

Агрохимия, 2019, № 6, стр. 64-71
ЗАВИСИМОСТЬ ЭФФЕКТИВНОСТИ ИСПОЛЬЗОВАНИЯ СОЛОМЫ ЗЕРНОВЫХ КУЛЬТУР С ДОПОЛНИТЕЛЬНЫМИ КОМПОНЕНТАМИ ОТ ПОГОДНЫХ УСЛОВИЙ ГОДА
И. В. Черепухина 1, 2, *, Н. В. Безлер 1, М. В. Колесникова 1
1 Всероссийский научно-исследовательский институт сахарной свеклы и сахара им. А.Л. Мазлумова
396030 Воронежская обл., Рамонский р-н, пос. ВНИИСС, 86, Россия
2 Воронежский государственный университет
394018 Воронеж, Университетская площадь, 1, Россия
* E-mail: icherepukhina@gmail.com
Поступила в редакцию 30.05.2018
После доработки 09.07.2018
Принята к публикации 10.12.2018
Аннотация
Показано, что запашка соломы зерновых культур совместно с целлюлозолитическим микромицетом (Humicola fuscoatra ВНИИСС 016) в различных гидротермических условиях была одним из способов ускорения трансформации соломы и поддержания эффективного плодородия почвы. В стационарном полевом опыте показано, что при ускорении процесса деструкции соломы увеличивалось содержание энергетического материала для микроорганизмов, благодаря чему активизировалась их жизнедеятельность. Целлюлозолитический микромицет способствовал ускорению гумификационных процессов, что подтверждено соотношением численности зимогенной и автохтонной групп микроорганизмов, которое выросло в 2015 и 2016 гг. соответственно до 4.82 и 4.95. Было отмечено развитие целлюлозолитических микроорганизмов при использовании H. fuscoatra, и если в 2013 г. была выявлена тенденция к росту их численности, то в 2015, 2016 и 2017 гг. отмечено достоверное ее увеличение на 25.9, 24.2 и 51.7% соответственно. Обнаружено увеличение численности диазотрофов, что непосредственно влияло на накопление доступных для растений форм соединений азота, в частности, щелочногидролизуемого. Благодаря использованию H. fuscoatra и других компонентов, высокой активности микрофлоры почвы, вовлекавшей в круговорот элементы питания, был отмечен рост урожайности сахарной свеклы: прибавки составили от 5.5 до 12.0 т/га.
ВВЕДЕНИЕ
Использование соломы зерновых культур в настоящее время может быть одним из способов сохранения и поддержания почвенного плодородия, т.к. она является наиболее доступным органическим удобрением. Кроме того, в производственных условиях на полях, где соблюдают зернопаропропашной севооборот, наименее энергозатратным может быть использование 2-х видов соломы в количестве, которое остается после уборки.
В зависимости от местных условий время разложения соломы зависит от типа почвы, ее физических, химических и биологических свойств, климатических условий, а также от вида и состава запахиваемого растительного материала [1, 2].
Структура и состав органических остатков, а также соотношение в них углерода к азоту влияют на интенсивность их последующего превращения. Ускоренная минерализация и быстрое возникновение в почвах дефицита свежего органического вещества усиливает микробиологическую нагрузку на гумус как источник питания, что приводит к более интенсивному его разложению и снижению плодородия [3].
В составе соломы озимой пшеницы и ячменя соотношение углерода к азоту очень широкое и составляет 70–80 : 1, это замедляет скорость ее разложения [4]. Поэтому на начальных этапах ее деструкции происходит значительное биологическое закрепление азота, и возделываемые растения испытывают дефицит этого элемента [5, 6]. Для компенсации потерь азота в процессе трансформации соломы необходимо дополнительное внесение азотного удобрения, а также внедрение способов, ускоряющих процесс разложения растительного материала [2, 7]. При соблюдении этих рекомендаций почва обогащается органическим веществом и в ней активизируются мобилизационные процессы [8].
Цель работы – оценка активности процессов трансформации соломы двух видов зерновых культур в паровом звене зернопаропропашного севооборота под влиянием дополнительных компонентов и формирования эффективного и потенциального плодородия.
МЕТОДИКА ИССЛЕДОВАНИЯ
В 2001 г. в лаборатории эколого-микробиологических исследований почв Всероссийского научно-исследовательского института сахарной свеклы и сахара им. А.Л. Мазлумова из чернозема выщелоченного был выделен аборигенный штамм целлюлозолитического микромицета (Humicola fuscoatra ВНИИСС 016). Лабораторные и полевые исследования показали его положительное влияние на скорость разложения соломы: у соломы озимой пшеницы она увеличилась на 50, у соломы ячменя – на 56% [9, 10].
В 2010 г. на опытном поле ВНИИСС был заложен многолетний стационарный полевой опыт с запашкой соломы озимой пшеницы и ячменя в зернопаропропашном севообороте с чередованием культур: пар–озимая пшеница–сахарная свекла–ячмень. Почва опытного участка – чернозем типичный тяжелосуглинистый малогумусный на покровных лессовидных суглинках. Севооборот представляет собой паровое звено зернопаропропашного севооборота с чередованием культур: пар–озимая пшеница–сахарная свекла–ячмень – наиболее распространенный 4-польный севооборот в Центрально-Черноземном регионе. Каждый год в 2-х полях севооборота вносили солому озимой пшеницы и ячменя. Солому озимой пшеницы и ячменя вносили по схеме, представленной в табл. 1. Опыт был начат в 2010 году, севооборот развернут и в пространстве, и во времени.
Таблица 1.
Схема внесения соломы зерновых культур
| Год отбора почвенных образцов в агроценозе сахарной свеклы | Год внесения соломы | |
|---|---|---|
| Солома озимой пшеницы | Солома ячменя | |
| 2013 | 2012 | 2010 |
| 2014 | 2013 | 2011 |
| 2015 | 2014 | 2012 |
| 2016 | 2015 | 2013 |
| 2017 | 2016 | 2014 |
Общая площадь полевого опыта составила 1210 м2, площадь делянки – 75.6 м2. Повторность опыта четырехкратная. Доза внесения соломы – 4–5 т/га (при запашке соломы после уборки зерновых культур из расчета их средней урожайности). Дополнительные компоненты (целлюлозолитический микромицет, минеральное удобрение и питательную добавку) вносили вручную непосредственно перед вспашкой. В качестве удобрения, содержащего азот, была использована АФК из расчета 40 кг д.в./га. В качестве питательной добавки (ПД) применяли патоку, которая была использована при разведении 1 : 1000. Расход рабочей жидкости – 200 л/га. Целлюлозолитический микромицет вносили на делянки в виде инокулюма (344 тыс. КОЕ/м2), предварительное компостирование проводили согласно методу инфицирования почвы [9, 10].
Схема опыта, варианты: 1 – контроль (без внесения соломы), 2 – солома озимой пшеницы и ячменя (в соответствии с севооборотом), 3 – солома + минеральное удобрение (солома + N), 4 – солома + минеральное удобрение + Humicola fuscoatra ВНИИСС 016 + патока (солома + N + + H. fuscoatra + ПД).
В ранее проведенных исследованиях [11] были учтены все факторы, влияющие на скорость разложения соломы. В опытах, включавших все варианты взаимодействия соломы, целлюдозолитического микромицета и патоки, было установлено, что для наилучшего освоения трудноразлагаемого субстрата целлюлозолитическому микромицету необходимо дополнительное питание, в качестве которого можно использовать патоку, кроме того, без дополнительного азота процесс разложения соломы может затянуться. На основании этих исследований при закладке стационарного многолетнего опыта были выбраны только 4 наиболее контрастных варианта, которые отражали влияние микромицета H. fuscoatra, азота и ПД (патоки) на процесс разложения соломы. При этом были учтены экономические показатели проведения опыта, т.е. при снижении затрат получить максимум информации.
Почвенные образцы отбирали в посевах сахарной свеклы в мае, июле, сентябре с глубины 0‒15 см. В них был проведен учет численности микроорганизмов различных физиологических, таксономических и эколого-трофических групп методом высева почвенной суспензии разной степени разведения на элективные питательные среды. Целлюлозолитические микроорганизмы выделяли на среде Виноградского. Автохтонную группировку определяли на нитритном агаре (НА), зимогенную микрофлору – расчетным методом по сумме аммонификаторов (на среде МПА) и иммобилизаторов азота (на среде КАА), количество диазотрофов – на среде Эшби [12, 13].
В лабораторных условиях в образцах почвы также было определено общее содержание гумуса по методу Тюрина, содержание щелочногидролизуемого азота (Nщг) – по методу Корнфилда [14].
Технология возделывания сахарной свеклы – общепринятая для ЦЧР. Высевали сахарную свеклу гибрида Рамоза. Культуру убирали поделяночно вручную с последующим подсчетом и взвешиванием корнеплодов, их сахаристость определяли на автоматической линии VENEMA [15].
Обработку полученных данных проводили методом дисперсионного анализа и с использованием пакета программ Microsoft Excel.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Влагообеспеченность вегетационного периода какой-либо территории можно охарактеризовать гидротермическим коэффициентом (ГТК) по Селянинову [16]. Наблюдения метеостанции ВНИИСС им. А.Л. Мазлумова за количеством осадков и температурой воздуха показали, что в течение 5-ти лет проведения полевого опыта гидротермические условия менялись в достаточно широких пределах (табл. 2). В 2013 г. ГТК составил 1.6, что характеризовало условия вегетационного периода (с апреля по сентябрь) как влажные. ГТК за вегетационный период 2014 г. составил 0.78. Экстремальными по соотношению влажности и температур в этом году были июль и сентябрь (ГТК 0.05 и 0.08 соответственно). В связи с неблагоприятными погодными условиями была получена самая низкая урожайность озимой пшеницы за последние несколько лет. С 2015 по 2017 гг. ГТК варьировал от 1.33 до 1.83, т.е. в эти годы складывались условия достаточного увлажнения.
Таблица 2.
Гидротермический коэффициент 2013–2017 гг.
| Год наблюдений | ГТК (апрель–сентябрь) | Характеристика условий увлажнения |
|---|---|---|
| 2013 | 1.60 | Влажный |
| 2014 | 0.78 | Очень засушливый |
| 2015 | 1.33 | Обеспеченного увлажнения |
| 2016 | 1.80 | Влажный |
| 2017 | 1.83 | Влажный |
Внесение соломы увеличивает содержание энергетического материала для микроорганизмов, что способствует более активной деятельности почвенной микрофлоры и обогащению ее видового состава. В процессе микробиологического разложения органических удобрений в почве формируется значительное количество самых разнообразных промежуточных продуктов их распада, которые используются в новообразовании гумусовых веществ [17, 18].
В процессе формирования гумусовых соединений принимает участие зимогенная группа микроорганизмов, она поставляет материалы (продукты полураспада органических веществ), из которых синтезируются молекулы гумуса, а также ферменты, катализирующие синтез образующихся молекул [19]. При запашке соломы в почву поступает значительное количество органических веществ, которые служат субстратом для жизнедеятельности зимогенной микрофлоры.
Деструкцию гумуса активизирует автохтонная микрофлора. Представители этой группы способны обеспечить себя минимумом питательных веществ даже в самых неблагоприятных почвенных условиях, в частности, они могут использовать такое стойкое органическое соединение, как гумус [20]. Соотношение численностей этих двух групп микроорганизмов может показать направленность процесса гумификации в сторону синтеза или распада гумусовых веществ.
Результаты исследования показали, что в 2013 г. соотношение зимогенной и автохтонной микрофлоры в почве составило 6.22, внесение соломы и соломы с различными компонентами снизило его до 3.58–4.42 (рис. 1). В 2014 г. складывались экстремальные погодные условия, в результате чего развитие обеих групп микроорганизмов сдерживалось, а их соотношение увеличивалось в ряду: контроль (0.38) – солома (0.35) – солома + N (0.50) – солома + N + Humicola fuscoatra (0.59).

В 2015 и 2016 гг. была зафиксирована однонаправленная динамика, при которой ингибирующий эффект на зимогенную микрофлору оказывало внесение соломы: численность этих микроорганизмов снижалась, а отношение их к автохтонной группе составляло соответственно 3.34 и 3.28. Добавление минерального удобрения к соломе восстановило соотношение зимогенной и автохтонной микрофлоры до уровня контроля. Целлюлозолитический микромицет способствовал усилению гумификационных процессов, а соотношение этих групп микроорганизмов выросло в 2015 и 2016 гг. соответственно до 4.82–4.95 в пользу зимогенной микрофлоры.
В процессе разложения соломы происходит упорядоченный и направленный процесс изменения микробного сообщества, обусловленный специализацией функций микроорганизмов. В первую очередь разлагаются наиболее доступные компоненты соломы, а именно: пентозаны, простые сахара, белки. Вслед за разложением пектиновых веществ начинается интенсивное разложение целлюлозы и близких к ней соединений, появляются типичные целлюлозоразрушающие микроорганизмы [21].
В ходе проведенного исследования было установлено, что их численность в 2013 г. составила 2.82 млн КОЕ/г абсолютно сухой почвы (а.с.п.), использование соломы и соломы с минеральным азотом задерживало рост их количества (рис. 2). Запашка соломы с Humicola fuscoatra способствовала появлению тенденции к повышению численности этой группы микроорганизмов до 3.12 млн КОЕ.
Рис. 2.
Численность целлюлозоразрушающих микроорганизмов в агроценозе сахарной свеклы (НСР05 = 0.84).
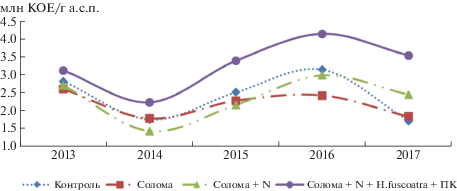
В 2014 г. было отмечено общее снижение численности целлюлозолитических микроорганизмов в почве. В контроле их стало меньше в 1.6 раза, при запашке соломы – в 1.5, соломы и N – в 1.9, соломы и Humicola fuscoatra – в 1.4 раза относительно прошлого года в связи с неблагоприятными засушливыми условиями, сложившимися в этот период.
Использование соломы и соломы с азотом на протяжении 2015 и 2016 гг. несколько снижало количество целлюлозоразрушающих микроорганизмов, которое менялось в пределах ошибки опыта. В 2017 г. при запашке соломы с азотом наметился рост численности этой группы микроорганизмов, однако он не превысил наименьшую существенную разницу. Использование H. fuscoatra в 2013 г. способствовало повышению численности целлюлозолитических микроорганизмов: в 2015, 2016 и 2017 гг. прослежен достоверный рост их количества соответственно на 0.88, 1.00, 1.83 млн КОЕ/г а.с.п.
Увеличение числа целлюлозоразрушающих микроорганизмов, которые требуют дополнительного азота, указывало на то, что микробиологические процессы были направлены на увеличение азотфиксирующей способности почв, которую косвенно выявили по наличию в почве диазотрофов и накоплению щелочногидролизуемого азота.
При деструкции соломы в почве накапливались продукты разложения целлюлозы (низкомолекулярные углеводы), которые являются энергетической основой и источником углерода для жизнедеятельности диазотрофов. Было установлено, что численность диазотрофов во влажном 2013 г. составила 2.05 млн КОЕ/г а.с.п. (рис. 3). Использование с соломой азота снизило этот показатель до 0.99 млн КОЕ/г а.с.п.

В засушливом 2014 г. количество диазотрофов в почве снизилось: в контроле – до 0.80, при запашке соломы – до 0.49, при внесении соломы с азотом – до 0.73 млн КОЕ/г а.с.п. В этих условиях H. fuscoatra способствовал сдерживанию сокращения численности диазотрофов в почве, которая составила 1.20 млн КОЕ/г а.с.п. Это объясняется ускорением разложения соломы и накоплением в почве низкомолекулярных соединений углерода, которые диазотрофы эффективно использовали в осенне-зимний период при достаточном содержании влаги. В 2015 г. (влажном) численность диазотрофов выросла как в контроле, так и после запашки просто соломы и при использовании дополнительных компонентов. В 2016 г., несмотря на оптимальные условия по количеству осадков и суммы эффективных температур, существенных изменений численности диазотрофов не выявлено. Сохранение в 2017 г. температурно-влажностных условий на уровне предыдущего года способствовало росту численности микроорганизмов, фиксирующих азот, при запашке соломы с азотом и H. fuscoatra и с другими дополнительными компонентами.
В целом за 5 лет наблюдений выявлено, что запашка соломы с минеральным азотом и целлюлозолитическим микромицетом (H. fuscoatra) увеличивала численность азотфиксирующих микроорганизмов на 27% относительно контроля и на 30 и 37% относительно вариантов внесения соломы и соломы с азотом соответственно. Увеличение численности диазотрофов непосредственно сказывалось на накоплении доступных для растений форм соединений азота.
В агрономическом отношении большой интерес представляет определение в почве фракции щелочногидролизуемого азота по Корнфилду. Она является наиболее ценной среди форм азота, т.к. по ее запасам можно судить о потенциальном содержании этого элемента в доступном для растений состоянии, что имеет значение для характеристики уровня плодородия. Определение именно этой формы дает возможность учесть часть минерального азота и ближайшие резервы усваиваемых аминосахаров, амидов и моноаминокислот. Если минеральные формы характеризуют текущий запас усваиваемого азота, то щелочногидролизуемый азот дает представление о мобильных органических соединениях, используемых растениями в течение всего вегетационного периода [22, 23].
Было установлено, что уже в 1-й год наблюдений накопление азота происходило при внесении соломы с целлюлозолитическим микромицетом (рис. 4). Негативного влияния внесения соломы выявлено не было: содержание Nщг в почве превышало контроль на 22%.
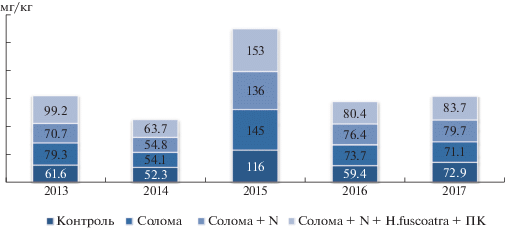
Внесение минерального азота с соломой способствовало достоверному снижению содержания этой формы почвенного азота. Оно было меньше на 8.6 мг/кг, чем при запашке одной соломы. Микромицет H. fuscoatra оказывал положительное влияние на скорость трансформации трудноразлагаемых соединений соломы, в результате чего в почве появились соединения, необходимые для питания растений, в том числе и щелочногидролизуемый азот: его содержание превысило контроль в 1.6 раза, варианты с внесением соломы – в 1.3, соломы + N – в 1.4 раза.
В 2014 г. содержание щелочногидролизуемого азота снизилось до 52.3 мг/кг, однако положительное действие микромицета прослеживалось даже в такой неблагоприятный по климатическим условиям сезон. Содержание его в почве превышало контроль на 11.4 мг/кг, вариант с внесением соломы – на 9.6, соломы и минерального удобрения – на 8.9 мг/кг.
В каждый из последующих 3-х лет такая динамика сохранялась: воздействие целлюлозолитического микромицета способствовало накоплению щелочногидролизуемого азота в почве, содержание которого в 2015, 2016 и 2017 гг. было больше, чем в контроле, соответственно на 24, 26 и 13%.
Почвенная микрофлора осуществляет процесс образования гумусовых веществ, который происходит на ранних стадиях трансформации растительных остатков. Внесение соломы увеличивает содержание энергетического материала для микроорганизмов, что способствует более активной деятельности почвенной микрофлоры и обогащению ее видового состава. В процессе микробиологического разложения органических удобрений в почве формируется значительное количество самых разнообразных промежуточных продуктов их распада, которые используются в новообразовании гумусовых веществ [23, 24].
В соответствии с изменениями в микробиологических процессах трансформации соломы менялось и формирование гумусного состояния почвы. Установлено, что запашка соломы как органического удобрения способствовала накоплению гумусовых веществ в почве на протяжении 5‑ти лет исследования: содержание гумуса увеличилось в 2013 г. на 0.20% (абсолютных), в 2014 г. – на 0.23, в 2015 г. – на 0.05, в 2016 г. – на 0.02, в 2017 г. – на 0.15%, что находилось в пределах наименьшей существенной разницы, а значит, наметилась лишь тенденция к улучшению гумусного состояния почвы (рис. 5).
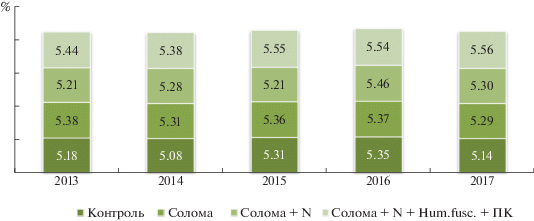
Такая же закономерность прослежена и при запашке соломы с минеральным удобрением. Наилучший эффект получен при дополнительном внесении с соломой целлюлозолитического микромицета H. fuscoatra: превышение содержания гумуса в сравнении с контролем составило в 2013 г. 0.26, в 2014 г. – 0.30 и в 2017 г. – 0.42%. Достаточно сложно проследить изменения в таком стабильном показателе как гумус на данном временнóм этапе, поэтому отмечают только тенденцию к улучшению гумусного состояния почвы.
Высокая активность микрофлоры, вовлекающей в круговорот азот, который затем становится доступным растениям, способствует наилучшему росту и развитию культурных растений, что характеризует интегральный показатель эффективности приема – урожайность (табл. 3).
Таблица 3.
Урожайность и сахаристость корнеплодов сахарной свеклы при запашке соломы зерновых культур
| Вариант | 2013 г. | 2014 г. | 2015 г. | 2016 г. | 2017 г. | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 1 | 2 | 1 | 2 | 1 | 2 | 1 | 2 | |
| Контроль | 39.2 | 16.0 | 23.2 | 18.0 | 24.8 | 18.7 | 29.1 | 16.5 | 30.4 | 18.5 |
| Солома | 39.3 | 16.1 | 25.7 | 17.6 | 27.2 | 18.5 | 26.4 | 16.7 | 29.8 | 18.4 |
| Солома + N | 38.3 | 15.8 | 27.2 | 18.1 | 26.0 | 18.3 | 32.2 | 16.7 | 34.6 | 18.7 |
| Солома + N + H. fuscoatra + ПД | 44.7 | 15.8 | 33.8 | 18.0 | 35.8 | 19.0 | 36.4 | 16.4 | 42.4 | 18.7 |
| НСР05 | 3.2 | 0.4 | 3.7 | 0.2 | 5.7 | 0.8 | 4.7 | 0.7 | 2.3 | 0.2 |
Запашка соломы способствовала либо снижению урожайности (в 2016 и 2017 гг.), либо сохранению ее на уровне контроля. В течение 5-ти лет в соответствии с накоплением щелочногидролизуемого азота и повышением содержания гумуса в почве наблюдали рост урожайности корнеплодов сахарной свеклы при использовании H. fuscoatra и других компонентов: в 2013 г. – на 5.5 т/га, в 2014 г. – на 10.6, в 2015 г. – на 11.0, в 2016 г. – на 7.3, в 2017 г. – на 12.0 т/га.
ЗАКЛЮЧЕНИЕ
Солома зерновых культур, как показало исследование, может быть использована в качестве органического удобрения. При этом введение аборигенного штамма целлюлозолитического микромицета и дополнительных компонентов в систему способствовало ускорению разложения соломы. Это один из способов оптимизации микробного сообщества почвы и повышения ее эффективного плодородия.
В ходе исследования, проведенного в многолетнем полевом опыте, было установлено, что благодаря запашке в почву соломы и дополнительных компонентов активизировалось развитие зимогенной микрофлоры, а также целлюлозолитических микроорганизмов и диазотрофов. В результате в почве накапливались необходимые для питания растений соединения азота. Вследствие улучшения азотного питания была отмечена прибавка биологической урожайности сахарной свеклы в среднем за 5 лет наблюдений: в сравнении с контролем – на 9.6, с использованием одной соломы – на 9.3, соломы с азотным удобрением – на 7.3 т/га.
Список литературы
Щербаков А.П., Рудай И.Д. Плодородие почв, круговорот и баланс питательных веществ. М.: Колос, 1983. 189 с.
Щербаков А.П., Володин В.М. Концепция регулирования почвенного плодородия на биоэнергетической основе // Почвоведение. 1990. № 11. С. 90–103.
Солдатова С.С. Роль сидерации и соломы в формировании экологически устойчивых агробиоценозов в южно-таежной зоне: Автореф. дис. … канд. биол. наук. М., 2011. 20 с.
Зезюков Н.И., Дедов А.В. Роль растительных остатков, соломы и сидератов в формировании плодородия выщелоченного чернозема // Почвы Среднего Поволжья и Урала. Теория и практика их использования и охраны: Тез. докл. XII конф. почвоведов, агрохимиков и земледелов Среднего Поволжья и Урала. 1991. Ч. 1. С. 122–124.
Зезюков Н.И., Острецов В.Е. Сохранение и повышение плодородия черноземов. Воронеж: Центр.-Чернозем. кн. изд-во, 1999. 312 с.
Комаревцева Л.Г., Пчельникова Ф.И. Использование соломы в качестве источника органического вещества почвы // Свойства почв и рациональное использование удобрений. Сб. науч. тр. Перм. СХИ. 1986. С. 120–127.
Мишустин Е.Н. Использование соломы в качестве удобрения // Почвоведение. 1971. № 8. С. 49–54.
Емцев В.Т., Ницэ Л.К. Влияние соломы на микробиологические процессы в почве при ее использовании в качестве органического удобрения // Использование соломы как органического удобрения. М.: Наука, 1980. С. 70–99.
Колесникова М.В., Безлер Н.В., Агапов Б.Л. Формирование плодородия чернозема выщелоченного при интродукции аборигенного штамма целлюлозолитического микромицета и дополнительных компонентов при запашке соломы озимой пшеницы // Агрохимия. 2014. № 8. С. 17–25.
Черепухина И.В., Безлер Н.В. Использование соломы зерновых культур с Humicola fuscoatra ВНИИСС 016 для повышения продуктивности культур зернопаропропашного севооборота // Земледелие. 2018. № 1. С. 35–41.
Колесникова М.В. Продуктивность сахарной свеклы в зависимости от разложения соломы озимой пшеницы под воздействием микромицетов-целлюлозолитиков: Дис. … канд. с.-х. наук. Рамонь, 2009. 152 с.
Звягинцев Д.Г., Асеева И.В., Бабьева И.П., Мирчинк Т.Г. Методы почвенной микробиологии и биохимии. М.: Изд-во МГУ, 1980. 224 с.
Теппер Е.З., Шильникова В.К., Переверзева Г.И. Практикум по микробиологии. М.: Дрофа, 2004. 255 с.
Практикум по агрохимии. Уч. пособ., 2-е изд., перераб. и доп. / Под ред. Минеева В.Г. М.: Изд-во МГУ, 2001. 689 с.
Методика исследований по сахарной свекле. Киев: ВНИС, 1986. 292 с.
Шашко Д.И. Агроклиматические ресурсы СССР. Л.: Гидрометиздат, 1985. 247 с.
Егоров М.А. Подвижное органическое вещество как один из показателей степени ее окультуренности // Зап. Харьков. СХИ. 1938. Т. 1. Вып. 2. С. 3–38.
Верниченко Л.Ю., Мишустин Е.Н. Влияние соломы на почвенные процессы и урожай сельскохозяйственных культур // Использование соломы как органического удобрения. М.: Наука, 1980. С. 3–24.
Шлегель Г. Общая микробиология. М.: Мир, 1987. 567 с.
Лобков В.Т. Биоразнообразие в агроэкосистемах как фактор оптимизации биологической активности почв // Почвоведение. 1999. № 6. С. 732–737.
Адерихин П.Г., Щербаков А.П. Азот в почвах Центрально-Черноземной полосы. Воронеж, 1974. 169 с.
Звягинцев Д.Г. Почва и микроорганизмы. М.: Изд-во МГУ, 1987. 256 с.
Войнова-Райкова Ж., Раиков В., Ампова Г. Микроорганизмы и плодородие. М.: Агропромиздат, 1986. 120 с.
Семенов В.М., Ходжаева А.К. Агроэкологические функции растительных остатков в почве // Агрохимия. 2006. № 7. С. 63–81.
Дополнительные материалы отсутствуют.