

Агрохимия, 2020, № 2, стр. 84-91
ВЛИЯНИЕ ХЛОРИДА НАТРИЯ НА РОСТ ПРОРОСТКОВ
Н. А. Хабиева 1, 2, *, З. М. Алиева 2, К. У. Куркиев 3
1 Прикаспийский институт биологических ресурсов ДНЦ РАН
367000 Махачкала, ул. М. Гаджиева, 45, Республика Дагестан, Россия
2 Дагестанский государственный университет
367000 Махачкала, ул. Гаджиева, 43-а, Республика Дагестан, Россия
3 Дагестанская опытная станция – филиал Федерального исследовательского центра
“Всероссийский институт генетических ресурсов растений им. Н.И. Вавилова”
368612 с. Вавилово, Дербентский р-н, Республика Дагестан, Россия
* E-mail: nadira.xabieva@mail.ru
Поступила в редакцию 18.03.2019
После доработки 15.05.2019
Принята к публикации 10.11.2019
Аннотация
Изучено влияние хлоридного засоления на энергию прорастания и всхожесть семян, ростовые показатели, содержание пролина и ионов натрия у озимого тритикале (×Triticosecale Wittm.) сортообразцов Алмаз, Сотник и ПРАГ530л-1934. Энергию прорастания и всхожесть семян определяли, проращивая их в растворах NaCl (85–350 мМ) и дистиллированной воде (контроль). Ростовые показатели (высоту надземной части, длину и количество корней, сырую и сухую биомассу), содержание пролина и ионов натрия определяли у проростков в вариантах 85 и 165 мМ NaCl. В условиях засоления отмечено подавление роста и накопления биомассы, в большей степени выраженное у корней по сравнению с надземной частью проростков. При оценке ростовых показателей самым чувствительным оказался сортообразец Алмаз, более устойчивыми – сортообразцы Сотник и ПРАГ530л-1934. Чувствительный сортообразец Алмаз характеризовался бóльшим содержанием натрия и менее интенсивной аккумуляцией пролина в условиях засоления.
ВВЕДЕНИЕ
Проблема солеустойчивости растений давно привлекает внимание исследователей. Засоленными, по средним оценкам, являются до 25% возделываемых почв, а в некоторых регионах, в частности, в Дагестане – до 38%. Наблюдается и постоянный рост уровня вторичного засоления, которое развивается при нерациональном орошении [1, 2]. В условиях засоления отмечено изменение комплекса физиолого-биохимических параметров, что приводит к подавлению роста и продуктивности растений. По степени и направленности этих изменений судят об солеустойчивости растений, разрабатывая на их основе различные методы ее диагностики.
Засоление вызывает в растениях осмотический стресс и ионный дисбаланс, связанный с избыточным содержанием ионов. Повышение содержания Na+ в цитоплазме растительных клеток определяет токсические эффекты, а системы транспорта Na+, поддерживающие его низкое содержание в цитоплазме при засолении, играют ключевую роль в устойчивости растений [3]. Одним из критериев оценки солеустойчивости растений служит накопление пролина, уровень которого при стрессе значительно возрастает. Пролин – один из самых типичных осмолитов – наиболее изученный протектор и осморегулятор. Интенсивное накопление в тканях пролина рассматривается как критерий солеустойчивости генотипов или как показатель их адаптивности к стрессовому воздействию [4].
Изучение солеустойчивости особенно актуально для широко возделываемых злаковых культур. Многочисленные работы посвящены изучению солеустойчивости пшеницы [5–8], ячменя [9, 10], ржи [11]. В то же время солеустойчивость такой ценной и перспективной культуры, как тритикале (Triticosecale Wittm.), имеющего высокую продуктивность и устойчивость к стрессам, остается малоизученной. В Дагестане озимое тритикале выращивают в низменных районах, где засоленность почвы достаточно высока, что делает актуальным изучение его солеустойчивости и выявление устойчивых сортообразцов. Однако исследования, посвященные этой проблеме, пока немногочисленны [12–15].
В связи с этим цель работы – изучение сравнительной солеустойчивости проростков сортообразцов озимого тритикале с применением различных морфометрических и физиологических методов оценки.
МЕТОДИКА ИССЛЕДОВАНИЯ
В качестве объекта исследования были выбраны 3 сортообразца озимого тритикале (×Triticosecale Wittm.) – Алмаз, Сотник и ПРАГ530л-1934 из коллекции Дагестанской опытной станции ВИР им. Н.И. Вавилова. Для оценки прорастания семян их проращивали в чашках Петри на фильтровальной бумаге [16], смоченной растворами хлорида натрия и дистиллированной водой (контроль). В связи с ограниченностью исследований солеустойчивости тритикале для оценки сортообразцов проростки культивировали в широком диапазоне концентраций NaCl (85–350 мМ). Объем выборки составлял 50 семян в трехкратной повторности в каждом варианте. Чашки Петри помещали на 7 сут в климатическую камеру (Sanyo MLR–352H, Япония) для моделирования условий: температура 24 ± 1°С, освещение – 3000 лк и влажность 80%. Энергию прорастания (на 3-и сут) и всхожесть (на 7-е сут) определяли согласно ГОСТ 12038–84. Число, длину и массу корней, длину и массу надземной части определяли на 7-е сут в вариантах засоления 85 и 165 мМ NaCl. Сырую и сухую (после фиксации материала при 105°С в течение 3 ч и доведения до постоянного веса) биомассу растительного материала оценивали гравиметрическим методом.
Содержание свободного пролина определяли по методу [17], в модификации Калинкиной с соавт. [18]. Навеску растительного материала массой 100 мг растирали в 5 мл дистиллированной воды. Брали по 5 проб, каждую в 3-х повторностях. Содержимое пробирки кипятили в течение 10 мин и охлаждали, затем центрифугировали 20 мин при 6 тыс. об./мин. К 2 мл надосадочной жидкости добавляли 2 мл нингидринового реактива (1.25 г нингидрина, 20 мл 6 М Н3РО4, 30 мл ледяной уксусной кислоты) и 2 мл ледяной уксусной кислоты. Полученный раствор тщательно перемешивали, инкубировали в течение 1 ч на кипящей водяной бане и охлаждали на ледяной бане. В контрольном образце экстракт заменяли 2 мл дистиллированной воды. Оптическую плотность полученных окрашенных растворов измеряли на спектрофотометре (Beckman Coulter DU–730, США) при длине волны 520 нм. Содержание пролина рассчитывали по калибровочной кривой. Для анализа брали по 5 проростков из 3-х чашек Петри каждого варианта.
Определение содержания ионов Na+ в надземной части и корнях проводили в водной вытяжке [19]. Для определения содержания ионов Na+ надземную часть и корни высушивали при температуре 105°С, измельчали в ступке до порошкообразного состояния и разводили 10 мл дистиллированной воды. Измерения проводили в 3-х повторностях для 4-х проб на атомно-адсорбционном спектрометре (МГА–915 МД, Россия).
Статистическая обработка данных. Обработку данных проводили с помощью программы Microsoft Excel 2007. Для сравнения независимых выборок использовали параметрический t-критерий Стьюдента (р < 0.05). В таблицах и рисунках приведены средние арифметические и их стандартные ошибки.
Исследование выполнены на базе биологического факультета и Центра коллективного пользования “Аналитическая спектроскопия” Дагестанского государственного университета. Часть исследования проведена на Дагестанской опытной станции ВИР им. Н.И. Вавилова в рамках работы по теме НИР 0662–2019–0006.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
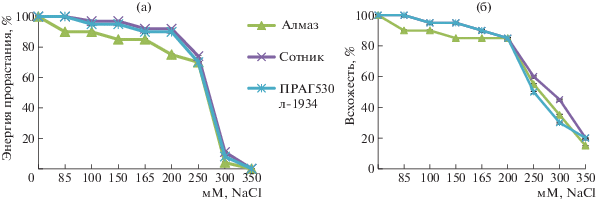
Результаты лабораторных опытов показали, что интенсивность снижения энергии прорастания и всхожести семян в условиях засоления зависела от сортообразца и от концентрации соли. В диапазоне концентраций 85–250 мМ NaCl бóльшую чувствительность проявил сортообразец Алмаз, чем образцы Сотник и ПРАГ530л-1934. Всхожесть и энергия прорастания семян у этого сортообразца при засолении снижались на 10–25%, тогда как у образцов Сотник и ПРАГ530л–134 – на 5–10%. Выраженное снижение энергии прорастания и всхожести семян у всех сортообразцов наблюдали, начиная c концентрации 250 мМ. При проращивании семян тритикале в этом варианте энергия прорастания в среднем составляла 70, всхожесть – 55%. При действии высокой концентрации NaCl (350 мМ) более сильное уменьшение всхожести также отмечено у сортообразца Алмаз (рис. 1).
Рис. 1.
Влияние засоления NaCl на энергию прорастания (а) и всхожесть (б) семян сортообразцов озимого тритикале.
Одним из наиболее чувствительных и интегральных физиологических процессов, отражающих состояние растений в условиях стресса, является их рост [20]. В растворе NaCl с концентрацией 85 мМ выраженных различий между сортообразцами по ростовым показателям не наблюдали. Только у сортообразца Сотник длина корня уменьшалась в меньшей степени. Ростовые показатели под влиянием 165 мМ хлорида натрия в меньшей степени ингибировались у проростков сортообразцов ПРАГ530л-1934 и Сотник: длина корня составила у них 43% по сравнению с контролем, высота надземной части – 46 и 44% соответственно. Наиболее сильное ингибирование роста в этом случае наблюдали у сортообразца Алмаз: длина корня составляла 30, высота надземной части – 32% к контролю (табл. 1).
Таблица 1.
Влияние засоления NaCl на ростовые показатели сортообразцов озимого тритикале
| Вариант | Длина корня, мм | Количество корней, шт. | Длина надземной части, мм |
|---|---|---|---|
| Сортообразец Алмаз | |||
| Контроль | 73.0 ± 0.4 | 5.8 ± 0.1 | 80.0 ± 0.3 |
| 85 мМ NaCl | 47.4 ± 0.3* | 5.7 ± 0.2 | 66.4 ± 0.3* |
| 165 мМ NaCl | 22.0 ± 0.2* | 5.6 ± 0.1 | 25.4 ± 0.2* |
| Сортообразец Сотник | |||
| Контроль | 117 ± 0.4 | 5.8 ± 0.1 | 115 ± 0.3 |
| 85 мМ NaCl | 94.0 ± .3 | 5.5 ± 0.1 | 97.1 ± 0.4 |
| 165 мМ NaCl | 50.0 ± 0.3 | 5.0 ± 0.1 | 50.3 ± 0.2 |
| Сортообразец ПРАГ530л-1934 | |||
| Контроль | 112 ± 0.5 | 6.2 ± 0.1 | 113 ± 0.3 |
| 85 мМ NaCl | 74.3 ± 0.3 | 6.1 ± 0.1 | 95.3 ± 0.4 |
| 165 мМ NaCl | 48.0 ± 0.3 | 6.3 ± 0.1 | 52.0 ± 0.2 |
Примечание: Представлены средние 3-х опытов, каждый вариант которых включал 25 растений, и их стандартные ошибки (при р < 0.05). То же в табл. 2.
Снижение сырой биомассы корней и надземной части в условиях засоления в большей степени было выражено также у образца Алмаз. Например, в варианте 165 мМ NaCl сырая биомасса корней и надземной части у проростков этого сортообразца снижалась в 2.4 и 2.0 раза, Сотник – в 2.4 и 1.8 раза, ПРАГ530л-1934 – в 2.3 и 1.4 раза по сравнению с контролем (табл. 2). По такому показателю, как накопление сухой биомассы в условиях засоления, преимущество имел сортообразец Сотник: в варианте 85 мМ NaCl накопление биомассы надземной части снизилось у него на 5%, в варианте 165 мМ NaCl – на 24% по сравнению с контролем, тогда как снижение этого показателя у сортообразца Алмаз составило 26 и 41%, у ПРАГ530л-1934 – 25 и 42% соответственно. В варианте засоления 85 мМ NaCl биомасса корня у сортообразца ПРАГ530л-1934 снижалась в меньшей степени, чем у образца Сотник.
Таблица 2.
Влияние засоления NaCl на сырую и сухую биомассу корней и надземной части проростков озимого тритикале
| Вариант | Сырая биомасса | Сухая биомасса | ||||
|---|---|---|---|---|---|---|
| надземная часть | корень | надземная часть | корень | |||
| мг | мг | % к контролю | мг | % к контролю | ||
| Сортообразец Алмаз | ||||||
| Контроль | 66.1 ± 3.0 | 73.1 ± 3.4 | 6.1 | 100 | 7.9 | 100 |
| 85 мМ NaCl | 53.2 ± 3.0 | 45.4 ± 2.0 | 4.5 | 74 | 4.8 | 61 |
| 165 мМ NaCl | 32.4 ± 2.0 | 30.0 ± 2.0 | 3.6 | 59 | 3.3 | 42 |
| Сортообразец Сотник | ||||||
| Контроль | 86.3 ± 4.4 | 108 ± 3.3 | 5.9 | 100 | 12.4 | 100 |
| 85 мМ NaCl | 72.3 ± 4.3 | 78.1 ± 3.4 | 5.6 | 95 | 8.5 | 69 |
| 165 мМ NaCl | 46.3 ± 2.1 | 45.0 ± 2.5 | 4.5 | 76 | 6.3 | 51 |
| Сортообразец ПРАГ530л-1934 | ||||||
| Контроль | 75.0 ± 4.8 | 98.1 ± 5.3 | 7.7 | 100 | 12.0 | 100 |
| 85 мМ NaCl | 65.2 ± 3.8 | 83.2 ± 4.3 | 5.8 | 75 | 10.3 | 86 |
| 165 мМ NaCl | 54.0 ± 3.5 | 42.0 ± 2.7 | 4.5 | 58 | 5.0 | 42 |
Следует отметить, что у всех изученных сортообразцов корневая система проявила большую чувствительность к засолению, чем надземная часть проростков: рост и накопление биомассы корней в условиях всех вариантов засоления снижались в большей степени (табл. 1, 2). При этом достоверных различий в количестве корней выявлено не было.
В литературе встречаются многочисленные сведения о более интенсивном накоплении пролина как в устойчивых, так и в чувствительных сортах [4, 9, 21]. В наших опытах в условиях низкого уровня засоления (85 мМ NaCl) содержание свободного пролина в большей степени повышалось в растениях сортообразцов ПРАГ530л-1934 (с 1.5 до 9.7 мкмоль/г) и Сотник (с 0.8 до 3.1 мкмоль/г, или в 3.8 раза), в меньшей степени – в растениях сортообразца Алмаз (с 0.8 до 2.3 мкмоль/г, или в 2.8 раза) (табл. 3). При концентрации 165 мМ NaCl абсолютные величины и интенсивность накопления пролина были наиболее высокими в сортообразце ПРАГ530л-1934 (с 1.5 до 21.8 мкмоль/г). Таким образом, более высокая пролинобразующая способность листьев в условиях как более слабого, так и сильного уровня засоления была больше у образцов ПРАГ530л-1934 и Сотник. Эти сортообразцы проявили бóльшую устойчивость при оценке роста и накопления биомассы проростков. Полученные данные согласуются с литературными сведениями о том, что в листьях более чувствительных к стрессу сортов содержание пролина повышается в меньшей степени по сравнению с устойчивыми [9].
Таблица 3.
Влияние засоления NaCl на содержание свободного пролина в листьях озимого тритикале
| Вариант | Сортообразцы | |||||
|---|---|---|---|---|---|---|
| Алмаз | Сотник | ПРАГ530л-1934 | ||||
| мкмоль/г | % к контролю | мкмоль/г | % к контролю | мкмоль/г | % к контролю | |
| Контроль | 0.8 ± 0.0 | 100 | 0.8 ± 0.0 | 100 | 1.5 ± 0.0 | 100 |
| 85 мМ NaCl | 2.3 ± 0.0 | 280 | 3.1 ± 0.1 | 390 | 9.7 ± 0.0 | 647 |
| 165 мМ NaCl | 11.0 ± 0.3 | 1100 | 15.2 ± 0.1 | 1900 | 21.8 ± 2.3 | 1450 |
Известно, что процессы поступления ионов Na+ в растение и его распределение в клетках и тканях строго контролируются на разных уровнях организации – от клеточного до организменного [3, 22, 23]. Низкая концентрация Na+ в цитоплазме и компартментация его в вакуоли поддерживается различными транспортными белками плазмалеммы и тонопласта [24, 25]. У высших растений клеточные механизмы дополняются механизмами дальнего транспорта. Большинство гликофитов стремится ограничить накопление Na+ в метаболически активных надземных органах путем торможения его транспорта из корней в побеги, накопления в старых листьях и стеблях, рециркуляции из побега в корни, компартментации избытка ионов в вакуолях и в свободном пространстве клеток корня или стебля [22, 24, 26]. В целом более высокое накопление ионов натрия характерно для чувствительных сортов [26, 27]. Устойчивые сорта часто характеризуются высокой эффективностью барьерной функцией корней и поддерживают относительно низкий уровень натрия в надземной части [26, 28–30]. Однако такая картина отмечена не для всех видов и сортов [31, 32].
В наших опытах в условиях засоления содержание ионов натрия значительно повышалось в проростках всех сортообразцов уже в варианте 85 мМ NaCl (табл. 4). В наибольшей степени его содержание при засолении возрастало как в корнях, так и в надземной части у чувствительного сортообразца Алмаз. У проростков этого образца в варианте 85 мМ NaCl в надземной части содержание NaCl составило 17.5, в корнях – 49.0 мг/кг. Проростки образцов Сотник и ПРАГ530л-1934 характеризовались более низким и почти одинаковым содержанием Na+ и в надземной части (6.0 и 6.3 мг/кг), и в корнях (29 и 26 мг/кг).
Таблица 4.
Влияние засоления NaCl на содержание ионов натрия в проростках сортообразцов тритикале
| Образец | Вариант | Корни | Надземная часть | Соотношение содержания Na+ корни/надземная часть | ||
|---|---|---|---|---|---|---|
| мг/кг | % к контролю | мг/кг | % к контролю | |||
| Алмаз | Контроль | 0.9 ± 0.08 | 100 | 0.2 ± 0.01 | 100 | 4.5 |
| 85 мМ NaCl | 49.5 ± 0.4 | 5400 | 17.5 ± 0.9 | 7950 | 2.8 | |
| 165 мМ NaCl | 61.0 ± 1.2 | 6600 | 19.0 ± 0.1 | 8600 | 3.2 | |
| Сотник | Контроль | 0.6 ± 0.08 | 100 | 0.09 ± 0.005 | 100 | 6.7 |
| 85 мМ NaCl | 29.0 ± 1.1 | 4750 | 6.0 ± 0.7 | 6700 | 4.8 | |
| 165 мМ NaCl | 34.5 ± 1.5 | 5600 | 6.6 ± 0.2 | 7300 | 5.2 | |
| ПРАГ530л-1934 | Контроль | 0.59 ± 0.2 | 100 | 0.11 ± 0.1 | 100 | 5.4 |
| 85 мМ NaCl | 26.0 ± 1.0 | 4400 | 6.3 ± 0.9 | 5700 | 4.1 | |
| 165 мМ NaCl | 29.0 ± 4.0 | 4900 | 7.9 ± 0.6 | 7200 | 3.6 | |
Увеличение концентрации раствора соли существенно не влияло на дальнейшее повышение содержания ионов Na+ в тканях надземной части проростков у всех сортообразцов – его уровень в варианте 165 мМ возрастал по сравнению с вариантом 85 мМ на 0.6–1.6 мг/кг. Повышение интенсивности засоления в большей степени влияло на содержание ионов Na+ в корнях. В этом случае проявилась и сортовая специфика: содержание ионов Na+ в сортообразце ПРАГ530л-1934 возрастало на 7.0, Сотник – на 5.5 мг/кг, максимальным было в чувствительном сортообразце Алмаз – на 11.5 мг/кг по сравнению с вариантом слабого засоления.
Большее содержание ионов Na+ во всех сортообразцах при засолении наблюдали в корнях, меньшее – в надземной части. При этом наиболее интенсивным его накоплением как в условиях слабого, так и сильного засоления, характеризовался чувствительный сортообразец Алмаз. У него в большей степени возрастала и интенсивность накопления Na+ по сравнению с контролем. Например, в надземной части этот показатель вырос у сортообразца Алмаз в варианте 85 мМ NaCl в 79, в корнях – в 53 раза. Наименьшее накопление было отмечено у образца ПРАГ530л-1934, у которого показатель возрастал в 57 раз в надземной части, в 44 раза – в корнях.
Различия в накоплении натрия между сортообразцами были более выражены в надземной части проростков по сравнению с корнями: в варианте засоления 85 мМ NaCl у чувствительного сортообразца Алмаз в надземной части проростков содержание натрия (17.5 мг/кг) превышало показатели устойчивых образцов ПРАГ530л-1934 (6.0 мг/кг) и Сотник (6.3 мг/кг) в 2.8 и 2.9 раза, в корнях – соответственно в 1.7 и 1.9 раза. Подобную закономерность наблюдали и при более сильном засолении.
Варьировало также у сортообразцов соотношение содержания Na+ в корнях к содержанию Na+ в надземной части. Например, в условиях слабого засоления (85 мМ NaCl) в корнях проростков сортообразца Сотник натрия накапливалось в 5 раз больше, чем в надземной части (29 и 6 мг/кг соответственно), у образца ПРАГ530л-1934 – в 4.1 раза (26 и 6.3 мг/кг), у образца Алмаз – в 2.8 раза (49.5 и 17.5 мг/кг соответственно). В условиях более интенсивного засоления (165 мМ NaCl) наиболее высоким соотношением содержания натрия в корнях к содержанию в надземной части также характеризовался сортообразец Сотник, у которого оно возрастало в 5.2 раза, меньшим – Алмаз (3.2 раза). Таким образом, независимо от уровня засоления чувствительный сортообразец Алмаз характеризовался меньшим соотношением накопления натрия в корнях по сравнению с надземной частью. Это согласуется с литературными данными [26] и может свидетельствовать о меньшей эффективности барьерной функции корней, связанной, в частности, с возможностью локализации засоляющих ионов в клеточных компартментах клеток корня [24].
Таким образом, сортообразцы тритикале Сотник и ПРАГ530л-1934 отличались меньшим накоплением натрия в тканях надземной части по сравнению с чувствительным сортообразцом Алмаз, а также бóльшим соотношением содержания натрия в корнях к его содержанию в надземной части проростков. Наиболее интенсивное повышение содержания натрия, как и пролина, наблюдали у более чувствительного сортообразца Алмаз, характеризующегося более выраженным подавлением роста и накоплением биомассы проростков, менее – у сортообразцов Сотник и ПРАГ530л-1934.
ЗАКЛЮЧЕНИЕ
Проведенное исследование показало, что засоление, вызванное действием растворов NaCl, в большей степени подавляло рост и накопление биомассы надземной части и корней сортообразца тритикале Алмаз, в меньшей степени – сортообразцов Сотник и ПРАГ530л-1934. Рост и накопление биомассы корней в условиях засоления снижались в большей степени, чем надземной части проростков. По накоплению свободного пролина в условиях засоления выделились сортообразцы ПРАГ530л-1934 и Сотник, меньше всего его содержание возрастало в проростках образца Алмаз. И у более устойчивых (Сотник, ПРАГ530л-1934) и у чувствительного (Алмаз) сортообразцов отмечено более интенсивное накопление натрия в корнях по сравнению с надземной частью проростков. Сортовая специфика была выражена в изменении общего содержания натрия в проростках при разных уровнях засоления: максимальным оно было у чувствительного сортообразца. Также она проявилась в различиях величины соотношения содержания Na+ в корнях к его содержанию в надземной части: в большей степени этот показатель увеличивался у устойчивых сортообразцов тритикале. Наиболее интенсивное повышение содержания натрия как в корнях, так и в надземной части наблюдали у более чувствительного сортообразца Алмаз, характеризующегося более выраженным подавлением роста и накопления биомассы проростков, меньшим – у сортообразцов ПРАГ530л-1934 и Сотник, проявивших бóльшую солеустойчивость по морфометрическим показателям.
Список литературы
Строгонов Б.П. Физиологические основы солеустойчивости растений. М.: АН СССР, 1962. 366 с.
Ковда В.А. Проблемы опустынивания и засоления почв аридных регионов мира. М.: Наука, 2008. 415 с.
Балнокин Ю.В. Ионный гомеостаз и солеустойчивость растений. 70-е Тимирязевское чтение. М.: Наука, 2012. 102 с.
Кузнецов Вл.В., Шевыкова Н.И. Пролин при стрессе: биологическая роль, метаболизм, регуляция // Физиология растений. 1999. Т. 46. № 2. С. 321–336.
Ахиярова Г.Р., Сабиржанова И.Б., Веселов Д.С., Фрике В. Участие гормонов в возобновлении роста побегов пшеницы при кратковременном засолении NаCl // Физиология растений. 2005. Т. 52. № 6. С. 891–896.
Шихмурадов А.З. Биоресурсный потенциал и эколого-генетические аспекты устойчивости представителей рода Triticum L. к солевому стрессу: Автореф. дис. … д-ра биол. наук. Владикавказ: Горский ГАУ, 2014. 37 с.
Луценко Э.К., Галактионова М.В. Влияние солевого стресса на некоторые цитофизиологические и биохимические показатели проростков пшеницы // Сел.-хоз. науки и агропром. комплекс на рубеже веков. 2013. № 1. С. 17–20.
Хусаинов А.Т., Сыздыкова Г.Т., Андреева Ю.А. Влияние степени сульфатного засоления на ростовые процессы генотипов яровой мягкой пшеницы // Аграрн. вестн. Урала. 2014. № 1 (119). С. 23–26.
Терлецкая Н.В. Неспецифические реакции зерновых злаков на абиотические стрессы in vivo и in vitro. Алматы: Ин-т биол. и биотехнол. растений КН МОН РК, 2012. 208 с.
Веселов Д.С., Шарипова Г.В., Кудоярова Г.Р. Сравнительное изучение реакции растений ячменя (Hordeum vulgaris) и пшеницы (Triticum durum) на кратковременное и длительное действие натрий-хлоридного засоления // Агрохимия. 2007. № 7. С. 41–48.
Напреенко Н.В. Влияние засоления на рост и развитие зерновых культур // Мат-лы XI Международ. научн. конф. “Почва, урожай и экология”. Горки, 2010. С. 139–141.
Жуков Н.Н. Исследование физиолого-биохимических механизмов солевого стресса у тритикале на ранних этапах онтогенеза: Автореф. дис. … канд. биол. наук. Пущино: Ин-т фундамент. пробл. биол. РАН, 2013. 22 с.
Хабиева Н.А., Куркиев К.У. Морфо-физиологические изменения у проростков озимого тритикале (Triticosecale) в условиях хлоридного засоления // Усп. совр. науки. 2017. Т. 2. № 10. С. 144–148.
Хабиева Н.А. Рост и интенсивность перекисного окисления липидов у проростков озимого тритикале (Triticosecale) при засолении среды // Вестн. Дагестан. гос. ун-та. Сер. 1. Естеств. науки. 2016. Том 31. Вып. 1. С. 114–118.
Куркиев К.У., Алиева З.М., Темирбекова С.К., Хабиева Н.А. Устойчивость мягкой пшеницы и тритикале к высокому уровню хлоридного засоления // Достиж. науки и техн. АПК. 2017. Т. 31. № 2. С. 26–28.
Удовенко Г.В. Диагностика устойчивости растений к стрессовым воздействиям. Метод. рук-во. Л.: ВИР, 1988. 227 с.
Bates L.S., Waldran R.P., Teare I.D. Rapid determination of free proline for water stress studies // Plant Soil. 1973. № 39. P. 205–208.
Калинкина Л.Г., Назаренко Л.В., Гордеева Е.Е. Модифицированный метод выделения свободных аминокислот для определения на аминокислотном анализаторе // Физиология растений. 1990. Т. 37. № 3. С. 617–621.
Халилова Л.А. Пути транспорта Cl– в системе целого растения у галофита Suaeda altissima (L.) Pall.: Автореф. дис. … канд. биол. наук. М.: ИФР РАН, 2008. 25 с.
Шевелуха В.С. Рост как показатель адаптивных возможностей растений и посевов и использование его характеристик в селекции и растениеводстве // Регуляция адаптивных реакций сельскохозяйственных растений. Кишинев: Штинница, 1987. С. 3–11.
Dashek W.V., Erickson S., Sharon S. Isolation, assay, biosynthesis, translocation, and function of proline in plant cells and tissues // Botan. Rev. 1981. V. 47. № 3. P. 349–381.
Tester M., Davenport R. Na+-Tolerance and Na+-transport in higher plants // Annal. Bot. 2003. № 91. P. 503–527.
Керимов Ф.А., Кузнецов Вл.В., Шамина З.Б. Организменный и клеточный уровни солеустойчивости двух сортов хлопчатника (133, ИНЭБР–85) // Физиология растений. 1993. Т. 40. № 1. С. 128–131.
Кривошеева А.Б. Получение и анализ солеустойчивости трансгенных растений арабидопсиса и картофеля, экспрессирующих гетерологические гены вакуолярных антипортеров HvNHX2 или HvNHX3: Автореф. дис. … канд. биол. наук. М.: ИФР РАН, 2015. 22 с.
Rodríguez-Rosales M.P., Galvez F.J., Huertas R., Aranda M.N., Baghour M., Cagnac O., Venema K. Plant NIIX cation/proton antiporters // Plant Signal. Behavior. 2009. V. 4. P. 265–276. https://doi.org/10.4161/psb.4.4
Омельченко А.В., Кабузенко С.Н., Белоусов А.А., Сериков В.А. Локализация натрия в компартментах корней и наземной части гибридов кукурузы нового поколения в сязи с их солеустойчивостью // Уч. зап. Таврического нац. ун-та им. В.И. Вернадского. Сер. биология, химия. 2009. Т. 22 (61). № 4. С. 112–121.
Ибрагимова У.Ф., Азизов И.В., Мамедова М.Г. Реакция сортов твердой и мягкой пшеницы на хлоридное засоление // Физиол. Раст. и генетика. 2013. Т. 45. № 5. С. 399–407.
Мохамед А.М., Ралдугина Г.Н., Холодова В.П., Кузнецов Вл.В. Аккумуляция осмолитов растениями различных 338 генотипов рапса при хлоридном засолении // Физиология растений. 2006. Т. 53. № 5. С. 732–738.
An Ping, Shinobu Inanaga, Jun Li Xiang, Engrinya Eneji A., Wen Nan Zhu Interactive effects of salinity and air humidity in two tomato cultivars differing in salt tolerance // J. Plant Nutr. 2005. V. 28. № 3. P. 459–473.
Yasar F., Uzal O., Tufenkci S., Yildiz K. Ion accumulation in different organs of green bean genotypes grown under salt stress // Plant Soil Environ. 2006. V. 52. P. 476–480.
Кабузенко С.Н., Омельченко А.В., Михальская Л.Н., Швартау В.В. Накопление и локализация ионов натрия и калия в растениях кукурузы в условиях почвенного засоления // Вестн. Днепропетровск. ун-та. Биол., экол. 2013. № 21 (1). С. 28–32.
Cuin T.A., Betts S.A., Chalmandrier R., Shabala S. A root’s ability to retain K+ correlates with salt tolerance in wheat // J. Exp. Bot. 2008. V. 59 (10). P. 2697–2706. https://www.ncbi.nlm.nih.gov/pmc/articles/ PMC2486465/
Дополнительные материалы отсутствуют.