красный и ближний инфракрасный каналы) (Congedo, 2014).
Каждый тип местообитания определяли как отдельный макрокласс в модуле. Для них создавали тренировочный полигон – набор обучающих пикселей или ROI – как наиболее типичную площадь растительности с использованием вегетационного индекса NDVI. Всего на территории было выделено пять типов местообитаний, наиболее точно описывающих соответственно пять типов местообитаний на территории гнездового участка. К первому типу относятся сплошные лесные массивы, колки и небольшие по площади участки леса на заброшенных сельскохозяйственных полях (далее – лес); ко второму – зарастающие кустами поля, лесополосы вдоль канав и ивняки (далее – кустарники); к третьему – сенокосные, залежные луга, а также непахотные луга (далее – луга); к четвертому типу – пашни. Отдельно вручную были обрисованы населенные пункты с постоянным населением до 50 человек, которые составили пятый тип местообитания (далее – деревни). Кроме того, классификацией были выделены “ошибки космоснимка”, к которым относились непосредственно ошибки съемки земной поверхности, облака и их тени. Этот параметр в анализе не использовался.
Для классификации растительности на основании тренировочных полигонов был использован алгоритм классификации “Land Cover Signature Classification”. Для неклассифицированных данных использовали дополнительный алгоритм классификации “Spectral Angle Mapping” с пороговым значением 0.0. На выходе получали один растровый слой, содержащий все 5 типов местообитаний. Точность классификации оценивали функцией “Accuracy” с вариациями от 83 до 100%. Классификацию считали точной при значении от 80% (Congedo, 2014).
Для того чтобы генерализовать несущественно малые площади и ошибки классификации, полученный растровый файл преобразовывали, с использованием алгоритма “Classification sieve” с объединением пикселей по 4 и пороговым значением 12. Далее результат конвертировали в векторный формат и преобразовывали инструментом программы QGIS “Исправление геометрии” для возможности получения пространственных данных.
Для каждого гнездового участка определяли суммарную площадь всех типов местообитаний, а с помощью коэффициента ландшафтной неоднородности (формула 1, по: Виноградов, 1998) оценивали разнообразие структуры местообитаний.
(1)
$K = \frac{m}{{m - 1}}\left[ {1 - \sum\limits_{i{\kern 1pt} - {\kern 1pt} 1}^n {\mathop {\left( {\frac{{{{S}_{i}}}}{S}} \right)}\nolimits^2 } } \right],$Также было качественно оценено наличие открытой воды на гнездовом участке (есть или нет) и пересечение его территории с асфальтированным шоссе или грунтовыми дорогами для сельскохозяйственной техники.
Анализ влияния типов местообитания был проведен в программе R версия 3.6.3 (The R Project, 2020), с использованием модулей “gbm” версия 2.1.8 и “dismo” версия 1.3-3 с применением алгоритма машинного обучения дерева решений (“Boosted Tree Classifier”) (Ridgeway, 2020). Подход, который мы использовали, предполагает, что отношения между зависимой переменной и предикторами являются сложными, и определяет относительную важность различных предикторов, включая нелинейные и интерактивные связи (Elith et al., 2008). В качестве зависимой переменной использовали качественное значение успеха гнездования (хотя бы один слеток). В качестве независимых переменных выступали: процент площади пяти типов местообитаний на гнездовом участке, коэффициент ландшафтной неоднородности, наличие открытой воды и тип дорог на гнездовом участке, высота гнезда над землей и наличие или отсутствие крыши. Тренд многолетних изменений структуры местообитания на гнездовом участке оценивали по пятилетним периодам с помощью коэффициента корреляции Кендалла (Tk). В каждый период входил один пиковый год, когда на территории наблюдали наибольшее число гнезд ушастой совы. Тип пространственного распределения гнезд определяли методом “ближайшего соседа” (R), так как данный метод намного точнее стандартных (Clark, Evans, 1954; Downey, 2006).
РЕЗУЛЬТАТЫ
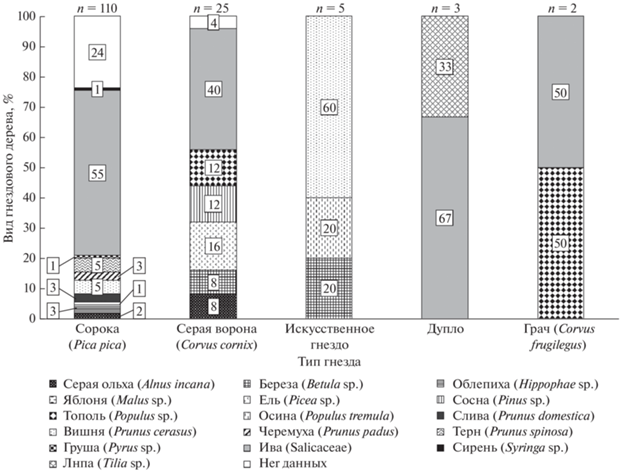
Описание гнездового участка. На территории исследовательской площадки ушастая сова гнездится ежегодно, численность пар колеблется от 1 до 41 пары, в среднем 4 (медиана) и только в 2005 г. сова не гнездилась (Volkov et al., 2009; Sharikov et al., 2010). За период 2001–2020 гг. описано 145 случаев гнездования ушастой совы, 64.1% (93 гнезда) из которых были успешны. Птицы были обнаружены в пяти различных типах гнезд и на 16 видах деревьев. В большинстве случаев гнездование было отмечено в гнезде сороки – 77%, и гнезда в основном располагались на иве (50%) (рис. 1). Из 110 сорочьих гнезд количество целых с крышей составило 35, а гнезд со сломанной крышей 75. Высота гнезда над землей варьировала от 1.5 до 22 м, в среднем 4 м (медиана), в диапазоне 3–5 м (±25% квантиль). Из 23 гнезд, расположенных ниже 3 м, чуть больше половины гнезд – 12 (52%) – были неудачные.
Рис. 1.
Соотношение (%) видов гнездового дерева (цифровые значения в квадратах) и типов гнезд (ось X), занятых ушастой совой для гнездования за весь период исследования. n – общее количество гнезд каждого типа гнезда.
Пространственное распределение гнезд ушастой совы по методу “ближайшего соседа” оказалось агрегированным (R = 0.4, p < 0.005), при этом 59% гнезд были в радиусе 500 м от центра деревни и все гнездовые участки, за исключением одного, пересекались в большей или меньшей степени с дорогами, в основном с грунтовыми. Тот факт, что гнезда здесь располагаются группами и, соответственно, чаще используются, вероятно, указывает на высокую значимость этих местообитаний для ушастой совы.
Основная территория гнездовых участков (n = = 145) за весь период исследования в среднем была занята лугами – 51%. Вторым по представленности на территории местообитанием стали кустарники – 16%. Остальные местообитания были представлены в меньшей степени: пашни (3.7%), деревни (3.5%), лес (3.1%), другое (<0.1%). На территории 114 гнездовых участков (78.6%) находились водоемы; грунтовые дороги пересекали 109 гнездовых участков (75%), а асфальтированные дороги – 35 (24%).
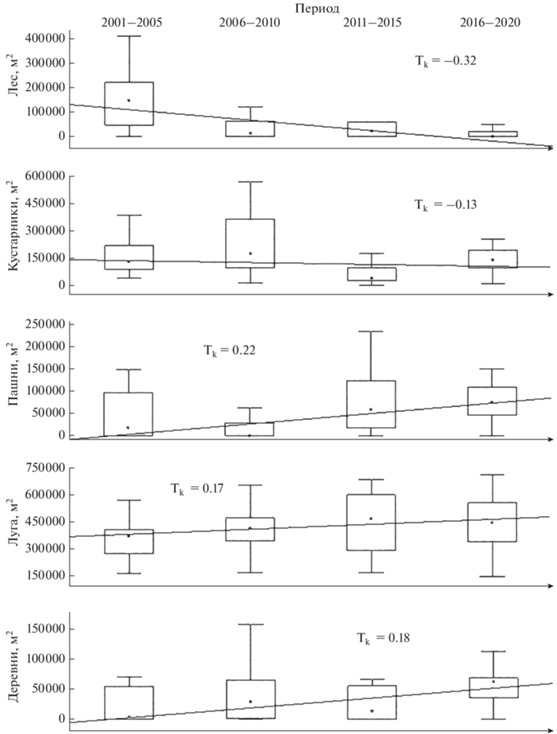
Изменение структуры гнездовых местообитаний с течением времени. За период исследования на гнездовых участках ушастой совы произошло статистически достоверное изменение соотношений местообитаний, а именно: уменьшилась площадь леса и кустарников и увеличилась площадь лугов, деревень и пашен (рис. 2). При этом на всей территории исследовательской площадки наблюдается тенденция к увеличению площади лугов и кустарников, а также уменьшению площади лесов и пашен. Наиболее значимо на гнездовых участках уменьшилась площадь леса и увеличилась площадь деревень.
Рис. 2.
Тренды изменения площади местообитаний на гнездовых участках за период исследования (для всех значений критерия Tk уровень достоверности p < 0.05).
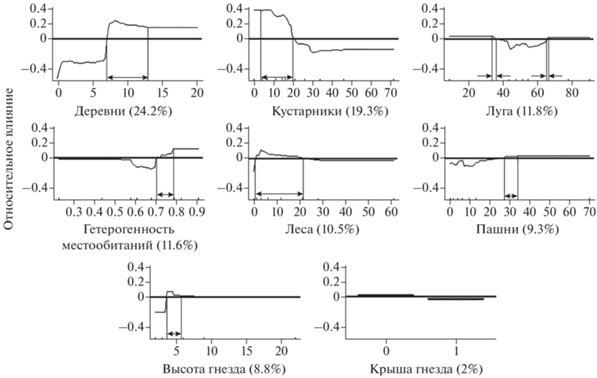
Успех гнездования в зависимости от структуры местообитаний на гнездовом участке. В целом за период исследования устойчивый тренд изменения успеха гнездования ушастой совы не выявлен (Tk = 0.06, p > 0.05). На успех гнездования ушастой совы наибольшее влияние оказывают такие местообитания внутри гнездового участка, как: площадь деревень (24.2%), кустарников (19.3%), лугов (11.8%), гетерогенность местообитаний (11.6%). При этом каждый из параметров местообитаний влиял на успех гнездования ушастой совы в определенном диапазоне значений (рис. 3). Площадь деревень оказывает влияние при представленности на гнездовой территории в диапазоне 7.5–12.5%. При этом максимальная вероятность успешного гнездования наблюдается при значении 8%. Кустарниковые ассоциации обусловливают успешное гнездование при представленности их на территории от 4 до 20%, с максимумом 5–8%; при 20–47% вероятность успешного гнездования снижается. Индекс ландшафтной гетерогенности (K) от 0.55 до 0.72 на гнездовом участке определяет низкую вероятность успеха гнездования, но при значении выше 0.72 – высокую, с максимумом при значении 0.79. Площадь леса в диапазоне 1–25% обусловливает высокий успех гнездования, с максимумом при 4%. Площадь пашен обусловливает высокий успех гнездования при 28–34% представленности их на территории. Наличие лугов в диапазоне 33–35% (с большей вероятностью) и 65–66% (с меньшей вероятностью) положительно влияет на успех гнездования, а при промежуточных значениях вероятность успешного гнездования сильно снижается. Высота расположения гнезда над землей в диапазоне 3–5 м положительно влияет на вероятность высокого успеха гнездования. Наличие крыши у гнезда, наличие дорог или воды на гнездовом участке определяет успех гнездования очень слабо (2%, 0.1 и 0.1% соответственно).
Рис. 3.
Влияние выбранных независимых переменных на успешность гнездования ушастой совы. В скобках – относительный вклад каждой переменной в конечную модель “Boosted regression tree”. Оси Y находятся на логит-шкале, а жирные горизонтальные линии указывают на эффект нуля (над линиями – успешное гнездование, под линиями – неуспешное). Прямая горизонтальная линия на каждом графике указывает на отсутствие данных.
ОБСУЖДЕНИЕ
Репродуктивный успех у птиц большей частью зависит от выживаемости гнезда, поэтому гнездящиеся птицы должны выбирать местообитание, которое обеспечивает их безопасность и достаточное количество кормовых ресурсов для выведения потомства (Frommhold et al., 2019). Таким образом, выбор ушастой совой места для гнездования – это, как правило, компромисс между защищенностью гнезда от потенциальных хищников и обилием кормовых ресурсов на гнездовом участке (Lack, Newton, 1981; Green, Anthony, 1989; Rodríguez et al., 2006; Frommhold et al., 2019).
Наше исследование показало, что в среднем за 20 лет наблюдений наибольшая площадь на гнездовом участке ушастой совы была занята лугами и кустарниками. Также за этот период имеется статистически достоверный тренд к изменению предпочтения местообитаний ушастой совой при выборе гнездового участка: от лесных к более открытым территориям, вплоть до участков с высокой антропогенной нагрузкой. Эту тенденцию отмечают и в других исследованиях, проведенных ранее на изучаемой территории и в Европе (Sharikov et al., 2010; Lövy, Riegert, 2013).
В нашей работе наличие населенного пункта на гнездовой территории ушастой совы в наибольшей степени влияло на вероятность успешного гнездования птицы, при этом площадь населенного пункта составляла 7.5–12.5% от всей площади гнездового участка. Вероятно, причиной является синантропизация сороки. Так как в Европе наблюдается выраженная тенденция к заселению сорокой более населенных людьми территорий, на нашей исследовательской площадке также возможно проявление таких же тенденций у этой птицы. На нашем стационаре в большинстве случаев ушастая сова предпочитала занимать гнезда сороки, что соотносится с данными других авторов (Craig, Trost, 1979; Marks, Yensen, 1980). Предполагается, что ушастая сова стала активно осваивать территории населенных пунктов вслед за сорокой, однако, имея на территории много потенциально пригодных сорочьих гнезд, ушастая сова занимает только определенную долю этих гнезд, то есть потенциальных мест для гнездования всегда избыточное количество (Craig, Trost, 1979; Rodríguez et al., 2006; Sharikov et al., 2010; Lövy, Riegert, 2013). Согласно полученным нами результатам, увеличение вероятности успешного гнездования ушастой совы связано с наличием на гнездовом участке населенного пункта, поэтому можно предположить, что ушастая сова выбирает гнездовой участок, ориентируясь именно на этот фактор.
Более частый выбор ушастой совой для гнездования населенных пунктов можно объяснить повышенной численностью и разнообразием мелких мышевидных грызунов, которых сова использует в пищу, в антропогенных ландшафтах. Так как в сильно фрагментированном антропогенном ландшафте присутствует опушечный эффект (и он вероятно намного выше природного), это может влиять на бо́льшую доступность и разнообразие добычи в период размножения и, соответственно, повышать успех гнездования (Holt, 1997; Henrioux, 2002; Lövy, Riegert, 2013). В то же время в естественных природных сообществах сильно фрагментированный ландшафт вместе с краевым эффектом способствуют увеличению доли разоренных гнезд (Donovan et al., 1997). При этом риск разорения гнезда в населенных пунктах, вероятно, ниже, так как наземные хищники меньше заходят на территорию деревень, о чем свидетельствуют результаты нашего предыдущего исследования на этой же модельной площадке (Sharikov et al., 2010). Гетерогенность антропогенного ландшафта, с одной стороны, в некоторой степени уменьшает угрозу хищничества в деревнях и, с другой стороны, может увеличивать доступность корма за счет опушечного эффекта населенных пунктов, что предположительно привело к увеличению вероятности успешного гнездования ушастой совы в деревнях на нашей исследовательской площадке. И наоборот, снижение успеха гнездования при увеличении лесистости и закустаренности на гнездовых участках обусловлено, вероятно, усилением факторов хищничества и исчезновения охотничьих угодий.
Результат нашей работы показал, что для успешного гнездования ушастой сове необходимы разнообразные местообитания: кустарники и луга – обширные открытые пространства для охоты. По нашим данным, если коэффициент ландшафтной неоднородности высок, то высока и вероятность успешного гнездования, что подтверждается выбором птицей более неоднородного ландшафта. Большие открытые пространства, такие как пашни и луга, несомненно, обеспечивают сову достаточным количеством корма в период размножения, но сова избегает обширных однородных открытых территорий и территорий, на которых отсутствуют кустарники. Как показано зарубежными исследователями (Henrioux, 2000; Benton et al., 2003; Moreno-Mateos et al., 2011; Emin et al., 2018), такие открытые пространства отрицательно влияют на гнездование и не обеспечивают ушастую сову необходимыми для этого условиями, поэтому птицы выбирают более закрытые участки, заросшие кустарником и лесом. Видимо поэтому в нашей работе ушастая сова предпочитала при выборе участки, занятые кустарниками (4–20%).
Кустарники, помимо того, что являются удобным местом для гнездования, вероятно также обеспечивают защиту от хищничества из-за плотной и разветвленной кроны (Glue, 1977; Marks, 1986; Bull et al., 1989; Henrioux, 2002). Выбор такого более защищенного гнездового участка подтверждается также данными зарубежных исследователей о воробьеобразных, где авторы указали, что именно защита от хищничества может быть главным критерием выбора более ветвистого кустарника, чем деревьев (Forstmeier, Weiss, 2004). В ландшафтах, где преобладают хвойные породы, плотность кроны обеспечивает хорошие защитные условия, и тогда ушастая сова предпочитает занимать хвойные деревья (Glue, 1977; Bull et al., 1989; Tome, 2003; Rodríguez et al., 2006). На территории нашего исследования мало хвойных лесов, зато много зарастающих ивой сельскохозяйственных угодий, поэтому функцию более плотного покрова, которое скрывает гнездо, выполняют ивы, то есть кустарниковые местообитания. Вероятно, выбор ивы, как гнездового дерева, связан и с выбором гнезда сороки, так как большинство сорочьих гнезд, занятых совой, располагалось на иве. Возможно, сочетание плотной кроны кустарника и сорочьего гнезда, которое, как известно имеет крышу, помогает избегать разорения гнезда хищниками. Однако данное предположение нашими исследованиями не подтвердилось, так как из всех сорочьих гнезд количество построек со сломанной крышей составило 68%, а также не обнаружилось достоверного различия в вероятности успеха гнездования между гнездами с крышей и без. Это соответствует результатам исследования в США, в которой ушастая сова занимала 67% сорочьих гнезд со сломанной крышей (Marks, 1986).
Высота расположения гнезда над поверхностью земли в нашем случае оказалась значимым параметром для успеха гнездования. На высоте 3–5 м вероятность успешного гнездования была самой большой, поэтому гнезда, расположенные выше или ниже, вероятно, чаще разоряют хищники (Rodríguez et al., 2006). Сходные результаты были получены ранее в США, при высоте ниже 3 м все попытки гнездования ушастой совы были неудачные (Marks, 1986), однако на нашей исследовательской площадке всего лишь половина гнезд, расположенных ниже 3 м, были неудачные. В целом, влияние на вероятность успеха гнездования таких параметров, как высота гнезда и наличие крыши, было самым незначительным, в отличие от влияния наличия на гнездовом участке определенных типов местообитаний (населенные пункты, кустарники и луга). Согласно данным, полученным за 20-летний период, для ушастой совы характерно групповое расположение гнезд и преимущественно на территории деревень, что также может подчеркивать важность антропогенных местообитаний за счет того, что они чаще используются и свидетельствовать о неслучайном их выборе птицами для гнездования.
Наличие водоемов оказалось не значимым фактором для успешного гнездования ушастой совы, хотя некоторые авторы отмечают положительное влияние водоемов на этот вид в засушливых ландшафтах (Moreno-Mateos et al., 2011). Однако в умеренном поясе на исследовательской территории наличие воды (при густой сети водоемов) стало незначительным параметром. Наличие дорог также не оказалось значимым фактором в нашей работе, как и различия между наличием на гнездовом участке асфальтированных дорог с достаточно интенсивным движением или грунтовых дорог с практически нулевой интенсивностью движения. Это также не соответствует исследованию из Испании, в котором ушастая сова реже занимала участки с асфальтированными дорогами на них (Martínez, Zuberogoitia, 2004).
На территории нашей модельной площадки за 20-летний период прослеживается тенденция к зарастанию сельскохозяйственных полей из-за упадка сельского хозяйства. Вероятно, это приводит к снижению площади охотничьих участков, увеличению гетерогенности территории и, возможно, к возрастанию пресса хищничества. За тот же период ушастая сова проявила отчетливую тенденцию к перемещению из лесистых участков в более мозаичные антропогенные. Населенные пункты обладают высоким краевым эффектом, что приводит к увеличению разнообразия добычи, а близость человеческого жилья, вероятно, приводит к уменьшению пресса хищничества со стороны диких животных. В совокупности эти факторы обусловливают выбор ушастой совой определенного оптимального местообитания с гетерогенным, преимущественно антропогенным ландшафтом, который обеспечивает ее повышенной вероятностью успешного размножения. Вероятно, именно структура гнездового местообитания, а не параметры микроместообитания (тип гнезда, вид гнездового дерева, высота гнезда над землей), в первую очередь определяет успех гнездования птицы на изучаемой территории.