Фаунистические исследования в России традиционно составляют значительный пласт зоологических работ. Такая тенденция понятна, учитывая огромную территорию страны. Кроме исследовательского интереса, долгое время фаунистика представляла интерес и для обеспечения хозяйственных нужд человека. Не будет преувеличением сказать, что современная российская территория сформировалась в процессе уточнения ареала соболя – высокая цена на пушнину XVI–XVII веков стала причиной поиска русскими землепроходцами новых территорий, способных обеспечить казну “мягкой рухлядью” (Кречмар, 2021). После утраты интереса к пушнине, фаунистика получила новый толчок в процессе формирования охотничьего хозяйства (Колосов, 1938; Томилин, 1953), а затем в процессе эпизоотологических исследований (Коренберг, 1979; Егоров и др., 2014; Корзиков и др., 2019). Интенсивность фаунистических исследований, поддерживаемая академическим интересом, не снизилась и позднее.
Какие задачи стоят перед фаунистикой? Очевидно, что базовая ее задача – это сбор информации для описания ареалов животных. Успех сбора фаунистической информации зависит от трех компонент (без учета очень важной в фаунистике квалификации исследователя): выбор места проведения исследования (перспективный участок на карте), выбор точек для сбора информации на местности (расположение ловушек в конкретных биотопах или на их стыке, позволяющее решить задачи по сбору информации по конкретным видам), выбор методов сбора информации (отловы разными типами ловушек, маршрутные или стационарные наблюдения, установка фотоловушек, ловушек для сбора волос, ДНК-проб и многие другие). Методам сбора фаунистической информации посвящена обширная литература (Новиков, 1949; Приклонский, 1973; Pocock, Jennings, 2006; Hinlo et al., 2017; Шефтель, 2018; Подольский и др., 2019). Выбор конкретных локаций на местности во многом зависит от полевого опыта исследователя и его умения воспринимать публикации по экологии интересующих его видов. Эта работа посвящена в большей степени выбору места проведения фаунистического исследования.
Как и в случае любых пространственных исследований (Wang et al., 2012), точки сбора информации можно выбирать случайно, по ячейкам регулярных сетей или располагать их в ключевых участках в рамках некой концепции описания ареала (здесь и далее слово “ареал” используется как краткий синоним области распространения). Естественно, в последнем случае выбор точек исследования должен быть полностью определяем теоретической концепцией, т.е. фаунистическое исследование оказывается зависимым от зоо- или биогеографической теории.
Поэтому здесь нельзя обойти стороной смысловую нагрузку понятия ареал. Согласно классическому определению, переходящему из публикации в публикацию уже более полутора веков (Wallace, 1876; Beddard, 1895; Гептнер, 1936; Пузанов, 1938; Абдурахманов и др., 2003; Лисовский, Оболенская, 2014), ареал – это область распространения вида, заселенная не сплошь, а лишь в пределах подходящих биотопов. Это определение блестяще обходит все основные теоретические проблемы выявления области распространения вида. Таких проблем существует как минимум две. Первая из них – это динамика ареалов во времени. Ареал может пульсировать в результате разного наполнения субоптимальных местообитаний в процессе естественной динамики численности. Он может смещаться как следствие изменения природных ландшафтов: локально (пожары, вырубки, сукцессии и т.п.) или глобально (изменение ландшафтов вследствие климатических флуктуаций). Наконец, ареал может изменяться в результате сдвига экологической ниши, занимаемой видом. Поэтому любые ареалогические построения должны сопровождаться указанием временно́го периода, в котором эти построения имеют смысл.
Вторая проблема – это дискретность понятия “обитает” в отличие от реально существующего в природе явления “появляется с определенной вероятностью”. Свободно передвигающиеся организмы не находятся в какой-либо точке пространства постоянно. Участок пространства может входить в экологический оптимум или пессимум вида, в индивидуальный участок животного, может быть местонахождением основного убежища, может посещаться во время миграций, может быть местом сезонного скопления особей или, наоборот, объектом избегания, хотя участок находится в центре ареала.
Таким образом, ареал – это трудно познаваемая динамичная сущность. Он представляет собой пульсирующую во времени картину пространственного распределения вероятности обитания вида; такая картина может оставаться стабильной до тех пор, пока стабильны заселенные видом ландшафты и занимаемая видом экологическая ниша (Лисовский и др., 2020). Задача познания ареала дополнительно усложняется невозможностью прямой оценки вероятности обитания, предсказываемой либо путем оценки вероятности встреч, либо путем оценки экологической емкости биотопов (Guillera-Arroita et al., 2015). Сегодня основным и, по всей видимости, оптимальным способом познания ареалов является карта. Карта позволяет видеть характер пространственного распределения явлений, однако любая карта редуцирует реальность до некоторой модели, заложенной в карту ее автором (Емельянова, Огуреева, 2006; Лисовский и др., 2020). Доминирующая модель отображения ареала неизбежно влияет на работу фауниста, подталкивая его к выбору тех или иных мест исследования.
Выбор мест исследования и смена ареалогических концепций
Как осуществляется выбор точек исследования на практике? В XVIII–XIX веках выбор точек был скорее случайным. Фаунистическая информация собиралась руками путешественников или специалистов, маршруты которых были больше связаны с политическими или географическими интересами (Н. Пржевальский, А. Миддендорф, Р. Маак). Специальные зоологические экспедиции были редки, но и в этом случае их маршруты определялись существующими дорогами, соображениями безопасности и т.п. (П. Паллас, А. Романов, Г. Радде). Фаунистической информации в тот период было еще мало, поэтому каждая новая [случайная] находка поступательно увеличивала наши знания об ареале видов. Очертания ареалов складывались из крупных региональных блоков. При этом до второй четверти XX века зоологов больше интересовали общие принципы неоднородности фауны (зоогеографическое районирование), чем ареалы отдельных видов (Житков, 1937).
Во второй четверти XX века задачи описания видовых ареалов начали выходить на первый план, в публикациях появились карты ареалов (Огнев, 1928, 1931). При этом впервые исследователи начали наносить точки находок животных на карту (Шнитников, 1936; Бобринский и др., 1944; Огнев, 1948). Это позволило сформировать не высказанную явно, но интуитивно очевидную гипотезу – вид живет “где-то там, между точками на карте”. Здесь надо заметить, что представление о неоднородности ареала существовало давно. Это неудивительно, ведь уже первобытные охотники не могли не понимать, что искать нужного зверя следует не “к востоку от Волги”, а в лесах или степях определенного типа. Исследователи также прекрасно понимали, что ареал не сплошь заселен видом, а вид живет в свойственных ему ландшафтах внутри некой географической области (Wallace, 1876; Beddard, 1895; Гептнер, 1936; Пузанов, 1938). Промптов (1934) называет эту неоднородность на локальном уровне кружевом ареала; Гептнер (1936) – топографией вида. Карты ареалов (региональный или глобальный уровень) и знания о биотопической приуроченности видов (локальный уровень) существовали параллельно. Этому было две причины. Первая – еще оставались неизученными крупные участки земной поверхности, поэтому переходить на локальный уровень описания ареалов было рано. Вторая – отсутствовал инструмент для познания и описания ареала на локальном уровне. Крупномасштабные ландшафтные карты отсутствовали (Солодянкина и др., 2021), да и анализ крупномасштабных карт на территорию площадью более миллиона квадратных километров был технически очень сложен. Публикация карт ареалов, учитывающих локальный уровень, была невозможна – достаточно представить пачку карт СССР даже масштаба 1 : 500 000 с ареалом каждого вида. В качестве дополнительной причины можно упомянуть, что фаунистические, ареалогические и экологические работы чаще всего выполнялись разными людьми.
Ко второй половине XX века региональная изученность России и сопредельных территорий сильно возросла. Общие очертания большинства ареалов видов млекопитающих стали более или менее ясны, начался этап локального уточнения распространения. Значительное внимание было уделено и выявлению кружева ареалов, путем познания связи распространения видов с ландшафтами и их картированию (Тупикова, Комарова, 1979). Целый ряд региональных фаунистических сводок был опубликован в этот период (Лаптев, 1958; Верещагин, 1959; Тавровский и др., 1971; Воронов, 1974; Юдин и др., 1976, 1979; Швецов, 1977; Кривошеев, 1984; Чернявский, 1984; Ревин, 1989).
Надо сказать, что подавляющее большинство фаунистических публикаций не содержит описания принципа выбора места исследования. Видимо, выбор был в достаточной степени случайным или был определен иными, не зависящими от цели познания фауны интересами. Зоолог мог оказаться в каком-то регионе и попутно собрать фаунистическую информацию; регион мог заинтересовать его как таковой (расположением, ландшафтами), а фаунистическое исследование играло вторичную роль в выборе места; место исследования было выбрано из соображений транспортной доступности внутри некоего условного региона; фаунистическое исследование было выполнено попутно с другими зоологическими работами. Наконец, фаунист мог работать по четкому плану, который казался ему настолько очевидным, что он не нашел нужным указать его в публикации. На другой стороне такого подхода лежат региональные фаунистические сводки, в которых выбор точек исследования определен полнотой обследования региона.
Несмотря на большой объем прилагаемых в XX веке усилий, вопрос о подходах к выбору точек для фаунистического исследования так и не был поставлен. В методических пособиях описаны любые этапы исследования от отлова и правил формирования полевого журнала до подходов к картографированию ареалов (Новиков, 1949; Тупикова, Комарова, 1979; Емельянова, Огуреева, 2006; Нумеров и др., 2010; Тимошкина, 2012), но принцип выбора места оставался интуитивным. В то же время, сколько нужно точек подтверждения присутствия вида в большом регионе, чтобы считать, что он полностью заселен этим видом? Насколько детально должен быть обследован каждый ландшафт? Да и нужно ли обследовать каждый ландшафт, если проводится исследование распространения одного вида? Мне не удалось найти ответы на эти вопросы в териологической литературе.
Отсутствие постановки вопроса о географическом распределении и плотности точек регистрации вида для возможности формулирования аргументированных выводов о его распространении вполне естественно отражает качественный характер ареалогических исследований. Выбор точек для сбора информации осуществляется интуитивно, построение карты ареала также полностью зависит от понимания автором особенностей распространения. В ареалогии не было гипотез, доказательств, достоверностей, поэтому не было и количественной оценки исходных фаунистических данных.
По сути основной концепцией ареала на протяжении XX века оставалось упомянутое выше “живет между точками на карте”. Соответственно, с точки зрения фаунистики, для уточнения информации об ареале вида можно было исследовать территорию “между точками” или по краю воображаемого ареала. Каждая новая точка повышала изученность ареала. Подробность фаунистической информации определялась возможностью нанесения условных символов в мелкомасштабных картах публикаций (Верещагин, 1959).
Способы описания ареала
До XIX века целостные описания ареалов были редкостью в силу слабой фаунистической изученности территорий. Описания распространения были очень общими или очень частными и приводились чаще всего среди прочих признаков в общих систематических списках (Erxleben, 1777; Gmelin, 1788). В XIX и начале XX веков описание ареала представляло собой длинный список регионов; степень подробности и упорядоченность этого списка полностью зависели от понимания автором географии территории и характера распространения вида (Симашко, 1851). В свою очередь, восприятие читателя также зависело от знания географии (может быть даже в большей степени, чем у автора – ведь читатель был обязан знать все топонимы, упоминаемые в публикации) и внутреннего понимания локальных деталей распространения. Таким образом, путь от распространения животного до понимания ареала широким кругом зоологов был четырехчленным: распространение–случайная выборка точек находок–степень знания автором публикации региональной географии–степень знания читателем региональной географии.
Со второй четверти XX века начали применять картографическое отображение ареалов, в т.ч. с указанием мест находок видов символами на карте (Шнитников, 1936; Бобринский и др., 1944; Огнев, 1948). Это резко понизило потерю информации в четырехчленном пути понимания ареала. Ведь теперь из этого пути выпадали региональные обобщения автора публикации: читатель стал волен сам решить, живет зверь “на Алтае” или всего лишь в районе Бийска. Схема понимания стала выглядеть примерно следующим образом: распространение–случайная выборка точек находок–точность отображения автором точек находок–способность читателя воспринять ареал по точкам. Последний пункт, несомненно, очень сложен и является причиной индивидуального восприятия ареала у каждого исследователя. Тем не менее картографическое отображение позволило перейти от догадок на региональном уровне к догадкам на локальном уровне.
В дальнейшем способы описания ареала стали развиваться несколькими независимыми путями. С одной стороны, в публикациях, особенно обобщающих, активно использовали карты ареалов, изображенных способом замкнутых полей. С точки зрения развития ареалогии и фаунистики, видимо, это можно расценивать как регресс. С одной стороны, это возвращало процесс познания ареалов на стадию восприятия глазами автора. Исходные данные опять стали неясны, а карты рисовались широкими мазками. С другой стороны, замкнутое поле психологически воспринимается как нечто завершенное, не вызывающее желания посмотреть, “что там между точками”. Побочным эффектом этого способа отображения стала иллюзия существования границ ареалов; уточнению этих “границ” посвящено множество публикаций.
Другие направления зоологической картографии продолжали развитие способов описания ареалов в крупном масштабе (Тупикова, Комарова, 1979). Основная теоретическая проблема, с которой столкнулись исследователи на предыдущем этапе, это произвол восприятия ареала пользователем при чтении карты. Эта проблема успешно решается добавлением на карту сопроводительной информации, например физико-географической, которая позволяет привязать внимание читающего к определенным, важным для распространения конкретного вида объектам (Lissovsky et al., 2017), но на практике такие карты встречаются нечасто.
Другая, техническая, проблема состояла в том, что бумажные карты всегда имеют слишком мелкий масштаб для отображения кружева ареала на локальном уровне. Увеличение масштаба карт позволяло отразить более мелкие детали, но при этом неизбежно сокращало размеры отображаемой территории (в рамках того же бюджета). В качестве логического финала этого подхода можно привести работу Реймерса (1966), схемы в которой содержат исчерпывающую информацию о распространении видов в исследованном регионе. Но размер этого региона при этом настолько мал, что очень сложно связать опубликованную информацию с ареалом в целом. Другим способом вместить максимум информации в ареал на бумаге оказались кадастровые карты. Этот способ весьма трудоемок, кадастровые карты не очень хорошо читаются, наверное, поэтому таких публикаций относительно немного (Рыбакова, 2007; Емельянова, Оботуров, 2018).
Отдельное направление составили попытки изобразить на карте структуру ареала, т.е. не только “границы” распространения видов, но и оптимумы, пессимумы и локальные разрывы (Тупикова, Комарова, 1979; Емельянова, 2018). Очевидно, что идея создания такого рода карт является большим шагом вперед. Но фактическая их реализация натолкнулась на необходимость сбора огромных массивов данных в течение длительного времени. Очевидно, что такая работа выполнима, как показывает пример создания крупномасштабных геологических и геоморфологических карт. Но бюджеты геологии и биогеографии несопоставимы, как несопоставимы число биологических видов и число факторов, интересующих практическую геологию.
Подытоживая этот раздел, можно констатировать, что использование карт для описания ареалов позволило оптимизировать как само понимание распространения животных, так и определение зон поиска для фаунистов. В то же время, бумажные карты ареалов на практике всегда имеют слишком мелкий масштаб для детального исследования, а отображение реальной структуры ареала, т.е. географического распределения плотности особей, требует неоправданных (финансовыми бюджетами биогеографии) затрат человеческих ресурсов.
Способы публикации фаунистической информации и их последствия
Объемы сбора и принципы представления фаунистической информации постепенно менялись во времени. В XVIII веке информация по фауне России собиралась в процессе единичных экспедиций. Информацию о местах находок видов можно было получить из опубликованных описаний маршрутов (Паллас, 1788); сохранность добытых экземпляров, по всей видимости, еще не представлялась чем-то особенно важным. В XIX и начале XX веков фаунистика стала в каком-то смысле “музейной” наукой. Публикация новых мест находок млекопитающих сопровождалась указанием музейных номеров подтверждающих экземпляров. Поэтому фаунистические публикации появлялись не после сбора соответствующих экземпляров, а после их поступления в тот или иной музей; автором такой публикации чаще всего становился куратор музейной коллекции, способный уследить за всей картиной распространения видов и корректно определить видовую принадлежность каждого экземпляра. Таким образом, музеи стали основными носителями фаунистической информации. Новые публикации обсуждали в первую очередь музейные экземпляры, упомянутые в предыдущих публикациях, а уж затем точку зрения авторов этих публикаций на определение видов или места их находок.
В советский период, в результате заметного увеличения числа научных работников, ситуация сильно изменилась. Многие специалисты перестали передавать свои сборы в музеи, публикуя вместо этого списки обнаруженных и определенных ими самими животных. По мере своего развития такой подход, как ни странно, стал сильно сдерживать изучение ареалов, несмотря на кажущееся увеличение объема фаунистической информации. Основной причиной замедления изучения ареалов на фоне роста объема такой информации, несомненно, можно считать дефицит специалистов и ограниченность человеческих возможностей. Ведь для построения ареала специалист должен обобщить максимально возможный объем имеющейся информации. По мере роста числа находок видов обработать их становилось все сложнее. Однако эта проблема не касалась музейных сборов. Музейные коллекции упорядочены, прирастают во времени в одном направлении, поэтому для использования новых данных достаточно просмотреть каталоги за последний период. С момента перехода фаунистики от музейной основы к публикационной основе временны́е затраты специалиста стали расти экспоненциально. Теперь вместо посещения нескольких музеев специалисту по фауне стало необходимо собирать множество разрозненных публикаций. Именно разрозненных, поскольку далеко не все фаунистические работы содержат в списке литературы все предшествующие исследования. Процесс перестал быть однонаправленным во времени, поскольку статьи стали не только публиковаться вновь, но и стало возможным “случайно” найти старые публикации с нужной информацией. Кроме того, видовая идентификация в публикациях имеет право вызвать сомнение у специалиста. Ситуация усугублялась тем, что редколлегии журналов не были способны справиться с потоком фаунистической информации, к тому же возникло понимание низкой эффективности публикации сырого фаунистического материала. В результате фаунистические работы переместились в сборники, где их стало еще сложнее искать. В итоге задача сбора актуальной информации по распространению того или иного вида превратилась в кошмар неизвестной продолжительности и непоследовательных, разрозненных поисковых действий. При отсутствии единого хранилища данных следующему специалисту приходилось начинать весь путь поиска информации сначала.
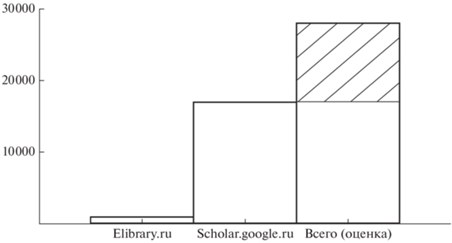
Сейчас нельзя сказать уверенно, сколько фаунистических публикаций было посвящено территории России. Оцифровка текстов научных библиотек пока не достигла того уровня, когда это можно было бы прямо посчитать. Можно попробовать косвенно оценить эти цифры (рис. 1). В базе данных https://elibrary.ru содержится 871 публикация со словами фауна и млекопитающие (палеонтологические работы были отфильтрованы). Это работы, опубликованные в индексируемых elibrary журналах, или работы, ссылки на которые были приведены в индексируемых журналах. База данных https://scholar.google.ru содержит 17 000 териофаунистических работ. Это число включает статьи в широком спектре изданий, в т.ч. процитированные в разнообразных сборниках. Несомненно, существуют и не цитировавшиеся работы. Для оценки их доли я посчитал долю статей, которые отсутствуют в https://scholar.google.ru и которые были опубликованы в нескольких фаунистических сборниках и одном библиографическом списке (Олькова, Башанов, 1970; Воронов и др., 1970, 1971, 1972, 1975, 1976; Бромлей, Костенко, 1976; Юдин, 1977; Контримавичус, 1978). Цифра составила 40%. Получается, что из общего предполагаемого числа фаунистических работ 40% не используются и только около 3% используются в журнальных статьях.
Подробность географической привязки данных
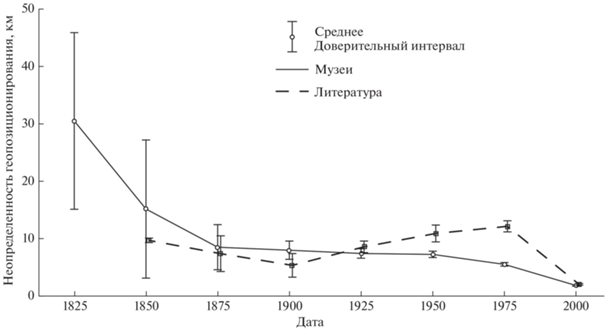
Подробность географической привязки мест обнаружения животных также менялась во времени, отражая основные тренды в фаунистике. Несомненно, подробность описания места регистрации вида сильно зависит от квалификации специалиста, но существуют и некоторые общие тенденции. Если в начале изучения фауны исследователю было достаточно приблизительно знать регион обитания животного, то по мере накопления данных, музейные этикетки становились все более точными (рис. 2): на них появлялись названия рек, хребтов, поселков; направления и расстояния до них. В последние десятилетия в результате активного использования ГИС-технологий в фаунистике, географическая привязка точки регистрации животного стала включать географические координаты. Возрастание неопределенности географической привязки в фаунистических публикациях во второй половине XX века связано с обилием региональных фаунистических сводок, содержащих очень генерализованные указания на места обитания животных.
Рис. 2.
Изменение неопределенности геопозиционирования мест находок млекопитающих за последние 200 лет по данным https://rusmam.ru. Неопределенность геопозиционирования рассчитывали как радиус круга, в котором помещаются все возможные варианты нахождения места, описанного на музейной этикетке или в литературе.
Подытоживая историческую часть статьи, нужно резюмировать основные проблемы, с которыми столкнулась фаунистика в докомпьютерное время. Одной из проблем стало отсутствие четкой концепции сбора фаунистических данных. Как результат, осуществлялся интуитивный выбор мест сбора информации, а способы оценки последствий появления новых данных были неясны. Вторая проблема – ограничение плотности публикации данных возможностями бумажных карт в биогеографии. И наконец, отсутствие единого хранилища фаунистических данных стало причиной потери больших объемов информации в “забытых” публикациях и общего замедления обработки фаунистической информации.
Изменения в фаунистике в последние 20 лет
Ситуация с фаунистическим картографированием коренным образом изменилась с развитием компьютерных технологий: баз данных (БД) и гео-информационных систем (ГИС). Базы данных позволили минимизировать усилия, затрачиваемые на ввод и поиск информации. Электронные карты сняли проблемы мелкомасштабных изображений. Теперь картографический продукт может существовать в любом масштабе (в т.ч. в разных одновременно), любой участок карты может быть просмотрен в крупном масштабе при увеличении (конечно, если исходные данные позволяют создавать карты в разных масштабах). Бюджеты создания разномасштабных карт многократно уменьшились. Наконец, совмещение ГИС и БД позволило создавать ряд карт автоматически. Естественно, это не могло не отразиться на фаунистике, как непосредственно, так и посредством влияния новых биогеографических концепций.
Упорядочивание фаунистической информации
Появление фаунистических баз данных сыграло революционную роль в развитии как фаунистики, так и биогеографии. С одной стороны, они дали возможность огромной экономии во времени каждого исследователя: каждую находку нужно ввести в БД только один раз (не важно, музейный это экземпляр или статья в сборнике), эту работу могут выполнять разные люди; единожды оказавшись в БД, находка за кратчайшее время может быть найдена разными способами интернет поиска. Проверка каждого наблюдения разными экспертами и внутренними средствами БД минимизируют возможность ошибок. С другой стороны, большие массивы данных БД и обилие пользовательских запросов позволили нащупать стандарт структуры фаунистических данных (Wieczorek et al., 2012). Иными словами, стало понятно, какие сопроводительные данные чаще всего бывают востребованы научным сообществом (и, наоборот, без каких данных наблюдение, скорее всего, не будет востребовано). Касательно фаунистических данных это, прежде всего, вид наблюдаемого животного, дата наблюдения, способ сбора информации, автор, описание места и географические координаты и т.п. (Wieczorek et al., 2012).
Мощнейшим подспорьем фаунистике оказалась возможность привлечения любителей для сбора фаунистических данных. Дефицит специалистов по фауне очевиден. Например, в России на последних съездах ВТО было зарегистрировано около 600 териологов. Понятно, что это не число фаунистов: не все териологи принимают участие в фаунистических исследованиях, как и не все фаунисты принимают участие в Съезде. Но по этой цифре можно составить общее представление о масштабе проблемы: на каждого из 600 териологов приходится 28 000 км2 территории России. Открытые БД позволили тысячам любителей вносить свой вклад в описание фауны. Использование любительских данных имеет свои ограничения (достоверность определения видов), но эта проблема имеет несколько решений: в научных исследованиях можно ограничивать набор видов, данные по которым собираются любителями, или использовать обязательное фотоподтверждение с экспертной оценкой точности определения (Lysaght, Marnell, 2016; Lissovsky et al., 2018).
В настоящий момент существуют как глобальные любительские БД (https://www.inaturalist.org, https://observation.org), так и базы данных отдельных стран, среди прочего позволяющие любителям вносить свою информацию (https:// artportalen.se, https://maps.biodiversityireland.ie, https://www.iop.krakow.pl/ssaki, https://nbnatlas.org, https://waarnemingen.be, https://www.ala.org.au, https:// www. artsobservasjoner.no, https://www.verspreidingsatlas.nl/zoogdieren). Более 12 000 статей по данным https://scholar.google.ru использовали информацию из этих ресурсов.
Ряд стран, воспринявших новые технологии, смог достаточно оперативно подготовить Атласы млекопитающих своей территории. Первым в 1999 г. выходит “Атлас млекопитающих Европы” (Mitchell-Jones et al., 1999). Работа начиналась еще без использования БД, но четкая систематизация данных позволила показать распространение млекопитающих в масштабе почти целой части света. В этом атласе использована стандартная картографическая сетка с размером ячейки 50 × 50 км. Со временем сложился стандарт использования ячейки 50 × 50 км при построении карт ареалов в континентальном масштабе и ячейки 10 × 10 км – в масштабе отдельных стран. Ряд стран издал атласы млекопитающих в таком масштабе (Anděra, 1995; Palomo et al., 2007; Lysaght, Marnell, 2016; Savoure-Soubelet et al., 2018; Crawley et al., 2020), но здесь следует обратить внимание, что благодаря планированию действий, фаунистические исследования через каждые 10 км действительно были проведены.
Изменение концепции ареала
Появление массивов геопривязанных фаунистических данных, а также многочисленных непрерывных в пространстве ГИС-ресурсов (данные дистанционного зондирования, климатические данные и рассчитанные по ним продукты и т.п.) закономерно привело к повышенному интересу к ареалам видов. В результате возникло целое направление “экологического моделирования” ареалов (species distribution modelling, environmental niche modelling), занимающееся реконструкцией структуры ареалов на основании точек регистрации видов и непрерывных данных среды (Peterson et al., 2011; Franklin, 2013; Guisan et al., 2017). Формально концепция ареала не изменилась (Лисовский, Оболенская, 2014). Экологическое моделирование все так же занимается поиском пригодных для вида местообитаний внутри единого ареала, как это было прежде. Но, благодаря совместным интенсивным усилиям широкого круга исследователей и появлению возможности верификации результатов моделирования, многие важные понятия были уточнены и развиты.
Прежде всего, стало понятно, что реально вычисляемым параметром на основании “экологических данных” (данных среды) является пригодность местообитаний для конкретного вида (Guillera-Arroita et al., 2015). Пригодность местообитаний через понятие емкости среды связано с обилием и плотностью населения вида. Поэтому стало возможно создавать карты обилия вида с тем пространственным разрешением, с которым существуют непрерывные данные среды (приблизительно и в среднем около 1 × 1 км) (Economov et al., 2020). Соответственно, создание карты кружева ареала со всеми локальными оптимумами, пессимумами и лакунами перестало быть мечтой. Кроме того, доступность непрерывных пространственных данных по абиотическим факторам (см. Пузаченко и др., 2010; Лисовский и др., 2020) позволила проводить типизацию ландшафтов по оптимальности индивидуально для каждого вида. Раньше на практике чаще всего для биогеографических построений использовали “готовую” типизацию ландшафтов, никак не связанную с экологическими предпочтениями того или иного вида.
Интенсивное развитие методов экологического моделирования позволило впервые сформулировать требования к пространственному распределению фаунистических данных (входящих данных для этого анализа). Было установлено, что фаунистические данные должны быть случайно распределены по всей территории ареала (Elith et al., 2011; Peterson et al., 2011; Guillera-Arroita et al., 2015). Любые агрегации данных приводят к сдвигу результатов анализа и должны быть устранены на подготовительном этапе (Fourcade et al., 2014; Inman et al., 2021). Аналогично, большие территории, не охваченные фаунистическими данными, негативно влияют на результаты анализа. С такой точки зрения, продуктивнее собирать данные с условным пространственным шагом или обследовать ячейки регулярной сети, чем досконально обследовать небольшой регион (размер шага и региона, несомненно, относительны и зависят от масштаба исследования). Изучение ареала на основании только краевых точек распространения вида можно сравнить с попыткой понять закономерность вылета теннисного мяча за пределы игровой площадки, при условии непрозрачности самой площадки.
Появилась возможность планировать фаунистическое исследование, благодаря доступности карт изученности территории (https://www.ala. org.au; https://rusmam.ru). Моделирование ареалов позволяет выявить участки, лишенные фаунистических данных и, таким образом, планировать сбор данных по конкретным видам. Возможность количественного сравнения разных моделей (Warren, Seifert, 2011; Radosavljevic, Anderson, 2014; Лисовский, Дудов, 2020), в том числе построенных на основании разных наборов данных, позволяют теперь выдвигать разные фаунистические гипотезы и количественно тестировать их.
Портал Млекопитающие России
Российская национальная БД распространения млекопитающих https://rusmam.ru была создана в 2017 г. рабочей группой Териологического общества при РАН (Lissovsky et al., 2018) и содержит по состоянию на октябрь 2022 г. более 200 000 записей. Каждая запись, кроме видовой идентификации, имени автора, даты, описания места и географических координат содержит точность определения географических координат, способ видовой идентификации, характер данных (фото, упоминание в литературе, музейный экземпляр, удаленная регистрация и т.д.), экспертно определяемый балл надежности видовой идентификации и ряд других параметров (Lissovsky et al., 2018). Благодаря структуре рубрикаторов, система позволяет оперативно создавать выборки фаунистических данных для широкого круга задач. Кроме того, обширный набор введенных в систему данных позволяет провести первичный статистический анализ фаунистической информации по территории России.
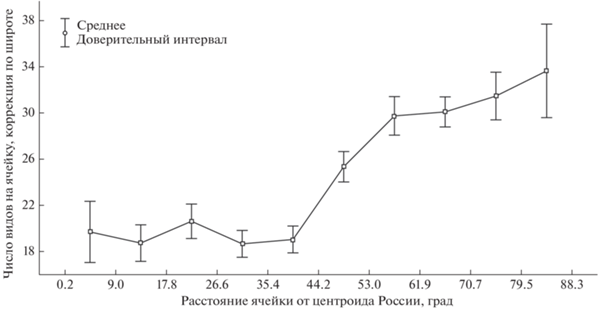
Прежде всего, можно констатировать, что территория России изучена далеко не полностью, при этом центральные части страны изучены намного слабее периферийных (рис. 3, 4). Кроме того, существуют точки, где собрано непропорционально много информации. Так, если рассматривать все наблюдения в проекции на регулярную сетку с ячейкой 50 × 50 км, то окажется, что 30.4% всех записей собраны всего лишь в одиннадцати ее квадратах. Остальные записи сделаны в 4429 квадратах из 7453, составляющих территорию страны. Около 10% собранных сейчас данных не могут быть геопривязаны с точностью до 2 км (условный порог, позволяющий использовать данные в большинстве исследований по экологическому моделированию). Также 10.1% всех данных собраны любителями, при этом 76.7% из них были уверенно определены специалистами по прилагаемым к наблюдениям фото. “Надежные” любительские наблюдения охватывают 192 вида из 323, зарегистрированных в России.
Рис. 3.
Распределение числа видов, обнаруженных в каждой ячейке размером 50 × 50 км на территории России по данным https://rusmam.ru. Естественные различия между числом видов в северных и южных широтах нивелированы при помощи линейной регрессии.
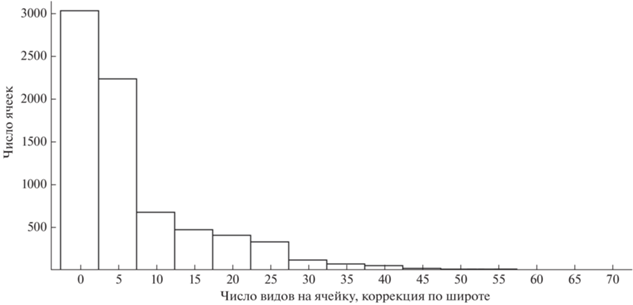
Рис. 4.
Распределение числа видов, обнаруженных в каждой ячейке размером 50 × 50 км на территории России, по удаленности от центроида страны по данным https://rusmam.ru. Естественные различия между числом видов в северных и южных широтах нивелированы при помощи линейной регрессии.
В качестве резюме можно констатировать, что технологические и теоретические изменения, произошедшие в последние десятилетия, позволили решить ряд проблем, с которыми сталкивалась фаунистика на протяжении длительного времени. Благодаря развитию фаунистических баз данных, были решены проблемы хранения и поиска фаунистической информации; разработаны подходы к использованию любительских данных; сформулирована необходимая структура собираемой фаунистической информации. Развитие ГИС сняло ограничения, связанные с мелким масштабом бумажных карт, и позволило планировать фаунистические исследования. Четкие представления о том, как должны быть пространственно распределены точки находок вида для возможного использования их в экологическом моделировании, позволяют надеяться, что такие данные будут собраны в недалеком будущем.