

Известия РАН. Серия биологическая, 2023, № 3, стр. 321-331
Морфо-физиологические детерминанты формирования урожая картофеля в условиях дефицита почвенной влаги
А. Л. Бакунов 1, Н. Н. Дмитриева 1, С. Л. Рубцов 1, А. В. Милехин 1, В. Н. Нестеров 2, Е. С. Богданова 2, О. А. Розенцвет 2, *
1 Самарский ФИНЦ РАН НИИ сельского хозяйства им. Н.М. Тулайкова
446254 Безенчук, ул. Карла Маркса, 41, Россия
2 Самарский ФИНЦ РАН, Институт экологии Волжского бассейна РАН
445003 Тольятти, ул. Комзина, 10, Россия
* E-mail: olgarozen55@mail.ru
Поступила в редакцию 24.01.2022
После доработки 25.04.2022
Принята к публикации 25.04.2022
- EDN: GZWLXU
- DOI: 10.31857/S1026347022700019
Аннотация
Исследован вклад морфо-физиологических параметров листьев и клубней 22 сортов картофеля (Solanum tuberosum L.) в формирование урожайности в условиях недостаточного увлажнения почвы и высоких температур воздуха. Дискриминантным анализом установлено, что размеры устьиц (12%), содержание хлорофилла (10%), количество клубней (29%) и средний вес клубня (21%) определяли градацию растений по урожайности. Наибольший вклад в дискриминацию по отношению к экологической пластичности вносят такие параметры как удельная поверхностная плотность листьев (38%), содержание хлорофилла (13%), каротиноидов (13%) и площадь листа (12%). Ключевыми характеристиками, определяющими высокую урожайность и адаптационную способность картофеля, являются число устьиц в единице площади листа, удельная поверхностная плотность листа и содержание фотосинтетических пигментов.
Картофель (Solanum tuberosum L.) – одна из основных незерновых сельскохозяйственных культур, которая играет важную роль в обеспечении продовольственной безопасности и улучшении питания людей во многих странах мира (George et al., 2018). Согласно данным по глобальному возделыванию картофеля, он выращивается на всех континентах, кроме Антарктиды (Rowe, Powelson, 2002). Показано, что повышение температуры воздуха, прогнозируемое различными сценариями изменения климата, приведет к снижению урожайности картофеля на 18–25%, что может обострить мировые экономические и социальные проблемы (Hijmans, 2003).
Картофель является культурой умеренно-прохладного климата. Эффективным диапазоном температур воздуха для роста его надземной массы является 18–25°С, а оптимальная температура почвы для роста клубней составляет 17–19°С (Давыденко, Лопух, 2019). Картофель является также одним из наиболее чувствительных видов к дефициту влаги в почве (Monneveux et al., 2013; Zarzynska et al., 2017). Засуха значительно снижает основные физиологические и биохимические процессы. Это приводит к снижению продуктивности надземной массы растений (Deblonde, Ledent, 2001), укорачивает цикл роста (Kumar et al., 2007) и снижает количество (Eiasu et al., 2007) и размер клубней (Gervais et al., 2021). В связи с изменением климата картофель все чаще выращивают в районах, подверженных засухе (Evers et al., 2010; Raymundo et al., 2014; Aliche et al., 2018). В этой связи существует необходимость в совершенствовании процесса создания сортов картофеля в направлении ресурсосбережения, биологизации и экологизации (Ashraf, 2010).
Выведение новых сортов требует времени. Одним из подходов наряду с традиционными селекционными программами может стать отбор сортов по ключевым характеристикам, важным для урожайности в неоптимальных условиях выращивания (Parry et al., 2005). Для этого необходимо идентифицировать признаки растений, которые придают толерантность или устойчивость к дефициту воды, высоким температурам воздуха и другим абиотическим факторам, а также установить взаимосвязь между этими характеристиками и урожайностью клубней (Schafleitner et al., 2007; Raymundo et al., 2014).
Урожайность является главным хозяйственно-ценным признаком сельскохозяйственных растений. Показателями, характеризующими адаптивные свойства сортов и селекционного материала картофеля, служат параметры пластичности (bi) и стабильности (Si2) урожайности (Eberhart, Russel, 1966). Формирование урожая осуществляется в ходе продукционного процесса – сложной и интегрированной функции растений, в основе которой лежат генетически детерминированные процессы роста и развития. Морфологические параметры листовой поверхности – одна из важных предпосылок накопления экономически полезной биомассы и формирования урожая картофеля – тесно связаны с физиологическими функциями (Головко, Табаленкова, 2019; Deblonde, Ledent, 2001). Содержание пигментов отражает фотосинтетическую способность листьев, а их пластичность – устойчивость к факторам среды (Lawlor, Tezara, 2009; Kapoor et al., 2020; Plich et al., 2020).
Проводимость устьиц регулирует поглощение СО2 и потерю воды через транспирацию и зависит от плотности и размера устьиц. Полагают, что растения, которые могут поддерживать адекватное содержание воды в течение длительного периода времени под воздействием неблагоприятных условий, будут иметь наибольшую вероятность продолжения метаболического функционирования и выживания (George et al., 2018). Например, сорта картофеля с более высокой устойчивостью к засухе способны поддерживать более высокую клеточную гидратацию (van Loon, 1981; Obidiegwu et al., 2015).
Таким образом, как морфологические так и физиологические параметры листьев важны в формировании урожая картофеля и зависят от генотипа и экологических факторов. В ряде работ сообщается о корреляции морфологических и биохимических признаков с урожайностью картофеля (Mahgoub et al., 2015; Alam et al., 2020). Вместе с тем использование листовых параметров и их взаимосвязи с урожайностью клубней в программах отбора сортов остается в значительной степени нереализованным.
Следует добавить, что оптимальный подбор сортимента для каждого конкретного региона является одним из основных факторов, влияющих на увеличение продуктивности и повышение качества продовольственного и семенного картофеля (Попова и др., 2021). В условиях Средневолжского региона РФ вегетационный период картофеля характеризуется повышенной стрессовой нагрузкой на растения; почвенной и воздушной засухой, повышенным температурным режимом, что приводит к снижению урожаев сельскохозяйственных культур, в том числе картофеля (Рубцов и др., 2019).
Цель настоящего исследования – выявить вклад морфо-физиологических параметров листьев и клубней в формирование урожайности растений картофеля различного генетического происхождения в условиях недостаточного увлажнения почвы.
МАТЕРИАЛЫ И МЕТОДЫ
Объектами исследования служили листья и клубни 22 сортов картофеля (S. tuberosum) различных групп спелости и генетического происхождения. Исследования проводили на опытном участке Самарского НИИСХ им Н.М. Тулайкова – филиала СамНЦ РАН в 2019–2021 гг. Высадку клубней проводили во второй декаде мая. Растения высаживали в четырех повторностях по 50 клубней каждая и выращивали без орошения; почва: чернозём террасовый, обыкновенный, малогумусный, среднемощный, тяжелосуглинистый.
Фенологические наблюдения и учеты продуктивности проводили согласно методическим указаниям (Симаков и др., 2006).
Отбор проб листьев проводили на десяти случайно выбранных растениях одного сорта в период полного цветения (июль). Для биохимических анализов из усредненной массы боковых долей листа составляли навески 0.1–0.5 г в трех повторностях для каждого вида анализа. Пробы листьев отбирали в первой половине дня.
Уборку урожая осуществляли одновременно для всех сортов картофеля в конце августа. Урожайность каждого сорта определяли в т на 1 га, количеству клубней на одно растение (шт.) и среднему весу одного клубня (г). Коэффициенты bi и Si2 определяли по формулам S. Eberhart, W. Russel (1966) в изложении В.З. Пакудина и Л.М. Лопатиной (1984):
bi = ${{{{Y}_{{ij}}}{{I}_{j}}} \mathord{\left/ {\vphantom {{{{Y}_{{ij}}}{{I}_{j}}} {\Sigma I_{j}^{2}}}} \right. \kern-0em} {\Sigma I_{j}^{2}}},$ где Yij – урожай i-того сорта в j‑году, Ij – индекс условий выращивания как отклонение среднего урожая в данном году от среднего значения в опыте; Si2 = [Σ$\delta _{{ij}}^{2}$/(n – 2)], где δij – отклонение фактических урожаев от теоретических, n – число степеней свободы.
Линейные размеры и площадь листа измеряли в 10 боковых долях листа у 5 растений каждого сорта. Удельную поверхностную плотность листьев (УППЛ) рассчитывали как отношение сухой массы листа (мг) к площади поверхности листа (см2).
Для подсчета числа устьиц и определение их длины листья предварительно фиксировались в 3.5%-ном глутаровом альдегиде, приготовленном на фосфатном буфере. Подсчет числа устьиц проводили на нижней стороне листа с помощью светового микроскопа не менее чем в пяти полях зрения для каждой поверхности листа и выражали в шт. на см2 листа. Одновременно с подсчетом устьиц определяли их длину и рассчитывали среднее значение (мкм).
Содержание воды в листьях рассчитывали после высушивания сырой массы (сыр. м.) листьев до постоянного веса при температуре 60°С и выражали в % от сыр. м.
Содержание пигментов определяли спектрофотометрически в ацетоновом экстракте (90%) при λ – 662, 645 и 470 нм. Расчет концентрации хлорофиллов (Хл) а, b и каротиноидов (Кар) проводили в соответствии с рекомендациями Lichtenthaler (1987). Листья (0.5 г сыр. м.) гомогенизировали в фарфоровой ступке вручную с 5 мл ацетона. Все операции проводили при температуре 4°С. Пигменты экстрагировали до полного обесцвечивания навески и фильтровали через стеклянный фильтр. Конечный объем экстракта составлял 50 мл. Количество Хл выражали как сумму Хл а + b.
Результаты представлены в виде средних значений и их стандартных ошибок, в отдельных случаях был рассчитан коэффициент вариации (cv). Взаимосвязь между показателями урожайности и условиями вегетации устанавливали с помощью корреляционного анализа Спирмена.
Для доказательства значимых различий между средними значениями морфо-физиологических показателей при нормальном распределении данных применяли однофакторный дисперсионный анализ (ANOVA) с использованием поправки Бонферрони, а при отклонении от нормального распределения – критерий Краскела–Уоллиса. Для установления различий между вычлененными группами растений картофеля по экологическим или экономическим (урожайность) признакам был использован дискриминантный анализ. Расчеты выполняли, используя программы Statistica 6.0 forWindows, Past 3, Statgraphics Centurion XVI и Microsoft Excel 2007.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
Климатические условия периода вегетации картофеля 2019–2021 гг. отличались неоднородностью. В 2019 г. период всходов, нарастания надземной массы растений и начала цветения (первая декада июня) характеризовался высокими температурами воздуха и недостаточностью увлажнения. Так, средняя температура июня была на 0.9°С выше среднего многолетнего значения, при этом за декаду выпало лишь 6 мм осадков, что было ниже климатической нормы в 9 раз (табл. 1).
Таблица 1.
Агроклиматические условия в период проведения экспериментов
| Месяц | Год | ||
|---|---|---|---|
| 2019 | 2020 | 2021 | |
| Среднемесячная температура, °С | |||
| Июнь | 20.6 (19.7) | 18.2 (19.7) | 22.0 (19.7) |
| Июль | 20.1 (21.4) | 23.8 (21.4) | 23.1 (21.7) |
| Август | 17.9 (19.3) | 18.5 (19.3) | 24.2 (19.7) |
| Среднемесячное количество осадков, мм | |||
| Июнь | 6.0 (54) | 40.0 (54) | 71.0 (46) |
| Июль | 64.0 (53) | 8.0 (53) | 31.0 (47) |
| Август | 42.0 (42) | 29.0 (42) | 6.0 (35) |
| Влажность почвы на глубине 20 см, % | |||
| На глубине 20 см | 18.3 | 13.5 | 13.3 |
Средняя температура воздуха в июле, напротив, была ниже нормы на 1.3°С, а осадков выпало на 11 мм больше. В августе средняя температура составила 17.9°С, что на 1.4°С было ниже нормы, однако осадков выпало в соответствии со средними многолетними значениями. Климатические условия вегетационного периода 2020 г. были крайне неблагоприятными для роста и развития растений картофеля. В периоды завязывания клубней и нарастания их массы – в июне и июле – осадков выпало ниже нормы на 26 и 85%, соответственно. При этом температура воздуха в июле была выше средних многолетних значений на 2.4°С, а в первой и второй декадах температура воздуха составила в среднем 25.1 и 24.9°С соответственно, что выше средних многолетних значений на 4.3 и 4.0°С. Вегетационный период 2021 г. в целом характеризовался более высокими температурами летних месяцев как по отношению к норме, так и по отношению к 2019 и 2020 гг. Кроме того, более низким в сравнении со средними многолетними значениями было количество осадков, за исключением июня. Как следствие влажность почвы в верхнем слое на уровне 20 см в исследуемый период постоянно уменьшалась. Например, в 2021 г. она была в 1.4 раза ниже в сравнении с 2019 г. Таким образом, в течение 3-х лет средняя температура воздуха в три летних месяца была выше нормы в среднем на 0.7°С, а количество осадков было ниже – в среднем на 33 мм.
Средние за три года значения урожайности образцов картофеля и ее элементов с разными сроками созревания – раннеспелые (р), среднеранние (ср), среднеспелые (сс) и позднеспелые (сп) представлены в табл. 2.
Таблица 2.
Характеристика урожайности сортообразцов растений картофеля в период 2019–2021 гг.
| Сорт | Урожайность т/га |
Количество клубней на 1 растение, шт. | Средний вес клубня, г | bi | Si |
|---|---|---|---|---|---|
| Ранние сорта | |||||
| Удача | 19.7 ± 3.7 (33)* | 5.5 ± 0 (6) | 83.6 ± 20.5 (40) | 1.96 | 0.05 |
| Барин | 14.7 ± 4.5 (53) | 4.0 ± 0.6 (27) | 80.5 ± 24.1 (52) | 2.30 | 6.12 |
| Среднеранние сорта | |||||
| Гранд | 18.9 ± 2.8 (25) | 6.9 ± 0.7 (24) | 61.7 ± 11.8 (33) | 0.05 | 45.68 |
| Дебют | 25.0 ± 3.9 (27) | 8.5 ± 0.6 (11) | 68.6 ± 13.4 (34) | 1.69 | 26.88 |
| Калибр | 15.0 ± 2.3 (27) | 5.4 ± 0.3 (9) | 65.7 ± 13.1 (35) | 1.06 | 7.05 |
| Красавчик | 19.6± 3.6 (32) | 5.8 ± 0.2 (6) | 77.0 ± 20.0 (45) | 1.82 | 5.30 |
| Краса Мещеры | 30.1 ± 2.2 (12) | 8.7 ± 0.6 (12) | 86.0 ± 7.5 (15) | 1.09 | 2.29 |
| Нарымская ночка | 15.1 ± 2.2 (26) | 6.9 ± 1.9 (48) | 56.6 ± 13.0 (40) | 0.21 | 31.56 |
| Сердолик | 20.1 ± 3.7 (32) | 5.9 ± 0.5 (13) | 77.5 ± 14.9 (33) | 1.52 | 29.84 |
| Сударыня | 23.0 ± 5.0 (37) | 9.1 ± 1.2 (23) | 57.6 ± 12.4 (37) | 2.00 | 59.06 |
| Третьяковка | 13.5 ± 2.4 (30) | 3.8 ± 0.3 (14) | 78.4 ± 12.8 (28) | 1.22 | 2.22 |
| Эликсред | 18.4 ± 4.0 (37) | 7.4 ± 0.4 (10) | 58.6 ± 17.6 (52) | 2.07 | 0.33 |
| Среднеспелые сорта | |||||
| Жигулевский | 20.4 ± 5.6 (48) | 5.0 ± 0.7 (24) | 89.0 ± 21.7 (42) | 2.94 | 0.26 |
| Августин | 15.6 ± 3.9 (43) | 5.8 ± 2.1 (62) | 65.2 ± 8.1 (22) | 1.71 | 33.14 |
| Брусника | 14.3 ± 2.5 (30) | 6.5 ± 1.2 (33) | 55.1 ± 16.0 (50) | 0.78 | 6.57 |
| Варяг | 21.0 ± 3.2 (26) | 7.4 ± 0.3 (8) | 65.2 ± 14.2 (38) | 1.67 | 0.02 |
| Кумач | 22.2 ± 1.7 (17) | 6.5 ± 0.6 (15) | 76.8 ± 5.3 (12) | 0.97 | 6.72 |
| Северное Сияние | 12.8 ± 2.4 (33) | 4.9 ± 0.5 (19) | 61.2 ± 16.4 (46) | 0.59 | 28.34 |
| Сиверский | 29.0 ± 5.5 (33) | 9.0 ± 0.8 (17) | 71.1 ± 10.7 (26) | 2.07 | 88.61 |
| Сигнал | 21.2 ± 3.0 (24) | 7.1 ± 0.8 (20) | 66.4 ± 5.4 (14) | 1.36 | 12.38 |
| Утро | 22.5 ± 7.7 (59) | 8.0 ± 0.5 (11) | 62.3 ± 21.8 (60) | 3.89 | 22.59 |
| Казачок | 14.6 ± 3.1 (37) | 4.9 ± 0.9 (33) | 70.1 ± 7.9 (19) | 0.30 | 57.17 |
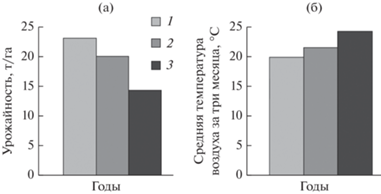
Средняя урожайность составила 19.4 т/га. Максимально стабильной урожайностью в различные по агроклиматическим условиям годы характеризовались с. Краса Мещеры (cv = 12.5) и с. Кумач (cv = 16.7). Наиболее вариативные показатели урожайности среди ранних и среднеранних сортов оказались у с. Барин (cv = 53.4) и с. Сударыня (cv = 37.4). Наибольшая урожайность выявлена у трех сортов – с. Краса Мещеры, с. Дебют и с. Сиверский. Количество клубней на одно растение и вес клубня по результатам трех лет у ранних, среднеранних и у среднеспелых сортов оказались одинаковыми: число клубней – 6.5 шт., вес клубня – 70.9 и 68.2 г, соответственно. Однако в контексте увеличения средней температуры в летние месяцы 2019–2021 гг. значение урожайности всех сортов картофеля снижалось (рис. 1). Урожайность картофеля складывается из количества клубней на одно растение и средней массы клубня. При общем снижении урожайности наблюдалось снижение среднего веса клубня с 95г (2019 г.) до 47 г (2021 г.) при одинаковом количестве клубней на 1 растение. Установлена отрицательная взаимосвязь между урожайностью и температурой воздуха (R = –0.63, при p = 0.06) и положительная – между урожайностью и содержанием влаги в почве (R = = 0.95, при p = 0.01).
Рис. 1.
Средние значения урожайности (а) 22 сортов картофеля и средней температуры воздуха (б) за три летних месяца. 1 – 2019, 2 – 2020, 3 – 2021.
Анализ коэффициента регрессии сорта на индекс среды (bi) показал существенные различия между сортами и группами спелости. Так среди ранних сортов значения bi варьировали в интервале 0.05–2.30, а среди среднеспелых – в интервале 0.3–3.89. Среди наиболее урожайных сортов урожайность снижалась в ряду с. Краса Мещеры → → с. Сиверский → с. Дебют, а показатель экологической пластичности увеличивался в ряду с. Краса Мещеры → с. Дебют → с. Сиверский (1.09 → 1.69 → → 2.07). Как правило, при значении bi >1, сорт относится к интенсивному типу. При значениях bi ≈ ≈ 1 сорт характеризуется высокой, а при bi < 1 – низкой экологической пластичностью.
Стабильность урожайности сорта Si2 в различных условиях определяется квадратическим отклонением фактических показателей урожайности от теоретически ожидаемых: чем меньше Si2, тем стабильнее урожайность. Согласно данным табл. 2 показатели стабильности различались на несколько порядков в зависимости от сорта (0.02–88.61). Половина исследованных сортов относится к стабильному и средне-стабильному типу (Si2 = 0.02–12.38).
Оценка роста по длительности прохождения фенологических фаз показала, что продолжительность фазы от посадки до полных всходов у большинства сортов составляла 23–25 сут. вне зависимости от группы спелости (табл. 3).
Таблица 3.
Длительность прохождения фенологических фаз (сутки) растениями картофеля различных сортов в вегетационные периоды 2019–2021 гг.
| Сорт | От посадки до полных всходов | От полных всходов до начала цветения | Продолжительность цветения |
|---|---|---|---|
| Ранние сорта | |||
| Удача | 22.7± 0.9 | 23.3 ± 1.6 | 19.3 ± 1.3 |
| Барин | 25.7 ± 2.0 | 14.0 ± 1.5 | 31.5 ± 3.3 |
| Среднеранние сорта | |||
| Гранд | 24.0 ± 2.0 | 24.7 ± 4.0 | 20.3 ±3.2 |
| Дебют | 22.5 ± 0.5 | 20.0 ± 1.0 | 23.3 ± 1.2 |
| Калибр | 27.5 ± 3.5 | 18.7 ± 2.0 | 26.7 ± 2.8 |
| Красавчик | 27.0 ± 0.5 | 23.0 ± 8.5 | 23.3 ± 8.4 |
| Краса Мещеры | 23.3 ± 0.3 | 20.7 ± 4.0 | 24.0 ± 4.6 |
| Нарымская ночка | 23.7 ± 2.5 | 22.0 ± 1.0 | 18.7 ± 0.8 |
| Сердолик | 21.0 ± 1.5 | 20.0 ± 0.5 | 31.0 ± 0.7 |
| Сударыня | 21.5 ± 0.5 | 26.0 ± 7.5 | 17.3 ± 4.9 |
| Третьяковка | 26.5 ± 0.5 | 22.3 ± 3.0 | 30.7 ± 4.1 |
| Эликсред | 28.7 ± 7.5 | 23.7 ± 3.0 | 16.5 ± 2.0 |
| Среднеспелые сорта | |||
| Жигулевский | 20.7 ± 2.0 | 20.7 ± 5.0 | 18.7 ± 4.5 |
| Августин | 23.7 ± 2.0 | 22 ± 4.0 | 17.0 ± 3.1 |
| Брусника | 23.5 ± 2.5 | 27.0 ± 1.0 | 22.0 ± 0.8 |
| Варяг | 23.3 ± 2.5 | 21.0 ± 1.0 | 26.0 ± 1.2 |
| Кумач | 21.7 ± 1.9 | 24.3 ± 2.2 | 22.3 ± 2.0 |
| Северное Сияние | 22.7 ± 0.7 | 27.0 ± 4.5 | 12.0 ± 1.9 |
| Сиверский | 21.3 ± 1.5 | 25.7 ± 2.0 | 21.3 ± 1.6 |
| Сигнал | 23.0 ± 2.1 | 25.3 ± 2.1 | 10.0 ± 0.8 |
| Утро | 22.7 ± 0.5 | 19.7 ± 1.0 | 32.3 ± 1.6 |
| Казачок | 24.5 ± 1.5 | 25.3 ± 5.0 | 18.3 ±3.6 |
Средняя продолжительность цветения у сортов с ранними сроками созревания (26.5 сут.) была выше, чем у более поздних сортов (24.9 сут.). В обеих группах спелости обнаружены как сорта с минимальной (с. Эликсред, с. Августин, с. Северное сияние), так и максимальной продолжительностью цветения (с. Барин, с. Третьяковка, с. Утро). Однако взаимосвязи между длительностью прохождения фенологических фаз и урожайностью в климатических условиях исследованного региона в период 2019–2021 гг. не установлено.
Прохождение отдельных фаз и их взаимообусловленность тесно связаны с ростом, и физиологическими процессами на уровне целого растения и отдельных органов. Листья растений с более ранними сроками созревания отличались большей удельной поверхностной плотностью листа (УППЛ) по сравнению с листьями более поздних сортов (табл. 4). Площадь листовой пластины у растений этой группы варьировала от 6 см2 (с. Калибр) до 21.3 см2 (с. Красавчик), в среднем площадь листьев оказалась меньше (11.6 см2) по сравнению с площадью листа у растений с более поздними сроками созревания (13.7 см2).
Таблица 4.
Средние значения физиологических параметров листьев сортов картофеля по результатам 2019–2021 гг.
| Сорт | Площадь листа, см2 | УППЛ, мг/см2 | Длина устьиц, мкм | Число устьиц, тыс. шт/см2 |
Вода, % | Хл а + б, мг/г сыр. м. | Кар, мг/г сыр. м |
|---|---|---|---|---|---|---|---|
| Ранние сорта | |||||||
| Удача | 15.8 ± 1.3 | 4.1 ± 0.4 | 24.0 ± 1.0 | 26.1 ± 1.4 | 79.7 ± 1.0 | 1.67 ± 0.17 | 0.36 ± 0.04 |
| Барин | 13.7 ± 1.2 | 5.9 ± 0.5 | 22.0 ± 2.0 | 29.3 ± 1.2 | 81.1 ± 0.3 | 1.63 ± 0.22 | 0.35 ± 0.06 |
| Среднеранние сорта | |||||||
| Гранд | 7.1 ± 0.6 | 5.4 ± 0.5 | 27.0 ± 2.0 | 28.9 ± 6.5 | 80.5 ± 0.0 | 1.48 ± 0.34 | 0.34 ± 0.08 |
| Дебют | 7.2 ± 0.6 | 13.9 ±1.1 | 25.0 ± 1.0 | 31.2 ± 1.2 | 79.4 ± 2.0 | 1.52 ± 0.01 | 0.31 ± 0.01 |
| Калибр | 6.0 ±0.6 | 5.0 ± 0.4 | 28.0 ± 2.0 | 27.2 ± 2.8 | 82.0 ± 0.7 | 1.50 ± 0.06 | 0.30 ± 0.03 |
| Красавчик | 21.3 ± 1.9 | 4.7 ± 0.4 | 24.0 ± 1.0 | 32.6 ± 2.4 | 80.5 ± 1.3 | 1.34 ± 0.41 | 0.33 ± 0.10 |
| Краса Мещеры | 10.9 ± 1.1 | 14.5 ± 1.0 | 27.0 ± 1.0 | 24.6 ± 2.2 | 81.4 ± 0.5 | 1.45 ± 0.30 | 0.33 ± 0.06 |
| Нарымская ночка | 11.5 ± 1.0 | 4.4 ± 0.4 | 25.0 ± 2.0 | 32.2 ± 4.2 | 82.4 ± 2.4 | 1.48 ± 0.09 | 0.31 ± 0.01 |
| Сердолик | 8.4 ± 0.6 | 6.5 ± 0.6 | 26.0 ± 0.0 | 30.0 ± 0.7 | 80.5 ± 0.6 | 1.47 ± 0.07 | 0.32 ± 0.01 |
| Сударыня | 12.6 ± 1.2 | 3.6 ± 0.4 | 25.0 ± 1.0 | 32.2 ± 1.8 | 81.3 ± 0.3 | 1.35 ± 0.41 | 0.26 ± 0.04 |
| Третьяковка | 13.1 ± 1.2 | 6.9 ± 0.5 | 25.0 ± 0.0 | 28.4 ± 0.4 | 82.1 ± 0.5 | 1.52 ± 0.22 | 0.34 ± 0.02 |
| Эликсред | 11.7 ± 1.0 | 6.6 ± 0.7 | 26.0 ± 1.0 | 24.0 ± 4.4 | 82.8 ± 0.9 | 1.53 ± 0.04 | 0.32 ± 0.01 |
| Среднеспелые сорта | |||||||
| Жигулевский | 16.9 ± 1.3 | 5.2 ± 0.5 | 25.0 ± 1.0 | 31.8 ± 4.3 | 81.3 ± 0.7 | 1.69 ± 0.09 | 0.36 ± 0.03 |
| Августин | 13.1 ± 1.2 | 5.2 ± 0.4 | 27.0 ± 0.0 | 25.1 ± 1.8 | 83.3 ± 0.3 | 1.35 ± 0.18 | 0.30 ± 0.03 |
| Брусника | 12.9 ± 1.3 | 5.2 ± 0.5 | 25.0 ± 1.0 | 32.9 ± 0.8 | 79.4 ± 0.3 | 1.29 ± 0.04 | 0.25 ± 0.01 |
| Варяг | 11.2 ± 1.1 | 5.5 ± 0.5 | 24.0 ± 1.0 | 32.9 ± 0.9 | 80.7 ±0.5 | 1.76 ± 0.44 | 0.36 ± 0.09 |
| Кумач | 13.7 ± 1.4 | 6.9 ± 0.6 | 25.0 ± 3.0 | 26.3 ± 4.6 | 81.5 ± 0.4 | 1.51 ± 0.18 | 0.36 ± 0.04 |
| Северное Сияние | 10.5 ± 0.8 | 3.7 ± 0.3 | 25.0 ± 1.0 | 25.7 ± 2.5 | 83.0 ± 1.9 | 1.33 ± 0.36 | 0.24 ± 0.08 |
| Сиверский | 16.8 ± 1.4 | 3.3 ± 0.3 | 21.0 ± 0.0 | 39.2 ± 0.8 | 82.1 ± 1.1 | 1.82 ± 0.04 | 0.40 ± 0.0 |
| Сигнал | 11.6 ± 1.5 | 3.6 ± 0.2 | 27.0 ± 3.0 | 23.8 ± 1.5 | 80.4 ± 0.0 | 1.38 ± 0.12 | 0.29 ± 0.04 |
| Утро | 11.5 ± 0.9 | 4.9 ± 0.4 | 25.0 ± 2.0 | 28.3 ± 7.6 | 83.4 ± 1.9 | 1.27 ± 0.39 | 0.30 ± 0.06 |
| Казачок | 19.5 ± 1.7 | 3.5 ± 0.3 | 24.0 ± 0.0 | 30.9 ± 3.8 | 80.6 ± 0.8 | 1.64 ± 0.26 | 0.37 ± 0.07 |
Содержание воды в листьях ранних сортов картофеля было на 10% ниже. Количество устьиц по средним показателям мало различалось между группами, но зависело от конкретного сорта. Общее содержание фотосинтетических пигментов также мало различалось между группами, с разным сроком созревания. В листьях ранних и среднеранних сортов количество пигментов было лишь на 10% выше по сравнению со среднеспелыми сортами, но разница между сортами могла достигать до 40%.
При использовании дискриминантного анализа по всему комплексу морфо-физиологических параметров обнаружено четкое ранжирование растений по урожайности и экологической стабильности. Высоко- (Дебют, Краса Мещеры, Сиверский), средне- (Августин, Варяг, Гранд, Жигулевский, Калибр, Красавчик, Кумач, Нарымская ночка, Сердолик, Сигнал, Сударыня, Удача, Утро, Эликсред) и низкоурожайные сорта (Барин, Брусника, Казачок, Северное сияние, Третьяковка) образовали три четких независимых кластера. Аналогичное разделение установлено для высоко- (Августин, Барин, Варяг, Дебют, Жигулевский, Красавчик, Сиверский, Сударыня, Удача, Утро, Эликсред), средне- (Брусника, Калибр, Краса Мещеры, Кумач, Сердолик, Сигнал, Третьяковка) и низкопластичных сортов (Гранд, Казачок, Нарымская Ночка, Северное сияние) (табл. 1, рис. 2а, 2б).
Рис. 2.
Результаты дискриминантного анализа морфо-физиологических параметров в зависимости от урожайности (а) и пластичности (б). 1 – низкоурожайные сорта, 2 – среднеурожайные сорта, 3 – высокоурожайные сорта; 5 – низкопластичные сорта, 6 – среднепластичные сорта, 7 – высокопластичные сорта; 4, 8 – центроиды групп.
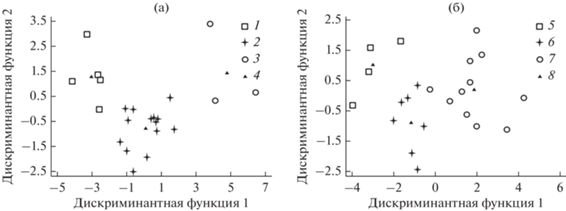
Распределение исследованных сортов, соответствующих 3 группам разной урожайности, в пространстве 1-й и 2-й дискриминантных функций показало, что наибольший вклад в кластеризацию (83%, p = 0.03) вносит первая дискриминантная функция (рис. 2а). Различия между сортами, выделенными по значениям урожайности, обуславливаются в первую очередь количеством клубней (29%) и средним весом клубня (21%), затем следуют размеры устьиц (12%), общее содержание Хл (10%). В отношении экологической пластичности установлено, что первая дискриминантная функция определяет 88% (p = 0.05) всех различий между вычлененными группами сортов (рис. 2б). Наибольший относительный вклад в дискриминацию вносит параметр УППЛ (38%), за которым следуют содержание Хл (13%) и Кар (13%), площадь листа (12%). Подобной закономерности в отношении стабильности урожайности сорта не выявлено.
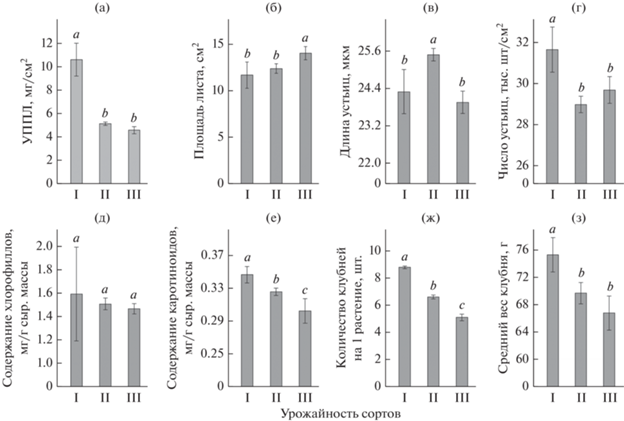
Более подробный анализ отдельных морфо-физиологических параметров листа показал, что высокоурожайные сорта в среднем имеют большее число устьиц в единице площади листа (рис. 3). Площадь листовой пластины у высокоурожайных сортов ниже, чем у низкоурожайных, а показатель УППЛ напротив более чем в два раза выше.
Рис. 3.
Средние значения морфо-физиологических параметров листьев (а–ж) и среднего веса клубней (з) сортов картофеля и их стандартная ошибка в зависимости от урожайности по результатам 2019–2021 гг.: I – высокоурожайные сорта (n = 3), II – среднеурожайные сорта (n = 14), III – низкоурожайные сорта (n = 5). Значимость различий между средними значениями признака устанавливали с использованием с однофакторного дисперсионного анализа (ANOVA). При нормальном распределении данных применяли поправку Бонферрони, а при отклонении от нормального распределения – критерий Краскела–Уоллиса. Разными буквами (a–c) обозначены достоверные различия между средними значениями признака (при p < 0.05).
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Анализ климатических условий (табл. 1) в периоды проведения экспериментов показывает, что район исследования, расположенный в степной зоне Средней полосы России, является удобным экспериментальным полигоном для исследования влияния засушливых условий на сельскохозяйственные растения в полевых условиях (Рубцов и др., 2019; Бакунов и др., 2020). Нами установлено, что увеличение температуры воздуха в летние месяцы 2019–2021 гг. в сравнении с климатической нормой региона привело к увеличению дефицита влаги в почве и, как следствие, к снижению средней урожайности картофеля. Потери урожая, в среднем, составили ~40% в основном за счет снижения массы клубней. При изучении 56 генотипов картофеля, выращиваемых в условиях полива с последующим водным стрессом, было обнаружено снижение, как числа, так и веса клубней (Gervais et al., 2021). Стресс засухи также значительно снизил вес клубней и их количество у сортов S. tuberosum, S. tuberosum × S. tuberosum subsp, однако снижение урожайности клубней варьировало в зависимости от генотипа (Schafleitner et al., 2007).
Чувствительность картофеля к засухе в основном объясняется малой эффективностью растений усваивать воду из-за неглубокого залегания корневой системы в почве и низкой способностью фотосинтетических механизмов восстанавливаться после водного и теплового стресса (Monneveux et al., 2013; Zarzynska et al., 2017). Содержание воды в листьях растений обуславливает осмос и тургор, от которых зависит рост, поддержание структуры, движение устьиц, фотосинтез и пр. Растения картофеля используют несколько стратегий для выживания в условиях дефицита воды и образования клубней (Plich et al., 2020), например, уменьшение потери воды или поддержание водопоглощения (Obidiegwu et al., 2015). Судя по содержанию воды в листьях, все исследованные нами растения были способны поддерживать достаточно высокий уровень в тканях листьев, несмотря на экстремально низкую влагообеспеченность почвы. Это свидетельствует о том, что урожайность клубней для разных генотипов картофеля при недостатке влаги в почве были связаны не с различиями в содержании воды в листьях, а скорее, с морфологией, физиологическими и биохимическими процессами.
Фотосинтетические пигменты имеют первостепенное значение для фотосинтеза и зависят от водного дефицита (Kapoor et al., 2020). Количество Хл, как правило, связано с продуктивностью сорта (Obidiegwu et al., 2015; Бакунов и др., 2020). Большее содержание Хл и Кар у высокоурожайных сортов способствует большему поглощению света и защите фотосинтетического аппарата от окислительных процессов, возникающих при дефиците влаги (Lawlor, Tezara, 2009; Dahal et al., 2019). При этом параметр содержание Хл оказался связан как с экологической пластичностью, так и с урожайностью картофеля, что показывает его интеграционную значимость, как в формировании урожая, так и в адаптации растений к экологическим условиям.
Высокоурожайные сорта отличались также низкой площадью листа по сравнению с низкоурожайными. В их листьях содержится больше сухого вещества в единице площади листа (показатель УППЛ), большое число некрупных устьиц (рис. 3). Эти особенности в структуре листьев способствует ограничению потери воды и большей ассимиляции СО2, что положительно влияет на продуктивность растений. Полученные нами данные согласуются с результатами полевых экспериментов с использованием культуры ячменя: контроль над потерей воды осуществлялся за счет снижения индекса листовых параметров (Lawlor, Tezara, 2009).
Показатель УППЛ важен для таких процессов как отношение процессов фотосинтез/азотфиксация, скорость фотосинтетической ассимиляции, продолжительность жизни листа, а также для разделения растений на функциональные группы в зависимости от условий окружающей среды (Cheng et al., 2014). Это отразилось на разделении групп сортов по коэффициенту пластичности bi.
Из всех исследованных сортообразцов картофеля с. Сиверский оказался и высокоурожайным (29 т/га) и высокопластичным (bi = 2.07). Однако коэффициент стабильности его урожайности достаточно высокий (Si2 = 88.61). Это означает, что сорт хорошо отзывается на улучшение условий выращивания, но в неблагоприятные годы урожайность может снизиться. Характерными признаками этого сорта являются наибольшее число устьиц в единице площади листа, высокая гидратационная способность листьев и высокое содержание фотосинтетических пигментов. Из всех высокоурожайных сортов он имел наименьшее значение УППЛ. Как показали наши предыдущие исследования, с. Сиверский отличался большей изменчивостью белкового и липидного метаболизма, а также более интенсивными процессами окисления и антиоксидантной защиты по сравнению с малоурожайным с. Третьяковка (Розенцвет и др., 2021). В свою очередь, с. Краса Мещеры также является высокоурожайным (30.1 т/га), имеет коэффициент bi близкий к 1, а Si2 равный 2.29, то есть характеризуется, как генотип с высокой экологической пластичностью и высокой стабильностью генотипа. Учитывая, что в основе реакции растений на стресс лежат сотни генов, можно полагать, что оба сорта могут служить источником генетического материала для молекулярно-физиологических и селекционных исследований с целью отбора сортов для интенсивного или стабильного урожая.
Таким образом, морфо-физиологические показатели листьев и клубней определяют урожайность и экологическую пластичность сортов картофеля. Характеристиками, способствующими высокой урожайности и адаптационной способности, являются число устьиц в единице площади листа, УППЛ и содержание фотосинтетических пигментов. В целом полученные результаты детализируют концепцию “адаптация/продуктивность”, согласно которой либо достигается наибольшая защита (экологическая пластичность), либо максимум продукции (урожайность), что необходимо учитывать при подборе сортов к конкретным почвенно-климатическим условиям.
Финансирование. Данная работа выполнена в рамках Комплексного плана научных исследований ”Развитие селекции и семеноводства картофеля” и “Структура, динамика и устойчивое развитие экосистем Волжского бассейна” (№ 021060107217-0-1.6.19).
Благодарности. Авторы выражают благодарность ФГБНУ “Федеральный исследовательский центр картофеля им. А.Г. Лорха” и создателям сортов S. tuberosum за предоставленный для исследований семенной материал.
Список литературы
Бакунов А.Л., Милехин А.В., Рубцов С.Л., Шевченко С.Л. Содержание фотосинтетических пигментов как косвенный признак устойчивости сортов картофеля к высоким температурам воздуха и недостаточному увлажнению // Изв. Сам. гос. сельскохоз. акад. 2020. № 2. С. 8–13.
Головко Т.К., Табаленкова Г.Н. Донорно-акцепротные связи в растении картофеля // Физиология растений. 2019. Т. 66, 34. С. 313–320.
Давыденко О.В., Лопух П.С. Влияние термических условий на урожайность картофеля в административных районах республики Беларусь // Журн. Белорус. гос. универ. Геог. Геол. 2019. № 1. С. 46–62.
Пакудин В.З., Лопатина Л.М. Оценка экологической пластичности и стабильности сортов сельскохозяйственных культур // Сельскохозяйственная биол. 1984. № 4. С. 109–113.
Попова Л.А., Головина Л.Н., Гинтов В.В., Шаманин А.А. Оценка адаптивности сортообразцов картофеля в условиях северных территорий Архангельской области // Карт. и овощи. 2021. № 1. С. 34–37.
Розенцвет О.А., Богданова Е.С., Нестеров В.Н., Шевченко С.Н., Бакунов А.Л., Милехин А.В., Рубцов С.Л. Продуктивность и динамика морфологических и физиолого-биохимических параметров картофеля в условиях засушливого климата // Док. Рос. акад. наук. Науки о жизни. 2021. Т. 497. С. 143–147.
Рубцов С.Л., Бакунов А.Л., Милехин А.В., Дмитриева Н.Н. Критерии отбора новых сортов картофеля для Средневолжского региона // Изв. Оренбург. гос. аграр. универ. 2019. № 1(75). С. 52–55.
Симаков Е.А., Склярова Н.П., Яшина И.М. Методические указания по технологии селекционного процесса картофеля. М.: Достижения науки и техники АПК, 2006.
Aliche E.B., Oortwijn M., Theeuwen T.P.J.M., Bachem C.W.B., Visser R.G.F., van der Linden C.G. Drought response in field grown potatoes and the interactions between canopy growth and yield // Agricult. Water Manag. 2018. V. 206. P. 20–30.
Alam M.S., Islam M.F., Rahman M.S., Molla M.M., Uddin M.Z., Mian M.A. Varietal evaluation and selection of yield-associated traits of potato (Solanum tuberosum) using correlation and path analysis in haor area of Moulvibazar Bangladesh // J. Agric. Sci. Engin. Innov. 2020. V. 1(10). P. 2–9.
Ashraf M. Inducing drought tolerance in plants: recent advances // Biotechnol. Adv. 2010. V. 28(1). P. 169–183.
Cheng T., Rivard B., Sanchez-Azofeifa A.G., Feret J.-B., Jacquemoud S., Ustin S.L. Deriving leaf mass per area (LMA) from foliar reflectance across a variety of plant species using continuous wavelet analysis // ISPRS J. Photogram. Rem. Sens. 2014. V. 87. P. 28–38.
Dahal K., Li X.Q., Tai H., Creelman A., Bizimungu B. Improving potato stress tolerance and tuber yield underclimate change scenario – a current overview // Front. Plant Sci. 2019. V. 10. P. 563.
Deblonde P.M.K., Ledent J.F. Effects of moderate drought conditions on green leaf number, stem height, leaf length and tuber yield of potato cultivars // Europ. J. Agron. 2001. V. 14(1). P. 31–41.
Eiasu B.K., Soundy P., Hammes P.S. Response of potato (Solanum tuberosum) tuber yield components to gelpolymer soil amendments and irrigation regimes // New Zealand J. Crop and Horticul. Sci. 2007. V. 35(1). P. 25–31.
Eberhart S., Russel W. Stability parameters for comparing varieties // Crop. Sci. 1966. V. 6. № 1. P. 36–42.
Evers D., Lefèvre I., Legay S., Lamoureux D., Hausman J.F., Rosales R.O., Marca L.R.T., Hoffmann L., Bonierbale M., Schafleitner R. Identification of drought-responsive compounds in potato through a combined transcriptomic and targeted metabolite approach // J. Exp. Bot. 2010. V. 61(9). P. 2327–2343.
George T.S., Taylor M.A., Dodd I.C., White P.J. Climate change and consequences for potato production: a review of tolerance to emerging abiotic stress // Pot. Res. 2018. V. 60. P. 239–268.
Gervais T., Creelman A., Li X-Q, Bizimungu B., De Koeye D., Dahal K. Potato response to drought stress: physiological and growth basis // Front. Plant Sci. 2021. V. 12. 698060.
Hijmans R.J. The effect of climate change on global potato production // Amer. J. Pot. Res. 2003. V. 80. P. 271–279.
Kapoor D., Bhardwaj S., Landi M., Sharma A., Ramakrishnan M., Sharma A. The impact of drought in plant metabolism: how to exploit tolerance mechanisms to increase crop production // App. Sci. 2020. V. 10(16). P. 5692.
Kumar S., Asrey A., Mandal G. Effect of differential irrigation regimes on potato (Solanum tuberosum) yield and post-harvest attributes // Indian J. Agricult. Sci. 2007. V. 77(6). P. 366–368.
Lawlor D.W., Tezara W. Causes of decreased photosynthetic rate and metabolic capacity in water-deficient leaf cells: a critical evaluation of mechanisms and integration of processes // Ann. Bot. 2009. V. 103(4). P. 561–579.
Lichtenthaler H.K. Chlorophylls and carotenoids pigments of photosynthetic biomembranes / Eds. Douse R., Packer L. // Methods in enzymology. 1987. N.Y.: Academic Press Inc. P. 350–382.
Mahgoub H.A.M., Eisa G.S.A., Youssef M.A.H. Molecular, biochemical and anatomical analysis of some potato (Solanum tuberosum L.) cultivars growing in Egypt // J. Gen. Engin. Biotech. 2015. V. 13(1). P. 39–49.
Monneveux P., Ramírez D.A., Pino M.-T. Drought tolerance in potato (S. tuberosum L.). Can we learn from drought tolerance research in cereals? // Plant Sci. 2013. V. 205. P. 76–86.
Obidiegwu J.E., Bryan G.J., Jones H.G., Prashar A. Coping with drought: stress and adaptive responses in potato and perspectives for improvement // Front. Plant Sci. 2015. V. 6. P. 1–23.
Parry M.L., Rosenzweig C., Livermore M. Climate change, global food supply and risk of hunger // Philosoph. Trans. Royal Soc. B: Biol. Sci. 2005. V. 360(1463). P. 2125–2136.
Plich J., Boguszewska-Mańkowska D., Marczewski W. Relations between photosynthetic parameters and drought-induced tuber yield decrease in Katahdin-derived potato cultivars // Pot. Res. 2020. V. 63(4). P. 463–477.
Raymundo R., Asseng S., Cammarano D., Quiroz R. Potato, sweet potato, and yam models for climate change: A review // Field Crops Res. 2014. V. 166. P. 173–185.
Rowe R.C., Powelson M.L. Potato early dying: management challenges in a changing production environment // Plant Dis. 2002. V. 86. P. 1184–1193.
Schafleitner R., Gutierrez R., Espino R., Gaudin A., Pérez J., Martínez M., Bonierbale M. Field screening for variation of drought tolerance in Solanum tuberosum L. by agronomical, physiological and genetic analysis // Pot. Res. 2007. V. 50. P. 71–85.
van Loon C.D. The effect of water stress on potato growth, development, and yield // Am. Pot. J. 1981. V. 58(1). P. 51–69.
Zarzynska K., Boguszewska-Mankowska D., Nosalewicz A. Differences in size and architecture of the potato cultivars root system and their tolerance to drought stress // Plant Soil Environ. 2017. V. 63. P. 159–164.
Дополнительные материалы отсутствуют.
Инструменты
Известия РАН. Серия биологическая