

Исследование Земли из Космоса, 2023, № 3, стр. 27-43
Оценка индекса NDVI как источника информации о надземной фитомассе в степях
А. В. Хорошев a, *, О. Г. Калмыкова b, Г. Х. Дусаева b
a Московский государственный университет имени М.В. Ломоносова
Москва, Россия
b Институт степи УрО РАН
Оренбург, Россия
* E-mail: avkh1970@yandex.ru
Поступила в редакцию 16.09.2022
- EDN: TYJJZE
- DOI: 10.31857/S020596142303003X
Аннотация
Трудоемкость измерений фитомассы на пробных площадках сдерживает экстраполяцию точечных данных на территории, сопоставимые с хозяйственными угодьями или ландшафтными единицами. Вегетационные индексы, рассчитываемые по космическим снимкам, обычно рассматриваются как индикаторы зеленой фитомассы и используются для ее площадных оценок. В исследовании решается задача установить информативность нормализованного разностного вегетационного индекса NDVI в зависимости от фракционной структуры живой и мертвой надземной фитомассы, сезонной динамики биологического круговорота, гидротермических условий и ландшафтной позиции. Использованы результаты ежемесячных измерений фракций надземной фитомассы на 13 площадках в фитоценозах залесскоковыльной и ковылковой формаций в Буртинской степи (заповедник “Оренбургский”) с мая по сентябрь в 2015–2020 гг. Для каждого срока по космическим снимкам Landsat рассчитаны значения NDVI на всех площадках. Гипотезы о геоботанических, гидротермических, фенологических и ландшафтных факторах информативности NDVI проверялись расчетом коэффициентов корреляции Спирмена, средствами дисперсионного и мультирегрессионного анализа. Несовпадение сезонных пиков NDVI и зеленой фитомассы не согласуется с распространенным представлением о прямом индикационном значении NDVI. Общая живая фитомасса более четко коррелирует с индексом в июне и июле, слабее – в конце сезона. NDVI оказался чувствительным не столько к запасам зеленой фитомассы как таковой, сколько к массе и доле разнотравья и соотношению живой и мертвой фитомасс. В поздневесеннее и раннелетнее время NDVI наиболее тесно связан с разнотравьем, в июле – со злаками. Подтвердилась гипотеза о возможности экранирования зеленой массы ветошью, что приводит к снижению NDVI несмотря на сохранение или рост зеленой фитомассы. NDVI может занижать реальную зеленую фитомассу, если происходит резкий прирост массы ветоши, обычно – во второй половине лета и начале осени. NDVI более адекватно отражает состояние надземной фитомассы степных сообществ, длительное время не подвергавшихся воздействию пожаров, по сравнению с горевшими сообществами и залежами.
ВВЕДЕНИЕ
Динамические ряды измерений фитомассы, особенно ее фракционного состава, относятся к числу наиболее трудоемких видов исследования функционирования ландшафтов (Базилевич и др., 1978; Титлянова и др., 2018). В то же время потребность в данных о фитомассе велика. Во-первых, это связано с ее ресурсным значением (урожайность сельскохозяйственных культур, качество пастбищ, запасы трав на сенокосах, запасы древесины и др.). Во-вторых, фитомасса – надежный индикатор экологических условий и их антропогенных трансформаций. В-третьих, она выступает как индикатор защищенности ландшафта от экзодинамических процессов (эрозии, дефляции, селей, термокарста и др.) и нарушений режима стока. Сезонная и многолетняя динамика фитомассы и ее составляющих важна для оценки устойчивости экосистем как способности выдерживать тот или иной диапазон естественных и антропогенных колебаний среды. Трудоемкость измерений фитомассы на немногочисленных пробных площадках и невозможность охватить все составляющие ландшафтного разнообразия сдерживают получение оценок параметров функционирования, которые позволяли бы экстраполировать точечные результаты на территории, сопоставимые с хозяйственными угодьями или ландшафтными единицами.
В последние два–три десятилетия острота этой проблемы, казалось бы, сглаживается благодаря доступности многоканальной космической информации и появлению разнообразных индексов состояния компонентов ландшафтов. Нормализованный разностный вегетационный индекс (NDVI) довольно давно трактуется как индикатор зеленой надземной фитомассы, отражающий интенсивность фотосинтеза. Связь NDVI с реальной надземной фитомассой чаще всего исследуется при выявлении факторов варьирования урожайности сельскохозяйственных культур (Nagy et al., 2018), в частности – причин внутриполевого варьирования (Verhulst, Govaerts, 2010; Гулянов, 2019). Одной из традиционных областей применения NDVI стали исследования межгодового варьирования значений NDVI и связи с климатическими флуктуациями и трендами (de Jong et al., 2012; Елсаков, Телятников, 2013; Курганович, Голятина, 2015; Han et al., 2019), изменениями землепользования (Peng et al., 2019). Установлена пространственная нестационарность отношений между NDVI и климатическими факторами (Gao et al., 2012; Zhao et al., 2015).
В то же время накопление литературных данных свидетельствует о неоднозначности трактовок этого индекса. Описано немало ограничений на прямолинейную интерпретацию и уточнений для разных географических условий. Так, высокая корреляция NDVI с зеленой массой получена для засушливых районов (Золотокрылин и др., 2020). Для лесов результаты противоречивы: для разных регионов получены как сильные связи (Елсаков, 2012), так и слабые (Залиханов и др., 2010). Получены данные о неравноценной информативности индекса в разные сезоны (Lyle et al., 2013), при экстремальных погодных условиях (Piao et al., 2014; Nagy et al., 2018). Установлена высокая чувствительность NDVI к экологическим характеристикам местообитания – почвам, влажности, рельефу (Gamon et al., 2013; Araya et al., 2017; Maynard, Levi, 2017; Piedallu et al., 2019; Гопп и др., 2016). Обращается внимание на неоднозначные соответствия между NDVI и фитомассой, связанные с большим количеством одревесневающих и усыхающих стеблей (Лиджиева и др., 2012), временной изменчивостью аспектов посевов (Жуков и др., 2013), различной архитектоникой доминантных видов растений по сезонам (Ерошенко и др., 2018), “насыщением” при высокой густоте растительного покрова (Савин и др., 2015), несовпадением сезонных максимумов NDVI и продуктивности (Рулев и др., 2016). В работе В.В. Михайлова с соавторами (2021) для тундровых сообществ средствами регрессионного моделирования выполнен переход от безразмерных показателей NDVI к метрическим величинам хлорофиллового индекса, по которому оценена масса автотрофных органов растений и определена фитомасса сообщества с учетом особенностей ее накопления и распределения в растениях. В связи с этим становится ясной необходимость и обратной задачи: строгой верификации NDVI по наземным измерениям фракций надземной фитомассы – живой и мертвой и связанной с разными агробиологическими группами растений.
В нашем исследовании поставлена задача установить зависимость NDVI от фракционной структуры живой и мертвой надземной фитомассы, сезонной динамики биологического круговорота, гидротермических условий года и ландшафтной позиции на основании многолетних сезонных измерений в заповедном степном ландшафте.
МАТЕРИАЛЫ И МЕТОДЫ
В качестве модельной территории для проведения исследований был выбран участок Буртинская степь заповедника “Оренбургский” и его охранная зона. В ботанико-географическом отношении этот заповедный кластер расположен в подзоне разнотравно-дерновиннозлаковых Заволжско-Казахстанских степей (Сафронова, Калмыкова, 2012), в почвенно-географическом – в подзоне черноземов южных (Красная книга почв …, 2001).
На территории Буртинской степи было заложено 6 стационарных мониторинговых степных участков в разных экологических условиях (Дусаева и др., 2016; Максутова и др., 2016; Дусаева, Максутова, 2017; Dusaeva et al., 2019). Растительные сообщества на участках относились к различным группам ассоциаций двух формаций – Stipeta zalesskii и Stipeta lessingiana, – сообщества которых наиболее распространены (Калмыкова, 2012) на исследуемом заповедном кластере). Участки № 1 и 4 в наибольшей степени пострадали от перевыпаса в дозаповедный период, а участок № 5, расположенный в охранной зоне заповедника – старовозрастная залежь. В пределах участков выбирались пары мониторинговых площадей, разделенные границей гари 2014 г. (горевшая (А), негоревшая (Б)), где ежегодно проводились геоботанические описания и укосы надземной фитомассы (Дусаева и др., 2020).
Геоботанические описания выполнялись с использованием стандартных геоботанических методик (Полевая геоботаника, 1964, Ярошенко, 1961, Работнов, 1992). Учет надземной фитомассы проводился методом укосных площадей (Базилевич, Титлянова и др., 1978) в середине месяца с мая по сентябрь с 2015 г. Растения срезались вровень с почвой, на площадках по 0.25 м2 в 3-кратной повторности. Полученные образцы высушивались до воздушно сухой массы. Отбор ветоши в каждой группе и взвешивание образцов осуществлялись в лабораторных условиях. Подстилку отбирали вручную на укосных площадках, после того как с них срезали растения. Пробы очищались от почвы до и после высушивания.
Расчет NDVI для территориальных единиц, соответствовавших по географическим координатам площадкам опробования, проводился по безоблачным космическим снимкам Landsat с разрешением 30 м за сроки, соответствовавшие датам полевых определений надземной фитомассы. Из 25 сроков измерений фитомассы в 2015–2018 и 2020 гг. космическими снимками и расчетами NDVI обеспечено 16, в том числе 3 – для мая, 3 – для июня, 3 – для июля, 4 – для августа, 3 – для сентября.
На основании имеющихся данных средствами программы Statistica 7.0 методами корреляционного и дисперсионного анализа проверялась серия гипотез о геоботанических, гидротермических, фенологических и ландшафтных факторах, определяющих информативность NDVI как индикатора зеленой фитомассы.
Гипотеза 1: агробиологические группы растений вносят неодинаковый вклад в пространственное и временно варьирование NDVI. Инструментом проверки гипотезы был расчет непараметрических коэффициентов корреляции Спирмена между NDVI за каждый срок и фитомассой групп растений – злаков, разнотравья, осок, бобовых, полукустарничков и кустарничков.
Гипотеза 2: теснота связи и вид зависимости между NDVI и фитомассой определяется фазой развития сообщества в годовом цикле. Сравнивались модуль и знак коэффициентов корреляции Спирмена для разных месяцев независимо от конкретного года. Делалось допущение, что пространственное варьирование показателей фитомассы и NDVI в конкретный месяц не зависит от гидротермических условий года.
Гипотеза 3: теснота связи и вид зависимости между NDVI и фитомассой может меняться в зависимости от гидротермических условий года. Ход температур и осадков в течение вегетационного периода с мая по сентябрь, различающийся от года к году, может определять характер связи исследуемых показателей. Сравнивались корреляции, полученные раздельно по годам (2015, 2016, 2017, 2018, 2020) независимо от месяца измерения. Проверялось предположение, что теснота связи, т.е. правило сопряженного пространственного варьирования фитомассы и NDVI, возрастает в годы с некоторым специфической комбинацией тепло- и влагообеспеченности.
Гипотеза 4: теснота связи и вид зависимости между NDVI и фитомассой специфичны для сообщества независимо от сезонной фазы его развития и гидротермических условий года, но определяются “историей” сообщества, т.е. былыми (до установления заповедного режима) способами землепользования и длительностью послепожарной сукцессии. Сравнивались корреляции для отдельных площадок независимо от года и месяца наблюдений с предположением, что варьирование фитомассы и NDVI во времени подчиняется единому правилу независимо от ландшафтной позиции.
Гипотеза 5: теснота связи и вид зависимости между NDVI и фитомассой в каждый момент времени определяются различиями ландшафтной позиции сообществ, сходных по видовому составу, а именно – приуроченностью к выпуклой или плоской мезоформе рельефа, принадлежностью к возвышенной (структурно-эрозионной) или низменной (долинно-балочной) местности, характером почвообразующих пород и гранулометрическим составом почв. Сравнивались корреляции для разных сроков наблюдения с предположением, что пространственное варьирование фитомассы и NDVI в ландшафте определяется одним и тем же правилом независимо от года и месяца наблюдений.
РЕЗУЛЬТАТЫ
Сезонная динамика фракций фитомассы и NDVI
Средние значения показателей надземной фитомассы и NDVI по месяцам показаны в табл. 1. Данные свидетельствуют о несовпадении сезонных пиков NDVI (май), зеленой фитомассы (июнь), ветоши (июль) и подстилки (август). Смещение сезонных пиков на более поздние сроки от живой фитомассы к ветоши и подстилке отражает нормальный для степи ход процесса трансформации фитомассы (Титлянова и др., 2018). Однако несовпадение пика NDVI и зеленой фитомассы не согласуется с распространенным представлением о прямом индикационном значении NDVI, что приводит к необходимости более детального анализа; его результаты приводятся ниже. В частности, в 2015 г. такое несовпадение отмечалось на 3-х площадках из 12, а в 2020 г. – на 2-х (причем, других). Согласно результатам дисперсионного анализа (по тестам LSD, Newman-Keuls, Tukey) все изученные площадки не отличаются по значениям NDVI в каждом месяце с мая по сентябрь 2015–2018 и 2020 гг., что согласуется с их выбором как представителей одного типа растительности и одинаковых или близких типов сообществ в парах площадок. Исключение составляют 5–6 пар площадок (из 144) в мае, июле и августе; в июне и сентябре достоверных отличий не было совсем.
Таблица 1.
Средние по 12 площадкам значения зеленой и мертвой надземной фитомассы и NDVI по месяцам (наблюдения 2015–2018 и 2020 гг.).
| Месяц | Май | Июнь | Июль | Август | Сентябрь |
|---|---|---|---|---|---|
| Кол-во измерений фитомассы | 62 | 61 | 54 | 62 | 62 |
| Средняя общая зеленая фитомасса (G), г/м2 | 106.32 | 139.39 | 106.13 | 89.33 | 62.91 |
| Стандартное отклонение значений фитомассы G, г/м2 | 33.38 | 41.79 | 31.7 | 29.01 | 23.52 |
| Средняя общая масса ветоши (D), г/м2 | 120.9 | 131.24 | 153.05 | 145.92 | 156.83 |
| Средняя масса подстилки (L), г/м2 | 124.7 | 133.01 | 151.02 | 160.89 | 154.18 |
| Кол-во измерений NDVI | 37 | 38 | 38 | 49 | 37 |
| NDVI | 0.56 | 0.43 | 0.33 | 0.28 | 0.23 |
| Стандартное отклонение значений NDVI | 0.05 | 0.09 | 0.05 | 0.03 | 0.03 |
Общая надземная фитомасса, включающая зеленую массу (G), ветошь (D) и подстилку (L), за весь период измерений подчиняется нормальному распределению, равно как и отдельно за каждый месяц при p > 0.10. Это свидетельствует об однородности исследуемой выборки и варьировании растительных сообществ во времени и пространстве под уравновешенным влиянием большой группы факторов, ни один из которых не является главным. Иными словами, сезонные и межгодовые отклонения от “нормативной” фитомассы разнотравно-типчаково-ковыльных степей взаимно компенсируют друг друга. Общая масса ветоши (D) характеризуется нормальным распределением за весь период наблюдений, однако для каждого месяца по отдельности отличия от нормального распределения достоверны. Следовательно, каждый период вегетационного сезона характеризуется неравновесием процесса отмирания зеленой фитомассы фитоценоза и, вероятно, ее потребления консументами и редуцентами (прямыми данными о котором мы не располагаем). Это означает возможность существенных отличий в протекании этого процесса от года к году в зависимости от гидротермических условий. Зеленая фитомасса, ее фракции по агробиологическим группам и масса подстилки нормальному распределению не подчиняются ни в один из месяцев и в целом за период измерений. По этой причине в дальнейшем для статистического анализа использовались непараметрические методы.
Сезонная динамика доли агробиологических групп в фитомассе
По данным 301 измерения на 12 площадках в течение 5 лет (пять раз год) и еще на одной – в течение 2 лет медианное значение доли злаков в зеленой фитомассе – 73% (диапазон от 32 до 96%), а доли разнотравья – всего 12% при максимуме 63%. Зеленая фитомасса в наибольшей степени определяется массой злаков: непараметрический коэффициент корреляции Спирмена (ККс) составляет 0.85. На втором месте по вкладу разнотравье (ККс = 0.62), на третьем – полукустарнички (ККс = 0.37). При этом статистическая связь фитомассы злаков и разнотравья достаточно слабая, хотя и достоверная (ККс = 0.30) Увеличение фитомассы разнотравья резко снижает долю злаков в фитомассе (ККс = –0.67), что, видимо, отражает запаздывание фенофазы наиболее активной вегетации злаков, по сравнению с разнотравьем; в начале сезона это особенно характерно для горевших сообществ. Однако обратное неверно: увеличение фитомассы злаков никак не отражается на доле разнотравья в фитомассе (ККс = 0.02).
Доля запасов живой фитомассы разнотравья в фитоценозе по массе всегда уменьшается от мая, когда в его составе присутствуют эфемеры и эфемероиды (Pedicularis physocalyx Bunge, P. sibirica Vved., Iris pumila L., Gagea pusilla (F.W. Schmidt) Schult. & Schult. f., Tulipa biebersteiniana Schult. & Schult. f., Valeriana tuberosa L.,), играющие особенно значительную роль в сообществах после пожаров, к концу вегетационного сезона (рис. 1). На некоторых площадках (7а, 7б, 5а) слабое увеличение доли разнотравья в некоторые годы отмечалось в сентябре. На площадке 5а (низменная аккумулятивно-денудационная равнина) это происходит за счет вегетации и цветения Taraxacum serotinum (Waldst. & Kit.) Poir., достигающего значительного обилия в сообществе в связи с залежным характером растительного покрова на этом участке. На площадках 7а и 7б (пологонаклонные денудационные увалы) в конце лета и начале осени основу фитомассы разнотравья формирует Galatella villosa (L.) Rchb. f., но абсолютные значения живой фитомассы разнотравья небольшие; увеличение доли разнотравья в общих запасах живой надземной фитомассы определяется существенным снижением запасов живой фитомассы злаков. Доля злаков в фитомассе не обнаруживает однозначной тенденции в течение вегетационного периода, хотя на некоторых площадках (2, 4, 6) она заметно возрастает в сентябре. При этом на указанных площадках запасы фитомассы злаков осенью снижаются или незначительно возрастают (преимущественно за счет Stipa capillata L.), а доля злаков в запасах живой надземной фитомассы увеличивается за счет снижения фитомассы разнотравья, а иногда и полукустарничков.
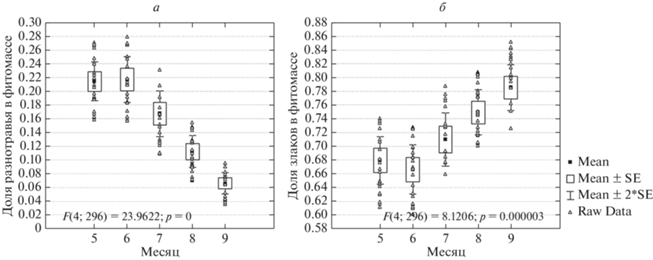
Зависимость NDVI от сезонных изменений состава фитоценоза (гипотеза 1)
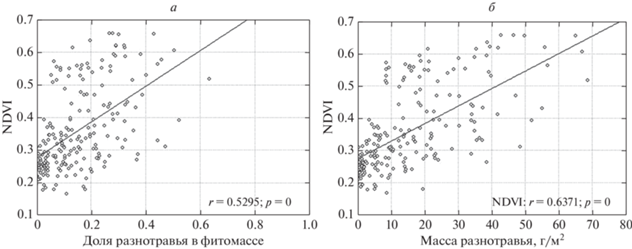
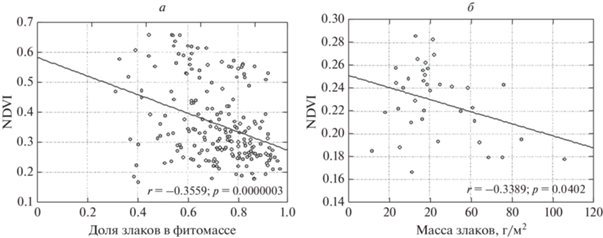
Оценка корреляционной связи по всей совокупности площадок без учета года и сезона показала, что NDVI наиболее достоверно коррелирует с массой разнотравья (ККс = 0.68) (рис. 2) и долей разнотравья в фитомассе (ККс = 0.57), хотя зеленая фитомасса как таковая определяется в основном злаками (ККс = 0.85). Более того, при доле злаков в фитомассе более 0.8 и при большой фитомассе злаков (обычно в августе и сентябре) NDVI имеет тенденцию к снижению: исключаются значения более 0.3 (рис. 3). В связи с этим требует проверки гипотеза о частичном экранировании преобладающих в живой массе злаков ветошью, результаты которой приводятся ниже.
Зависимость информативности NDVI от фазы вегетационного периода (гипотеза 2)
Оценка проведена по совокупности измерений на 13 площадках в течение 5 лет за 16 сроков, обеспеченных безоблачными космоснимками. Выявлены помесячные различия в информативности NDVI как индикатора общей фитомассы и фитомассы отдельных агробиологических групп (табл. 2). Обращает на себя внимание, что единой тенденции роста или снижения в течение года информативности NDVI как показателей фитомассы нет. В мае и июне зависимость NDVI от массы разнотравья более тесная, чем от массы злаков. В июле NDVI определяется в основном массой злаков. В августе эти две группы вносят примерно одинаковые вклады. Несколько более сильная зависимость NDVI от массы разнотравья проявляется в невозможности низких значений NDVI (менее 0.27) при большой массе разнотравья. При малой массе разнотравья (до 15 г/м2), возможен большой диапазон значений NDVI – от 0.22 до 0.35, т.е. вносят вклад другие группы растений, особенно полукустарнички, что характерно в некоторые годы для площадок 2Б, 7Б, 1А, находящихся в разных условиях рельефа.
Таблица 2.
Коэффициент корреляции Спирмена между значениями NDVI и показателями фитомассы по совокупности наблюдений на всех площадках в 2015–2018, 2020 гг. (245 измерений)
| Показатель | Май | Июнь | Июль | Август | Сентябрь |
|---|---|---|---|---|---|
| Общая живая фитомасса G | 0.38 | 0.67 | 0.64 | 0.54 | –0.37 |
| Злаки Z | 0.07 | 0.25 | 0.59 | 0.35 | –0.20 |
| Разнотравье R | 0.61 | 0.54 | 0.18 | 0.40 | –0.27 |
| Бобовые | –0.16 | 0.18 | 0.15 | 0.10 | 0.04 |
| Осоки | 0.29 | 0.14 | 0.10 | 0.13 | –0.12 |
| Полукустарнички | 0.36 | 0.07 | 0.04 | 0.10 | –0.25 |
| Кустарнички | 0.03 | 0.09 | 0.23 | 0.02 | – |
| Z/G | –0.51 | –0.36 | 0.10 | –0.30 | 0.22 |
| R/G | 0.36 | 0.38 | 0.01 | 0.29 | –0.15 |
| Ветошь злаков Dz | –0.01 | –0.20 | 0.08 | –0.12 | 0.30 |
| Ветошь разнотравья Dr | 0.40 | 0.38 | 0.12 | 0.42 | –0.19 |
| Подстилка L | –0.03 | –0.36 | 0.09 | –0.04 | 0.23 |
| Ветошь общая Dt | 0.05 | –0.18 | 0.08 | 0.02 | 0.23 |
| Мортмасса M | 0.04 | –0.25 | 0.13 | –0.01 | 0.25 |
| Фитомасса надземная F = G + V + L | 0.10 | –0.04 | 0.29 | 0.10 | 0.13 |
| Dt/G | –0.12 | –0.53 | –0.22 | –0.28 | 0.51 |
| L/G | –0.16 | –0.53 | –0.26 | –0.28 | 0.43 |
| M/G | –0.17 | –0.55 | –0.32 | –0.30 | 0.51 |
| G/F | 0.17 | 0.55 | 0.32 | 0.30 | –0.51 |
В сентябре NDVI не зависит от массы и доли разнотравья и злаков, но при этом появляется парадоксальная положительная (в отличие от других месяцев) связь с показателями, характеризующими превышение мортмасс (ветоши, подстилки, суммарной мортмассы) над живой массой, что требует отдельного пояснения.
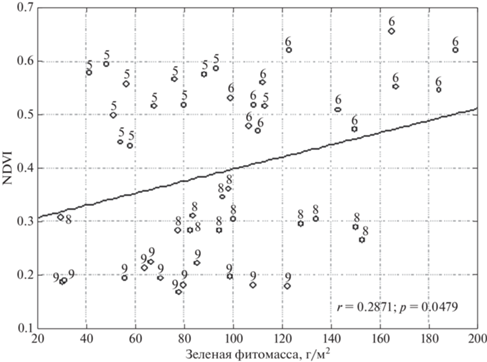
Логика сезонного ритма степей подсказывает обратное: в сентябре усиленное отмирание живой массы и, соответственно, накопление ветоши должно приводить к экранированию фотосинтезирующих органов сухими и снижением NDVI. Однако полученный неожиданный результат целиком обусловлен особенностями фитопродукционного процесса в 2015 г по сравнению с 2017–2018 гг. В 2015 г. все измерения показали низкие NDVI при одновременно малом отношении мортмасс (D, G, M) к живой массе. Иными словами, ветошь не могла оказать большое экранирующее влияние на живые зеленые органы, масса которых была достаточно большой для начала осени благодаря наиболее теплому за 2000–2021 гг. сентябрю (+15.6°). Сравнение структуры фитомассы показало, что в 2015 г., по сравнению с 2017 и 2018 гг., на некоторых площадках была существенно выше масса и доля живых злаков (5А, 5Б, 7Б) и полукустарничков (1А, 1Б, 2А). Однако площадки с высокой (по сравнению с 2017 и 2018 гг.) массой злаков и полукустарничков отличались низкими значениями NDVI. Иными словами, возобновление вегетации злаков (но не разнотравья) в очень теплом сентябре 2015 г. сопровождалось ростом фитомассы, но снижением NDVI. Хорошо видно, что при сопоставимых значениях живой фитомассы (40–80 г/м2) в мае и сентябре 2015 г. (рис. 4), фотосинтетическая активность, приводящая к росту NDVI, была существенно выше весной.
Зависимость информативности NDVI от гидротермических условий (гипотеза 3)
Известна высокая чувствительность NDVI к температурам и осадкам (Рулев и др., 2016). Проверка гипотезы была несколько затруднена неодинаковой представленностью данных NDVI по месяцам из-за отсутствия безоблачных космоснимков близких к сроку наземного измерения фитомассы. Среднемесячные гидротермические показатели (температуры и осадки) ни для одного из пяти месяцев сезона не дали однозначных объяснений повышенных или пониженных корреляций по сравнению с другими годами. Межгодовые различия в информативности NDVI показаны в табл. 3.
Таблица 3.
Непараметрические коэффициенты корреляции Спирмена между показателями фитомассы и NDVI по годам (все площадки, все сезоны)
| Год | 2015 | 2016 | 2017 | 2018 | 2020 |
|---|---|---|---|---|---|
| Месяцы, обеспеченные данными NDVI | 5, 6, 8, 9 | 5, 8 | 7, 8, 9 | 6, 7, 8, 9 | 5, 6, 7 |
| Количество измерений | 48 | 24 | 36 | 51 | 39 |
| ГТК Селянинова за период май–сентябрь | 0.31 | 0.58 | 0.30 | 0.17 | 0.30 |
| Показатели | Коэффициент корреляции | ||||
| Общая живая фитомасса G | 0.23 | 0.47 | 0.76 | 0.82 | 0.78 |
| Злаки Z | 0.00 | –0.05 | 0.72 | 0.79 | 0.72 |
| Разнотравье R | 0.66 | 0.76 | 0.42 | 0.74 | 0.37 |
| Бобовые | 0.33 | –0.20 | 0.37 | 0.15 | 0.29 |
| Осоки | 0.16 | –0.01 | 0.02 | 0.13 | 0.08 |
| Полукустарнички | –0.11 | –0.19 | 0.03 | 0.23 | 0.04 |
| Кустарнички | –0.06 | 0.14 | 0.25 | Н.д. | 0.27 |
| Доля злаков в живой фитомассе Z/G | –0.42 | –0.31 | 0.00 | –0.34 | 0.01 |
| Доля разнотравья в живой фитомассе R/G | 0.63 | 0.66 | 0.25 | 0.65 | 0.09 |
| Доля полукустарничков в живой фитомассе | –0.17 | –0.24 | –0.26 | –0.19 | –0.18 |
| Ветошь злаков Dz | –0.17 | –0.05 | 0.29 | 0.02 | –0.29 |
| Ветошь разнотравья Dr | –0.31 | 0.29 | 0.08 | 0.12 | –0.26 |
| Ветошь бобовых | –0.29 | –0.16 | 0.07 | –0.04 | 0.12 |
| Ветошь осок | –0.11 | 0.08 | –0.13 | –0.03 | 0.09 |
| Ветошь полукустарничков | –0.23 | –0.36 | –0.25 | –0.12 | –0.14 |
| Подстилка L | –0.17 | –0.24 | –0.09 | 0.11 | 0.06 |
| Ветошь общая Dt | –0.18 | –0.08 | 0.24 | 0.01 | –0.30 |
| Мортмасса M | –0.29 | –0.26 | 0.06 | 0.12 | –0.06 |
| Фитомасса надземная F = G + D + L | –0.12 | –0.09 | 0.28 | 0.33 | 0.20 |
| Dt/G | –0.40 | –0.25 | –0.58 | –0.69 | –0.71 |
| L/G | –0.21 | –0.33 | –0.60 | –0.59 | –0.44 |
| M/G | –0.36 | –0.29 | –0.66 | –0.68 | –0.60 |
| G/F | 0.33 | 0.29 | 0.66 | 0.68 | 0.60 |
| Коэффициент детерминации мультирегрессионного уравнения | |||||
| G, Z, R, Dt, L, M | 0.40 | 0.58 | 0.70 | 0.73 | 0.62 |
Из всех показателей живой фитомассы наиболее устойчивой от года к году оказалась положительная связь NDVI с массой разнотравья, хотя с долей разнотравья связь проявляется не каждый год. Зависимость NDVI от общей живой массы и массы злаков положительна, но не каждый год достоверна. Информативность NDVI как индикатора живой фитомассы и ее соотношений с мортмассами оказалась наиболее высокой в самый засушливый 2018 г. В наиболее влажный и теплый 2016 г. оказались более слабыми связи NDVI с живой фитомассой и недостоверными – с показателями мортмасс, однако нет достаточных оснований объяснять это именно гидроклиматическими условиями, поскольку мы располагаем данными NDVI только за май и август.
Устойчивы от года к году положительная связь NDVI с долей зеленой массы в общей надземной фитомассе и, наоборот, отрицательные связи NDVI с показателями, характеризующими отношение мортмасс (ветоши, подстилки, общей) к зеленой массе. Эти факты иллюстрируют зависимость NDVI не только от живой фитомассы как таковой, но и от степени перекрытия зеленых органов отмершими (прежде всего – ветошью), которая варьирует по месяцам. Поэтому возникает необходимость установить, бывают ли случаи, когда NDVI и живая фитомасса меняются за временной интервал в противоположных направлениях (что противоречит ожидаемой закономерности) и можно ли это объяснить экранированием зеленых органов ветошью или (для низких растений) подстилкой.
Для каждого года было составлены мультирегрессионные линейные модели методом прямого пошагового выбора (Forward stepwise regression, F = 1), где зависимая переменная – NDVI, независимые переменные – G, R, Z, Dt, L, M (см. табл. 2, 3). Они показали, что во все годы значения NDVI описываются значениями либо живой фитомассы, либо массы разнотравья, либо совместным эффектом массы разнотравья и массы ветоши (либо общей мортмассы). Максимальное значение доли объясненной дисперсии (коэффициент детерминации R^2) составило 0.73 в 2018 г. (табл. 4), минимальное – 0.40 в 2015 г. В 2018 и 2020 гг. при близких значениях живой массы разнотравья (основного фактора NDVI, как было показано выше) увеличение массы ветоши сопровождалось снижением NDVI. При одинаковой зеленой фитомассе накопление ветоши также способствует снижению NDVI, например, в 2020 г. – в июне по сравнению с маем, в 2016 г. – в августе по сравнению с маем, в 2015 г. – в сентябре по сравнению с маем.
Таблица 4.
Мультирегрессионное уравнение зависимости NDVI от показателей фитомассы в 2018 г. (май–сентябрь, 13 площадок). Метод прямого пошагового выбора при F = 1. Достоверные коэффициенты выделены жирным шрифтом. Коэффициент детерминации R^2 = 0.73, Подправленный R^2 = 0.71. F = 31.624, p = 0.00000. Beta – стандартизованный регрессионный коэффициент; Std.Err. Beta – ошибка коэффициента Beta; Std.Err. В – регрессионный коэффициент; B – ошибка коэффициента B, t – критерий Стьюдента; p – уровень значимости
| Beta | Std.Err. Beta | B | Std.Err. B | t | p-level | |
|---|---|---|---|---|---|---|
| Свободный член уравнения | 0.24 | 0.01 | 16.09 | 0.00 | ||
| Живая фитомасса | 0.19 | 0.42 | 0.00 | 0.00 | 0.45 | 0.66 |
| Живая масса разнотравья | 0.40 | 0.16 | 0.00 | 0.00 | 2.45 | 0.02 |
| Живая масса злаков | 0.43 | 0.35 | 0.00 | 0.00 | 1.23 | 0.22 |
| Суммарная масса ветоши | –0.28 | 0.08 | –0.00 | 0.00 | –3.40 | 0.00 |
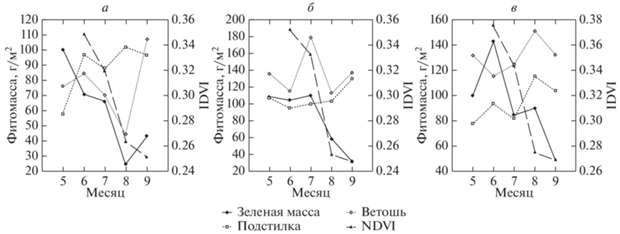
При общем правиле положительной корреляции между зеленой фитомассой и NDVI, массой разнотравья и NDVI обнаружены эпизодические отклонения от этого правила, которые во всех случаях объясняются экранирующей ролью ветоши. Так, в 2018 г. на т. 1А и 1Б падение NDVI от августа к сентябрю происходило несмотря на рост зеленой фитомассы, но сопряженно с ростом массы ветоши, которая, вероятно, экранировала зеленую фитомассу (рис. 5, а). На т. 2А тот же эффект наблюдался от июня к июлю (рис. 5, б), на т. 4А и 4Б от июля к августу (рис. 5, в).
Рис. 5.
Сезонная динамика живой фитомассы (FM_green), массы подстилки (Litter), массы ветоши (V_SUM) и NDVI в 2018 г. 5, 6, 7, 8, 9 – номера месяцев. а – площадка 1А (низменное пологоувалистое днище балки). б – площадка 2А (возвышенная слабонаклонная приводораздельная поверхность). в – площадка 4А (низменная плоская денудационно-аккумулятивная равнина).
Зависимость информативности NDVI от индивидуальных особенностей сообщества конкретного урочища (гипотеза 4)
Рассчитывались корреляции для каждой модельной площадки отдельно. Согласно результатам, приведенным в табл. 5, общая закономерность сохраняется, подтверждается большими по модулю значениями коэффициентов и является единой для большинства площадок. Основной вклад в значения NDVI на любой площадке вносит масса разнотравья, вклад злаков всегда меньше. Разница в тесноте связей на горевших и негоревших площадках наиболее четко проявляется в урочищах приводораздельных поверхностей возвышенных равнин (2А и 2Б, 6А и 6Б): на негоревших площадках (с индексом Б) связи NDVI являются более надежным индикатором соотношения живой и мертвой массы, чем на горевших. На остальных площадках это различие тоже есть, но очень незначительное. Рост доли ветоши и подстилки в надземной фитомассе сопровождается снижением значений NDVI. Из общего правила четко выделяются слабыми корреляциями площадки 5А и 5Б, которые, в отличие от остальных, расположены на залежном участке и относятся к ковылковой (Stipeta lessingianae) формации. Фитомасса злаков оказывает наибольшее влияние на NDVI (обычно за счет июня-июля) на площадках 7А и 7Б. Эти площадки отличается наибольшим разнообразием ковылей: Stipa capillata, S. lessingiana, S. pulcherrima, S. zalesskii. Помимо ковылей в сообществах встречаются другие плотнодерновинные (Festuca valesiaca Koeleria cristata) и рыхлодерновинные (Poa transbaicalica, Agropyron pectinatum) злаки. Эти площадки расположены у северо-восточной границы заповедника на пологовыпуклом увале северного простирания и отличаются наименьшей массой и долей разнотравья по сравнению с другими площадками. Однозначной закономерности различий тесноты связи между горевшими и негоревшими площадками не обнаружено.
Таблица 5.
Непараметрические коэффициенты корреляции Спирмена между характеристиками фитомассы и NDVI для 12 площадок (май–сентябрь, 2015–2020 гг.). Буквенный индекс “А” соответствует горевшим площадкам, индекс “Б” – не испытавшим пожарного воздействия в 2014 г.
| 1А | 1Б | 2А | 2Б | 4А | 4Б | 5А | 5Б | 6А | 6Б | 7А | 7Б | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Общая живая фитомасса G | 0.59 | 0.62 | 0.71 | 0.74 | 0.66 | 0.64 | 0.34 | 0.21 | 0.69 | 0.77 | 0.82 | 0.64 |
| Злаки Z | 0.44 | 0.56 | 0.59 | 0.49 | 0.54 | 0.30 | 0.33 | 0.15 | 0.38 | 0.51 | 0.66 | 0.61 |
| Разнотравье R | 0.80 | 0.76 | 0.69 | 0.70 | 0.80 | 0.83 | 0.29 | 0.62 | 0.84 | 0.86 | 0.70 | 0.81 |
| Бобовые | –0.10 | 0.12 | 0.62 | 0.33 | 0.07 | 0.01 | 0.28 | 0.43 | 0.50 | 0.78 | 0.06 | 0.17 |
| Осоки | 0.49 | 0.29 | 0.26 | 0.36 | 0.08 | –0.42 | – | –0.24 | 0.35 | 0.34 | –0.02 | – |
| Полукустарнички | –0.16 | –0.11 | 0.18 | 0.59 | –0.30 | –0.32 | –0.38 | –0.11 | 0.50 | 0.60 | 0.14 | 0.06 |
| Кустарнички | 0.14 | 0.36 | – | – | – | – | – | – | – | 0.31 | 0.42 | 0.14 |
| Ветошь злаков Dz | –0.14 | 0.05 | –0.28 | –0.05 | –0.29 | –0.53 | –0.17 | –0.12 | –0.14 | –0.27 | –0.27 | –0.05 |
| Ветошь разнотравья Dr | –0.16 | 0.07 | –0.58 | 0.02 | 0.14 | –0.64 | –0.23 | –0.20 | –0.34 | –0.16 | –0.50 | –0.46 |
| Ветошь бобовых | –0.27 | –0.06 | –0.27 | –0.04 | –0.25 | 0.03 | –0.33 | 0.04 | –0.25 | –0.22 | –0.06 | –0.37 |
| Ветошь осок | 0.06 | –0.41 | –0.20 | –0.13 | 0.08 | –0.20 | – | – | –0.30 | –0.10 | –0.18 | – |
| Ветошь полукустарничков | –0.31 | 0.18 | –0.46 | 0.11 | –0.40 | –0.27 | –0.47 | –0.04 | –0.44 | –0.64 | –0.29 | –0.24 |
| Подстилка L | –0.44 | –0.40 | –0.48 | –0.28 | –0.22 | –0.14 | –0.34 | –0.19 | –0.11 | –0.26 | –0.13 | –0.15 |
| Ветошь общая Dt | –0.22 | 0.08 | –0.35 | –0.11 | –0.32 | –0.64 | –0.23 | –0.18 | –0.15 | –0.44 | –0.27 | –0.05 |
| Мортмасса M | –0.31 | –0.20 | –0.47 | –0.25 | –0.31 | –0.31 | –0.28 | –0.18 | –0.15 | –0.42 | –0.28 | –0.14 |
| Фитомасса F = G + V + L | –0.08 | 0.14 | –0.12 | 0.06 | –0.04 | –0.24 | –0.23 | –0.11 | –0.06 | –0.21 | 0.02 | 0.36 |
| Доля злаков Z/G | –0.22 | 0.16 | –0.36 | –0.55 | –0.52 | –0.76 | 0.04 | –0.35 | –0.79 | –0.79 | –0.38 | –0.13 |
| Доля разнотравья R/G | 0.67 | 0.57 | 0.47 | 0.60 | 0.70 | 0.76 | 0.25 | 0.68 | 0.74 | 0.73 | 0.57 | 0.74 |
| Доля бобовых | –0.11 | 0.10 | 0.61 | 0.26 | 0.03 | 0.00 | 0.20 | 0.28 | 0.46 | 0.64 | 0.04 | 0.07 |
| Доля полукустарничков | –0.35 | –0.47 | –0.04 | 0.33 | –0.47 | –0.40 | –0.58 | –0.15 | 0.23 | 0.31 | –0.16 | –0.38 |
| Dt/G | –0.58 | –0.51 | –0.74 | –0.79 | –0.76 | –0.87 | –0.39 | –0.36 | –0.59 | –0.81 | –0.69 | –0.58 |
| L/G | –0.56 | –0.76 | –0.59 | –0.64 | –0.60 | –0.46 | –0.44 | –0.43 | –0.50 | –0.70 | –0.42 | –0.55 |
| M/G | –0.60 | –0.71 | –0.65 | –0.82 | –0.78 | –0.65 | –0.43 | –0.43 | –0.58 | –0.81 | –0.58 | –0.61 |
| G/F | 0.60 | 0.71 | 0.65 | 0.82 | 0.78 | 0.65 | 0.43 | 0.43 | 0.58 | 0.81 | 0.58 | 0.61 |
Зависимость информативности NDVI от ландшафтной позиции сообществ (гипотеза 5)
Предполагалось, что различия фракций фитомассы между аналогичными сообществами разных условий рельефа и микроклимата могут в той или иной степени отражаться вегетационными индексами.
Коэффициенты корреляции Спирмена, полученные по выборке 13 площадок отдельно для каждого срока, показывают, на первый взгляд, парадоксальные результаты. Для некоторых майских, июньских и июльских сроков получены положительные корреляции (0.58–0.74) NDVI с показателями мортмасс (ветошь, подстилка, общая мортмасса) и их отношениями к живой фитомассе. По смыслу вегетационного индекса такого не должно быть: NDVI должен увеличиваться с ростом массы зеленых органов и уменьшаться – с ростом массы отмерших органов, которые к тому же, как показано выше, могут маскировать живые листья низких растений. Однако обращает на себя внимание, что для тех же сроков зафиксированы положительные достоверные корреляции (0.64–0.80) с общей живой фитомассой и/или массой живого разнотравья и злаков. Иначе говоря, чем больше зеленой фитомассы накопило степное сообщество к конкретному сроку наблюдения, тем больше оно содержит и мортмасс. Достоверные связи между живой фитомассой и NDVI определяются различиями сообществ по положению в рельефе. Исследованные площадки образуют три группы. Наиболее высокие значения фитомассы и NDVI характерны для сообществ возвышенных (380–410 м) увалов и приводораздельных поверхностей (тт. 6А, 6Б, 2А, 2Б). Самые низкие значения живой фитомассы и NDVI в разные месяцы и годы обычно характерны для сообществ низменных пологоувалистых равнин с наследием пахотного режима (тт. 5А, 5Б) и увалистого днища балки (тт. 1А, 1Б). Возможно, этот факт отражает негативное влияние стекания холодного воздуха в понижения с высотой 260–280 м на фитопродукционный процесс. Третью группу образуют площадки низменных равнин с наследием пастбищной дигрессии (тт. 4А, 4Б) и пологой поверхности увала (тт. 7А, 7Б), которые имеют промежуточные характеристики.
В сентябре наблюдается ослабление информативности NDVI как индикатора различий фитомассы на площадках. Это объясняется, видимо, минимальной вариабельностью (оцененной по среднеквадратическому отклонению) NDVI и живой надземной фитомассы между площадками по сравнению с другими месяцами. Фактически, по этим двум показателям модельные площадки в сентябре ничем не отличаются друг от друга. В целом за год пространственная вариабельность NDVI в группе модельных площадок убывает от мая и июня к сентябрю. В то же время для вариабельности живой надземной фитомассы как таковой сезонная тенденция отсутствует. По значениям среднеквадратического отклонения выделяется только июнь: вариабельность больше, чем в мае и сентябре, но не отличается от вариабельности в июле и августе.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Исследование показало, что NDVI как индикатор зеленой фитомассы определяется соотношением массы агробиологических групп. Вклад этих групп в фитомассу определяется различием в сезонной динамике запасов фитомассы злаков и разнотравья, а также с некоторой ролью полукустарничков. При этом если сезонный тренд запасов живой фитомассы злаков и разнотравья нередко совпадает, то абсолютные значения и доли, напротив обычно существенно различаются. Полукустарнички во второй половине лета поддерживают формируемую за счет разнотравья тенденцию к снижению доли злаков в общих запасах живой надземной фитомассы. В ходе сезонной динамики запасы фитомассы разнотравья и полукустарничков либо достигают наиболее высоких значений одновременно, совместно уменьшая долю злаков в укосах, либо компенсируют пиковые значения запасов фитомассы друг друга. Увеличение запасов фитомассы полукустарничков, активный прирост которых приходится на летнее время (Astragalus macropus Bunge, Eremogone koriniana (Fisch. ex Fenzl) Ikonn., Onosma simplicissima L., Thymus marschallianus Willd.) позволяет сохранять эту закономерность при увеличении запасов живой фитомассы злаков или некотором снижении живой фитомассы разнотравья. В позднелетний и осенний периоды значения запасов живой фитомассы полукустарничков, активный прирост которых приходится на это время (Artemisia austriaca Jacq., A. marschalliana Spreng.) достигают максимальных пиковых значений, в то время как запасы живой фитомассы разнотравья минимальны. Таким образом, полукустарнички в конце сезона принимают на себя роль “регулятора” доли злаков в общих запасах живой надземной фитомассы. Это подтверждается статистически значимой корреляцией долей злаков и полукустарничков (ККс = –0.49).
Вегетационный индекс NDVI оказался весьма чувствительным не столько к запасам зеленой фитомассы как таковой (которые в изученном степном ландшафте контролируется, главным образом, злаками), сколько к массе и доле разнотравья и соотношению живой и мертвой фитомассы. На площадках с наибольшими статистически значимыми показателями корреляции NDVI и живой фитомассы разнотравья в этой группе наиболее обильны: Achillea nobilis, Artemisia armeniaca, Centaurea marschalliana, Dianthus campestris, Falcaria vulgaris, Galatella villosa, Galium octonarium, G. ruthenicum, Medicago romanica, Oxytropis spicata, Potentilla humifusa, P. orientalis, Salvia stepposa, Scorzonera stricta. На горевших площадках в первые годы после пожара велико обилие Sisymbrium polymorphum, Tulipa biebersteiniana, Scorzonera austriaca, Ferula tatarica, Eremogone biebersteinii, Scorzonera stricta.
Следует принимать во внимание, что информативность NDVI варьирует в течение теплого периода. Этот результат согласуется с литературными данными по фитомассе степной растительности (Немцева, 2018). Общая живая фитомасса более четко коррелирует с индексом в июне и июле, слабее – в конце сезона. В поздневесеннее и раннелетнее время NDVI наиболее тесно связан с разнотравьем. Это объясняется тем, что в этот период большинство видов разнотравья характеризуются наиболее активной вегетацией и цветением. В мае в составе разнотравья присутствуют эфемеры и эфемероиды, дающие значительную часть фитомассы этой группы, особенно на недавно выгоревших участках. В июле, несмотря на наступление закономерной для степей паузы вегетации, основная фотосинтетическая активность, отражаемая NDVI, приходится на злаки. В августе и сентябре функцию поддержания фитомассы при слабом развитии разнотравья берут на себя полукустарнички, переходящие в фазу цветения и всплеска вегетации. Итак, NDVI может принимать большие значения либо при высокой доле разнотравья, либо при высокой доле полукустарничков.
Наши данные показали, что не всякий рост зеленой фитомассы приводит к росту NDVI, а лишь соответствующий определенной фазе вегетационного периода. В 2015 г. при одинаковой фитомассе NDVI имело более высокие значение в мае, чем в сентябре, который отличался высокой температурой и влажностью и активной вегетацией злаков. Для пшеницы также отмечалось, что одни и те же значения NDVI могут соответствовать разной площади ассимиляционной поверхности в зависимости от фазы развития (Гулянов, 2019). Следует отметить, что не только на исследуемых площадках, но и во всем ландшафте междуречья Урты-Бурти и Бурли (в том числе в заповедной Буртинской степи) в сентябре 2015 г. была повышена встречаемость низких значений NDVI (0.1–0.2) по сравнению с другими годами десятилетия, за исключением экстремально жаркого 2010 г. и экстремально холодного сентября 2019 г. (+10.6°).
В целом подтвердилась гипотеза о возможности экранирования зеленой массы ветошью, что приводит к снижению NDVI несмотря на отсутствие уменьшения зеленой фитомассы. Это далеко не всегда является общим правилом; скорее, свидетельствует о возможности (но необязательности) искажений ожидаемых положительных корреляций между NDVI и живой фитомассой (прежде всего – разнотравья, в меньшей степени – злаков), особенно в сентябре.
Таким образом, приведенные результаты показывают, что NDVI отражает не столько реальную зеленую фитомассу, сколько информацию о балансе производства новой зеленой массы и производства мортмассы. Вегетационные индексы NDVI могут занижать реальную зеленую фитомассу, если происходит резкий прирост массы ветоши, обычно – во второй половине лета и начале осени, но иногда и в июне. Немаловажное влияние оказывает история развития фитоценозов: для фитоценозов залежей и бывших гарей индекс NDVI существенно менее информативен как индикатор живой фитомассы, чем для ненарушенных участков. На более низкую информативность вегетационных индексов на залежных участках степей, по сравнению с коренными сообществами имеются указания и в литературе (Аюржанаев и др., 2021).
Анализ состава и структуры фитоценозов показал, что каждое сообщество использует свои механизмы для достижения необходимого уровня живой надземной фитомассы. Так, площадки, поддерживающие стабильно высокую (относительно других) фитомассу (тт. 6А, 6Б, 2А, 2Б), характеризуются всегда большим, чем на других площадках количеством видов полукустарничков, присутствием плотнодерновинного злака Helictotrichon desertorum (Less.) Nevski, наибольшими значениями проективного покрытия Koeleria cristata (L.) Pers. По-видимому, менее стабильно на поддержание высокого уровня живой фитомассы влияет присутствие в составе сообществ рыхлодерновинного злака – Agropyron pectinatum (M. Bieb.) P. Beauv., высокое обилие которого характерно для участков 4 и 7, повышенное обилие Stipa capillata L., свойственное растительным сообществам на участках 4, 6, 7, присутствие корневищных злаков стабильно отмеченное в фитоценозах площадок 4А, 4Б и 6А и в отдельные годы – 2Б и 7Б. При этом относительно высокое разнообразие разнотравья (по количеству видов), а также высокая доля его в формировании общего проективного покрытия фитоценоза характерны как для площадок со стабильно высокими показателями запасов живой фитомассы и NDVI (6А, 6Б), так и с промежуточными (7А, 7Б) и низкими (1Б). Сходная закономерность наблюдается и при рассмотрении отдельно многолетнего разнотравья. Присутствие однолетников и двулетников, видимо, определяет низкие значения живой фитомассы и NDVI. Большее их количество и проективное покрытие на участках 1, 4, 7.
Геоморфологические и микроклиматические факторы пространственного варьирования NDVI наиболее ярко проявляются в период нарастания фитомассы, а с наступлением летней паузы вегетации различия фитопродукционного функционирования между площадками сглаживаются. Иными словами, различия между площадками, обусловленные ландшафтными факторами, наиболее ярко проявляются в июне, т.е. в период максимального накопления фитомассы.
Приведенные факты доказывают, что запасы живой надземной фитомассы и NDVI несут несколько разную информацию. NDVI индицирует не столько зеленую фитомассу как таковую, сколько ее соотношение с мортмассой, прежде всего – с ветошью. Имеющиеся временные ряды данных, характеризующих интенсивность процессов образования и минерализации фитомассы в горевших и негоревших сообществах позволяют на следующих этапах исследования проверить гипотезу о связи между NDVI и параметрами продукционного процесса.
ВЫВОДЫ
1. В степи с заповедным режимом NDVI отражает отношение живой надземной массы к надземной мортмассе, преимущественно ветоши, а не живую надземную фитомассу. Большая масса высокой ветоши может занижать значения NDVI как индикатора живой фитомасы.
2. Хотя основную надземную массу составляют злаки, основной вклад в формирование значения NDVI среди агробиологических групп вносит надземная масса живого разнотравья.
3. Информативность NDVI как индикатора надземной фитомассы меняется по сезонам года и уменьшается в раннеосенний период.
4. NDVI более адекватно отражает состояние надземной фитомассы степных сообществ, длительное время не подвергавшихся воздействию пожаров, по сравнению с горевшими сообществами.
Список литературы
Аюржанаев А.А., Алымбаева Ж.Б., Жарникова М.А., Содномов Б.В. О возможности оценки надземной фитомассы степной растительности с помощью цветных вегетационных индексов (по данным съемки с беспилотных систем) // Вестник Бурятского государственного университета. Биология, география. 2021. № 4. С. 45–53.
Базилевич Н.И., Титлянова А.А., Смирнов В.В., Родин Л.Е., Нечаева Н.И., Левин Ф.И. Методы изучения биологического круговорота в различных природных зонах. Москва: Мысль, 1978. 185 с.
Гопп Н.В., Нечаева Т.В., Савенков О.А., Смирнова Н.В., Смирнов В.В. Оценка влияния мезорельефа склона на пространственную изменчивость свойств почвы и характеристики растительного покрова по данным дистанционного зондирования Земли // Исслед. Земли из космоса. 2016. № 3. С. 66–74.
Гулянов Ю.А. Мониторинг фитометрических параметров с использованием инновационных методов сканирования посевов // Таврический вестник аграрной науки. 2019. № 3(19). С. 64–76. https://doi.org/10.33952/2542-0720-2019-3-19-64-76
Дусаева Г.Х., Калмыкова О.Г., Дусаева Н.В. Влияние пожара на динамику подземной фитомассы степных фитоценозов на участке “Буртинская степь” Оренбургского заповедника // Экосистемы. 2020. № 24(54). С. 83–92.
Дусаева Г.Х., Максутова Н.В. Сезонная динамика запасов надземной фитомассы в разнотравно-овсецово-типчаково-залесскоковыльном сообществе с Poa transbaicalica и Spiraea crenata // Вестник Оренбургского государственного университета. 2017. № 11(211). С. 79–83.
Дусаева Г.Х., Максутова Н.В., Калмыкова О.Г. Сезонная динамика надземной фитомассы разнотравно-овсецово-типчаково-залесскоковыльного сообщества // Сохранение разнообразия растительного мира Тувы и сопредельных регионов Центральной Азии: история, современность, перспективы. Кызыл: ТувИКОПР СО РАН, 2016. С. 67–69.
Елсаков В.В. Спутниковая съемка в оценке продуктивности экосистем Европейского Севера // Современные проблемы дистанционного зондирования Земли из космоса. 2012. Т. 9. № 1. С. 71–79.
Елсаков В.В., Телятников М.Ю. Межгодовые изменения индекса NDVI на территории европейского северо-востока России и западной Сибири в условиях климатических флуктуаций последних десятилетий // Современные проблемы дистанционного зондирования Земли из космоса. 2013. Т. 10. № 3. С. 260–271.
Ерошенко Ф.В., Барталев С.А., Лапенко Н.Г., Самофал Е.В., Сторчак И.Г. Возможности дистанционной оценки состояния и степени деградации природных кормовых угодий // Современные проблемы дистанционного зондирования Земли из космоса. 2018. Т. 15. № 7. С. 53–66
Жуков А.В., Кунах О.Н., Задорожная Г.А., Андрусевич Е.В. Ландшафтная экология как основа пространственного анализа продуктивности агроценозов // Ecology and noospherology. 2013. V. 24. № 1–2.
Залиханов М.Ч., Коломыц Э.Г., Шарая Л.С., Цепкова Н.Л., Сурова Н.А. Высокогорная геоэкология в моделях. М.: Наука, 2010. 487 с.
Золотокрылин А.Н., Черенкова Е.А., Титкова Т.Б. Аридизация засушливых земель Европейской части России и связь с засухами // Изв. Российской академии наук. Серия географическая. 2020. № 2. С. 207–217. https://doi.org/10.31857/S258755662002017X
Калмыкова О.Г. О растительном покрове Госзаповедника “Оренбургский” // Известия Самарского научного центра Российской академии наук. 2012. Т. 14. № 1(4). С. 1024–1026.
Красная книга почв Оренбургской области. Екатеринбург, 2001. 295 с.
Курганович К.А., Голятина М.А. Пространственно-временной отклик NDVI на изменение климатических характеристик в Забайкальском крае за период 2000–2014 гг. // Вестник ЗабГУ 2015. № 09(124). С. 10–20.
Лиджиева Н.Ц., Уланова С.С., Федорова Н.Л. Опыт применения индекса вегетации (NDVI) для определения биологической продуктивности фитоценозов аридной зоны на примере региона Черные земли // Известия Саратовского университета. 2012. Т. 12. Сер. Химия. Биология. Экология. Вып. 2. Научный отдел 94.
Максутова Н.В., Дусаева Г.Х., Калмыкова О.Г. Сезонная динамика надземной фитомассы разнотравно-типчаково-красивейшековыльно-залесскоковыльного сообщества // Сохранение разнообразия растительного мира Тувы и сопредельных регионов Центральной Азии: история, современность, перспективы. Кызыл: ТувИКОПР СО РАН, 2016. С. 77–80.
Михайлов В.В., Спесивцев А.В., Соболевский В.А. Многомодельное оценивание динамики фитомассы растительных сообществ тундры на основе спутниковых снимков // Исследование Земли из космоса. 2021. № 2. С. 15–30.
Немцева Л.Д., Беспалова Л.А., Голубева Е.И., Михайлов С.И. Оценка состояния растительного покрова сухостепных ландшафтов в условиях выпаса с применением методов дистанционного зондирования Земли // Труды ЮНЦ РАН. Т. VII. 2018. С. 151–164.
Полевая геоботаника. Под общ. ред. Е.М. Лавренко и А.А. Корчагина. М.-Л.: Наука, 1964. Т. 3. 442 с.
Работнов Т.А. Фитоценология. Москва: Изд-во МГУ, 1992. 350 с.
Рулев А.С., Канищев С.Н., Шинкаренко С.С. Анализ сезонной динамики NDVI естественной растительности Заволжья Волгоградской области // Современные проблемы дистанционного зондирования Земли из космоса. 2016. Т. 13. № 4. С. 113–123.
Савин И.Ю., Танов Э.Р., Харзинов С. Использование вегетационного индекса NDVI для оценки качества почв пашни (на примере Баксанского района Кабардино-Балкарии) // Бюллетень Почвенного института им. В.В. Докучаева. 2015. Вып. 77. С. 51–65.
Сафронова И.Н., Калмыкова О.Г. Вопросы зональности и роль заповедников в их решении // Изв. Самарского научного центра Российской академии наук. 2012. Т. 14. № 1(6). С. 1638-1641.
Титлянова А.А., Базилевич Н.И., Шмакова Е.И., Снытко В.А., Дубынина С.С., Магомедова Л.Н., Нефедьева Л.Г., Семенюк Н. В., Тишков А.А., Ти Тран, Хакимзянова Ф.И., Шатохина Н.Г., Кыргыс Ч.О., Самбуу А.Д. Биологическая продуктивность травяных экосистем. Географические закономерности и экологические особенности. Новосибирск: ИПА СО РАН, 2018. 110 с.
Ярошенко П.Д. Геоботаника. Москва–Ленинград: Изд-во Акад. Наук СССР, 1961. 474 с.
Araya S., Ostendorf B., Lyle G., Lewis M. Remote sensing derived phenological metrics to assess the spatio-temporal growth variability in cropping fields // Advances in Remote Sensing. V. 6. 2017. P. 212–228.
de Jong R., Verbesselt J., Schaepman M.E., Bruin S.D. Trend changes in global greening and browning: contribution of short-term trends to longer-term change // Glob. Chang. Biol. V. 18(2). 2012. P. 642–655.
Dusaeva G.Kh., Kalmykova O.G., Dusaeva N.V. Fire influence on dynamics of above-ground phytomass in steppe plant communities in the Burtinskaya steppe (Orenburg state nature reserve, Russia) // Nature Conservation Research. 2019. V. 4. № S1. P. 78–92. https://doi.org/10.24189/ncr.2019.050
Gamon J.A., Huemmrich K.F., Stone R.S., Tweedie C.E. Spatial and temporal variation in primary productivity (NDVI) of coastal Alaskan tundra: Decreased vegetation growth following earlier snowmelt. Remote Sensing of Environment. V. 129. 2013. P. 144–153.
Gao Y., Huang J., Li S., Li Sh. Spatial pattern of non-stationarity and scale-dependent relationships between NDVI and climatic factors – A case study in Qinghai-Tibet Plateau, China // Ecological Indicators. V. 20. 2012. P. 170–176.
Han J.C., Huang Y., Zhang H., Wu X. Characterization of elevation and land cover dependent trends of NDVI variations in the Hexi region, northwest China // J. Environmental Management. V. 232. 2019. P. 1037–1048.
Lyle G., Lewis M., Ostendorf B. Testing the temporal ability of Landsat imagery and precision agriculture technology to provide high resolution historical estimates of wheat yield at the farm scale // Remote Sensing. V. 5. 2013. P. 1549–1567.
Maynard J.J., Levi, M.R. Hyper-temporal remote sensing for digital soil mapping: characterizing soil-vegetation response to climatic variability // Geoderma. V. 285. 2017. P. 94–109.
Nagy A., Fehér J., Tamás T. Wheat and maize yield forecasting for the Tisza river catchment using MODIS NDVI time series and reported crop statistics // Computers and Electronics in Agriculture. V. 151. 2018. P. 41–49.
Peng W., Kuang T., Tao T. Quantifying influences of natural factors on vegetation NDVI changes based on geographical detector in Sichuan, western China // J. Cleaner Production. V. 233. 2019. P. 353–367.
Piao S.L., Nan H.J., Huntingford C., Ciais P., Friedingstein P., Sitch S., Peng S.S., Ahlstrom A., Canadell J.G., Cong N., Levis S., Levy P.E., Liu L.L., Lomas M.R., Mao J.F., Myneni R.B., Peylin P., Poulter B., Shi X.Y., Yin G.D., Viovy N., Wang T.,Wang X.H., Zaehle S., Zeng N., Zeng Z.Z., Chen A.P. Evidence for a weakening relationship between interannual temperature variability and northern vegetation activity // Nat. Commun. V. 5. 2014. 5018.
Piedallu C., Chéret V., Denux J.P., Perez V., Azcona J.S., Seynave I., Gégout J.C. Soil and climate differently impact NDVI patterns according to the season and the stand type // Science of The Total Environment. V. 651. P. 2. 2019. P. 2874–2885.
Verhulst N., Govaerts B. The normalized difference vegetation index (NDVI) GreenSeekerTM handheld sensor: Toward the integrated evaluation of crop management. Part A: Concepts and case studies. Mexico, D.F.; CIMMYT. 2010.
Zhao Z., Gao J., Wang Y., Liu J., Li S. Exploring spatially variable relationships between NDVI and climatic factors in a transition zone using geographically weighted regression // Theoretical and Applied Climatology. V. 120(3–4). 2015. P. 507–519 https://doi.org/10.1007/s00704-014-1188-x
Дополнительные материалы отсутствуют.
Инструменты
Исследование Земли из Космоса