

Геоморфология и палеогеография, 2020, № 4, стр. 45-57
ПАЛЕОСЕЙСМОГЕННЫЕ ТЕКТОНИЧЕСКИЕ РВЫ – УНИКАЛЬНЫЕ АРХИВЫ ИСТОРИИ БЕЛОГО МОРЯ В ГОЛОЦЕНЕ
Н. Е. Зарецкая 1, 2, 3, *, А. В. Лудикова 4, С. В. Шварев 1, 3, Д. Д. Кузнецов 4, С. А. Кутенков 5
1 Институт географии РАН
Москва, Россия
2 Геологический институт РАН
Москва, Россия
3 Институт физики Земли имени О.Ю. Шмидта РАН
Москва, Россия
4 Институт озероведения РАН
Санкт-Петербург, Россия
5 Институт биологии Карельского Научного Центра РАН
Петрозаводск, Россия
* E-mail: n_zaretskaya@inbox.ru
Поступила в редакцию 21.01.2020
После доработки 05.03.2020
Принята к публикации 16.06.2020
Аннотация
На основе изучения различных седиментационных палеоархивов – осадков, заполняющих палеосейсмогенный тектонический ров, разрезов морских террас и литоральных ракушечных линз – получены новые данные об особенностях перемещения береговой линии Белого моря в голоцене в районах с блоковыми движениями земной коры (полуостров Киндо, полигон ББС МГУ, карельский берег Белого моря). Применение палеосейсмологического подхода во время детальных полевых исследований с выявлением характерных морфологических, структурных и динамических признаков древних землетрясений позволило установить сейсмогенное происхождение системы изученных рвов, заполненных голоценовыми осадками. Землетрясение, их породившее, произошло в позднеледниковье или раннем голоцене с образованием сейсморазрыва под водой. Впоследствии ров “раскрылся” в результате размыва и выноса тектонически раздробленных пород при гляциоизостатическом поднятии дна незадолго до заполнения его голоценовыми осадками. Литостратиграфическое изучение рыхлых отложений верифицировалось данными радиоуглеродного, диатомового анализов и изучения ботанического состава торфа, а также потерь при прокаливании. Осадки, заполнившие сейсморвы, накапливались в течение голоцена с перерывом на время перехода через береговую зону и демонстрируют последовательную смену морских обстановок осадконакопления (серый суглинок с раковинами Hiatellа arctica, возрастом 9 тыс. кал. лет назад) прибрежно-морскими (песок с обломками разной окатанности), затем изолирующегося, постепенно опресняющегося морского залива лагунного типа (слюдистый ил, возраст 3 тыс. кал. лет назад) и в конечном итоге – болота, заросшего еловым лесом (древесный торф, возраст 2 тыс. кал. лет назад). Строение верхней части разреза могло быть обусловлено: а) слабым поднятием уровня моря в позднем голоцене или б) слабым опусканием блока земной коры с расположенным на нем сейсморвом.
Также выявлены особенности формирования субфоссильных тафоценозов малакофауны в Кандалакшском заливе Белого моря, смены седиментационных обстановок в условиях поднимающегося берега и следы сильной сейсмичности в поздне- и постгляциальный период. Проведенное исследование показывает, что при изучении сейсмогенных форм весьма продуктивен комплексный подход, при котором выявляется полный спектр возрастных маркеров различных палеогеографических событий.
ВВЕДЕНИЕ
Белое море – внутреннее море Арктического бассейна севера европейской части России – представляет собой молодую экосистему, в современном виде начавшую формироваться в начале голоцена, после ухода с территории его бассейна последнего покровного ледника. Белое море представляет собой уникальную геодинамическую структуру: его северо-западные берега находятся на территории Балтийского щита и в голоцене испытывали и продолжают испытывать интенсивное, по преимуществу изостатическое поднятие, в то время как юго-восточное побережье, расположенное на Русской плите, в голоцене оставалось почти стабильным [1]. На фоне общего воздымания, характерного для щита, в северо-западной части Белого моря происходят дифференцированные движения земной коры, проявляющиеся как в неравномерном поднятии тектонических блоков на побережье, так и в опускании морского дна в сочетании с латеральными смещениями из-за неотектонической активизации Кандалакшского грабена [2]. Такие движения имели место как в голоцене, так и в позднем плейстоцене, проявляясь в палеосейсмических событиях. Это привело к формированию на территории северо-западного Прибеломорья многочисленных нарушений в скальных породах, среди которых особую роль для палеогеографических реконструкций имеют такие специфические формы рельефа, как тектонические рвы, относящиеся к первичным сейсмогенным дислокациям – следам подвижек вдоль плоскости разрыва, образующимся при выходе на поверхность очага землетрясения [3, 4]. Образование рвов в геологическом временном исчислении происходит мгновенно, а постсейсмогенная денудационная трансформация в условиях кристаллического субстрата весьма замедлена, поэтому они во многих случаях представляют собой “ловушки” для осадков. С целью изучения потенциальных палеоархивов природной среды, формирующихся в подобных “ловушках”, и дополнения голоценовой геологической истории Белого моря нами были проведены комплексные исследования одного из таких рвов и его заполнения.
РАСПОЛОЖЕНИЕ ОБЪЕКТА И МЕТОДЫ ИССЛЕДОВАНИЙ
Объект исследований расположен на полуострове Киндо (полигон Беломорской биологической станции (ББС) МГУ), в районе мыса Зелёный, на юго-западном берегу Кандалакшского залива Белого моря, на абсолютной отметке около 14.5 м. (рис. 1). Деформации рельефа были здесь обнаружены ранее в ходе специальных исследований, в результате которых был определен их сейсмический генезис и предположительно позднеголоценовый возраст [5]. Для изучения следов древних землетрясений нами использованы палеосейсмологический подход [6, 7] и разработанные на его основе методы идентификации сейсмогенных деформаций в рельефе, скальных породах и рыхлых отложениях [8]. В условиях стабильной платформенной территории с отсутствием современной сильной сейсмичности корректное использование палеосейсмогеологического подхода опирается на детальные полевые исследования с выявлением характерных морфологических, структурных и динамических признаков древних землетрясений, пространственно сопряженных с выраженными в морфоструктуре потенциально сейсмогенерирующими разломами.
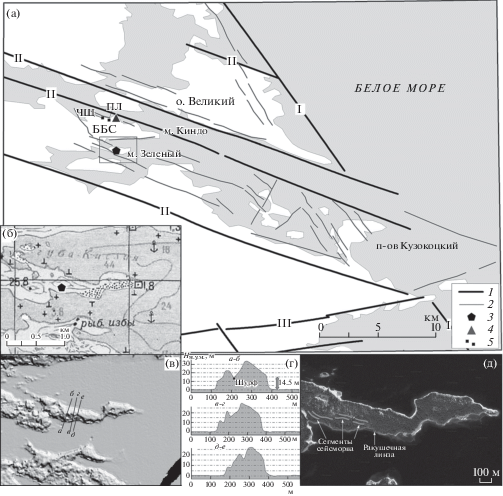
Рис. 1.
Палеосейсмотектонический ров на мысе Зеленый.
(а) – морфолинеаменты, определяющие блоковую структуру территории: 1 – основные, 2 – второстепенные; 3 – расположение сейсморва; 4 – Беломорская биологическая станция; 5 – ПЛ (“Палеонтологическая линза”); ЧЩ – Чёрные Щали; (б) – крупномасштабная топографическая карта участка; (в) – цифровая модель рельефа, созданная с использованием данных ArcticDEM, и поперечные профили через полуостров; (г) – поперечные профили через полуостров и сейсморов и место расположения шурфа с голоценовыми осадками; (д) – детальный космоснимок участка исследований (сервис GOOGLE) с фрагментами палеосейсмотектонического рва и местоположение ракушечной линзы.
Дно рва перекрыто отложениями разного генезиса, общей мощностью около 1 м (рис. 2). Строение разреза следующее (сверху вниз): 1) 0–5 см – дернина (моховая подушка); 2) 5–55 см – торф коричневый (до черного); 3) 55–90 см – ил от оливково-серого до коричнево-бурого, слюдистый; 4) 90–100 (110) см – линзовидный прослой невыдержанной мощности слабоокатанной гальки и мелких валунов в заполнителе из грубозернистого песка; 5) 100 (110)–105 (120) см – серый суглинок, очень плотно набитый мелкими раковинами и ракушечным детритом двустворчатых моллюсков; залегает непосредственно на скале (дне рва).
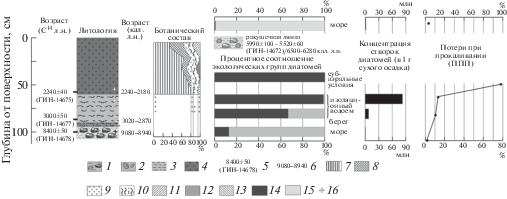
Рис. 2.
Результаты исследования заполнения сейсморва.
Литология: 1 – суглинок с ракушей, 2 – галька и валуны в заполнителе из грубозернистого песка, 3 – ил слюдистый, 4 – торф; возраст: 5 – радиоуглеродная дата, лабораторный индекс и номер, 6 – калиброванный; ботанический состав: 7 – древесина (ель, береза), 8 – кустарнички, 9 – хвощ, 10 – травы, 11 – мхи, 12 – мицелий грибов, 13 – пушица; экологические группы диатомей: 14 – пресноводные (галофилы+индифференты), 15 – солоноватоводно-морские и морские; 16 – места отбора образцов.
Заполнение рва изучалось комплексом методов.
Раковины моллюсков определялись до вида11. В качестве сравнительных материалов привлекались данные по ракушечным тафоценозам, обнаруженным и исследованным ранее на полигоне ББС МГУ [9, 10]. Все исследованные ракушечные местонахождения были продатированы радиоуглеродным методом по стандартной методике, применяющейся в лаборатории геохимии изотопов и геохронологии Геологического института РАН: раковины отмывались от вмещающих отложений и обрабатывались раствором соляной кислоты для снятия внешнего слоя с внутренней и внешней поверхности. Дальнейшая процедура определения 14С возраста раковин описана в работе [11]. По образцам из ила и торфа также были получены 14С даты по методике, описанной в упомянутой работе. Калибровка радиоуглеродных дат проводилась с использованием калибровочной программы Calib 704 [12]; даты, полученные по образцам раковин, калибровались с использованием калибровочной кривой IntcalMarine, даты по торфу – Intcal13, по илам – с помощью кривой для смешанных образцов.
Методами диатомового анализа и анализа содержания органического вещества были изучены четыре пробы из заполнения рва, а также одна проба из ракушечной линзы, расположенной в 100 м к югу от разреза и вскрытой в тыловом шве современного пляжа. Пробоподготовка для диатомового анализа и расчет концентрации створок в 1 г сухого осадка выполнены по стандартной методике [13].
Анализ потерь при прокаливании проводился по принятой в Институте озероведения РАН методике [14]. С целью ботанического анализа торфа было отобрано 5 образцов с шагом 10 см. Ботанический анализ торфа на макроостатки и степень его разложения выполнен микроскопическим методом (с точностью до 5%) в лаборатории болотных экосистем Института биологии КарНЦ РАН22.
РЕЗУЛЬТАТЫ: МОРФОСТРУКТУРА И ПОЛУЧЕННЫЕ АНАЛИТИЧЕСКИЕ ДАННЫЕ
Мыс Зеленый представляет собой коленообразно изогнутую в плане прерывистую скальную гряду, кулисообразные фрагменты которой, протяженностью до нескольких сотен метров, вытянуты в ЗСЗ направлении (рис. 1, врезка). Морфоструктура мыса сформирована с преобладанием разрывов ЗСЗ простирания, определяющих ориентировку гряды, ее осевой зоны и флангов, и с участием второстепенных разрывов ССВ простирания, смещающих фрагменты гряды относительно друг друга к югу и северу. Ров или, скорее, система коротких (первые десятки метров), но широких (до 10–20 м по бровкам и до 10 м по днищу) и глубоких (до 5–6 м) рвов рассекает скальную гряду в ее осевой части. Широкие продольные рвы сопрягаются с узкими “щелями” и вертикальными стенками, ориентированными в поперечном направлении. Высота гряды составляет 15–20 м над у. м., а днище основного рва (с заполнением) расположено на высоте около 14.5 м над у. м.
Простирание основных (ЗСЗ) и второстепенных (ССВ) элементов рва полностью отвечает общему морфоструктурному плану территории, что подтверждает его тектоническую обусловленность, а продольное рассечение вершинной части скального гребня со следами ледниковой обработки при отсутствии ледниковых отложений в днище определяет его сейсмический генезис и позднеледниковый возраст. Особенность системы рвов – сочетание значительных величин глубины и ширины и ограниченного продольного развития при почти полном отсутствии коллювия и плоском днище. Очевидно, что такой характер связан с последующей экзогенной переработкой сейсмически подготовленного материала. Выход сейсморазрыва на поверхность проявился в узкой плоскости собственно разрыва и сопряженной широкой зоне мелкодисперсного дробления, материал из которой в виде дресвы, щебня и мелких глыб вынесен в результате интенсивной волноприбойной деятельности.
Подобные рвы в различных “стадиях” – от широких (до нескольких метров) до узких трещин можно наблюдать в непосредственной близости – на островах Кузокоцкого архипелага, где они находятся либо в зоне волнового воздействия, либо совсем недавно вышли из него. Характерная особенность рвов – резкий переход от широкой “траншеи” с плоским днищем, расположенной выше уровня моря (где имеется или было волновое воздействие), к узкой трещине-разрыву ниже уровня моря (где его пока не было). Исходя из этих предпосылок, можно констатировать, что землетрясение, породившее систему сейсморвов на мысе Зеленый, произошло в позднеледниковье или раннем голоцене с образованием сейсморазрыва под водой, а ров “раскрылся” в результате последующего подъема дна, размыва и выноса раздробленных пород в волноприбойной зоне незадолго до заполнения его голоценовыми осадками.
Изучение заполнения сейсморва выявило следующие особенности его строения, состава и возраста. Слой серого суглинка, выстилающего днище рва, и залегающий непосредственно на биотитовых и эпидот-содержащих гранито-гнейсах керецкой толщи нижнего архея, плотно набит мелкими раковинами вида Hiatellа arctica, размер которых близок (5–7 мм) к современным раковинам и существенно (в два-три раза) меньше раковин, найденных в осадках, формировавшихся во время климатического оптимума голоцена [10]. Эти моллюски встречаются на глубинах от 2 до 140 м, на илистых и смешанных грунтах, при температурах от –0.9 до 20.3°С и солености 16.4–30.1‰, однако максимум обилия наблюдается на глубине 20 м, а наибольшая плотность поселения – на глубине 8 м в биоценозе Laminaria digitata [15]. По раковинам из разреза получена радиоуглеродная дата 8400 ± 50 (ГИН-14678, 9080–8940 кал. л. н.) (табл. 1). Таким образом, слой формировался на дне рва в раннем бореальном периоде голоцена. На полигоне ББС МГУ это самое древнее местонахождение погребенной ракуши.
Концентрация створок диатомей в пробе из серого суглинка, насыщенного раковинами, крайне низка – 19 тыс. в 1 г сухого осадка. В составе диатомовых комплексов отмечаются обитатели морских мелководий – солоноватоводно-морские Grammatophora oceanica, реже Cocconeis scutellum и Rhabdonema minutum. Присутствуют также единичные створки пресноводного обрастателя Fragilaria pinnata, индифферентного по отношению к солености, и изредка – споры морских планктонных Chaetoceros spp. Преобладание бентосных видов указывает на то, что отложения формировались в условиях морской сублиторали (по данным изучения малакофауны на глубине около 8 м). Низкая концентрация створок и обедненный видовой состав диатомовых комплексов свидетельствуют об обстановках, неблагоприятных для развития диатомей (низкая продуктивность) и аккумуляции их створок (терригенное разбавление, воздействие течений).
На суглинке с ракушей линзами залегает, возможно – с размывом, прослой слабоокатанной гальки и мелких валунов в заполнителе из грубозернистого песка, который, в свою очередь, перекрыт оливково-серыми илами с листоватой слоистостью, насыщенными слюдой, и с заметным содержанием рассеянной органики. Анализ макроостатков растений показал присутствие корней хвоща, очевидно, проникших в слой из вышележащих отложений. Из этого слоя с глубины 0.85 м была получена дата 3000 ± 50 (ГИН-14677, 3020–2870 кал. л. н.). Концентрация створок диатомей возрастает до 8.8 млн. В составе диатомовых комплексов преобладают разнообразные обрастатели Fragilariaceae (в сумме 63%), массовое развитие которых сопровождает процесс изоляции от моря [16]. Численности субдоминантов достигают литоральные солоноватоводно-морские Cocconeis scutellum и Opephora mutabilis. В верхней части данного горизонта происходит дальнейшее увеличение концентрации створок диатомей до 84 млн. Суммарная численность обрастателей Fragilariaceae достигает 98%. Лишь единично отмечены створки морских литоральных диатомей. Диатомовые комплексы с доминированием обрастателей Fragilaria sensu lato отмечены в донных отложениях разновозрастных изоляционных бассейнов беломорского побережья [17]. Таким образом, накопление слюдистых илов происходило в условиях сильно опресненного изолирующегося морского залива, периодически (во время высоких приливов или штормовых заплесков) сообщавшегося с морем. При этом по данным диатомового анализа влияние морских вод заметно ослабело на этапе формирования верхней части отложений. Значения потерь при прокаливании характерны как для прибрежных морских, так и для озерных обстановок (10–13%). По-видимому, это была узкая котловина лагунного типа с благоприятными условиями для произрастания прибрежно-водной растительности и аккумуляции тонкого материала.
Дальнейшее осадконакопление во рву шло практически без перерыва – верхняя часть его заполнения представлена еловым торфом, по которому была получена дата 2240 ± 40 (ГИН-14675, 2240–2180 кал. л. н.). Диатомовые водоросли в нижней части торфяной залежи практически отсутствуют, за исключением единичных обломков пресноводных бентосных Pinnularia spp и неопределимых мелких фрагментов. Формирование отложений происходило в условиях, неблагоприятных для развития диатомей, по-видимому, в результате дефицита увлажненности. Значение потерь при прокаливании достигает 80%, что характерно для торфяных отложений.
По-видимому, во рву произошла закономерная эвтрофикация изолировавшегося от моря мелкого водоема. Склоновое положение участка обусловило проточный тип увлажнения атмосферными и поверхностными водами и развитие здесь елового сообщества на переувлажненном грунте. Данным сообществом сформировано 55 см торфа, состоящего преимущественно из остатков ели, а также березы, черники, вороники, хвоща и разнотравья (рис. 2). Состав торфа отражает однородный состав материнского сообщества весь период его существования. Степень разложения высокая – 40% и более, скорость торфонакопления, напротив, низкая – 0.25 мм/год, что наряду с развитым древостоем свидетельствует о преобладании лесообразовательных процессов над болотообразовательными.
В 100 м к юго-востоку от изученного разреза, на берегу моря в тыловом шве пляжа (в верхней части современной литорали 0.5 м ниже у. м., рис. 1, д) была обнаружена выходящая на дневную поверхность ракушечная линза, представленная серым суглинком с богатым комплексом субфоссильной малакофауны более крупной размерности. Здесь двустворчатые моллюски представлены как раковинами Hiatellа arctica, так и Tridonta borealis (рис. 3). Последняя в настоящее время встречается на глубинах от 0 до 40 м на илистых, песчаных и смешанных грунтах при температуре от –1.4 до 14.2°С и солености 23.0–27.8‰, при максимуме обилия на глубине 15 м и наибольшей плотности популяции на глубине 5 м в биоценозе Laminaria saccharina [15]. Размеры раковин обоих видов в линзе превышают размеры современных моллюсков в 2–3 раза; по ним получены даты в диапазоне от 5990 ± 100 (ГИН-14672) до 5390 ± 70 (ГИН-14673), что составляет 6500–5700 кал. л. н. (табл. 1). Диатомовый анализ вмещающих раковины глинистых отложений показал преобладание солоноватоводно-морских и морских диатомей. Среди первых наиболее многочисленны бентосные виды (46%), вторые преимущественно представлены спорами планктонных (40%). Диатомовые комплексы формировались в условиях морского залива, сообщающегося с открытым морем. По-видимому, это был один из неглубоких (по данным изучения малакофауны, около 5–8 м) заливов среднеголоценового моря Тапес, характеризовавшегося длительным стоянием воды на уровне выше современного и более высокой температурой [1].
Рис. 3.
Субфоссильные раковины двустворчатых моллюсков.
(а) – Hiatella arctica; (б) – Tridonta borealis [21].

Субаэральные местонахождения, подобные описанному заполнению рва, в Западном Прибеломорье достаточно редки. Причина, по-видимому, в плохой обнаженности четвертичных осадков и малом количестве естественных “коллекторов”, в которых могут накапливаться и сохраняться органогенные отложения. На поднятых участках побережья такие местонахождения иногда вскрываются карьерами или во время дорожных работ. Так, при прокладке трассы на территории ББС МГУ в 1982 г. на высоте 24.5 м над у. м. было вскрыто местонахождение субфоссильной малакофауны во вмещающих серых суглинках (т.н. “Палеонтологическая линза”, или “Палеонтологический объект”, рис. 1, а). Строение разреза следующее (сверху вниз): 1) 0–0.03 м – дернина; 2) 0.03–1.1 м – пачка серо-желтых песков, преимущественно крупнозернистых, с косой субгоризонтальной и наклонной слоистостью, с прослоями гравия, с четкой и ровной нижней границей; 3) 1.1–2.0 м – песчано-гравийная толща с большим количеством гальки и валунов диаметром до 30 см (видимые); 4) 2.0–2.3 м – серый суглинок с ракушей; 5) 2.3–2.6 м (видимая мощность) – сизый тяжелый суглинок (рис. 4).
Рис. 4.
Сопоставление разрезов заполнения сейсморва и “палеонтологической линзы”.
1 – глина; 2 – суглинок с ракушей; 3 – ил слюдистый; 4 – торф; 5 – песок; 6 – галька и валуны в заполнителе из грубозернистого песка; 7 – радиоуглеродная дата, лабораторный индекс и номер.
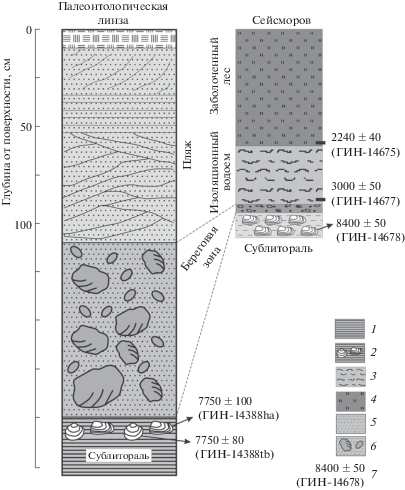
В составе погребенной малакофауны доминируют те же два вида – H. arctica и Т. borealis, встреченные нами в местонахождении на литорали у сейсморва. По радиоуглеродным образцам из каждого вида были получены даты – 7750 ± 100 и 7700 ± 80 (8400–8600 кал. л. н., ГИН-14388) (табл. 1). Ранее для этого слоя были проведены палинологические исследования [18]; состав палинокомплекса показал, что слой формировался в сублиторальных условиях, а характер реконструируемой растительности мог соответствовать началу атлантического периода голоцена. Полученные нами радиоуглеродные даты подтверждают этот вывод.
На берегу моря неподалеку от разреза с “палеонтологической линзой” (в 500 м к СЗ), в урочище Чёрные Щали (рис. 1, а), в верхней части литорали нами был обнаружен выход на поверхность размываемой линзы серых глин с ракушей. В составе ракуши преобладали створки Tridonta borealis (при наличии единичных экземпляров Hiatella arctica), по которым была получена радиоуглеродная дата 4590 ± 110 (4640–4950 кал. л. н., ГИН-14387).
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Итак, в позднеледниковье – раннем голоцене в районе современного мыса Зелёный под водой в результате сейсмического события сформировался сейсморазрыв, который “раскрылся” в результате последующего подъема дна. Размыв и вынос сейсмоколлювия (раздробленных пород) могли произойти при переходе сейсморва через волноприбойную зону во время раннеголоценовой регрессии Белого моря, возраст которой, по данным ряда исследователей, определяется как 10.2–9.3 кал. тыс. л. н. [1, 19, 20]. Заполнение рва голоценовыми осадками началось после трансгрессии Белого моря, когда его дно оказалось под водой на глубине около 8 м, достаточной для массового развития там моллюсков Hiatella arctica, ∼9.1–8.9 кал. тыс. л. н.
При дальнейшем гляциоизостатическом подъеме территории при переходе через береговую линию дно сейсмотектонического рва вместе с ракушечным слоем существовало, по-видимому, в условиях практически нулевой седиментации. Возможно, верхняя часть слоя была размыта, так как в осадконакоплении наблюдается перерыв, зафиксированный радиоуглеродными датами (∼9–3 кал. тыс. л. н.), а сам слой перекрыт 5-сантиметровым слоем отмостки (слабоокатанной галькой с песчаным заполнителем). Однако в то время, как во рве доминировали прибрежно-морские процессы, на подводном склоне, поднявшемся до уровня сублиторали, в непосредственной близости ото рва на глубине около 5 м 6.5–5.7 кал. тыс. л. н. происходило формирование новой ракушечной линзы с более богатой и крупной фауной. Как было установлено ранее [10], подобные линзы формируются вблизи скал в местах распространения ламинарников – единственном биогеоценозе, в котором оба вида (H. arctica и T. borealis) могут существовать одновременно.
Дальнейшая смена характера осадконакопления в сейсмотектоническом рве предполагает два сценария развития. Первый – слабое поднятие уровня моря до формирования небольшой лагуны около 3 кал. тыс. л. н., которая впоследствии отчленилась от основного бассейна, и превратилась сначала в пресное озерцо, впоследствии (около 2.2 кал. тыс. л. н.), с падением базиса эрозии, заболотившееся. Второй сценарий – слабое опускание блока земной коры с расположенным на нем сейсморвом, впоследствии стабилизировавшееся на фоне общего поднятия территории и регрессии Белого моря. Таким образом, лагунные обстановки пришли на смену условиям нулевой седиментации либо из-за повышения уровня моря, либо из-за опускания блока земной коры.
Сходную историю имеет “Палеонтологический объект”, расположенный с другой стороны мыса Киндо. Здесь формирование слоя морских серых глин происходило до 8.6 кал. тыс. л. н., а затем в период 8.6–8.4 кал. тыс. л. н. на глубине 8–5 м ниже уровня моря сформировалась линза с ракушей (H. arctica и T. borealis). При поднятии территории и прохождении через береговую линию здесь формировался бенч, выстланный валунами и галькой в песчаном заполнителе, перекрытый затем пляжевыми отложениями (песками с разнообразной слоистостью), а в урочище “Чёрные Щали” в то же время (4.9–4.6 кал. тыс. л. н.) на глубине 5 м формировалась линза с Tridonta borealis. Отсутствие на “Палеонтологическом объекте” седиментационной “ловушки” в виде сейсморва обусловило формирование пляжа, а не лагуны с органогенными осадками и более полной летописью.
Таким образом, мы видим, что расположенные недалеко друг от друга седиментационные архивы формировались практически сходным образом, но в разное время и на разных абсолютных отметках (“Палеонтологическая линза” на высоте 24.5 м “запаздывала” относительно сейсморва, расположенного на высоте 14.5 м). Причина такой разницы, по-видимому, заключается в расположении этих объектов в пределах разных тектонических блоков, которые в голоцене испытывали дифференцированные движения (разная скорость + разные направления?). Дать точную оценку скорости движения блоков в голоцене пока не представляется возможным из-за неполноты полученных летописей.
ВЫВОДЫ
Следы сильной сейсмичности в виде первичных сейсмодеформаций – тектонических рвов в местах поверхностных разрывов, образовавшихся во время землетрясений, традиционно являются источником данных о палеосейсмичности территории, поскольку дают информацию о разных ее параметрах – генерирующих структурах, интенсивности, возрасте и повторяемости. Последние два параметра основываются на анализе возраста коррелятных событию (или последовательным событиям) отложений, захороненных в таких формах рельефа.
Проведенное исследование показывает, что весьма продуктивным является комплексный подход при исследовании сейсмогенных форм, при котором выявляется полный спектр возрастных маркеров различных палеогеографических событий.
Диатомовый и ботанический анализы отложений, заполняющих тектонический ров, выявили смену седиментационных обстановок, обусловленную перемещениями относительного уровня Белого моря в голоцене и осложненную локальными тектоническими движениями, положенную, благодаря радиоуглеродным датам, на хронологическую канву. Динамика содержания органического вещества отражает смену седиментационных обстановок от морских к прибрежно-морским и далее субаэральным.
Изучение седиментационных палеоархивов разного генезиса подтвердило наличие блоковых движений на относительно небольшой территории в голоцене.
Список литературы
Кошечкин Б.И. Голоценовая тектоника восточной части Балтийского щита. Л.: Наука, 1979. 157 с.
Балуев А.С., Журавлев В.А., Терехов Е.Н., Пржиялговский Е.С. Тектоника Белого моря и прилегающих территорий // Труды Геологического института. Вып. 597. 2012. 104 с.
Солоненко В.П. Землетрясения и рельеф // Геоморфология. 1973. № 4. С. 3–12.
Никонов А.А. Терминология и классификация сейсмогенных нарушений рельефа // Геоморфология. 1995. № 1. С. 3–9.
Мараханов А.В., Романенко Ф.А. Новые данные о послеледниковых сейсмодислокациях Северной Карелии (Карельский берег Белого моря) // Юдахинские чтения. Геодинамика и экология Баренц-региона в XXI в. / Мат-лы докл. Всерос. конф. с междунар. участием. Архангельск. 2014. С. 137–140.
Флоренсов Н.А. О неотектонике и сейсмичности Монголо-Байкальской горной области // Геология и геофизика. 1960. № 1. С. 74–90.
Солоненко В.П. Палеосейсмогеология // Изв. АН СССР. Физика Земли. 1973. № 9. С. 3–16.
Paleoseismology. McCalpin J.P. (Ed.). San-Diego: Acad. Press. 1996. 585 p.
Зарецкая Н.Е., Шевченко Н.В., Басилян А.Э., Покровский Б.Г., Максимов Ф.Е., Кузнецов В.Ю. Голоценовая малакофауна полуострова Киндо (Кандалакшский залив Белого моря): геолого-геохронологические исследования // Мат-лы XIX междунар. науч. конф. (школы) по морской геологии. М.: ГЕОС, 2011. Т. 3. С. 154–159.
Зарецкая Н.Е., Шевченко Н.В., Хайтов В.М. Голоценовые тафоценозы малакофауны Белого моря: особенности формирования и радиоуглеродная хронология // Мат-лы третьей Всерос. науч. конф. с междунар. участием “Динамика современных экосистем в голоцене”. Казань. 2013. С. 160–164.
Zaretskaya N.E., Hartz Sönke, Terberger Thomas, Savchenko S.N., and Zhilin M.G. Radiocarbon Chronology of the Shigir and Gorbunovo Archaeological Bog Sites, Middle Urals, Russia // Radiocarbon. 2012. Vol. 54 (3). P. 783–794.
Reimer P.J., Bard E., Bayliss A., Beck J.W., Blackwell P.G., Ramsey C.B., Buck C.E., Cheng H., Edwards R.L., Friedrich M., Grootes P.M., Guilderson T.P., Haflidason H., Hajdas I., Hatte C., Heaton T.J., Hoffmann D.L., Hogg A.G., Hughen K.A., Kaiser K.F., Kromer B., Manning S.W., Niu M., Reimer R.W., Richards D.A., Scott E.M., Southon J.R., Staff R.A., Turney C.S.M., and Plicht J. IntCal13 and Marine13 radiocarbon age calibration curves 0-50.000 years cal. BP // Radiocarbon. 2013. Vol. 55 (4). P. 1869–1887.
Давыдова Н.Н. Диатомовые водоросли – индикаторы природных условий водоемов в голоцене. Л.: Наука, 1985. 244 с.
Кузнецов Д.Д. Динамика содержания органического вещества в колонках озерных отложений Европейской России и реконструкция голоценовых климатических изменений // Paleolimnology of Northern Eurasia. Experience, Methodology, Current Status: Proceedings of the International Conference. Yakutsk, 22–27 August, 2016 / Eds. S. Levina, R. Gorodnichev, I. Yadrikhinski, P. Davydova. Yakutsk: North-Eastern Federal University, 2016. P. 146–149.
Наумов А.Д. Двустворчатые моллюски Белого моря: опыт эколого-фаунистического анализа. СПб.: Зоологический институт РАН, 2006. 351 с.
Stabell B. The development and succession of taxa within the diatom genus Fragilaria Lyngbye as a response to basin isolation from the sea // Boreas. 1985. Vol. 14. P. 273–286.
Лудикова А.В. От моря к озеру: биоиндикационные признаки изоляционных процессов // Мат-лы Всерос. научной конф. “Поздне- и постгляциальная история Белого моря: геология, тектоника, седиментационные обстановки, хронология”: Сборник статей. М.: “КДУ”, Университетская книга, 2018. С. 104–108.
Зюганова И.С. Палинологическое исследование линзы голоценовых отложений в районе Беломорской станции МГУ // Вестн. Моск. ун-та. Сер. 4. Геология. 1999. № 3. С. 47–49.
Колька В.В., Евзеров В.Я., Мёллер Я.Й., Корнер Г.Д. Перемещение уровня моря в позднем плейстоцене–голоцене и стратиграфия донных осадков изолированных озер на южном берегу Кольского полуострова, в районе поселка Умба // Изв. РАН. Сер. геогр. 2013. № 1. С. 73–88.
Зарецкая Н.Е. Голоценовая история дельты р. Северной Двины // Геоморфология. 2018. № 1. С. 3–17.
Мычко Э.В. Комплексное изучение фауны и условия формирования линзы глин в районе ББС. Курсовая работа кафедры палеонтологии Геологического факультета МГУ. 2006. 48 с.
Дополнительные материалы отсутствуют.
Инструменты
Геоморфология и палеогеография