

Известия РАН. Серия биологическая, 2023, № 2, стр. 172-185
К изучению водяной полевки (Arvicola amphibious) на северо-западном пределе ареала
1 Петрозаводский государственный университет
185910 Петрозаводск, просп. Ленина, 33, Россия
2 Карельский научный центр РАН
185035 Петрозаводск, Пушкинская ул., 11, Россия
* E-mail: Ivanter@petrsu.ru
Поступила в редакцию 18.02.2022
После доработки 06.08.2022
Принята к публикации 08.08.2022
- EDN: HAFIRN
- DOI: 10.31857/S1026347022700044
Аннотация
На основе многолетних экспедиционных и стационарных исследований на таежном Северо-Западе России (1958–2018 гг.) охарактеризованы численность, биотопическое размещение, питание, размножение, пространственная и экологическая структура популяции и динамика численности водяной полевки, в том числе связанные с ее обитанием у северо-западных границ ареала. В европейской части ареала, особенно вблизи северо-западных границ, водяная полевка становится все более узким стенотопом с отчетливым проявлением синантропизма, и, в частности, усиленным тяготением к огородам и приусадебным участкам. В репродукции участвуют взрослые и молодые полевки ранних весенне-летних выводков, принося за сезон до двух выводков, в среднем, из 6.5 ± 0.37 детенышей (зимовавшие) и 5.7 ± 0.41 (сеголетки). Численность колеблется по годам в значительных пределах под влиянием резких изменений гидрологического режима водоемов
Представленная статья посвящена обобщенному анализу собственных (1958–2018 гг.) и литературных данных о распространении, численности и биологии водяной полевки (Arvicola amphibious) на северо-западной периферии ареала. Распространена она в Евразии чрезвычайно широко – от лесотундры до пустынных степей включительно. Это типичный обитатель побережий водоемов: в поймах больших и малых рек, ручьев и озер различного типа, где она образует в основном линейные поселения, а также на обширных заболоченных пространствах водоразделов и междуречий северной части Евразии, где типичны диффузные поселения (Карасева и др., 2008). На территории бывшего СССР – от балтийского до черноморского побережья, Закавказья, северного Прикаспия и южного Прибалхашья до Алае-Саянской горной страны, Забайкалья и Восточного Приалданья (Громов, Ербаева, 1995). Водяная полевка повсюду предпочитает берега рек, озера различных типов, верховые и пойменные болота; селится на сплавинах, лугах, среди кустарниковых зарослей и в болотистом мелколесье. При этом осенью водяная полевка переселяется с берегов рек и озер на луга или огороды и в течение всего холодного времени года ведет подземный образ жизни, роя глубокие и сложные норы, нанося при этом существенный ущерб в основном огородным культурам. Полевка изучена неравномерно. В Сибири, особенно в Западной ее исследовали достаточно полно (Зверев, Пономарев, 1930; Шварц и др. 1957; Максимов, 1959, 1966; Максимов, Андрусевич, 1959; Пантелеев, 1968; Ердаков, 1972, 1984, Соломонов, 1980; Евсиков и др., 1991, 1997; Рогов, 1999; Назарова, Проскурняк, 2017, и мн. др.), тогда как в европейской части ареала фрагментарно и эпизодически (Жарков, Теплов, 1931; Тупикова, Швецов, 1956; Доброхотов, Неронов, 1965; Соловьев, 1994). Представленная статья призвана хотя бы частично восполнить этот пробел.
МАТЕРИАЛ И МЕТОДИКА
Материал собирали в процессе специальных учетов численности зверьков на ловушко-линиях, составленных из капканчиков (отработано 6900 капкано-суток) и четырех 30-метровых ловчих канавок с врытыми вровень с их дном тремя металлическими конусовидными цилиндрами (980 канавко-суток). Всего на линиях ловушек было добыто 124 водяные полевки, а ловчими канавками – 334. Кроме того, в соответствии с рекомендациями Пантелеева и др., (2001), применяли визуальный учет зверьков по отверстиям нор, погрызам и кормовым столикам. Этот учет проводили на береговой полосе шириной 1 м, а единицей учета служило число учетных объектов в расчете на 1 км берега.
Возраст пойманных зверьков определяли по морфологическим особенностей черепа, а генеративное состояние животных оценивали при вскрытии в соответствии с указаниями Карасевой и др. (2008). Лактацию фиксировали по состоянию молочных желез и сосков. Питание анализировали по учетам поедей на кормовых столиках и содержимому желудков, а также экспериментально – на основе опытов, проведенных на животных, содержавшихся в неволе.
Статистический анализ материала (расчет средних, статистических ошибок, коэффициента вариации, хи-квадрата, достоверности различий, показателя асимметрии, величины и значимости корреляций и др.) производили по общепринятым методикам (Ивантер, 1979; Ивантер, Коросов, 1992, 2003).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Распространение и численность. В европейской части ареала, особенно близ северо-западных границ, водяная полевка становится все более узким стенотопом с отчетливым проявлением синантропизма (Громов, Ербаева, 1995).
Согласно данным Пантелеева (1968), территорию рассматриваемого региона населяет карело-финская географическая популяция этого вида, для которой характерны единство условий существования и сравнительно низкий общий уровень численности, несмотря на обилие здесь озер и болот. Это связано с ограниченной емкостью угодий из-за их относительной малокормности и неблагоприятного гидрологического режима. Все же и в Карелии водяная полевка достигает местами довольно высокой численности. Так, в заповеднике “Кивач” на реках Ниве и Сандалке и по Чечкиному ручью, впадающему в Мунозеро, в процессе учетов 1958–1960 гг., мы насчитывали до 20 жилых нор и до 30 кормовых столиков этого зверька на 1 км береговой линии, а в период с 2009 по 2011 гг. там же эти показатели составили соответственно 27 нор и 42 кормовых столика. Высокая численность водяной полевки (5–15 жилых нор и 10–20 кормовых столиков на 1 км берега) отмечена также на ряде водоемов Пудожского района (рр. Водла, Гакукса, Летняя и Сума, озера Сюзикозеро, Ведлозеро, Колодозеро и Муромозеро, Ершовский ручей), Медвежьегорского (р. Кумса и Йемена, оз. Семчезеро), Олонецкого (р. Олонка и Тумакса, оз. Сюськоярви), Муезерского (оз. Унусозеро, р. Тумба и Сидра), Прионежского (озера Паросозеро и Машезеро, р. Шопша и Лососинка), Сегежского (Лисья губа Сегозера) и Калевальского (озера Нюк и Кимасозеро, рр. Ногеукс и Черная) районов. С меньшей плотностью (обычно до 3 жилых нор и до 5 кормовых столиков на 1 км берега) населяет этот вид водоемы Лоухского, Беломорского, Кемского и Суоярвского районов.
Вполне показательны и проведенные нами в 1959–1960 и 2013 гг. специальные количественные учеты водяных полевок ловушко-линиями и ловчими канавками на побережье р. Суны (заповедник “Кивач”). Согласно полученными данным (табл. 1), средние за эти годы показатели численности зверьков составили там соответственно 1.80 экз. на 100 лов.-сут и 3.41 на 10 кан.-суток.
Таблица 1.
Учеты численности водяной полевки в заповеднике “Кивач” (побережье р. Суны)
| Годы учетов | Учеты ловушко-линиями | Учеты ловчими канавками | ||||
|---|---|---|---|---|---|---|
| число лов-сут. | число добытых зверьков | на 100 лов-сут | число кан-сут. | число добытых зверьков | на 10 кан-сут | |
| 1959 | 2120 | 43 | 2.00 | 310 | 102 | 3.28 |
| 1960 | 1800 | 36 | 2.00 | 330 | 78 | 2.36 |
| 2013 | 2980 | 45 | 1.51 | 340 | 134 | 4.52 |
| Итого | 6900 | 124 | 1.80 | 980 | 334 | 3.41 |
Биотопическое размещение, особенности норения и участки обитания. Обитающие на таежном Северо-Западе России водяные полевки относятся в основном к разным типам т. н.пойменных популяций, различающихся между собой, прежде всего, степенью связи вида с водной средой (Пантелеев и др., 2001). В течение всего лета они достаточно тесно связаны с различными по характеру и размерам водоемами и заселяют речные поймы, верховые и пойменные болота, прибрежные участки небольших речек и ручьев, берега мелководных губ различного типа озер, заросшие водно-болотной растительностью, мелиоративные канавы по окраинам болот, заболоченные осоковые кочкарники, сырые пойменные луга и т.п. Полевки селятся на сплавинах, среди кустарниковых зарослей, в болотистом мелколесье по берегам лесных ручьев и речек, на полях и огородах, расположенных вблизи водоемов. Все это – постоянные весенне-летние местообитания зверьков, где они живут вплоть до периода расселения молодняка. Ближе к осени, обычно начиная с конца июля, водяные полевки в массе покидают летние убежища и перебираются на более сухие участки – прилегающие к водоемам и болотам луга, огороды и залежи. Здесь они перезимовывают, а весной (обычно в мае) вновь возвращаются к водоемам. На местах, где летние и зимние стации находятся по соседству друг с другом (например, сенокосные луга по берегам р. Суны и Сандалки в заповеднике “Кивач”), сезонные кочевки растягиваются на продолжительное время и не принимают массового характера. Напротив, в районах, где эти стации разобщены, наблюдается хорошо выраженное массовое передвижение зверьков (озера Нюк и Унусозеро, рр. Тумба, Кумса, Нива, Черная и др.). На берегах водоемов, не замерзающих зимой (например, на ручье, впадающем в Мунозеро), водяные полевки живут иногда круглый год и сезонных перемещений у них не наблюдается. Полевки реже живут небольшими разрозненными группами, но чаще образуют либо ленточные, либо (реже) сплошные мозаично расположенные поселения.
Почти половину года полевки проводят в русловой части водоема и только в конце–начале октября выселяются на возвышенные участки пойменного берега, не удаляясь от русла более 20–30 м, выкапывая здесь сложную систему ходов, в основном, подземных, запасая вегетативные части пойменных растений. Низинные луга надпойменных террас обитаемы в течение всего года. Только весной во время затопления их талыми водами зверьки смещаются ближе к лесу, к менее подтопленным кустарникам.
В культурном ландшафте, вблизи деревень и дачных участков с огородами, поселения полевок отличаются даже большей устойчивостью, чем в естественных местообитаниях. Зверьки здесь смещаются к кустарникам, где под их корнями устраивают зимние норы, запасая в них картофель, морковь или корневища сорных растений, спрессовывая их в боковых отнорках в виде “колбасок” по их диаметру. По сообщению Соловьева (1994), ему не раз приходилось находить в таких местах до двух ведер запасенных грызунами на зиму клубней картофеля.
Во время кочевок водяные полевки встречаются во всех биотопах, в том числе и в лесных участках, расположенных на значительном удалении от воды. Однако лесные стации представляют для них лишь миграционные пути, постоянно они здесь не живут.
Круглый год полевки обычно обитают в норах, вырытых по берегам водоемов, близ самой воды, но в теплое время года, особенно при резких подъемах воды, в наземных и надземных гнездах, размещенных в кочках, образованных корневыми частями водно-болотных растений: осок, рогозов и тростника. Постоянные норы обычно неглубокие, кормовые же ходы прокладываются на большей глубине – до 1 м, а иногда и глубже. Роющая деятельность наиболее активна в осенне-зимний период, когда большую часть времени зверьки проводят в неглубоких подземных убежищах и в различных естественных укрытиях, а также во время массового расселения молодняка. Обследованные нами береговые норы водяных полевок располагались, как правило, на глубине 10–20 см от поверхности земли, но в отдельных случаях (например, на некоторых участках в пойме р. Сандалки) значительно глубже (до 50–60 см). Обычно они представляли собой несложную систему подземных галерей с главным ходом, открывающимся чаще всего выше уровня воды, реже – под водой. Помимо главного, магистрального, хода в норе почти всегда имеется 2–4 (редко больше) дополнительных хода, часто с отдельным выходом наружу. Все они обязательно соединены между собой и с главным ходом. Вся система подземных галерей лежит обычно в одной плоскости, в большей или меньшей степени наклоненной к воде. Однако там, где водоемы характеризуются резкими колебаниями уровня воды, полевки строят многоэтажные жилища с 2–4 гнездовыми камерами, сооруженными на разной высоте. Иногда встречаются и колониальные поселения (например, в устье р. Сандалки, на р. Ниве, на озерах Сюськоярви и Семчезеро), где по сути дела нет отдельных нор: весь берег изрыт магистральными и соединительными ходами, которыми зверьки пользуются сообща. Число выходных отверстий зависит от характера берега и типа поселения.
Каждая нора имеет 1–3 гнездовые камеры, расположенные на глубине 5–20 см. В них находятся гнезда, сделанные из травы. Размеры гнездовых камер (их измерено 15): высота 12–18, длина 15–34, ширина 14–20 см. Число выходных отверстий зависит от характера берега и типа поселения. На Чечкином болоте (заповедник “Кивач”) мы нашли наземное гнездо водяной полевки, устроенное в зарослях голубики, багульника и других трав на вершине кочки.
Встречается в Карелии и другой тип норения этих зверьков – длинные подземные ходы, отрытые горизонтально на сенокосных лугах и огородах с частыми выходами на поверхность. Это т. н. зимовочные норы – основной осенне-зимний тип убежища, но иногда водяные полевки живут в них и летом. Для нор водяной полевки характерны похожие на кротовые поверхностные земляные выбросы, а после зимы – образующиеся под снегом своеобразные земляные “колбаски”.
Множество исследователей измеряли и описывали индивидуальные участки водяной полевки в разных точках ее ареала (Громов, 2008). Однако, знакомясь с этими материалами, нельзя не заметить, что эта характеристика чрезвычайно изменчива и связана с множеством факторов: пол, возраст, размеры животного, участие в размножении, географическая точка, время года, характер местообитания и т.д. Тем не менее наличие индивидуального участка представляется для водяной полевки весьма характерной и достаточно важной видовой особенностью.
Как показывают и наши наблюдения в природе, и эксперименты на площадках живоотлова, большинство взрослых особей этого грызуна занимают до конца сезона размножения относительно постоянные индивидуальные участки обитания. С помощью мечения и повторных отловов установлено, что участки обитания самок, длина которых в ленточных поселениях достигает 80 м, почти не перекрываются. Напротив, более крупные индивидуальные участки самцов, длина которых варьирует от 20 до 310 м, перекрываются в значительной степени. Индивидуальный участок каждого взрослого самца накладывается на участки нескольких самок (максимально – до восьми). Вследствие зарастания водоема, а также по мере его высыхания в летний период, конфигурация участков обитания может меняться. При этом и обособленность индивидуальных участков самок и агрегированность участков самцов, каждый из которых обычно перекрывает участки двух–трех других особей того же пола, не вызывает сомнений.
Исследования, проведенные с помощью радиотелеметрии, показали, что в ленточных поселениях оседлые зверьки перемещаются вдоль узкой прибрежной полосы шириной не более 1–2 м, при этом площадь индивидуального участка, оцененная на основе радиотелеметрических данных, оказалась на 13% больше, чем при оценке методом повторных отловов (Stoddart, 1971; Рогов, 1999).
В некоторых биотопах, например, на болотах, ленточные поселения преобразуются в диффузные, особенно в годы высокой численности. Характерной особенностью диффузных поселений является совместное использование пространства несколькими взрослыми особями (Пантелеев и др., 2001).
Расположение участков обитания взрослых особей регулярно меняется, поскольку они время от времени перемещаются вдоль береговой линии водоемов на значительное расстояние. Частота перемещений составляет 8% от общей численности популяции в сутки (Рогов, 1999) Таким образом, периоды оседлости взрослых особей относительно кратковременны. Регулярная же смена участков обитания объясняется, по-видимому, относительно быстро меняющимися условиями существования этого вида вблизи водоемов.
В апреле–мае внутри ленточных поселений обособляются более мелкие репродуктивные группировки, объединяющие от двух до десяти зверьков. В этот период происходит относительное укрепление социальных связей между самцами и самками, вследствие чего образуются “квази-моногамные” пары (Плюснин, Евсиков, 1983; Громов, 2008). Позднее у беременных и кормящих самок проявляется более отчетливое территориальное поведение, они становятся агрессивными по отношению к самцам, из-за чего часть последних откочевывает. В результате число взрослых особей в репродуктивных группировках уменьшается, а границы между группировками становятся менее четкими. В дальнейшем подвижность самцов увеличивается в еще большей степени.
Суточная активность и перемещения. Суточная активность водяной полевки, как и у большинства мелких грызунов (Громов, 2008), круглосуточная и полифазная. Тем не менее как показали наблюдения за мечеными животными у их нор и на индивидуальных участках, ее отличает наличие характерных подъемов и спадов. Абсолютный минимум активности приходится обычно на сумерки и ночь, а максимально длительный и высокий подъем – на утренние часы. При этом наибольшую часть активной деятельности эти полевки тратят на кормежку около нор. По оценке Громова (2008), средняя продолжительность вне норовой активности составляла у этих зверьков 30 мин. В то же время конкретный ход суточной активности, ее периодичность и продолжительность фаз, длительность пауз и т.д., – все это зависит от возраста зверьков, сезона года и даже от географических особенностей популяции. И хотя изучение изменений активности зверьков в зависимости от возраста представляет определенную методическую сложность, тем не менее многое в этом отношении удалось выяснить. В частности, при всем сходстве соответствующих хронограмм, у молодых самцов было выявлено хорошо заметное повышение активности в утренние часы, у взрослых же в это время наблюдается отчетливый ее спад (Ердаков, 1984). Кроме того, у молодых полевок подъемы активности в течение суток происходят чаще, т.е. полифазность их ритма больше, чем у взрослых.
Более точные результаты дало нам количественное сравнение активности взрослых и молодых зверьков по распределению в течение суток: если средняя часовая активность у разных возрастных групп самцов на этих материалах статистически неразличима, то интенсивность активности в светлое время суток у молодых самцов оказалась достоверно больше, чем у взрослых (t = = 3.3, P = 0.99). Основное различие, видимо, состоит в том, что молодые самцы активнее взрослых днем, но пассивнее ночью.
Гораздо отчетливее и более понятны в плане вызывающих их физиологических причин сезонные изменения активности. Так, у водяных полевок показана строгая монофазная цикличность с периодом около года в активности половых желез (Ивантер, 2018): максимальная их активность у половозрелых животных отмечена с марта по октябрь, а минимальная – в декабре–январе. Кроме того, рассматривая сезонные изменения в организме водяных полевок, Г.А. Кондрашкин и Л.И. Кузнецова (1959) нашли “хорошо проявляющуюся годовую цикличность уровня потребления кислорода при "критической точке”, а также общего числа лейкоцитов в крови животных" (цит. по Зудовой, 2014). Таким образом, имеется и вполне очевидная физиологическая основа для сезонных изменений в деятельности водяных полевок.
Сезонная изменчивость суточной активности у них проявляется в сокращении ее бюджета и уменьшении интенсивности активного состояния в светлое время суток к осени. Осенью же амплитуда колебаний суточной активности у этих зверьков чаще всего снижается, а сама активность становится более ночной.
Интересное экспериментальное изучение суточной активности водяных полевок в республике Коми, содержащихся в открытых вольерах вблизи водоема, а также в клетках, с помощью актографов провел профессор Сыктывкарского университета В.А. Соловьев (1994). При круглосуточных экспериментах в вольерах им проводился подробный хронометраж кормовой и двигательной активности животных. Исследования начались в конце июня и закончились в первой декаде июля. Анализируя полученные данные, автор прежде всего констатирует значительную лабильность активности водяных полевок. Не было замечено изменений активности в зависимости от температурного фактора, и только дождь синхронизировал активность всех подопытных животных – все они уходили в укрытия. Наблюдения показали, что водяной полевке свойствен полифазный тип активности. Периоды кормления, продолжающиеся от 1–3 до 5 мин, чередовались с периодами двигательной активности, длительность которых нередко достигала 15 мин. Средняя продолжительность кормовой и двигательной активности в течение суток составила 418 мин, или 29%. В среднем зверьки треть суток активны, а две трети отдыхают.
Согласно цитируемому автору, соотношение форм активности неодинаково у разных возрастных групп. У молодых зверьков двигательная активность относительно выше, чем кормовая: на прием корма зверек затрачивает в сутки 68, а на движение – 262 мин, соотношение – 20.6 : 79.4%. Перезимовавшие полевки кормятся дольше, чем движутся. Из 580 мин. на принятие корма тратится 303, а на движение –277 мин., соотношение – 52.2 : 47.8%. В целом, по соотношению активности и отдыха ювенильные особи менее подвижны. Их общая активность составила 22.9% от времени суток, тогда как у перезимовавших – 40.2%. Беременные самки оказались менее активными, чем самцы, а прием пищи у них составлял 61% от общей активности животного. Если сравнить этот показатель с аналогичными у ювенильных особей, то окажется, что кормовая активность беременных перезимовавших самок в 4.5 раза выше. Связано это, по всей видимости, с большим абсолютным количеством потребляемого корма перезимовавшими полевками, имеющими значительно большую массу тела.
Четкой приуроченности отдельных фаз активности и отдыха к восходу или заходу солнца В.А. Соловьеву обнаружить не удалось. Возможно, в условиях белых ночей световой режим не столь жестко определяет суточную ритмику полевок. У семи зверьков, находившихся под наблюдением, отмечалось от 5 до 11 смен фазовых циклов, чаще всего – от 5 до 6. Продолжительность фаз колебалась от 10 мин до 7 ч 15 мин, при этом продолжительность отдыха была от 40 мин до 5 ч 40 мин. В течение суток пики активности распределялись равномерно. Однако в ночное время наступает полоса подъемов пиков активности, которая начинается с 21 ч, продолжаясь с перерывами всю ночь. При этом фазы отдыха значительно уменьшаются. Любопытно, что беременные перезимовавшие самки чаще, чем другие, переходят от активности к покою. У них наблюдается до 11 циклов в течение суток. При этом продолжительность периодов активности резко сокращается, поэтому большую часть времени беременная самка проводит в укрытии. Тем не менее зверьки этой группы съедают наибольшее количество корма.
Общеизвестно, что водяной полевке свойственны приуроченные к определенному времени массовые сезонные перемещения (кочевки), называемые нередко миграциями. Наиболее отчетливые из них – весенняя, в результате которой зверьки перемещаются к воде: на берега рек, озер, к болотам; и осенняя, когда животные перемещаются на поля, залежи, занимают возвышения рельефа. Эти масштабные перекочевки особенно характерны для вида и не раз описывались в литературе (Зверев, Пономарев, 1930; Пантелеев, 1968; Stoddart, 1971, и др.).
Так, согласно данным литературы (Максимов, Ердаков, 1985), в Новосибирской области миграции и заселение зверьками луго-полевых стаций обычно начинается со второй половине июля. Это начало большой осенней миграции – ухода от воды, выхода из болот. Существенным фактором здесь служит необходимость питаться под снегом в течение зимы, а болота для подснежной и поземной зимовки не годятся. В окрестностях Центральной Барабы миграция водяных полевок ежегодно фиксируется в первой декаде августа. При этом между основными миграциями грызунов – осенней и весенней, – существует большое число перемещений зверьков по территории, иногда на значительные расстояния. По материалам П.А. Пантелеева и др., (2001), в Барабе перемещения самцов и самок водяных полевок на площадке живоловок не превышала 60–80 м. При расселении молодых животных этот автор приводит данные о поимке зверьков в 2.5 км от места мечения. На осенних поселениях большинство водяных полевок не отходят от своей норы более чем на 10 м, но случаются и более дальние переходы (в основном самцов) – на 80–120 м.
В общем в зависимости от мотивации, можно выделить передвижения водяных полевок не менее четырех типов. Первый включает постоянные перемещения вблизи норы: дистанция переходов здесь редко превышает 10 м. Второй тип – такие же передвижения по всему индивидуальному участку, дистанция их достигает 600 м. Третий тип передвижений – далекие расселение зверьков; дистанции отмечены до 2.6 км. Четвертый тип перемещений – это регулярные, но только по сезонам смены местообитаний. Последние могут иметь любые расстояния от 10 м, когда водяные полевки устраивают свои зимовочные норы на лугу, рядом с болотом, из которого они вышли, но могут передвигаться до 2–3 км, когда места размножения отделены большими непригодными для зимовки территориями.
Питание. В вегетационный период основу питания водяной полевки составляют различные виды в основном гидрофильных (водно-болотных и прибрежных) травянистых растений тех биотопов, в которых она обитает (рис. 1). Как показало изучение остатков поеденных водяной полевкой растений на 153 кормовых столиках, питание этого вида в Карелии существенно не отличается от уже описанного для большинства других областей ареала (Пантелеев и др., 2001). Ее пища состоит преимущественно из водно-болотной растительности, главным образом, осоки (57.3%), хвощей (38.5%), вахты (27.7%), тростника (12.6%), кубышки (9.9%), кувшинки (8.5%) и некоторых других, причем в отличие от ондатры, водяные полевки редко сгрызают растение под водой и поедают преимущественно его надводные части. В летние корма полевок входят также многие другие виды растений: клевера, таволга, одуванчик, подорожник, осот, лютики, борец, сныть, различные злаки. Они обнаружены на 60.6% исследованных кормовых столиках. Осенью важным компонентом рациона становится также выращиваемый на возделываемых полях и огородах картофель, запасы которого в норах грызунов достигают 10 кг и более.
Рис. 1.
Типы корма водяной полевки в разные сезоны (по: Ивантер, 1975). 1 – корневища, 2 – зеленые части растений, 3 – древесно-кустарниковые корма, 4 – бесхлорофильные побеги, 5 – ягоды.
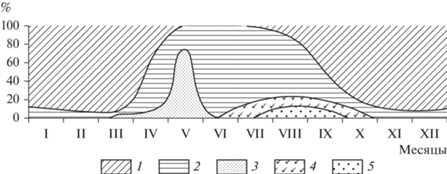
Некоторый интерес представляет опыт кормления молодой водяной полевки в условиях неволи, поставленный нами в заповеднике “Кивач” летом 1961 г. Поскольку аналогичный эксперимент М.Я. Марвина (1959) касался выяснения поедаемости главным образом гидрофильных растений, мы, чтобы избегнуть повторения, пытались изучить потребление в основном лишь суходольной растительности и корнеплодов. В результате удалось установить, что самыми предпочитаемыми видами корма из луговых и полевых растений являются злаки (душистый колосок, полевица, ежа сборная, овсяница красная, тимофеевка, мятлик), некоторые бобовые (клевера, чина луговая, мышиный горошек) и сочное луговое разнотравье (щавель, поповник, тысячелистник, манжетка, таволга); из корнеплодов (скармливали морковь, картофель, свеклу, репу и брюкву) наиболее охотно поедался картофель.
Исследованиями В.А. Соловьева (1994) установлено, что в условиях республики Коми водяная полевка поедает более 50 видов водно-болотных и прибрежных травянистых растений. В весенне-летний период грызуны переключаются на питание надземными частями растений, отдавая предпочтение сочным приземным частям. В этот период на кормовых столиках обычно встречаются остатки различных видов осок. Чаще всего это пузырчатка, осока стройная, осока лисья. Они составляют основу питания. Наблюдая это, нетрудно заметить, что в пищу употребляется однообразный корм, несмотря на широкий выбор кормовых объектов. При вскрытии желудков отловленных зверьков обнаружены три фракции растительной кашицеобразной массы, которые отличались по цвету: белая, светло-зеленая и темно-зеленая. Белая представляла картофель, потребляемый в качестве приманки в конусах, с помощью которых производился отлов, светло-зеленая – остаток переработанных влагалищных, а темно-зеленая – верхушечных частей растений. Никаких остатков животной пищи в желудках водяной полевки обнаружено не было.
Вместе с тем в условиях проведенных опытов был выявлен широкий спектр кормовых растений, причем с явным предпочтением осокам, как правило, удаленным от воды на значительное расстояние. Семи подопытным полевкам в течение нескольких дней предлагалось 25 видов растений в различных сочетаниях в виде букетиков. Полевки активнее поедали сочные луговые растения, особенно молодые побеги заячьей капусты. Чаще съедали таволгу вязолистную, все вегетативные части горошка заборного, чины весенней, листья и молодые побеги иван-чая узколистного. Менее охотно, но достаточно хорошо поедали хвощ лесной, листья клевера лугового, у которого съедобными оказались все наземные части; различные виды осок, камыш лесной. У камыша и осок охотно съедались влагалищные части. Не менее часто зверьки ели листья водокраса лягушачьего, листья и стебли вахты трехлистной. Белокрыльник болотный, зверобой четырехгранный, щавель кисловатый и подорожник средний поедались незначительно, и совсем плохо – черноголовка обыкновенная, валериана аптечная, пижма обыкновенная, подмаренник мягкий, щавель кислый.
В зимний период и в самом начале вегетации (конец мая–первая половина июня), по наблюдениям на северо-востоке Европейской части России (Бобрецов, 2004), водяная полевка особенно охотно поедает подземные части растений – корневища борца северного, чемерицы Лобеля, вейника пурпурного и др. С этой целью она выгрызает в земле специальные кормовые ходы, скусывает растения и затаскивает их под землю, где и кормится ими. В это время в их норах иногда находили фрагменты корневищ общим весом до 2 кг. По данным ряда авторов инстинкт запасания кормов в осенне-зимний период – довольно обычное явление для этого вида на севере Европейской части и Сибири (Жарков, Теплов, 1931; Формозов, 1948; Максимов, Андрусевич, 1959; Соломонов, 1980 и др.).
В небольшом количестве потребляют эти полевки и животный корм. В июне на кормовых столиках в поселениях этих зверьков, расположенных в подгольцовых лугах Печеро-Илычского заповедника, неоднократно находили кучки недоеденных наземных моллюсков (Бобрецов, 2004). По мнению П.А. Пантелеева (2001), необходимость в животной пище возникает у зверьков чаще всего весной для утоления белкового голода.
Размножение и структура популяции. Согласно данным литературы (Пантелеев и др., 2001), водяная полевка способна вступать в размножение в возрасте 40 дней. Однако, по нашим данным, полученным в Карелии, у животных, родившихся в конце лета–начале осени, половое созревание задерживается нередко до весны, и зрелость наступает лишь в возрасте 7–8 мес. и более в зависимости от экологических условий.
Возрастное половое созревание летом в год своего рождения раньше наступает у самок. Молодые самцы половозрелыми становятся позже, примерно на 7–10 дней. По данным Я. Пеликана (Pelikan, 1972), половая активность молодых самок наступает при массе тела от 40 до 60 г, молодых самцов – от 60 до 89 г; популяция достигает 50%-ной зрелости среди числа молодых, когда средний вес тела самок достигает 76 г, а самцов – 88 г.
Половая зрелость у перезимовавших зверьков, по нашим наблюдениям, наступает раньше у самцов. При этом начало полового созревания связано с фотопериодом и сопровождается ускорением роста животных (Евсиков и др., 1991). Кроме того установлено, что срок полового созревания самок-сеголеток зависит от фазы популяционного цикла численности (Пантелеев и др., 2001; Назарова, Проскурняк, 2017). На подъеме численности, по сравнению с пиком и спадом, самки становятся половозрелыми при меньшем весе тела и вступают в размножение в более ранние календарные сроки.
Срок беременности, установленный в лабораторных условиях, равен 22 дням, по сведениям же Г. Рейхштейна (Handbuch, 1982) – 20–23 дням, в среднем 21.6 (n = 23), а по данным Н.В. Тупиковой (Тупикова, Швецов, 1956) – 21 дню.
Сезон размножения от начала попадания первых беременных самок весной до последних только что разродившихся самок осенью длится 4–6 месяцев, в зависимости от состояния популяции и погодно-фенологических условий года. Таким образом, в принципе, водяную полевку следует считать сезонно размножающимся видом. При этом от продолжительности сезона размножения зависит и число пометов в году.
Первое спаривание зимовавших водяных полевок проходит в Карелии в начале мая. Семенники добытых в это время самцов имели в длину 16–21 мм, семенные пузырьки – 9–25 мм, вес двух семенников – 1.7–2.8 г; сперматогенез обнаружен у всех исследованных самцов (табл. 2). Все самки, пойманные во второй половине мая, были беременны (80%) или уже кормили выводок (20%). В июне эмбрионы найдены у половины взрослых самок, остальные были лактирующими. Иногда беременность и лактация у них совпадают. В первой декаде июня (самая ранняя дата 8.06.1951) отловлены первые молодые зверьки весом 33–75 г, но в массе они появляются во второй половине этого месяца. Беременные и лактирующие зимовавшие самки попадались до конца августа (в июле 44% беременные и 22% кормящие, в августе 50% беременные). Эти данные, а также тот факт, что молодые зверьки приблизительно одного и того же возраста ловились в совершенно разное время, показывают, что период размножения водяной полевки в Карелии сильно растянут. Пользуясь таблицами для определения возраста молодых водяных полевок, приведенными В.А. Поповым (1960) и П.Л. Пантелеевым (1968), мы установили, что отловленные нами в период с 8 июня по 24 июля сеголетки находились в возрасте 30–60 дней, т. е. родились в мае–первой половине июня (Ивантер, 1975, 2018). В то же время в августе вновь стали попадаться молодые зверьки 20–30-дневного возраста. Таким образом, у зимовавших водяных полевок имеется не менее двух пометов: первый в мае и второй (его рождение более растянуто во времени) в июле–августе. Не исключено, что у части взрослых самок имеется и третий помет, но прямого подтверждения этому нет. Наличие у большинства водяных полевок Карелии не менее двух выводков в течение года доказывается и результатами исследования половых органов взрослыx самок в июле–августе (на рогах матки – следы рождения двух пометов), а также поимкой в июле самок, одновременно подсосных и беременных.
Таблица 2.
Состояние половой системы и участие в размножении самцов и самок водяной полевки в Карелии
| Месяц | Число исследованных зверьков | Характеристика половой системы | Доля самцов со сперматогенезом, % от общего числа | Число самок | |||||
|---|---|---|---|---|---|---|---|---|---|
| длина семенника, мм | длина семенных пузырьков, мм | Вес двух семенников, г | беременных | кормящих | рожавших | без следов размножения | |||
| Взрослые (зимовавшие) | |||||||||
| Май | 15 | 17.4(15.6–20.1) | 18.9(9–24.5) | 2.0(1.1–2.8) | 100 | 4 | 1 | – | – |
| Июнь | 15 | 16.8(10.5–18.7) | 18.2(8–28.4) | 2.1(0.6–2.6) | 89 | 3 | 3 | – | – |
| Июль | 21 | 16.7(11–19.6) | 18.0(9.5–25.0) | 2.3(0.6–2.6) | 100 | 4 | 2 | 2 | 1 |
| Август | 5 | 12.8(9.8–15.9) | 10.3(8–13.1) | 1.4(0.2–2.1) | 67 | 1 | – | 1 | – |
| Сентябрь | 10 | 13.0(4.8–15.1) | 12.3(10–13.6) | 0.7(0.3–1.5) | 40 | – | – | 3 | – |
| Прибылые (сеголетки) ранних выводков | |||||||||
| Июнь | 12 | 13.5(3.5–14.9) | 7.5(3.7–10.2) | 1.0(0.8–1.3) | 82 | – | – | – | 6 |
| Июль | 28 | 9.3(3.6–17) | 10.1(2–23) | 0.6(0.05–2.6) | 58 | 2 | 2 | 1 | 5 |
| Август | 47 | 6.1(4.1–15.9) | 7.1(2.5–16) | 0.2(0.05–1.1) | 27 | 4 | 2 | 4 | 13 |
| Сентябрь | 12 | 7.3(4.1–15) | 3.2(2–9.8) | 0.4(0.03–1.4) | 17 | – | – | 3 | 3 |
| Прибылые (сеголетки) поздних выводков | |||||||||
| Июль–сентябрь | 82 | 4.7(3.5–8.6) | 3.0(2–4.5) | 0.08(0.04–0.2) | 0 | – | – | – | 40 |
При этом число пометов в году, помимо прочего, зависит от продолжительности сезона размножения. До середины 1950-х гг. число пометов определяли обычно расчетным числом: от начала сезона размножения прибавляли срок беременности, срок полового созревания, наступление новой беременности и так до срока окончания сезона репродукции. Лишь Н.В. Тупикова и Ю.Г. Швецов (1956) показали, что перезимовавшие самки и самки-сеголетки дают разное количество пометов, и что между пометами у самок бывает перерыв в размножении, имеющий больший срок, чем это допускалось при расчетах.
Прибылые водяные полевки размножаются в Карелии с гораздо меньшей интенсивностью, чем зимовавшие, и участие их в размножении зависит от времени рождения. Разделение сеголеток на две возрастные группы (родившиеся в мае–первой половине июня и в июле–августе) показало, что в год рождения размножается только 70% самок и 45% самцов ранних выводков, а прибылые, родившиеся во второй половине июня–августе, в текущем году вообще не достигают половой зрелости и не размножаются (табл. 2). При этом большинство размножающихся самок-сеголеток успевает принести за первый сезон только один выводок (в наших сборах их было 70.6%) и лишь около трети (29.4%) – два выводка. Тем не менее значение прибылых особей в воспроизводстве популяции и нарастании ее численности не менее существенно, чем зимовавших полевок.
При этом раннее начало размножения и низкая численность весенней популяции вызывают более активное размножение молодых и роль их в воспроизводстве популяции еще более возрастает. Таким образом, в Карелии, как и в других местах (Шварц и др., 1957; Максимов, 1966 Попов, 1960; Соломонов, 1980; Пантелеев, 1968, и др.), ведущее значение в нарастании численности популяции имеют зверьки первого весеннего помета, интенсивность размножения которых связана с весенними условиями существования и уровнем плотности популяции. Заканчивается размножение в августе, причем зимовавшие самки прекращают размножаться на 2–3 недели раньше, чем сеголетки.
По данным Г.Г. Назаровой и Л.П. Проскурняк, (2017), регулирование репродуктивного вклада сеголеток осуществляется в основном за счет изменения скорости полового созревания самок. На пике и спаде численности размножается не более 26% прибылых самок (имеется в виду, очевидно, вся популяция сеголеток); в период депрессии их доля начинает увеличиваться, а на подъеме достигает 60%. В результате около 50% прибылых зверьков на подъеме численности – это потомки сеголеток, тогда как на пике и спаде их доля не превышает 3% (Евсиков и др., 1997).
В общем же, весь сезон размножения от начала попадания первых беременных самок весной до последних только что разродившихся особей осенью длится у нас в отдельные годы от 4 до 6 мес. в зависимости от местных погодных условий (срока наступления весны, ее хода и т.п.).
Важным показателем репродуктивного потенциала популяции принято считать число детенышей в помете (разовая плодовитость) (табл. 3). Вообще-то этот показатель у водяной полевки отличается высокой изменчивостью (Пантелеев, 2001). Его колебания связаны с климатическими условиями, в которых находится данная популяция, также с географической широтой региона. Чем в более холодном климате – в более высоких широтах или выше над уровнем моря – находится популяция, тем большее среднее число детенышей в помете ей свойственно. Среди прочего это связано с необходимостью компенсировать репродуктивные потери, которые вынуждена нести популяция в связи с сокращением периода размножения и повышенной эмбриональной и постнатальной смертностью.
Таблица 3.
Плодовитость самок водяной полевки в Карелии
| Возрастная группа | Число исследованных самок | Количество самок (экз.) с числом эмбрионов | Среднее число эмбрионов на 1 самку | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | |||
| Зимовавшие | 27 | 1 | – | 2 | 3 | 8 | 6 | 4 | 2 | 1 | 6.5 (3.4 + 3.1) |
| Прибылые | 16 | – | 1 | 2 | 6 | 4 | 1 | – | 1 | 1 | 5.7 (3.0 + 2.7) |
Средняя величина выводка у зимовавших и прибылых самок водяной полевки в нашем регионе оказалась близка к видовой норме и составляет соответственно 6.5 ± 0.37 и плодовитости 5.7 ± 0.41 (табл. 3). В первом случае имеет место стабилизация средней величины плодовитости (показатель асимметрии А = –0.01), а во втором – процесс ее увеличения еще продолжается (Ивантер, 1975). Это говорит о том, что отбор на повышенную плодовитость затрагивает в основном молодых самок, а у взрослых он уже завершился.
Важным фактором, определяющим снижение репродуктивного потенциала водяной полевки, является эмбриональная смертность. Как убедительно показали специальные исследования в Северной Барабе (Евсиков и др., 1991), смертность эмбрионов способна изменяться в разные фазы многолетних циклов динамики численности популяции в 2–3 раза. В фазе спада численности уровень этой формы эмбриональной смертности достигал там 52%, а на подъеме он снижался до 4%.
В условиях таежного Северо-Запада России эмбриональная смертность водяной полевки, вычисленная по разности между числом желтых тел и живых эмбрионов, равна 2.7%. Это близко к данным по Западной Сибири (1.7%) и низовьям Волги (2.5%), но ниже, чем в Татарии (7.2%).
Эмбриональная и ранняя постнатальная смертность определяет фактическую плодовитость – число выращенных потомков. При этом и та, и другая обязаны своим существованием стрессовым явлениям в популяции (Евсиков и др., 1991). После того, как популяция в течение нескольких репродуктивных сезонов находится в фазе депрессии и на уровне низкой численности, она освобождается от стрессовых явлений, и при наличии высокой емкости угодий начинает нормально реализовывать свой репродуктивный потенциал. Для достижения нормального (среднего) уровня численности, популяция, очевидно, не нуждается в каких-то особых факторах: достаточно наличия определенной емкости угодий и снятия популяционного стресса.
Выше уже указывалось, что, продолжительность жизни водяной полевки, по наблюдении в виварии, составляет около 2 лет, в природных же условиях, видимо, не более 22 мес. Кроме того, размножаются у этого вида не только взрослые (перезимовавшие) и сеголетки ранних рождений, но и прибылые, родившиеся позднее, включая и появившихся на свет в конце лета. Это еще несколько усложняет возрастную структуру, характерную для данного вида. Представленные на рис. 2 и в табл. 4 сезонные изменения возрастного состава отловленных в природе водяных полевок показывают постепенное “разбавление” зимовавшей части популяции сеголетками. В июне на их долю приходится 47.8%, в июле – 54.6, в августе–сентябре – 90, в октябре – 100%. При этом вместе с зимовавшими полевками из популяции исчезают и прибылые ранних выводков. Их, очевидно, можно отнести к скороспелой весенней генерации, сочетающей в себе быстрое половое развитие с коротким жизненным циклом. Поэтому весеннее поголовье водяных полевок однородно в возрастном отношении и состоит преимущественно из зверьков, родившихся во второй половине лета прошлого года.
Рис. 2.
Сезонные изменения возрастного состава популяции водяной полевки по данным отлова. 1 – зимовавшие; 2 – прибылые ранних выводков; 3 – прибылые поздних выводков. По оси абсцисс: месяцы; по оси ординат: относительное число зверьков определенного возраста, % от общего числа.
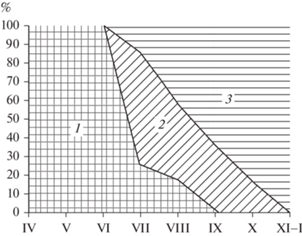
Таблица 4.
Возрастной и половой состав популяции водяной полевки в Карелии по данным многолетних учетов
| Месяц | Число прибылых | Число зимовавших | Общее число исследованных зверьков | |||||
|---|---|---|---|---|---|---|---|---|
| абс. | из них самцы, % | χ2 | абс. | из них самцы, % | χ2 | абс. | из них зимовавшие, % | |
| Май | – | – | – | 13 | 61.5 | 0.7 | 13 | 100.0 |
| Июнь | 13 | 53.8 | 0.1 | 14 | 58.3 | 0.3 | 27 | 48.0 |
| Июль | 43 | 58.1 | 1.1 | 22 | 54.5 | 0.2 | 65 | 33.8 |
| Август | 65 | 50.8 | 0.01 | 6 | 50.0 | – | 71 | 5.8 |
| Сентябрь | 59 | 54.3 | 0.6 | 10 | 70.0 | 1.6 | 69 | 14.5 |
| Октябрь | 2 | – | – | – | – | – | 2 | – |
| Декабрь | 2 | 50.0 | – | – | – | – | 2 | – |
| Всего | 184 | 53.3 | 0.8 | 65 | 59.0 | 2.0 | 249 | 26.1 |
Тем не менее, в северо-западных частях ареала, а значит, и на его периферии, куда относится и изучаемый нами регион, возрастная структура популяции водяной полевки имеет свои особенности. Как уже указывалось, это прежде всего относительная ее сложность, являющаяся следствием активного участия в размножении трех возрастных когорт, что может считаться и одной из весьма характерных для пограничных популяций адаптаций к повышенной здесь смертности зверьков.
Напомним, что период массового размножения водяных полевок в Карелии обычно начинается в первой декаде мая и заканчивается уже в августе. За эти четыре месяца большинство перезимовавших самок успевает принести два помета, размножается и часть молодых самок первых двух пометов. В отдельные годы с неблагоприятными погодно-климатическими условиями многие самки вообще не размножаются. С таким “замедленным” размножением связана и сложность возрастной структуры осенней популяции. Она представлена в два раза большим числом генераций, по сравнению, например, с западносибирскими популяциями из Барабинской низменности. Тем не менее, воспроизводство популяции водяной полевки в Карелии обеспечивается в основном за счет размножения перезимовавших самок. Из самок-сеголеток в размножении принимает участие лишь некоторая часть особей первой генерации и то не каждый год. Однако именно в годы более полного участия прибылых в воспроизводстве популяции создаются необходимые условия для заметного ее усложнения и увеличения численности вида.
Среди пойманных зимовавших полевок преобладают самцы, очевидно, в связи с их большей активностью в период размножения. Молодые самцы также количественно преобладают над самками, но диспропорция эта невелика и статистически недостоверна.
Изменения численности. Годовые изменения численности водяных полевок довольно значительны, что в свое время сказывалось и на динамике заготовок шкурок (рис. 3). Максимальная за 40 лет величина добычи (1940 г.) превышает минимальную (1961 г.) в 22 раза, причем за смежные годы объем промысла изменялся в 3–6 раз. Основными причинами динамики численности водяной полевки служат изменения гидрологического режима водоемов (неблагоприятный водный режим был, в частности, главной причиной снижения численности водяной полевки и ондатры в 1955–1961 гг.; Ивантер, 1965, 1975) и вспышки туляремии, что наблюдалось в Карелии в 1959–1960 гг. (Андреев, 1961). Последнему способствует высокая зараженность водяных полевок Карелии эктопаразитами (экстенсивность до 70 и даже 100% при общей интенсивности 0.5–2.2).
Рис. 3.
Кривая заготовок шкурок водяной полевки в Карелии в 1932–1962 гг. По оси абсцисс: годы; по оси ординат: число шкурок.
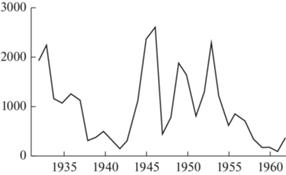
Вообще же, на севере европейской части страны (поймы рек Северная Двина, Печера, Вычегда, Сухона, Онега, а в Карелии – Суна и Кемь) по литературным данным (Пантелеев и др., 2001) вспышки массовых размножений водяной полевки отмечались в 1904, 1907, 1923 и в 1926–1928 гг. (пик в 1927 г.). Затем, судя по заготовкам шкурок, в поймах рек Архангельской обл. (вместе с Ненецким автономным округом, где основной помысел велся в дельте р. Печоры) и Вологодской обл. (р. Сухона и др.), максимально высокие подъемы численности грызунов происходили в середине и в первой половине 1950-х гг. (рис. 3). Сходная динамика имела место в Коми и у нас в Карелии (1927, 1934–1937, 1955–1956 гг. и менее значительные подъемы в 1942, 1949, 1952, 1960 и 1968 гг.).
Обнаруживаются и некоторые различия в динамике численности водяных полевок в поймах разных северных рек. Это подтверждают и данные Б.П. Доброхотова и В.М. Неронова (1965). Так, в долине р. Онеги (нижнее течение) в период с 1950 по 1962 г. по их данным наблюдалось три подъема численности зверьков: в 1952, 1956, 1959–1960 гг.; в долине р. Северной Двины, в основном ниже впадения в нее р. Пинеги и в дельте, отмечались три волны высокой численности с пиками в 1952, 1957, 1961 гг.; в долине р. Мезени и ее притока – р. Башки – было также три подъема численности вида: в 1952, 1956–1957, 1960 гг.; в пойме р. Печоры и по ее притокам Ижме, Пижме и Цильме – в 1952, 1955 и 1956 гг.
При этом интенсивность вспышек массового размножения в поймах северных рек достигала весьма высоких показателей. Так, в дельте Печоры, где складывались наиболее благоприятные на севере условия для жизни водяных полевок, заготовки превышали 0.5 млн шкурок в год (1953), в Коми – 380 тыс. (1934 г.). В Вологодской обл. максимальная заготовка была в 1935 г. (9 тыс.).
Однако при некоторых различиях периодов развития вспышек массового размножения в бассейнах разных рек наблюдались и периоды синхронных подъемов численности. Из 36 краев и областей европейской части страны, по которым имеются данные о заготовках шкурок за довоенные годы, пики численности помимо общего подъема в 1956–1959 гг. регистрировались в 1933–1935 гг. (в 21 регионе) и в 1940–1941 гг. (14 регионов).
Что же касается связи массовых размножений этого вида с цикличностью солнечной активности (Максимов, 1966; Пантелеев, 1968), то на нашем материале она не прослеживается. Из четырех достоверных подъемов численности водяной полевки в Карелии с максимумом солнечной активности совпал лишь один (1949–1950 гг.), а остальные (1932, 1945–1946 и 1953) пришлись на минимум или спад цюрихских среднегодовых чисел Вольфа. Это, конечно, не значит, что космические факторы вообще не влияют на численность водяной полевки, но они опосредованы многолетними колебаниями гидрологического режима водоемов и сильно затушевываются действием внутрипопуляционных и биоценотических факторов. В очагах массового размножения рост численности может затормаживаться авторегуляторно через стресс (Евсиков и др., 1991), а также путем компенсаторной перестройки возрастной структуры размножающихся зверьков. В данном случае речь идет о более массовом половом созревании и об участии в размножении сеголеток-первопометников в условиях разреженной популяции и о почти полном не созревании их при переуплотнении населения. Компенсаторная перестройка возрастной структуры за счет изменения скорости полового созревания у полевок чаще всего связана именно с эффектами хронического стресса, развивающейся при высокой плотности популяции.
Вместе с тем, как было показано С.С. Шварцем с соавторами (1957), интенсивность размножения популяции тесно связана с весенними условиями существования. Ранняя и благоприятная по гидроклиматическим условиям весна способствует более дружному и раннему началу размножения зимовавших полевок, раннему половому созреванию и более полному вступлению в размножение сеголеток весенне-летних выводков. При раннем начале сезона размножения как зимовавшие самки, так и сеголетки первого весеннего помета дают за лето по “лишней” генерации (Пантелеев, 1968). В то же время сроки и характер весны могут быть связаны с солнечной активностью. Таким образом, численность водяной полевки, как и других мелких млекопитающих Карелии, определяется совокупным действием многих факторов, каждый из которых может превалировать в определенные годы и определенных ситуациях.
Заключение. Как показали проведенные исследования, выявленные выше экологические особенности, свойственные водяной полевке в специфических условиях северо-западной периферии ареала, отличаются значительной лабильностью и гармоничной согласованностью как с плотностью и состоянием популяции, так и с внешними факторами среды. Разнообразие экологически приспособлений, их трансформация и смена во времени и пространстве, адаптивный динамизм и ориентированность на конкретную экологическую ситуацию, складывающуюся из взаимодействия экзогенных и эндогенных факторов, обуславливают относительную стабильность и оптимальное состояние популяций рассматриваемого вида в экстремальных для него приграничных условиях. Вместо узких специальных приспособлений индивидуального ранга в периферических его населениях успешно реализуются широкие структурно-популяционные адаптивные комплексы, отличающиеся динамичностью и высокой скоростью компенсаторной перестройки. Весьма характерны для обитающей на северо-западе ареала водяной полевки, имеющие существенное адаптивное значение синантропизм в биотопическом размещении, трехкогортность участия зверьков в репродукции и значительно более сложная, чем в оптимуме ареала, экологическая структура популяции. Определенную роль в жизни и особенно воспроизводстве периферических популяций водяной полевки могут играть и специфические регуляторные, в том числе стресс-факторные, механизмы направленного действия, осуществляющие поддержание численности на оптимальном уровне. Однако у северо-западных границ ареала действие эти механизмов более ограничено и поддержание динамического баланса популяции с ресурсами ее местообитания в основном достигается при более значительном участии внешних факторов. Все это подтверждает выдвинутую нами ранее концепцию периферических популяций (Ивантер, 1975, 1985, 2008, 2017), согласно которой в экологическом центре (оптимуме) видового ареала животных население вида благодаря функционированию внутрипопуляционных компенсаторных механизмов отличается не только высоким уровнем численности, но и большей устойчивостью и однородностью, тогда как у границ распространения действие этих механизмов более ограничено, и организация населения изменяется в более широком диапазоне и с большей амплитудой. Вместе с тем для периферически популяций водяной полевки характерны и так называемые “популяционные волны”, возникающие с чрезвычайно резкими и неритмичными изменениями численности. Соответственно более выражены и гораздо рельефнее проявляются вблизи границ ареала и специфические структурно-популяционные адаптации, направленные в частности на максимально эффективное воспроизводство населения, способное обеспечить виду и его популяциям необходимое преодоление экстремальных условий экологического пессимума. Таково, в частности, отмеченное нами для северо-западных зон ареала водяной полевки более полное и частое созревание и участие в размножении прибылых особей.
Финансирование. Работу проводили в соответствии с Государственным заданием федерального исследовательского центра “Карельский научный центр РАН”.
Список литературы
Андреев И.Ф. Численность микромаммалия в Карелии и факторы, определяющие ее // Тез. докл. конфер. по итогам работ Ин-та биол. Карельск. фил. АН СССР за 1960 г. Петрозаводск. 1961. С. 118–120.
Бобрецов А.В. Мышевидные грызуны // Млекопитающие Печоро-Илычского заповедника. Сыктывкар: Коми. книжн. изд-во, 2004. С. 206–301.
Громов И.М., Ербаева М.А. Млекопитающие фауны России и сопредельных территорий. Зайцеобразные и грызуны. – СПБ., 1995. 522 с
Громов В.С. Пространственно-этологическая структура популяций грызунов. М.: Т-во научн. изданий КМК, 2008. 581 с.
Доброхотов Б.П., Неронов В.Н. Поселения водяных крыс и природные очаги туляремии на Севере Европейской части СССР // Туляремия и сопутствующие инфекции // Омск. 1965. С. 211–216.
Евсиков В.И., Мошкин М.П., Назарова Г.Г. Реализация репродуктивного потенциала и стресс в цикле численности водяных полевок // Экология популяций. М., 1991. С. 213–228.
Евсиков В.И., Назарова Г.Г., Потапов М.А. Генетико-экологический мониторинг циклирующей популяции водяной полевки (Arvicola terrestris L.) на юге Западной Сибири // Генетика, 1997. Т. 33. № 8. С. 1133–1143.
Ердаков Л.И. Циркадный ритм активности и питание водяной полевки и обыкновенного хомячка в условиях эксперимента. Автореф. дис. канд. биол. наук. Новосибирск, 1972. 23 с.
Ердаков А.Н. Организация ритмов активности грызунов. Новосибирск: Наука, 1984. 181 с.
Жарков М.Д., Теплов В.П. Данные по биологии и промыслу водяной крысы в Татарской республике // Работы Волжско-Камской краевой промысловой станции. Казань.1931. Вып. С. 17–28.
Зверев М.Д., Пономарев М.Г. Биология водяных крыс и отравленные приманки в борьбе с ними // Изв. Сибкрайстазра. Новосибирск.1930. Т. 4(7). Вып. 1. С. 26–47.
Зудова Г.А. Сезонно-возрастная динамика состава лейкоцитов крови у водяной полевки // Актуальные вопросы ветеринарной биологии, 2014. № 3(23). С. 79–87.
Ивантер Э.В. Состояние запасов и причины снижения численности ондатры в Карельской АССР // Проблемы ондатроводства. М., 1965. С. 27–28.
Ивантер Э.В. Материалы по размножению ондатры карельской популяции // Уч. зап. Петрозаводск. гос. ун-та. Петрозаводск. 1966. Т. 14, вып. 3. С. 109–116.
Ивантер Э.В. Популяционная экология мелких млекопитающих таежного Северо-Запада СССР. Л.: Наука.1975. 318 с.
Ивантер Э.В. Основы практической биометрии. Введение в статистический анализ биологически явлений. Петрозаводск: Изд-во “Карелия”, 1079. 96 с.
Ивантер Э.В. Эволюционная роль периферических популяций // Микропопуляция. М. 1985. С. 80–84.
Ивантер Э.В. К изучении популяционной организации политипического вида // Уч. зап. Петр. ГУ. 2008. № 1. С. 39–60.
Ивантер Э.В. К разработке экологической концепции периферических популяций // Экология. 2017. № 1. С. 60–65.
Ивантер Э.В. Очерки популяционной экологии мелких млекопитающих на северной периферии ареала. М.: Т-во научн. изд. КМК. 2018. 70 с.
Ивантер Э.В, Коросов А.В. Основы биометрии. Введение в статистический анализ биологических явлений и процессов. Петрозаводск. Изд-во Петр. ГУ, 1992. 168 с.
Ивантер Э.В., Коросов А.В. Введение в количественную биологию: Учеб. пособие. Петрозаводск: Изд-во Петр. ГУ. 2003. 304 с. .
Карасева Е.В., Телецына А.Ю., Жигальский О.А. Методы изучения грызунов в полевых условиях. М.: Изд-во ЛКИ, 2008. 416 с.
Максимов А.А. Размножение и изменение численности водяной крысы в различных ландшафтах Западной Сибири. Распределение водяной крысы по зонам и ландшафтах Западной Сибири // Водяная крыса и борьба с ней в Западной Сибири. Новосибирск. 1959. С. 19–48, 71–121.
Максимов А.А. Цикличность массовых размножений водяной крысы в Западной Сибири в сопоставлении с солнечной активностью // Вопр. зоологии. Томск. 1966. С. 232–241.
Максимов А.А., Андрусевич Е.К. Убежища (норы и гнезда) и питание водяной крысы // Водяная крыса и борьба с ней в Западной Сибири. Новосибирск. 1950. С. 49–71.
Максимов А.А., Ердаков Л.Н. Циклические проблемы и сообщества животных. Новосибирск: Наука СО. 1985. 236 с.
Марвин М.Я. Млекопитающие Карелии. Петрозаводск.1959. 237 с.
Назарова Г.Г., Проскурняк Л.П. Возрастная изменчивость репродуктивных показателей водяной полевки (Arvicola terrestris L.) // Вестник Ир. ГСХА. 2017. № 83. С. 141–145.
Пантелеев П.А. Популяционная экология водяной полевки и меры борьбы. М., 1968. 254 с.
Пантелеев П.А. и др. Водяная полевка. М.: Наука, 2001. 387 с.
Петров П.А. Видовой состав и эволюция грызунов Волго-Ахтубинской поймы // Тр. Ростовского н/Д НИИПЧИТ. 1956. Т. 3. С. 127–146.
Плюснин Ю.М., Евсиков В.И. Сезонные изменения формы организации репродуктивных групп водяной полевки // Популяционная изменчивость вида и проблемы охраны генофонда млекопитающих. М.: Наука, 1983. С. 53–55.
Попов В.А. Млекопитающие Волжско-Камского края. Казань. 1960. 466 с.
Рогов В.Г. Динамика численности и демографические параметры популяции водяной полевки в подтаежной зоне Западной Сибири. Автореф. дис. … канд. биол. наук. Новосибирск. 1999. 23 с.
Соловьев В.А. Водяная полевка // Фауна Европейского Северо-Востока России. Млекопитающие. Т. II. Ч. I. СПб.: Наука. 1994. С. 224–236.
Соломонов Н.Г. Экология водяной полевки в Якутии. Новосибирск: Наука. 1980. 26 с.
Тупикова Н.В., Швецов Ю.Г. Размножение водяной крысы в Волго-Ахтубинской пойме // Зоол. журн. 1056. Т. 35. Вып. 1. С. 130–140.
Формозов А.Н. Мелкие грызуны и насекомоядные Шарьинского района Костромской области в период 1930–1940 гг. // Матер. по грызунам. М. Изд-во МОИП. 1948. Вып. 3. С. 3–110.
Шварц С.С., Павлинин В.И., Сюзюмова Л.М. Теоретические основы построения прогнозов численности мышевидных грызунов в лесостепном Зауралье // Тр. Ин-та биол. Уральск. фил. АНСССР. Свердловск. 1957. Вып. 8. С. 3–59.
Handbuch der Saugetiere Europas /Niethamme J. Krapp. F. (eds.). Bd. 2/1. Nagetiere 2. – Wiesbaden: Acad. Vetag., 1982. 649 s.
Pelican J. Arvicola terrestris (L.), indexes of reproduction i Czechoslovakia // Prirodoved pr Ustavu CSAV Brno. 1972. V. 6. № 11. 50 p.
Stoddart M.D. Breeding and survival in a population of Water voles // J. Ann. Ecol. 1971. V. 40. № 2. P. 487–494.
Дополнительные материалы отсутствуют.
Инструменты
Известия РАН. Серия биологическая