

Агрохимия, 2019, № 7, стр. 67-74
УСТОЙЧИВОСТЬ ЯРОВОГО ЯЧМЕНЯ К АБИОТИЧЕСКОМУ СТРЕССУ В ЗАВИСИМОСТИ ОТ УРОВНЯ МИНЕРАЛЬНОГО ПИТАНИЯ И ПРЕДОБРАБОТКИ СЕМЯН СЕЛЕНОМ И КРЕМНИЕМ
Л. В. Осипова 1, *, И. В. Верниченко 2, Л. В. Ромодина 2, Т. Л. Курносова 1, И. А. Быковская 1
1 Всероссийский научно-исследовательский институт агрохимии им. Д.Н. Прянишникова
127550 Москва, ул. Прянишникова, 31а, Россия
2 Российский государственный аграрный университет – МСХА им. К.А. Тимирязева
127550 Москва, ул. Тимирязевская, 49, Россия
* E-mail: legos4@yandex.ru
Поступила в редакцию 09.07.2018
После доработки 21.07.2018
Принята к публикации 10.04.2019
Аннотация
Приведены данные о продуктивности ярового ячменя, содержании фотосинтетических пигментов и уровне свободно-радикального окисления при различной обеспеченности минеральным питанием и предпосевной обработки семян селеном и кремнием в оптимальных условиях культивирования и при действии стресса, индуцированного ограниченным водообеспечением (лабораторные и вегетационные опыты). Показано, что эффективность действия селена и кремния зависела от уровня минерального питания.
ВВЕДЕНИЕ
Глобальные климатические изменения – важнейшая экологическая проблема, связанная с продовольственной безопасностью страны. Во многих климатических сценариях отмечена высокая вероятность негативных последствий изменения климата для производства сельскохозяйственной продукции [1–4]. Увеличение абиотических стрессов, действующих на зерновые культуры, приводит к низкой реализации генетического потенциала продуктивности и снижению урожаев зерна. Поиску средств, смягчающих негативное влияние стрессов, посвящены многочисленные исследования, в которых изучают роль микроэлементов, регуляторов роста, экзогенных гормоноподобных соединений и т.п. [5–10].
Разработка эффективных способов повышения адаптивности в условиях обострения погодных аномалий и комбинированного воздействия неблагоприятных факторов невозможно без изучения механизмов и стратегий приспособления растений к повреждающим воздействиям. В последние десятилетия было показано функционирование неспецифического механизма устойчивости при действии стрессов различной природы. Общей неспецифической реакцией на засуху, засоление, затопление, воздействие высокой и низкой температуры, УФ-радиацию, загрязнение почвы тяжелыми металлами является активация процессов свободнорадикального окисления (СРО) из-за избыточного накопления активных форм кислорода (АФК), что приводит к торможению роста и развития растения и повреждению структур клетки [11–17].
Неспецифическим ответом растений на активизацию СРО является развитие защитно-компенсаторных реакций, включающих синтез ферментов антиоксидантной защиты и низкомолекулярных соединений, обладающих протекторным действием. Биофильные элементы селен и кремний входят в состав антиоксидантной системы, их роль в повышении устойчивости в последние годы установлена многими исследователями [18–25]. Показано их влияние на физиолого-биохимические процессы при стрессовом воздействии, однако эффективность их применения в зависимости от обеспеченности основными элементами минерального питания изучена недостаточно.
В современных условиях хозяйствования при использовании различных технологий (низкозатратной, интенсивной, индустриальной, прогрессивной, зональной, сортовой, экологической, ресурсосберегающей, биологизированной) возникает вопрос об уровне обеспеченности растений основными элементами минерального питания. Роль минерального статуса растений в реализации адаптивного потенциала практически не изучена. Предполагают, что увеличение содержания элементов в растениях в условиях засухи обусловлено, в основном, бóльшим торможением роста, чем поступлением элементов. В наших исследованиях было показано, что низкая обеспеченность минеральным питанием не обеспечивала проявление физиологических защитных механизмов, не возрастала водоудерживающая способность, на низком уровне находилась активность фотосинтеза, не было сбалансировано соотношение фотосинтеза и дыхания, что приводило к низким показателям нетто-ассимиляции и резкому снижению продуктивности [26].
Вопрос о влиянии предпосевной обработки семян (ПОС) биофильными элементами селеном и кремнием изучали на среднеобеспеченных фонах минерального питания, где были установлены физиологические причины протекторного действия этих элементов. В связи с этим цель работы – изучение влияния ПОС на формирование продуктивности ярового ячменя при различной обеспеченности минеральным питанием.
МЕТОДИКА ИССЛЕДОВАНИЯ
Лабораторные и вегетационные опыты (3) проводили с яровым ячменем (Hordeum vulgare L.) сорта Надежный. В 6-ти лабораторных опытах выращивали растения на растворах стрессора в рулонной культуре в термокамере при постоянных температуре и влажности. Осмотический стресс создавали, используя раствор сахарозы.
Семена, обработанные водой, Na2SeO3 и Na2SiO3 · 9H2O, проращивали на воде первые 3-е сут, затем половину рулонов переносили на раствор сахарозы (3 атм.). Через 5, 6, 7 сут определяли линейные размеры проростков и рассчитывали степень депрессии роста по отношению к контролю (Н2О). Оценивали влияние концентраций селена и кремния для совместной обработки семян: 0.001 + 0.015, 0.01 + 0.15, 0.02 + 0.015, 0.02 + + 0.15. На 5-е сут определяли уровень свободно-радикального окисления.
Три вегетационных опыта проводили в почвенной культуре с дерново-подзолистой среднесуглинистой почвой со следующей характеристикой: Nмин – 9 мг/кг, Р2О5 – 93 мг/кг, K2О – 64 мг/кг, Нг – 3,8 мг-экв./100 г почвы; S – 12.0 мг-экв/100 г почвы; рНKCl 4.7. Растения выращивали в сосудах, содержащих 5 кг воздушно-сухой почвы. Перед посевом почву известковали по полной норме Нг и в половину сосудов вносили питательные элементы в виде водных растворов солей (NH4NO3, KH2PO4, NH4Н2РО4) из расчета (NPK)100, во второй половине сосудов растения выращивали без внесения удобрений и известкования. Повторность опыта четырехкратная.
Влажность почвы в вариантах с оптимальным водообеспечением поддерживали на уровне 70% от полной влагоемкости (ПВ). Почвенную засуху создавали путем прекращения полива растений на VI этапе органогенеза, когда происходила закладка цветков в колосе. Растения оставались без полива до достижения почвы влажности устойчивого завядания растений, что соответствовало 14% ПВ. Затем полив растений возобновляли и поддерживали влажность почвы на уровне 70% ПВ до конца вегетации растений.
О влиянии уровня минерального питания и действия биофильных элементов селена и кремния судили по состоянию конуса нарастания главного побега ярового ячменя на VI–VII этапах органогенеза, продуктивности растений и ее структуре.
В течение вегетации оценивали уровень свободнорадикального окисления (СРО). Интенсивность свободнорадикальных процессов определяли по содержанию продуктов перекисного окисления липидов мембран, реагирующих и дающих стойкое окрашивание с тиобарбитуровой кислотой (ТБКпр). Содержание ТБКпр определяли на спектрофотометре “Helios Omega UV-VIS” [27]. Содержание пигментов фотосинтетического аппарата – хлорофиллов а, б и каротиноидов определяли в 100%-ной ацетоновой вытяжке спектрометрическим методом [28].
Поглотительную способность корневой системы и ассимиляцию поступившего азота (15N) определяли методом изотопной индикации. Для определения активности корневой системы растений вместе с поливом вносили небольшое количество (25 мг/сосуд) высокообогащенного (95 АТ%) стабильным изотопом 15N нитратного азота в виде Ca(15NO3)2. Далее через короткие экспозиции отбирали и фиксировали растительные пробы. Образцы анализировали на изотопном масс-спектрометре “Delta V Advantage”, сопряженном с элементным анализатором Flash EA с двухреакторной схемой окисления и восстановления. Метод основан на сжигании растительного материала по методу Дюма в токе кислорода окислительного реактора (Сr2O3 + Co3O4 · AgO) при температуре 1030°С, затем восстановлении в реакторе чистой гранулированной восстановленной медью при 680°С соединений азота до N2, с последующей атомизацией и фракционированием атомов 14N и 15N под действием магнитного поля. Формула для расчета поступившего азота:
Процесс поглощения и ассимиляции нитратных форм азота изучали при коротких экспозициях после внесения $^{{15}}{\text{NO}}_{3}^{ - }$, с этой целью на VI этапе органогенеза ячменя вносили Ca(15NO3)2 с обогащением 95%. Растительные пробы отбирали через 4 и 9 суток. Экспозиция 4 суток соответствовала прямому действию стресса, 9 суток – восстановительному периоду.
Результаты опытов обработаны статистически. В таблицах и рисунках приведены среднеарифметические, стандартное отклонение которых не превышало 5–7% от средних величин.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
В серии лабораторных опытов были подобраны концентрации биофильных элементов для предпосевной обработки семян. Анализ прорастания семян и оценка линейных размеров корней и проростков позволила выявить оптимальные концентрации селена и кремния для их совместного применения. Критерием выбора служила минимальная депрессия роста при действии стресса, индуцированного осмотическим агентом (сахароза 3 атм.) (табл. 1). Оценивая действие совместного применения селена и кремния, можно отметить концентрацию Se0.02 + Si0.15 для предпосевной обработки семян, при которой наблюдали наибольшие размеры проростка и наименьшую депрессию роста при индуцированном стрессе.
Таблица 1.
Влияние ПОС селеном и кремнием на депрессию роста проростка ячменя при осмотическом стрессе (лабораторный опыт)
| Вариант ПОС | Депрессия роста, % | |||||
|---|---|---|---|---|---|---|
| проросток | корень | |||||
| Время, сут | ||||||
| 7 | 6 | 5 | 7 | 6 | 5 | |
| Н2О | 33.4 | 46.5 | 93.1 | 28.0 | 50.5 | 97.4 |
| Se0.001 + Si0.015 | 38.1 | 28.0 | 37.6 | 24.4 | 35.1 | 23.8 |
| Se0.01 + Si0.15 | 30.4 | 45.2 | 69.3 | 32.5 | 56.7 | 30.0 |
| Se0.02 + Si0.015 | 34.1 | 51.3 | 84.4 | 16.6 | 29.8 | 38.6 |
| Se0.02 + Si0.15 | 17.4 | 20.8 | 26.4 | 20.0 | 30.5 | 40.2 |
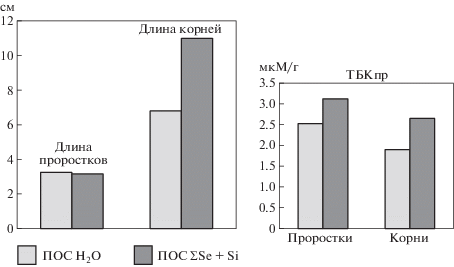
Также в проведенных лабораторных экспериментах было показано, что в оптимальных условиях культивирования предпосевная обработка семян селеном и кремнием приводила к повышению уровня СРО, что свидетельствовало об интенсификации метаболизма, приводящей к активации ростовых процессов (рис. 1).
Рис. 1.
Влияние ПОС селеном и кремнием на линейные размеры проростков ячменя и содержание тиобарбитуровой кислоты (ТБКпр) в благоприятных условиях прорастания.
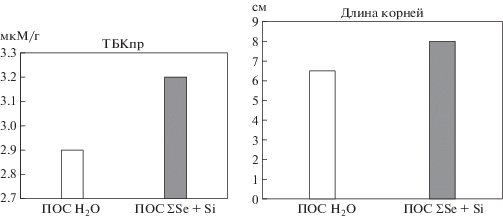
При действии осмотического стресса увеличивалось накопление свободных радикалов в корнях проростков на 15% по сравнению с контролем, возрастало содержание продуктов перекисного окисления липидов (ПОЛ). В варианте с предобработкой семян селеном и кремнием увеличение ТБКпр было выражено в меньшей степени и коррелировало с меньшим торможением роста корней (рис. 2).
Рис. 2.
Длина корней и содержание тиобарбитуровой кислоты (ТБКпр) в корнях ячменя при действии осмотического стресса при ПОС селеном и кремнием.
В исследованиях, проведенных в вегетационном эксперименте, было установлено, что растения ячменя, выращенные без применения удобрений, в течение всего онтогенеза характеризовались низким уровнем свободно-радикального окисления (табл. 2). При оптимизации минерального питания – известковании и внесении основных питательных веществ – уровень СРО был значительно выше.
Таблица 2.
Влияние ПОС и обеспеченности NPK на содержание тиобарбитуровой кислоты (тест на уровень свободнорадикального окисления) (вегетационный опыт)
| Вариант питания | V–VI этап органогенеза | Цветение | Молочная спелость | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Варианты ПОС | ||||||||||||
| Н2О | Se | Si | Se + Si | Н2О | Se | Si | Se + Si | Н2О | Se | Si | Se + Si | |
| Без удобрения | 3.1 | 4.8 | 3.7 | 8.2 | 3.0 | 2.9 | 2.9 | 3.0 | 5.1 | 5.9 | 12.9 | 5.2 |
| NPK | 9.5 | 10.3 | 6.0 | 12.3 | 11.6 | 13.0 | 13.6 | 11.8 | 18.2 | 22.6 | 15.4 | 20.6 |
Согласно современным представлениям, в растительном организме наряду с окислительно-восстановительными реакциями, полностью контролируемыми ферментами, имеет место СРО, которое при низкой интенсивности является нормальным метаболическим процессом.
В свете свободно-радикальной теории АФК рассматривают не только как повреждающие, но и как регуляторные молекулы. Свободно-радикальное окисление необходимо для нормального функционирования растительного организма. Свободные радикалы участвуют в обновлении структур, способствуют уничтожению отживших клеток, участвуют в транспорте ионов и регуляции проницаемости мембран [29–36].
В проведенном эксперименте условия минерального питания определяли интенсивность процессов свободно-радикального окисления. Содержание ТБКпр, по которому судят об активности метаболизма, в вариантах без внесения удобрений было в 3–4 раза меньше, чем на фоне внесения NPK и известкования.
Анализ морфометрических параметров растений на VI этапе органогенеза, ответственном за формирование зачаточного колоса, также показал значительные различия между вариантами питания. Ячмень на оптимальном фоне питания отличался большей высотой растений, большей облиственностью и величиной ассимиляционной поверхности. К концу VI этапа органогенеза в период формирования цветков у них были бóльшие линейные размеры конуса нарастания главного побега и в 3 раза больше заложившихся колосков (табл. 3).
Таблица 3.
Влияние ПОС и обеспеченности NPK на формирование зачаточного колоса ячменя (V–VI этап органогенеза) (вегетационный опыт)
| Вариант питания | Длина конуса нарастания, см | Количество колосков, шт. | ||||||
|---|---|---|---|---|---|---|---|---|
| Варианты ПОС | ||||||||
| Н2О | Se | Si | Se + Si | Н2О | Se | Si | Se + Si | |
| БУ | 2.8 | 2.1 | 2.5 | 2.6 | 8.1 | 12.0 | 9.0 | 8.0 |
| NPK | 6.1 | 6.1 | 6.6 | 6.5 | 20.0 | 20.3 | 20.1 | 21.5 |
Оценивая состояние пигментного комплекса растений ячменя в онтогенезе, можно отметить, что условия минерального питания повлияли на содержание хлорофилла б и каротиноидов. В варианте без внесения удобрений их содержание было на 15–30% меньше, чем на фоне NPK. При незначительных различиях в содержании хлорофилла а, соотношение форм хлорофилла различалось в зависимости от фона питания, что свидетельствовало о разных стратегиях адаптации растений к условиям питания (табл. 4) [37–40].
Таблица 4.
Влияние ПОС на соотношение хлорофилла a : б в оптимальных условиях и при действии стресса в зависимости от уровня обеспеченности NPK (вегетационный опыт)
| Вариант ПОС | Оптимальные условия | Стресс | ||
|---|---|---|---|---|
| a : б | ||||
| Варианты питания | ||||
| Без удобрения | NPK | Без удобрения | NPK | |
| Н2О | 1.25 | 0.61 | 0.77 | 0.51 |
| Se | 1.07 | 0.86 | 0.97 | 0.50 |
| Si | 1.10 | 0.90 | 0.78 | 0.50 |
| ΣSe + Si | 0.79 | 0.53 | 0.64 | 0.45 |
При низкой обеспеченности растений минеральным питанием доля хлорофилла б в сумме хлорофиллов была в 2.0–2.3 раза меньше, чем на оптимальном минеральном фоне. Для фотосинтетических процессов, поддержания роста и формирования продуктивности такого количества хлорофилла б и каротиноидов без опасности фотодеструкции было достаточно.
В вариантах с внесением удобрений требовалась трансформация светособирающего комплекса: увеличение содержания хлорофилла б и каротиноидов для дополнительного поглощения световой энергии, обеспечивающей более активный рост и фотосинтез.
В оптимальных условиях культивирования ПОС селеном и кремнием не оказывала влияния на уровень СРО в онтогенезе. Исключением было повышение содержания ТБКпр на VII этапе органогенеза при совместной обработке семян селеном и кремнием (табл. 2), что не сказалось на параметрах роста растений, закладке колосков и длине конуса нарастания (табл. 3), содержании хлорофиллов.
На недостаточно обеспеченной почве с низкой интенсивностью свободно-радикального окисления возросшая под влиянием ПОС селеном и кремнием на первых этапах органогенеза ростовая активность проростков ячменя ко времени перехода к генеративному развитию завершилась. ПОС не повлияла на реализацию адаптивного потенциала и продуктивность ячменя (табл. 5). На известкованном фоне при внесении рекомендуемых доз NPK ПОС биогенными элементами Se и Si, активируя общие неспецифические системы стрессорного ответа, способствовала запуску механизмов долговременной адаптации, что позволило растениям находиться в адаптированном состоянии. ПОС приводила к проявлению неспецифической защитной реакции, которая выражалась в увеличении пигментного фонда за счет каротиноидов, защищающих фотосинтетический аппарат от оксидативного стресса (рис. 3). Наибольшая эффективность ПОС отмечена при совместном применении селена и кремния, что привело к сохранению заложившихся элементов генеративной сферы и величины продуктивности.
Таблица 5.
Влияние ПОС и обеспеченности NPK на продуктивность ячменя в благоприятных условиях культивирования, г/сосуд
| Варианты ПОС | Н2О | Se | Si | Se + Si | НСР0.5 |
|---|---|---|---|---|---|
| Вариант питания | |||||
| Без удобрения | 4.2 | 4.7 | 3.8 | 4.4 | 0.4 |
| NPK | 11.2 | 13.1 | 13.3 | 13.1 | 0.6 |

В исследовании, проведенном с использованием метода изотопной индикации, получены аналогичные закономерности. Влияние биогенных элементов селена и кремния при совместном и раздельном применении в зависимости от обеспеченности почвы азотом, фосфором и калием изучали с использованием метода меченых атомов. Оценивали поглотительную способность растений ячменя при действии стресса, индуцированного недостатком водообеспечения в критический период формирования генеративной сферы и во время репарации, после прекращения действия стрессового фактора. В период стрессорного воздействия, когда растения подошли к состоянию завядания, поглотительная функция у них была сохранена, хотя и снижена по сравнению с аналогичными поливными вариантами.
Растения ячменя, семена которых были обработаны совместно селеном и кремнием, не только не снижали поглотительную деятельность корней, но и значительно активизировали ее в период действия стресса. Это свидетельствовало о реализации защитной функции у растений этого варианта предобработки семян. В репарационный период после окончания негативного влияния стресса активность поступления азота в растения увеличилась (рис. 4) и в наибольшей степени под влиянием Se и Si.

При действии стресс-фактора предпосевная обработка семян всеми изученными элементами способствовала реализации адаптивного потенциала ячменя. Применение селена и кремния во всех вариантах снижало негативное действие стрессора на поглощение растениями меченого нитратного азота с момента его внесения. Наибольшую активность корневой системы отметили при совместном применении селена и кремния (табл. 4). ПОС ячменя не только оказывала протекторное действие и нивелировала токсическое влияние стрессора, но и активизировала поступление азота в надземные органы растений. Полученные данные согласовались с результатами оценки свободно-радикального окисления и состоянием фонда фотосинтетических пигментов.
ЗАКЛЮЧЕНИЕ
В результате проведенного исследования было установлено, что эффективность предобработки семян ячменя селеном и кремнием зависела от обеспеченности растений элементами минерального питания. На среднекислой среднеобеспеченной фосфором и калием почве и низкой обеспеченности азотом применение селена и кремния не оказывало влияние на продуктивность и устойчивость растений ячменя к стрессу. Семена, прошедшие предпосевную обработку селеном и кремнием, в первый период роста и развития отличались более оптимальным физиологическим статусом: у них были активированы ростовые функции, увеличен уровень свободно-радикального окисления, что свидетельствовало о повышенной активности метаболизма. При низком уровне минерального питания (без внесения питательных веществ) активированные стартовые механизмы не реализовались, и при наступлении стрессовой ситуации протекторное действие селена и кремния не проявлялось. При увеличении уровня минерального питания за счет внесения азота, фосфора и калия активизированные стартовые механизмы отличались пролонгированным действием. При развитии окислительного стресса, вызванного ограничением водообеспечения, ПОС селеном и кремнием способствовала реализации адаптивного потенциала ячменя, снижению потерь продуктивности. Наиболее эффективным это было при совместном применении селена и кремния.
Список литературы
Оценочный доклад об изменениях климата и их последствий на территории Российской Федерации. М.: Росгидромет, 2008. 227 с.
Шульгин И.А. Энергетический баланс и физиологические критерии продуктивности посевов сельскохозяйственных культур в условиях изменения климата // Проблемы экологического мониторинга и моделирования экосистем. 2013. Т. 25. С. 224–250.
Шеламова Н.А. Влияние изменения климата на сельское хозяйство. М.: ЦИ и ТЭИ АПК ВНИИЭСХ, 2013. 81 с.
Jurrel H., Burke J., Faures J.M. Climate change, water and bood security. FAO. Imform. Dir. Electronic. Pube Policy and support brauch. 2011. V. XXI. 174 p.
Костин В.И., Мазурова О.Г., Маркелова Е.С. Использование природных рострегуляторов для регуляции адаптивных реакций озимой пшеницы к неблагоприятным условиям // Агроэкологические проблемы сельскохозяйственных производств. Пенза, 2006. С. 69–71.
Шаяхметов И.Т. Защитно-стимулирующие и адаптивные свойства препарата гуми – биоактивированной формы гуминовых кислот. Эффективность его использования в сельском хозяйстве. Уфа, 2000. 102 с.
Ямалеев А.М., Ямалеева А.А. Антистрессовый эффект гуми на посевах яровой пшеницы // Агрохим. вестн. 2007. № 2. С. 21–22.
Грехова И.В., Матвеева Н.В. Реакция яровой пшеницы на применение регуляторов и микроудобрений при протравливании семян // Аграрн. вестн. Урала. 2014. № 1(119). С. 6–8.
Шаповал О.А., Можарова И.Г., Барчукова А.Я., Коршунов А.Л., Мухина М.Т. Регуляторы роста растений в агротехнологиях основных сельскохозяйственных культур. М.: ВНИИА, 2015. 348 с.
Пахомова В.М., Даминова А.И., Гайсин И.А. Хелатные микроудобрения марки ЖУСС в устойчивости яровой пшеницы к комбинированному стрессу // Агрохим. вестн. 2015. Т. 6. С. 29–31.
Apel K., Hirt H. Reactive oxygen on wheat grouth and some physiological factors // Jnt. I. Forest Soil Erosoin. 2012. V. 2. № 1. P. 50–58.
Apel K. Reactive oxigen species metabolism, oxidative stress and signal transduction // Ann. Rev. Plant Biol. 2004. V. 55. P. 373–399.
Arova A. Oxidative stress and antioxidative system in plants // Current sci. 2002. V. 82. № 10. P. 1227–1238.
Полесская О.Г., Каширина Е.И., Алехина Н.Д. Влияние солевого стресса на антиоксидантную систему растений в зависимости от условий азотного питания // Физиология растений. 2006. Т. 53. № 2. С. 207–214.
Карпец Ю.В., Колупаев Ю.Е. Значение окислительного стресса в индуцировании теплоустойчивости проростков пшеницы кратковременным действием сублетальной температуры // Физиол. и биохим. культ. растений. 2008. Т. 40. № 3. С. 245–258.
Радюкина Н.Л., Шашукова А.Н., Шевякова Н.И., Кузнецов Вл.В. Участие пролина в системе антиоксидантной защиты при действии NaCl и параквата // Физиология растений. 2008. Т. 55. С. 721–730.
Гаридзянов А.Р., Жуков Н.Н., Пантюхин Ю.О., Иванищев В.В. Особенности NaCl-индуцированного окислительного стресса и динамики активности антиоксидантных ферментов в органах тритикале // Докл. РАСХН. 2012. № 2. С. 9–11.
Heather A.C., Carole C.P. Silica in plants: Biological, biochemical and chemical studies // Ann. Bot. 2007. December. № 100(7). P. 1383–1389.
Самсонова Н.Е., Зайцева З.Ф., Капустина М.В. Влияние соединений кремния и сложного NPK-удобрения на водный режим листьев и урожайность яровой пшеницы // Агрохимия. 2014. № 9. С. 58–66.
Давыдова О.Е., Велицкий В.А., Яровский П.П. Физиолого-биохимические и стресспротекторные функции селена в растениях // Физиол. и биохим. культ. раст. 2009. Т. 41. № 2. С. 109–123.
Vao X., Chu J., Wang G. Effect of selenium on wheat seedling under drought stress // Biol. Trace Element. Res. 2009. V. 130. P. 283–290.
Вихрева В.А., Блинохватов А.А., Клейменова Т.В. Селен в жизни растений. Пенза, 2012. 220 с.
Chader H. Effect of drought stress and selenium spraying on photosynthesis and antioxidant activity of spring barley // Acta Agricult. Slovenica. 2013. V. 101. № 1. P. 31–39.
Осипова Л.В., Курносова Т.Л., Быковская И.А. Повышение адаптивного потенциала ячменя ярового (Hordeum vulgare L.) при действии абиотического стресса // Пробл. агрохим. и экол. 2016. № 3. С. 48–51.
Верниченко И.В., Осипова Л.В., Яковлев П.А., Курносова Т.Л., Литвинский В.А. Изучение устойчивости растений и ассимиляция ими нитратного азота при действии абиотического стресса как теоретическая основа метода оценки адаптивного потенциала зерновых культур // Пробл. агрохим. и экол. 2016. № 4. С. 45–50.
Ниловская Н.Т., Осипова Л.В. Приемы управления продукционным процессом яровой пшеницы агрохимическими средствами в условиях засухи. М.: ВНИИА, 2009. 176 с.
Heath R.L., Pasker L. Photoperoxidation in isolated chloroplasts // Arch. Biochem. Biophys. 1968. V. 125. P. 180–198.
Lichtenthaler H.K. Chlorophylls and carotinoids pigments of photosynthetic biomembranes methods in enzymology // Methods in enzymology. 1987. V. 148. P. 350–382.
Waraich E.A., Ahmad R., Saifullah, Ashraf M.Y. Role of mineral nutrition in alleviation of drought stress in plants // Anstral. J. Crop Sci. 2011. T. 5. № 6. P. 764–777.
Allen R.G., Balin A.K. Oxidative influence on development and differentiation an overview of a free radical theory of development // Free Radic. Biol. 1989. V. 6. P. 631–661.
Hitchler M.Y., Domann F.E. An epigenetic perspective on the free radical theory of development // Free Radic. Biol. 2007. V. 43. P. 1023–1036.
Go Y.M., Jones D.P. Redox compartmentalization in eukaryotic cell // Biochim. Biophys. Acta. 2008. V. 1780. P. 1273–1290.
Мартанович Г.Г., Черенкевич С.Н. Окислительно-восстановительные процессы в клетках. Монография. Минск: БГУ, 2008. 159 с.
Jones D.P. Radical-free biology of oxidative stress // Am. J. Physiol. Cell Physiol. 2008. V. 295. P. 849–868.
Jones D.P. Redox theory of aging // Redox Boil. 2015. V. 5. P. 71–79.
Burhans W.C., Heintz N.H. The cell cycle is a redox cycle // Free Radic. Biol. 2009. V. 47. P. 1282–1293.
Tanaka R., Tanaka A. Chlorophyll b is not just an accessory pigment but a regulator of photosynthetic antenna // Porphyrins. 2000. V. 9. № 1. P. 240–245.
Tanaka R., Tanaka A. Chlorophyll cycle regulates the construction and destruction of the light-harvesting complexes // Biochim. Biophys. Acta. 2011. V. 1807. P. 968–976.
Тютерева Е.В., Иванова А.Н., Войцеховская О.В. К вопросу о роли хлорофилла b в онтогенетических адаптациях растений // Усп. совр. биол. 2014. Т. 134. С. 249–256.
Kirst H., Gabilly S.T., Niyogi K.K., Lemaux P.G., Melis A. Photosynthetic antenna engineering to improve crop yields // Planta. 2017. V. 245. P. 1009–1020.
Дополнительные материалы отсутствуют.