

Агрохимия, 2020, № 9, стр. 48-64
Потоки диоксида углерода при разном режиме использования сенокоса на осушенной торфяной почве
А. Г. Молчанов 1, Г. Г. Суворов 1, Д. В. Ильясов 1, А. А. Сирин 1, *
1 Институт лесоведения РАН
143030 Московская обл., Одинцовский р-н, с. Успенское, ул. Советская, 21, Россия
* E-mail: sirin@ilan.ras.ru
Поступила в редакцию 30.12.2019
После доработки 10.06.2020
Принята к публикации 10.06.2020
Аннотация
Проведено сравнение потоков CO2, измеренных в 2018 и 2019 гг., на сенокосе в разное время после кошения и залежи на осушенной торфяной почве в Московской обл. Сразу после кошения в 2019 г. дыхание экосистемы (Reco) и валовый обмен CO2 экосистемы (GEE) на сенокосе снизились в 2 раза и составили 26 и –18 г CO2/м2/сут соответственно, а удельный GEE (на надземную фитомассу) вырос в 2 раза до 0.56 г CO2 /м2/сут/граст. На залежи величины Reco и GEE в 2019 г. составили 50 и –45 г CO2/м2/сут. В 2018 г., через месяц после кошения показатели Reco и GEE на сенокосе (69 и –32 г CO2/м2/сут) и залежи (66 и –34) практически не отличались, хотя фитомасса была различной: надземная – 138 и 368, подземная – 1223 и 600 г/м2. Через месяц после кошения в 2018 г. нетто-экосистемный обмен СО2 (NEE) на сенокосе и залежи не отличался и составил 38 и 39 г CO2 г/м2/сут. С учетом изымаемой при кошении фитомассы общий за сезон NEE на сенокосе смещался в сторону ассимиляции СО2 в сравнении с залежью.
ВВЕДЕНИЕ
Известно, что из 5 млн га земель, осушенных для сельского хозяйства в Российской Федерации, большая часть исходно имела торфяные почвы [1], а значительную долю торфяников, частично выработанных при добыче торфа, также рекультивировали для этих целей [1–3]. В результате под нужды сельского хозяйства в стране была осушена значительная площадь торфяных болот [4], которые использовали под пропашные культуры, сенокосы и пастбища [1, 2, 5]. В последние десятилетия многие из них, вне зависимости от категории пользователей, были заброшены, стали объектами торфяных пожаров [6], подвергаются ветровой и водной эрозии. Они являются источником парниковых газов и теряют запасы углерода [7–9], не принося экономической пользы.
В настоящее время происходит возвращение части осушенных земель в сельскохозяйственный оборот, что особенно актуально в связи с обеспечением кормовой базы животноводства. Это поднимает вопрос об эмиссии парниковых газов этой категории земель. Осушенные торфяники – значительный источник диоксида углерода в атмосферу [10, 11] и объект учета, согласно Рамочной конвенции ООН об изменении климата [12].
С точки зрения сохранения физико-химических свойств торфяной залежи наиболее щадящим ее использованием является луговодство [11, 13, 14]. Наименьшие потери углерода таких почв характерны при лугово-полевом севообороте и многолетней травяной растительности [15], а при регулировании водного режима возможна их высокая продуктивность для производства кормов [14]. Залужение может препятствовать деградации осушенных торфяных почв [16], а их пастбищное и сенокосное использование в сравнении с распашкой – снижать потери углерода в 3–4 раза, азота – в 2 раза [11].
Отечественными исследованиями проблеме эмиссии парниковых газов из осушенных торфяных почв уделяется существенно меньше внимания, как по сравнению с естественными болотами, так и минеральными сельскохозяйственными почвами [17–22]. Осушенные торфяные почвы в России и странах ближнего зарубежья изучены меньше и в контексте углеродного баланса [7–9, 14, 15, 23–27], чем минеральные или неосушенные торфяные, хотя содержат значительный запас углерода, более уязвимый, чем в естественных болотах. Цель работы – сравнение потоков CO2 сенокоса на осушенной торфяной почве в Московской обл. в условиях залежи и в разное время после сенокошения.
МЕТОДИКА ИССЛЕДОВАНИЯ
Исследование проводили на Дубненском болотном массиве в Талдомском р-не Московской обл. (56°39.4′–56°43.6′ с.ш., 37°42.8′–37°54.6′ в.д., протяженность 8 км с севера на юг и 12 км с запада на восток). Из общей площади торфяника ≈8200 га частично осушено ≈2850 га для разных видов использования: добычи торфа, сельского и лесного хозяйства. Сохранились разнообразные естественные болотные биогеоценозы – от верховых кустарничково-сфагновых сосняков до низинных черноольшаников. Осушение проводили в конце 1970-х годов для добычи торфа фрезерным способом. Расстояние между картовыми каналами было преимущественно 40 м, а после выработки залежи на глубину 1–2 м многие площади были рекультивированы под пропашные культуры, однако в большей части залужены для создания сенокосов [7, 8]. Торфяные пожары 2010 г., которые охватили торфяники области [28], мало коснулись массива, но часть его была включена в программу обводнения пожароопасных торфяников Московской области 2010–2013 гг. с созданием переливных и водорегулирующих плотин, противопожарного водоема и др. [6, 29].
Объекты исследования расположены в южной части массива (56°40.6′ с.ш., 37°48.8′ в.д.), где вторичное обводнение не проводили, а осушенные площади все активнее используют как сенокосы. По имеющимся данным, торфяная залежь была выработана здесь на глубину, вероятно, 0.5–1.0 м; современная глубина осушительных каналов составляет 0.8–1.5 м, а края каналов заросли древесно-кустарниковой растительностью высотой 2–12 м.
Работы проводили на двух соседних картах шириной 50 и 110 м, из которых южную косили каждый год, а северная с 2015 г. и до конца лета 2019 г. была залежной и зарастала рудеральной растительностью, преимущественно крапивой (Urtica dioica L.), вытесняющей основную луговую культуру – мятлик (Poa pratensis L.). Для анализа распределения растительности использовали актуальные спектрозональные спутниковые данные и съемку с БПЛА в видимом диапазоне.
Через оба участка был проведен нивелировочный профиль. Для измерения уровней почвенно-грунтовых вод (УПГВ) в центре участков были установлены перфорированные на всю глубину (100 см) трубки из ПВХ ∅50 мм [30], на залежи – дополнительно автоматический датчик Mini Diver (Eijkelkamp, Нидерланды) с записью УПГВ и температуры воды с частотой 4 ч с охватом вегетационных сезонов 2018 и 2019 гг. Для характеристики метеоусловий периодов измерений были использованы данные сайта www.rp5.ru (ООО “Расписание Погоды”, Санкт-Петербург, Россия) для метеостанции г. Дмитров.
На обоих участках были заложены разрезы до глубины 50 см, из которых с шагом 5 см взяты образцы торфа на влажность, зольность, объемный вес, ботанический состав и элементный состав (CNS-анализ). Влажность почвы определяли термовесовым методом, плотность сложения – в абсолютно сухих образцах 5 × 5 × 5 см, зольность – озолением при температуре 850°C, ботанический состав и степень разложения торфа – микроскопическим методом и центрифугированием, содержание С, N и S – на элементном анализаторе Vario MICRO cube (Elementar, Германия) при сгорании образца при t = 1140°С, разделении газов с помощью метода программируемой ступенчатой десорбции (TPD – Temperature Programmed Desorption) в двукратной повторности.
Интенсивность разложения органического вещества в почве оценивали весовым методом по убыли массы хлопчатобумажной ткани [31], экспонированной в почве с 27.07.2019 по 01.09.2019. Для этого к стенке прикопок (по 2 на участке сенокоса и залежи) были прижаты стеклянные пластины, с закрепленными при помощи полимерной сетки тремя отрезками хлопчатобумажной ткани высотой 10 и шириной 15 см так, что они были экспонированы на глубинах 0–10, 10–20 и 20–30 см. Ткань предварительно прокипятили, высушили до воздушно-сухого состояния и взвесили на аналитических весах. По завершении эксперимента образцы ткани повторно промыли, высушили и взвесили.
Фитомассу растений (с пересчетом на сухое вещество после сушки при температуре 105°C) определяли в двукратной повторности: надземную (НФМ) методом “укосных квадратов” (площадь рамки 0.49 м2), подземную (ПФМ) промыванием корней почвенных образцов (10 × 10 × 20 см) на сите с размером ячейки 2 мм для верхнего почвенного горизонта 0–10 см [32]. Индекс листовой поверхности (Leaf area index – LAI) рассчитывали как отношение площади листьев к площади их произрастания. Площадь листьев определяли по палетке в выбранной навеске, после чего пересчитывали на общую массу укоса.
Потоки диоксида углерода измеряли в близкие даты 2018 и 2019 гг. – 27–28.08 и 28–29.08 соответственно. В эти же дни 1 раз в час автоматическими датчиками Thermochron iButton DS1921 (Dallas Semiconductor, США) на глубине 5, 10 и 20 см измеряли температуру почвы. Температуру и влажность воздуха, а также фотосинтетически активную радиацию (плотность фотосинтетического фотонного потока) измеряли 1 раз в минуту сенсором Minikin QTH (EMS Brno, Чехия). В 2018 г. измерения потоков на сенокосе выполнены спустя 1 мес. после кошения, в 2019 г. – на следующий день. Измерения проводили на участках с доминированием мятлика; в 2018 г. с разделением на высокий (≈50 см) и низкий (≈25 см) травостой. Потоки CO2 измеряли одновременно на залежи и сенокосе методом динамических камер [33] разного размера и круглого сечения, из прозрачного пластика толщиной 0.5 мм с использованием инфракрасных газоанализаторов LI-820/840 (Li-Cor, США).
Регистрацию концентраций СО2 проводили непрерывно с помощью оригинального автоматического устройства на основе трехходовых переключателей, благодаря которым воздух через камеры проходил непрерывно. Показания регистрировали логгером круглосуточно (EMS, Чехия) каждые 5 с автоматическим сохранением каждые 20–30 с. Переключение между камерами было автоматическим, время “опроса” – 20–30 мин. Для расчетов концентрации CO2 использовали разность средней (за 2 мин) величины в поступающем и выходящем из камеры воздухе. Поток CO2 (мг С/м2/ч) рассчитывали по формуле (подробнее методика и схема измерений представлена в [34]):
(1)
$\begin{gathered} F~ = (106 \times 273 \times V([{\text{C}}{{{\text{O}}}_{{\text{2}}}}{\text{к] - [C}}{{{\text{O}}}_{{\text{2}}}}{\text{а}}])){\text{/}}(22.4 \times \\ \times \;100(273 + T)S \times 60), \\ \end{gathered} $По данным измерений NEE и Rsoil было рассчитано дыхание экосистемы (Reco) – совокупное гетеротрофное дыхание почвы, подземных и надземных частей растений по формуле:
где Rsoil – измеренное дыхание почвы, $R_{{{\text{plant}}}}^{{\text{*}}}$ – среднее дыхание надземных частей растений, рассчитанное на основе ночных измерений NEE и Rsoil. Дыхание надземных частей растений Rplant, используемое в формуле (2), было рассчитано как разность между средним (за одинаковые промежутки времени) дыханием экосистемы (Reco) и дыханием почвы (Rsoil) по формуле: где NEE* – нетто-экосистемный обмен ночью (измерения ночью – в отсутствии фотосинтетической активности растений, де-факто характеризующего Reco), $R_{{{\text{soil}}}}^{{\text{*}}}$ – дыхание почвы ночью. Расчет валового газообмена экосистемы (Gross ecosystem-atmosphere exchange of carbon dioxide – GEE) – общей фотосинтетической ассимиляции экосистемы или количества СО2, которое поглотила экосистема за единицу времени, проводили по формуле: где NEE – измеренный дневной нетто-экосистемный обмен, Reco – расчетное дыхание экосистемы. Обработку данных полевых и лабораторных измерений (корреляционный анализ, расчет средних и стандартных отклонений, множественная линейная регрессия) проводили в программе Excel. По данным измерений были также рассчитаны суммарные за 1 сут потоки СО2 с использованием метода бутстрапа [35] в программе MATLAB R2010b (количество бутстрап-повторений 5000). В каждом бутстрап-повторении для потоков СО2 с учетом погрешности измерений была получена случайная реализация их величин. При допущении об их нормальном распределении рассчитывали подинтегральную площадь, среднее арифметическое и стандартное отклонение реализаций суточной оценки, полученных в отдельных бутстрап-повторениях. При аппроксимации динамики потока за 1 сут использовали линейную интерполяцию.РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Характеристика участков. На залежи преобладала крапива двудомная и мятлик луговой, проективное покрытие которых менялось в течение вегетационного сезона: в начале лета преобладал мятлик, к концу – крапива. На сенокосе представлен исключительно мятлик. Оба участка имеют ровную поверхность с перепадом высот до 20–30 см; среднее относительное превышение залежи над сенокосом равно 10 см. На обоих участках УПГВ весной и осенью равен 50–60 см, летом в периоды без дождей – 80–90 см.
Метеоусловия. Лето 2018 г. (01.06–01.09) было теплым и сухим: средняя, минимальная и максимальная температуры (°C) воздуха (Твозд) составили 18.2, 3.5 (09.06), 29.5 (29.08), количество осадков – 130 мм, дней с осадками – 39. Лето 2019 г. было холодным и влажным: средняя, минимальная и максимальная Твозд составили 16.3, 6.0 (05.08 и 29.08) и 30.0 (07.06), осадки – 294 мм, дней с осадками – 66. Сумма активных температур (>10°C) за лето в 2018 г. – 1641, в 2019 г. – 1483°C.
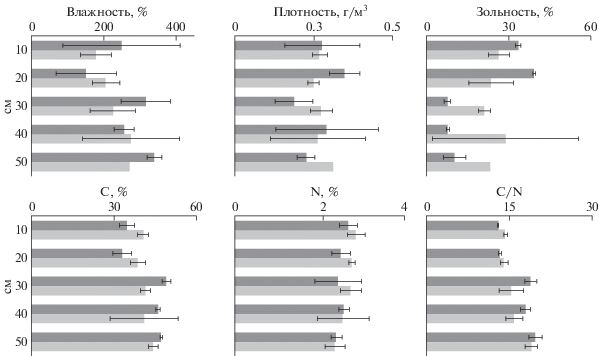
Почвенные условия. На залежи и сенокосе представлены осушенные низинные торфяные почвы. Ботанический состав торфа верхних 10 см отражает современную растительность: преобладают остатки корней злаков и крапивы (на залежи). Глубже – низинные древесно-травяные и древесные торфа: преобладают остатки ольхи, встречаются остатки березы, ивы, сосны и ели, отмечен эпидермис осок и камыша. Степень разложения на залежи и сенокосе менялась слабо: в среднем 35 и 50% в слое 0–30 см; 55 и 75% в слое 30–50 см соответственно. Влажность торфа (%) варьировала от (150 ± 84)–(178 ± 43) в верхнем (0–20 см) до (280 ± 134)–(340 ± 20) в нижнем (30–50 см) слое и плавно росла с глубиной на обоих участках (рис. 1). Плотность сложения существенно не менялась по профилю и варьировала от 0.19 ± 0.06 до 0.27 ± ± 0.04 г/см3 на обоих участках. Зольность в слое 0–20 см была немного меньше на сенокосе, чем на залежи (24 ± 8 и 39 ± 1%), а в слое 30–50 см наоборот – 21 ± 2 и 8 ± 1%. Незначительное повышение зольности на глубинах 30–50 см на сенокосе в сравнении с залежью слабо отражалось на содержании C и N в этом горизонте: 42–47 и 2.6–2.5% соответственно. В слое 40–50 см отмечено незначительное повышение содержания серы на сенокосе в сравнении с залежью – 0.8 и 0.5% соответственно. Отношение C : N не отличалось на сенокосе и залежи и слабо росло с глубиной: от 13.6 (0–10 см) до 19.3 (40–50 см) в среднем на обоих участках.
Рис. 1.
Влажность, плотность сложения и зольность почвы, cодержание углерода, азота в почве, соотношение C : N: залежь – темный тон, сенокос – светлый. То же на рис. 2–10.
Подземная и надземная фитомасса. На рис. 2 показаны изменения НФМ на залежи и сенокосе в 2019 г. в пересчете на сухое вещество. Вертикальной сплошной линией отмечена дата кошения на сенокосе – 24.08.19. На залежи 27.09.19 (пунктирная линия) впервые за несколько лет было проведено кошение для улучшения состояния травостоя. НФМ до кошения в 2019 г. слабо отличалась на обоих участках: возрастала с 24 в апреле до 267 г/м2 в июне на залежи и с 39 в апреле до 199 г/м2 в июле на сенокосе. После кошения 24 августа НФМ на сенокосе составила 32, спустя месяц, 27 сентября – 72 г/м2. В 2018 г. при более раннем кошении (28 июля) НФМ спустя 1 мес. (28 августа) на сенокосе составила 138 г/м2, т.е. в значительной степени восстановилась (например, в мае и июле 2019 г. она была лишь незначительно больше – 177 и 199 г/м2 соответственно). В августе 2018 г. НФМ на залежи составляла 368 г/м2.
Рис. 2.
Накопление надземной (круг) и подземной (квадрат) фитомассы растений на залежи и сенокосе (2019 г.).

ПФМ в верхних 10 см почвы в среднем за период с июня по сентябрь была в 2 раза больше (1220 г/м2) на сенокосе, чем на залежи (600 г/м2). Отмечено ее снижение на обоих участках от июня к сентябрю. Определение ПФМ проводили без разделения на мертвые и живые корни, поэтому, возможно, снижение величины ПФМ связано с преобладанием разложения мертвых корней над ростом живых.
Ранее (05.09.2008 г.) на неиспользуемом сенокосе в другой части этого массива НФМ составила 490, ПФМ (в слое 0–10 см) – 914 г/м2. Там же 25.09.2015 г. НФМ была равна 358 г/м2 [8], что близко полученным данным. Для сравнения, на сенокосных лугах в Костромской обл. НФМ составляла 147–324, а ПФМ – 647–1750 г/м2 [36]; на сенокосе выработанного низинного торфяника в Кировской обл. – 185 и 1580 г/м2 (слой 0–20 см) соответственно [37].
Показатель LAI на залежи в 2018 г. составил 2.5 для низкого и 3.9 для высокого травостоя, а на сенокосе – 1.5 и 1.8 соответственно. В 2019 г. этот показатель составил для залежи и сенокоса (после кошения) 1.3 и 0.5 соответственно. Полученные оценки LAI хорошо соотносятся с НФМ этих лет.
Целлюлозолитическая активность почвы. Потери органического вещества (%) в верхних 10 см были сопоставимы на сенокосе (19.6 ± 3.8) и залежи (12.9 ± 4.7), однако в среднем для слоя 0–30 см оказались меньше на сенокосе (8.2 ± 2.4), чем на залежи (14.4 ± 1.2) (табл. 1). Эти величины согласуются с данными для осушенных торфяников в конце периода вегетации [31]. Возможно, бóльшая скорость разложения на сенокосе в слое 0–10 см могла быть обусловлена в 2 раза большей массой корней: обитающие в прикорневой зоне микроорганизмы, лучшая аэрация и прогрев почвенного профиля способствовали интенсификации процессов разложения органического вещества. Это предположение подтверждено более интенсивным дыханием почвы на сенокосе в 2018 г.: после кошения почва прогревалась лучше, чем на залежи, что могло стимулировать корневое дыхание и разложение мертвых корней.
Таблица 1.
Целлюлозолитическая активность в профиле торфяной почвы на залежи и сенокосе, % потери массы целлюлозного полотна за 30 сут экспозиции
| Слой, см | Залежь | Сенокос |
|---|---|---|
| 0–10 | 12.9 ± 4.7 | 19.6 ± 3.8 |
| 10–20 | 21.2 ± 4.0 | 2.8 ± 2.1 |
| 20–30 | 9.2 ± 5.0 | 2.1 ± 1.2 |
| Среднее | 14.4 ± 1.2 | 8.2 ± 2.4 |
Нетто-экосистемный обмен (NEE). Показатель NEE в 2018 г. на залежи с высоким травостоем составлял ночью в среднем 4.0 и днем до –1.5 г CO2/м2/ч. Для низкого травостоя наблюдали эмиссию CO2 как ночью, так и днем (в среднем за 1 сут – 1.5 г CO2/м2/ч) (рис. 3). Изменения Твозд в течение суток достигали 20°C. Величина NEE была обратно пропорциональна Твозд и ФАР, что было особенно характерно для высокого травостоя. В 2018 г., через 1 мес. после кошения, на сенокосе с высоким травостоем наблюдали эмиссию CO2 ночью (≈2.0 г CO2/м2/ч) и поглощение днем (до –1.0), с низким травостоем – эмиссию CO2 в течение суток (≈2.5 г CO2/м2/ч).
Рис. 3.
Нетто-экосистемный обмен (NEE) на залежи и сенокосе спустя 1 мес. (27–28.08.2018) и сразу после кошения (28–29.08.2019); треугольники – низкий травостой, точки – высокий. Черная линия – температура почвы, серая – ФАР, пунктир – граница суток (00:00). То же на рис. 4.

В 2019 г. на залежи происходила эмиссия CO2 и ночью (≈4.0 г CO2/м2/ч) и днем (≈0.5 г CO2/м2/ч), только в утренние часы (7:00–9:00) было отмечено поглощение до –0.6 г CO2/м2/ч. В среднем за 1 сут залежь характеризовалась потерями углерода (≈1.0 г CO2/м2/ч). Суточные изменения Твозд на залежи также составили 20°C, хотя средняя температура была на 10°C меньше, чем в 2018 г. На сенокосе сразу после кошения отмечена эмиссия CO2 как ночью (≈0.6 г CO2/м2/ч), так и днем (≈0.5 г), суточная динамика была выражена слабо. Суточные изменения Твозд и ФАР не оказывали значительного влияния на величину NEE.
Дыхание почвы (Rsoil). Показатель Rsoil в 2018 г. менялся в течение 1 сут в пределах 0.7–2.0 и 1.0–2.3 г CO2/м2/ч на залежи и сенокосе соответственно, в 2019 г. – в пределах 0.2–2.5 и 0.3–2.9 г CO2/м2/ч. В 2018 г. суточная динамика Rsoil и Тпочв на глубине 5 см были идентичными (рис. 4). На сенокосе Rsoil было больше примерно на 25%, что, возможно, связано с более интенсивным дневным прогревом почвы или большей корневой биомассой. В 2019 г. суточная динамика Rsoil в меньшей степени совпадала с Тпочв.
Рис. 4.
Дыхание почвы на залежи и сенокосе через 1 мес. (27–28.08.2018) и сразу после кошения (28–29.08.2019).

Среднее за 1 сут Rsoil в 2019 г. было больше на сенокосе (1.8 г CO2/м2/ч), чем на залежи (1.3 г CO2/м2/ч), что может объяснить различиями в ПФМ (1223 и 604 г/м2 в среднем в течение сезона). Большая почти в 2 раза ПФМ на сенокосе могла быть обусловлена увеличением ассимиляции углерода растениями при регулярном кошении. Растения накапливают не только дополнительную НФМ, изымаемую при сенокосе, но и ПФМ. Корни растений, отмирая и разлагаясь, увеличивали показатель Rsoil.
Валовый обмен CO2экосистемы (GEE), дыхание экосистемы (Reco) и растений (Rplant). В 2018 г. средний GEE составил –0.8 для низкого и –1.7 г CO2/м2/ч для высокого травостоя на залежи (n = = 34); –1.8 и –1.9 для низкого и высокого травостоя на сенокосе (n = 52). При осреднении величин использованы одинаковые промежутки времени: 28.08.2018, 13:00–29.08.2018, 14:00 (за исключением ночи – до и после заката солнца) и 28.08.2019, 8:00–29.08.2019, 17:00. Несмотря на превышение НФМ на залежи в 2.7 раза над сенокосом (368 и 138 г/м2 соответственно), величины GEE на площадь произрастания были близкими. Средняя величина Reco (или ночной NEE) в 2018 г. на низком и высоком травостое залежи (n = 34) составила 1.9 и 3.6 г CO2/м2/ч, а на сенокосе (n = 62) – 1.9 и 4.0 г CO2/м2/ч. Reco не отличалось по величине на залежи и сенокосе, однако менялся вклад составляющих его компонентов: в первом случае преобладало дыхание растений, во втором – дыхание почвы, особенно на сенокосе с низким травостоем, где Rsoil практически равнялось Reco. Среднее дыхание растений (Rplant) в 2018 г. для залежи с низким травостоем составило 0.5, с высоким – 2.3 г CO2/м2/ч, на сенокосе – 0.1 и 2.2 г CO2/м2/ч соответственно.
Непосредственно после кошения на сенокосе в 2019 г. GEE для высокого травостоя отличался на залежи и сенокосе: –2.7 и –1.1 г CO2/м2/ч, соответственно. В сравнении с 2018 г. GEE на сенокосе был меньше в 1.6 раза, хотя НФМ сократилась после кошения практически в 6 раз. В 2019 г. Reco на залежи составило 3.2 г CO2/м2/ч (сопоставимо с высоким травостоем на залежи в 2018 г.), а на сенокосе после кошения – 1.7 г CO2/м2/ч, что сопоставимо с низким травостоем на сенокосе в 2018 г. (рис. 3). Величина Rplant в 2019 г. составила 1.7 г CO2/м2/ч для залежи, что сравнимо с результатами 2018 г. при высоком травостое, и 0.3 г CO2/м2/ч для сенокоса, что сравнимо с результатами 2018 г. при низком травостое.
Зависимость GEE, NEE, Reco, и Rsoil от температуры воздуха, почвы и ФАР. По данным 2018 г. для сенокоса (через месяц после кошения) и залежи были получены коэффициенты регрессионного уравнения (табл. 2):
где GEE – валовый обмен CO2 экосистемы, г CO2/м2/ч; a, b, c – безразмерные коэффициенты для ФАР, температуры воздуха и пересечения с осью ординат соответственно; Твозд – температура воздуха, °C; ФАР – фотосинтетически активная радиация, мкмоль/м2/с.Таблица 2.
Коэффициенты уравнений зависимости потоков CO2 от факторов среды
| Коэффициент | Залежь | Сенокос | ||
|---|---|---|---|---|
| травостой | ||||
| низкий | высокий | низкий | высокий | |
| GEE от температуры воздуха и ФАР | ||||
| a | –2 × 10–3 ± 1 × 10–3 | 1 × 10–3 ± 1 × 10–3 | –2 × 10–3 ± 0 × 10–3 | –1 × 10–3 ± 0 × 10–3 |
| b | 6 × 10–2 ± 4 × 10–2 | –1.0 ± 0.1 | 7 × 10–2 ± 2 × 10–2 | –1 × 10–2 ± 2 × 10–2 |
| c | –1.6 ± 1.4 | 11.8 ± 1.5 | –3.0 ± 0.4 | –1.4 ± 0.4 |
| R2 | 0.24 | 0.88 | 0.38 | 0.41 |
| NEE от температуры почвы, воздуха и ФАР (день) | ||||
| a | –1 × 10–3 ± 0 × 10–3 | –1 × 10–3 ± 0×10–3 | –1 × 10–3 ± 0 × 10–3 | –1 × 10–3 ± 0 × 10–3 |
| b | 1.0 ± 0.2 | –0.3 ± 0.3 | 1.0 ± 0.6 | 0.7 ± 0.6 |
| c | –0.3 ± 0.1 | –0.2 ± 0.1 | –0.3 ± 0.2 | –0.3 ± 0.2 |
| d | –7.8 ± 1.9 | 7.8 ± 2.4 | –2.9 ± 3.5 | –1.2 ± 3.4 |
| R2 | 0.63 | 0.84 | 0.50 | 0.69 |
| Reco от температуры почвы и воздуха (ночь) | ||||
| a | 0.5 ± 0.4 | 1.9 ± 0.8 | –0.1 ± 2.2 | 0.8 ± 1.2 |
| b | –0.3 ± 0.1 | –1.0 ± 0.3 | 0.1 ± 0.8 | –0.3 ± 0.4 |
| c | –2.1 ± 3.4 | –9.3 ± 6.1 | 3.8 ± 13.2 | –2.7 ± 7.1 |
| R2 | 0.22 | 0.53 | 0.07 | 0.05 |
| Rsoil от температуры почвы | ||||
| a | 0.1 ± 0.1 | 0.1 ± 0.1 | ||
| b | 0.2 ± 0.1 | 0.6 ± 0.1 | ||
| R2 | 0.56 | 0.69 | ||
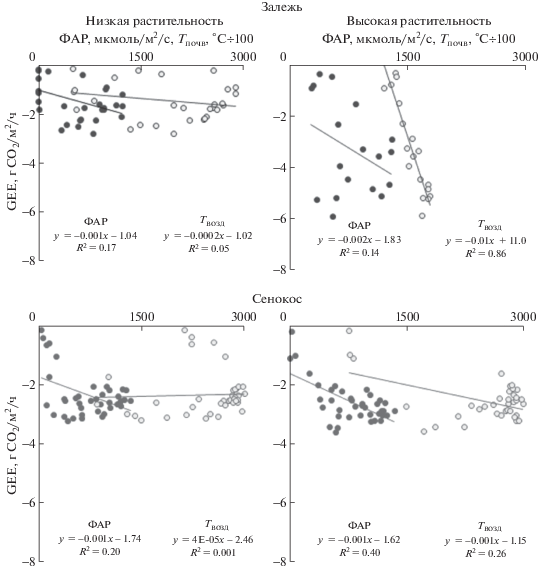
Установлена обратная связь (R2) GEE с ФАР – при росте ФАР GEE принимает “более” отрицательные величины, что свидетельствовало об увеличении фотосинтетической активности растений, и убывает в ряду: сенокос–высокий травостой (0.40), сенокос–низкий травостой (0.20), залежь–низкий травостой (0.17), залежь–высокий травостой (0.14) (рис. 5). Значимая корреляция была характерна только для высокого травостоя на сенокосе. Возможно, меньшее самозатенение растений определяло в этом случае более явный отклик на изменение поступающей радиации, тогда как на залежи только часть незатененных растений была способна отреагировать на изменение ФАР. Связь GEE с Tвозд была прямой для высокого травостоя и отсутствовала для низкого. R2 убывал в ряду: залежь–высокий травостой (0.86), сенокос–высокий травостой (0.26), залежь–низкий травостой (0.05), сенокос–низкий травостой (0.001). На залежи и сенокосе с высоким травостоем температура воздуха влияла сильнее, чем ФАР. Таким образом, более точно могут быть предсказаны GEE на сенокосе и залежи с высоким травостоем.
Рис. 5.
Связь GEE с ФАР (точки) и температурой воздуха (светлые точки) на залежи и на сенокосе с низким и высоким травостоем.
Согласно результатам регрессионного анализа, в дневное время (с 5:00 до 19:00) величина NEE зависела от ФАР, температуры почвы и воздуха. Коэффициенты уравнения (6) даны в табл. 2:
(6)
${\text{NEE}} = a \times {\text{ФАР}} + b \times {{T}_{{{\text{почв}}}}} + c \times {{T}_{{{\text{возд}}}}} + d,$Связь NEE с ФАР, как и для GEE, была обратной: при росте ФАР NEE принимает “более” отрицательные величины, что говорит о существенном вкладе GEE в нетто–экосистемный обмен. NEE демонстрирует аналогичную с GEE зависимость от Тпочв и Твозд. Связь (R2) NEE с ФАР убывает в ряду: сенокос–высокий травостой (0.65), сенокос–низкий травостой (0.37), залежь–низкий травостой (0.36), залежь–высокий травостой (0.21) (рис. 6). Значимая корреляция характерна для сенокоса с высоким травостоем, а для сенокоса и залежи с низким травостоем находится на границе значимости. Зависимость NEE от ФАР подтверждена показанной ранее связью GEE и ФАР, однако требует дальнейшей проверки для низкого травостоя на сенокосе. Для высокого травостоя на сенокосе это также касается связи NEE с температурой почвы и воздуха.
Рис. 6.
Корреляция дневного NEE с температурой почвы на глубине 5 см (светлые точки), воздуха (выколотые точки) и ФАР (точки) на залежи (сверху) и на сенокосе (снизу) на участках с низким (слева) и высоким (справа) травостоем.

Связь (R2) NEE с температурой почвы убывала в ряду: залежь–высокий травостой (0.81), сенокос–высокий травостой (0.32), на залежи и сенокосе с низким травостоем связь отсутствовала (0.03). Связь NEE с температурой воздуха была наибольшей на участках с высоким травостоем: 0.79 и 0.41 и отсутствовала на участках с низким травостоем (0.11 и 0.01 – залежь и сенокос соответственно). Во всех случаях, кроме низкого травостоя на сенокосе, характерно смещение NEE в сторону ассимиляции при повышении ФАР и температуры. Это связано со значительным вкладом GEE в NEE (в сравнении с Reco), что обусловило варьирование газообмена сенокоса и залежи при изменениях ФАР.
Для ночных потоков NEE, характеризующих Reco (дыхание экосистемы) в отсутствии фотосинтеза, было использовано уравнение (7), коэффициенты которого даны в табл. 2:
(7)
${{R}_{{{\text{eco}}}}}~ = a \times {{T}_{{{\text{почв}}}}} + b \times {{T}_{{{\text{возд}}}}} + c,$Рис. 7.
Связь ночного NEE (Reco) с температурой почвы (точки) и воздуха (выколотые точки) на залежи (сверху) и на сенокосе (снизу) на участках с низким (слева) и высоким (справа) травостоем.

Дыхание экосистемы является многокомпонентным параметром, и его составляющие могут иметь различную связь с факторами среды, что при совместном рассмотрении может привести к маскировке закономерностей. Также возможно, что расчетное дыхание растений (Rplant), используемое при расчете Reco, может вносить определенную погрешность в его оценку.
Показано, что Rsoil значимо зависело от Тпочв, R2 составил 0.60 и 0.70 на залежи и сенокосе соответственно (рис. 8). Коэффициенты уравнения (8) даны в табл. 2:
где Rsoil – дыхание почвы, г CO2/м2/ч, a, b – безразмерные коэффициенты регрессионного уравнения для температуры почвы и пересечения с осью ординат, Tпочв – температура почвы, °C.
Интегральная (за 1 сут) оценка NEE, GEE, Reco, Rsoil. Средние (для высокого и низкого травостоя) NEE на залежи и сенокосе через 1 мес. после кошения в 2018 г. не отличались – 39 и 38 г CO2/м2/сут соответственно (рис. 9). Однако на сенокосе наблюдали бóльшую пространственную неоднородность NEE, основной вклад в которую вносил, вероятно, газообмен надземных частей растений. Непосредственно после кошения в 2019 г. NEE на сенокосе была больше, чем на залежи: 9 и 5 г CO2/м2/сут соответственно.

При расчете для летнего периода продолжительностью 90 сут показатель NEE на залежи в 2019 и 2018 гг. составил 1.4 и 11.0 т C/га/год; на сенокосе – 2.6 и 10.9 соответственно. Это сопоставимо с оценками потерь углерода сенокосами на осушенных верховых и низинных торфяниках: от 2.5–3.5 [38, 39] до 4.6–6.0 [11] и на низинных торфяниках – 3.7–4.2 т C/га/год. Более широкий, относительно литературных данных, диапазон наших оценок можно объяснить тем, что летний сезон 2019 г. был влажным и аномально холодным, а 2018 г. – оптимально влажным и теплым. УПГВ (80–90 см), который наблюдали в 2018 г. на исследованных объектах, в сочетании с оптимальными термическими условиями мог способствовать максимальным потерям почвенного углерода [10]. Используемые камеры имеют небольшую площадь основания, и для корректного пересчета потоков диоксида углерода (в т C/га/год) необходимо учитывать неоднородность растительного покрова, дополняя это измерениями площади листьев и фитомассы растений, находящихся в камере. Съемка с БПЛА показала значительную неоднородность растительного покрова, что требует дальнейшего более пристального изучения. Средняя за 2 года величина NEE составляла 6.2 для залежи и 6.8 т C/га/год для сенокоса, что свидетельствовало о незначительных различиях между участками.
Величины Rsoil в 2018 г. (г CO2/м2/сут) были несколько выше на сенокосе, чем на залежи (42 и 32 г CO2/м2/сут), а в 2019 г. они были равны 21 и 22 г CO2/м2/сут соответственно. При этом ПФМ составила в среднем 1220 и 600 г/м2 в слое 0–10 см на сенокосе и залежи. Можно предположить, что вклад корневого дыхания и разложения мертвых корней был больше на сенокосе. Дыхание почвы на осушенных торфяниках характеризуется значительным варьированием: 0.1–2.5 г CO2/м2/сут в Московской и Ярославской обл. [24, 25], 1.7, 2.1 г CO2/м2/сут – в Карелии и Центральной России соответственно [40], 1.2–5.3 г CO2/м2/сут – в Швейцарии [41], 3.1–7.8 г CO2/м2/сут – в Беларуси [41, 42]. Но даже на минеральных почвах отмечают значительно бóльшие величины Rsoil: до 35–65 г CO2/м2/сут на залежных участках в Тверской обл. [43]. Потенциальная величина дыхания торфяной почвы, определенная в лабораторных условиях, также меняется широко и может достигать 53.3 г CO2/м2/сут [41]. Ранее на другом участке неиспользуемого сенокоса Дубненского болотного массива среднее за бесснежный период дыхание почвы варьировало от 10.7 ± 0.3 до 16.0 ± ± 2.5 г CO2/м2/сут [44], что согласуется с данными представленной работы, которые были получены летом и, вероятно, характеризуют близкие к максимальным величины Rsoil для осушенных органогенных почв.
Величины Reco в 2018 г. были близкими на сенокосе и залежи (в среднем 69 и 66 г CO2/м2/сут соответственно), но значительно зависели от высоты травостоя: на сенокосе с низкой растительностью – 94, с высокой – 44, на залежи – 45 и 87 г CO2/м2/сут соответственно. В 2018 г. Reco в среднем, как на залежи, так и на сенокосе в 2 раза больше GEE, и оба участка характеризовались потерями диоксида углерода. В 2019 г. (непосредственно после кошения на сенокосе) Reco на сенокосе составило 26, на залежи – 50 г CO2/м2/сут. Предположительно, более половины Reco обусловлено дыханием НФМ, кошение которой в 2019 г. на сенокосе снизило экосистемное дыхание более чем в 2 раза.
Суточные GEE на сенокосе и на залежи в 2018 г. был близкими: –32 и –34 г CO2/м2/сут при том, что НФМ отличалась почти в 3 раза: 138 и 368 г/м2 соответственно. Вероятно, прореживание растений в результате кошения приводит к уменьшению самозатенения растений и росту удельной (на величину НФМ – GEEуд г CO2/м2/сут/граст) ассимиляции, что обеспечивало сопоставимые величины GEE на сенокосе и залежи. Это подтверждено результатами 2019 г.: непосредственно после кошения на сенокосе GEE (–18 г CO2/м2/сут) в сравнении с залежью (–45 г CO2/м2/сут) отличался значительно меньше, чем НФМ – 32 и 293 г/м2.
НФМ в значительной степени определяла различия между компонентами углеродного баланса залежи и сенокоса: ее увеличение сопровождалось ростом Rplant и GEE с постепенным выходом на плато при НФМ $ \gtrsim $300 г/м2 (рис. 10) – черные и серые маркеры соответственно; знаки потоков даны с точки зрения накопления или потери органического вещества растениями, а не изменения концентрации в атмосфере, как это было ранее. GEEуд при этом достаточно быстро уменьшался, достигая минимума при наибольшей фитомассе.
Рис. 10.
Зависимость дыхания надземных частей растений Rplant (черные квадраты), удельного GEE (GEEуд – на единицу надземной фитомассы выколотые точки) и GEE на площадь произрастания (серые точки) от надземной фитомассы растений (Гнадзем. фитомасса) .

Оценка GEE, Rplant, Rroot, Rmort по динамике фитомассы растений. Была предпринята попытка сопоставить GEE с приростом НФМ до кошения, дыхания живой корневой массы и разложения мертвой на залежи и сенокосе, а также сравнить последнее со скоростью разложения, оцененной в полевом опыте по величине целлюлозолитической активности почвы. Прирост НФМ за вегетационный период 2019 г. был определен как разница между НФМ в августе и апреле для залежи и в июле и апреле для сенокоса. Так как прирост ПФМ неизвестен, было принято допущение о том, что он был равен относительному приросту НФМ. Величина прироста общей фитомассы (ПФМ + НФМ) была переведена в поток (г CO2/м2/сут, с учетом содержания углерода в сухой биомассе растений 45% [45, 46]) и, по существу, характеризовала NPP (Net primary production – чистую первичную продукцию) (табл. 3).
Таблица 3.
Расчетные (по приросту фитомассы) показатели GEE, Rplant,Rroot и Rmort для залежи (с апреля по август 2019 г.) и сенокоса (с апреля по июль 2019 г.)
| Дата | Залежь | Сенокос | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Надземная фитомасса | Rplant | GEE | Rplant + + GEE | Rroot : : Rmort | Надземная фитомасса | Rplant | GEE | Rplant + + GEE | Rroot : : Rmort | |||
| г/м2 | Δ г/м2 : Δ г CO2/м2/сут | г CO2/м2/сут | г/м2 | Δ г/м2 : Δ г CO2/м2/сут | г CO2/м2/сут | |||||||
| 25.04 | 24 | 270 : 7 | –3 | 16 | 13 | –2 : –30 | 39 | 160 : 5 | –9 | 21 | 12 | –4 : –38 |
| 19.05 | 88 | –18 | 28 | 10 | 177 | –26 | 34 | 8 | ||||
| 19.06 | 267 | –30 | 37 | 7 | 188 | –26 | 34 | 8 | ||||
| 31.07 | 192 | –27 | 34 | 8 | 199 | –27 | 35 | 8 | ||||
| 31.08 | 293 | –31 | 38 | 7 | – | – | – | – | ||||
На основе выявленных зависимостей GEE и Rplant с НФМ (рис. 10) были рассчитаны их величины с апреля по август для залежи и с апреля по июль для сенокоса, и усреднены за рассмотренный период. Разница между NPP и суммой средних величин GEE + Rplant характеризовала величину дыхания живых корней растений (Rroot), выраженную в г CO2/м2/сут. Разность между величинами среднего дыхания почвы (Rsoil) и Rroot являлась показателем гетеротрофного дыхания почвы, или потока диоксида углерода, сопровождающего разложение органики, в том числе корней растений (Rmort). Отношение скорости разложения целлюлозы на сенокосе к залежи составило 1 : 1.52 и оказалось близким к отношению расчетной скорости разложения мортмассы – 1 : 1.26.
ПФМ, отличающаяся в 2 раза на сенокосе и залежи, обусловила двукратное различие скорости дыхания живых корней (–4 и –2 г CO2/м2/сут) и 25%-ное различие скорости разложения мертвых корней. Возможно, что эти отличия определялись не только разницей в количестве ПФМ, но и температурой почвы, зольностью и др. Расчеты предварительные, однако могут отражать незначительные различия между компонентами баланса диоксида углерода на залежи и сенокосе, зависимые от динамики величины их НФМ и ПФМ в течение сезона.
Потери углерода на сенокосе слабо отличались от залежи, однако эта оценка не учитывала изъятие фитомассы при кошении (рис. 9). Использование осушенных торфяников под сенокос способно уменьшить потери углерода в сравнении с пашней примерно на 20% [11], а при оптимальном сочетании частоты и времени скашивания даже способствовать накоплению углерода и азота в почве [19]. При этом слишком частое кошение без внесения удобрений может привести к истощению углеродного запаса почв и ухудшению их физико-химических и агротехнических характеристик.
ЗАКЛЮЧЕНИЕ
Сравнение почвенно-гидрологических условий сенокосного и залежного участков осушенного торфяника не выявило заметных различий между ними, что позволило отнести различия в потоках СО2 к влиянию характера их использования.
Установлена зависимость валового обмена CO2 экосистемы (GEE) от температуры воздуха (R2 = 0.86 на залежи) и ФАР (R2 = 0.40 на сенокосе); нетто-экосистемного обмена CO2 с атмосферой (NEE) – от температуры воздуха, почвы (R2 = = 0.79 и 0.81 на залежи соответственно) и ФАР (R2 = 0.65 на сенокосе и 0.37 – на залежи). Для дыхания почвы (Rsoil) выявлена зависимость от ее температуры: R2 = 0.69 на сенокосе и 0.56 – на залежи. Полученные коэффициенты регрессионных уравнений могут быть использованы для интерпретации динамики NEE за вегетационный период.
На сенокосе непосредственно после кошения в 2019 г. экосистемное дыхание (Reco) было в 2 раза меньше, чем через 1 мес. после кошения на высоком травостое в 2018 г. (26 и 44 г CO2/м2/сут), аналогично менялся и показатель GEE (–18 и –34 г CO2/м2/сут). При этом на залежи в 2019 г. показатели Reco и GEE составили 50 и –45 г CO2/м2/сут соотвественно. Показатель GEEуд (GEE на единицу надземной фитомассы растений) на сенокосе сразу после кошения был в 2.4 раза больше, чем через 1 мес. после кошения (0.56 и 0.23 г CO2/м2/сут/граст), что возможно было связано с уменьшением самозатенения растений. Показатели NEE и Rsoil в 2019 г. составили 9 и 21 г CO2/м2/сут на сенокосе и 5 и 22 г CO2/м2/сут на залежи соответственно.
На сенокосе через 1 мес. после кошения в 2018 г. параметры Reco, Rsoil, GEE и NEE в среднем для высокого и низкого травостоя (69, 42, –32 и 38 г CO2/м2/сут соответственно) практически не отличались от залежи (66, 32, –34 и 39 г CO2/м2/сут), хотя надземная фитомасса (НФМ) на сенокосе была в 2.7 раза меньше (138 г/м2), чем на залежи (368 г/м2), а подземная (ПФМ) – наоборот (1223 и 600 г/м2).
Бóльшая величина ПФМ на сенокосе была связана, вероятно, с активным ростом растений, в том числе корневой системы, как приспособительной реакцией в ответ на кошение, о чем свидетельствовало увеличение показателя GEEуд.
Согласно расчетам, на основе прироста НФМ сенокошение незначительно влияло на величину NEE. Суммарно за вегетационный сезон 2019 г. он был близким для сенокоса и залежи и составил 5 и 7 г CO2/м2/сут. Однако в этом случае менялось соотношение входящих в NEE компонентов: увеличивался вклад дыхания живых и разложения мертвых корней, а непосредственно после уборки уменьшалось дыхание надземных частей растений и увеличивался GEEуд.
Использование осушенного торфяника под сенокос позволяет получить сельскохозяйственную продукцию и за счет контроля пользователем снизить опасность пожаров. При этом такое использование не оказывает существенного влияния на величину нетто-экосистемного обмена СО2 за вегетационный период, а с учетом изъятой фитомассы, вероятно, даже смещает его в сторону ассимиляции CO2 в сравнении с залежью.
Авторы признательны М.В. Смагиной за советы по изучению процесса разложения в почве, М.М. Метелевой – за анализ ботанического состава торфа, А.А. Маслову (ИЛАН РАН) – за подбор спутниковых снимков и Д.Г. Суворову – за съемку объектов исследования с БПЛА, А.В. Маркиной, М.В. Левчуку (ИЛАН РАН), М.В. Чистотину (ВНИИ агрохимии) – за помощь в проведении полевых работ.
Список литературы
Торфяные болота России: к анализу отраслевой информации // Под ред. Сирина А.А., Минаевой Т.Ю. М.: Геос, 2001. 190 с.
A Quick scan of peatlands in central and Eastern Europe / Eds. Minayeva T., Sirin A., Bragg O. wageningen, The Netherlands: Wetlands International, 2009. 132 p.
Sirin A., Minayeva, T., Yurkovskaya T., Kuznetsov O., Smagin V., Fedotov Yu. Russian Federation (European Part) // Mires and peatlands of Europe: Status, distribution and conservation /Eds. Joosten H., Tanneberger F. Moen A. Stuttgart: Schweizerbart Science Publishers, 2017. P. 589–616.
Tanneberger F., Tegetmeyer C., Busse S., Barthelmes A. The peatland map of Europe // Mires and Peat. 2017. V. 19 (22). P. 1–17. (Online: http://www.mires–and–peat.net/pages/volumes/map19/map1922.php); https://doi.org/10.19189/MaP.2016.OMB.264
Лукин С.М., Анисимова Т.Ю. К вопросу о сельскохозяйственном торфопользовании в России // Мелиорация. 2016. № 1 (75). С. 48–54.
Сирин А.А., Минаева Т.Ю., Возбранная А.Е., Барталев С.А. Как избежать торфяных пожаров? // Наука в России. 2011. № 2. С. 13–21.
Чистотин М.В., Сирин А.А., Дулов Л.Е. Сезонная динамика эмиссии углекислого газа и метана при осушении болота в Московской области для добычи торфа и сельскохозяйственного использования // Агрохимия. 2006. № 6. С. 54–62.
Суворов Г.Г., Чистотин М.В., Сирин А.А. Потери углерода при добыче торфа и сельскохозяйственном использовании осушенного торфяника в Московской области // Агрохимия. 2015. № 11. С. 51–62.
Ильясов Д.В., Сирин А.А., Суворов Г.Г., Мартыненко В.Б. Летние потоки диоксида углерода и метана на осушенном торфянике в условиях лесостепи республики Башкортостан // Агрохимия. 2017. № 1. С. 50–62.
Assessment on peatlands, biodiversity and climate change. Main report / Eds. Parish F., Sirin A., Charman D., Joosten H., Minayeva T., Silvius M., Stringer L. Kuala Lumpur and Wetlands Int., Wageningen: Global Environment Centre, 2008. 179 p.
Peatlands and climate change / Ed. Strack. S. Saarijarven Offset Oy, 2008. 223 p.
IPCC: 2013 Supplement to the 2006 IPCC Guidelines for National Greenhouse Gas Inventories: Wetlands / Eds. Hiraishi T., Krug T., Tanabe K., Srivastava N., Baasansuren J., Fukuda M., Troxler T.G. Switzerland: IPCC, 2014.
Зайдельман Ф.Р., Шваров А.П. Пирогенная и гидротермическая деградация торфяных почв, их агроэкология, песчаные культуры земледелия, рекультивация. М.: Изд-во МГУ, 2002. 168 с.
Уланов А.Н., Журавлева Е.Л. Экологические аспекты освоения и сельскохозяйственного использования торфяных почв и выработанных торфяников // Теор. и прикл. экол. 2009. № 3. С. 94–96.
Трускавецкий Р.С. Баланс углерода в осушенных торфяниках украинского полесья // Почвоведение. 2014. №. 7. С. 829–836.
Рабинович Г.Ю., Широкова Е.В., Поздняков Л.А., Позднякова А.Д., Анциферова О.Н., Пантелеева Т.Н. Деградационные процессы в мелиорированных низинных торфяных почвах ландшафтов Яхромской долины // Новые методы и результаты исследований ландшафтов в Европе, Центральной Азии и Сибири / Под ред. Сычева В.Г., Мюллера Л.М., 2018. С. 306–310.
Курганова И.Н., Ермолаев А.М., Лопес де Гереню В.О., Ларионова А.А., Келлер Т., Ланге Ш., Кузяков Я.В. Баланс углерода в залежных землях Подмосковья // Почвоведение. 2007. № 1. С. 60–68.
Лопес де Гереню В.О., Курганова И.Н., Ермолаев А.М., Кузяков Я.В. Изменение пулов органического углерода при самовосстановлении пахотных черноземов // Агрохимия. 2009. № 5. С. 5–12.
Ларионова A.A., Ермолаев А.М., Никитишен В.И., Лопес де Гереню В.О., Евдокимов И.В. Баланс углерода в пахотных серых лесных почвах при разных способах сельскохозяйственного использования // Почвоведение. 2009. №. 12. С. 1464–1474.
Шевцова Л.К., Романенков В.А., Благовещенский Г.В., Хайдуков К.П., Канзыва С.О. Структура баланса углерода и биоэнергетическая оценка его компонентов в агроценозах длительных полевых опытов // Агрохимия. 2015. № 12. С. 67–75.
Малханова Е.В., Егорова Р.А., Чимитдоржиева Г.Д. Баланс углерода в лугово-черноземных мерзлотных почвах юга Витимского плоскогорья // Агрохимия. 2011. № 1. С. 3–7.
Помазкина Л.В., Семенова Ю.В., Симакова А.А., Кириллова Н.Н., Башалханов И.А. Оценка баланса углерода почв в агроэкосистемах Иркутской области // Агрохимия. 2016. № 5. С. 65–71.
Зайдельман Ф.Р., Шваров А.П. Потоки диоксида углерода в осушенных торфяных почвах // Вестн. МГУ. Сер. 17. Почвоведение. 2001. № 3. С. 867–874.
Новиков В.В., Степанов А.Л., Поздняков А.И., Лебедева Е.В. Сезонная динамика эмиссии СО2, СН4, N2O и NO из торфяных почв поймы р. Яхрома // Почвоведение. 2004. № 7. С. 867–874.
Новиков В.В., Русаков А.В. Выделение и поглощение парниковых газов в мелиорированных торфяных почвах Ростовской низины // Почвоведение. 2005. № 7. С. 844–850.
Аладко С.В., Трибис В.П., Шкутов Э.Н. Минерализация органического вещества осушенных торфяников при длительном сельскохозяйственном использовании // Мелиорация переувлажненных земель. 2005. № 2 (54). С. 94–99.
Глаголев М.В., Чистотин М.В., Шнырев Н.А., Сирин А.А. Летне-осенняя эмиссия диоксида углерода и метана осушенными торфяниками, измененными при хозяйственном использовании, и естественными болотами (на примере участка Томской области) // Агрохимия. 2008. № 5. С. 46–58.
Сирин А.А., Маслов А.А., Валяева Н.А., Цыганова О.П., Глухова Т.В. Картографирование торфяных болот Московской области по данным космической съемки высокого разрешения // Лесоведение. 2014. № 5. С. 65–71.
Информационный выпуск “О состоянии природных ресурсов и окружающей среды Московской области в 2015 году”. М.: ИП “Алексашин А.А.”, 2016. 206 с.
Вомперский С.Э., Сирин А.А., Глухов А.И. Формирование и режим стока при гидролесомелиорации. М.: Наука, 1988. 168 с.
Смагина М.В. Активность целлюлозоразрушающих микроорганизмов торфяно-болотных почв // Лесоведение. 1984. № 1. С. 52–58.
Суворов Г.Г., Чистотин М.В., Сирин А.А. Влияние растительности и режима увлажнения на эмиссию метана из осушенной торфяной почвы // Агрохимия. 2010. № 12. С. 40–49.
Edwards N.N., Sollins P. Continuous measurement of carbon dioxide evolution from partitioned forest floor components // Ecology. 1973. V. 54. № 2. P. 406–412.
Молчанов А.Г. Газообмен диоксида углерода с поверхности сфагнума в заболоченном сосняке южной тайги // Динамика окруж. среды и глобал. изменения климата. 2017. Т. 8. № 1. С. 43–54.
Davison A.C., Hinkley D.V. Bootstrap methods and their application. Cambridge: Cambridge University Press, 1997. 582 p.
Владыченский А.С., Телеснина В.М., Румянцева К.А., Чалая Т.А. Органическое вещество и биологическая активность постагрогенных почв южной тайги (на примере Костромской области) // Почвоведение. 2013. №. 5. С. 518–529.
Ковшова В.Н. Средообразующая роль создания и использования сеяных долголетних сенокосов на низинных выработанных торфяных почвах в условиях Волго-Вятского региона // Висвітлено матеріали IV Міжнародної наукової конференції “Корми і кормовий білок” (26–27 червня 2006 року), які відбулися у м. Вінниці на базі Інституту кормів. 2006. С. 45.
Joosten H., Clarke D. Wise use of mires and peatlands. Jyväskylä, Finland: International Mire Conservation Group & International Peat Society, 2002. 304 p.
Schipper L.A., McLeod M. Subsidence rates and carbon loss in peat soils following conversion to pasture in the Waikato Region, New Zealand // Soil Use Manag. 2002. V. 18. № 2. C. 91–93.
Уланов А.Н., Смирнова А.В. Опыт использования 115 торфяных месторождений на Кировской луго-болотной опытной станции // Мелиорация. 2009. Т. 61. С. 191–201.
Bader C., Müller M., Schulin R., Leifeld J. Peat decomposability in managed organic soils in relation to land use, organic matter composition and temperature // Biogeosciences. 2018. V. 15. №. 3. P. 703–719.
Белковский В.И., Решетник А.П. Динамика эмиссии СО2 из торфяной почвы при различном ее использовании // Почвоведение. 1981. № 6. С. 57–61.
Комарова Т.В. Экологическая оценка почвенной эмиссии СО2 в сукцессионном ряду зарастания залежи на территории Центрально-лесного заповедника: Дис. … канд. биол. наук. М., 2019. 159 с.
Суворов Г.Г. Изменение потоков СО2, СН4 и запасов углерода лесоболотной экосистемой в результате добычи торфа и сельскохозяйственного использования (на примере Дубненского массива Московской обл.): Дис. … канд. биол. наук. М., 2018. 128 с.
Poorter H., Bergkotte M. Chemical Composition of 24 Wild Species Differing in Relative Growth Rate // Plant Cell Environ. 1992. V. 15. P. 221–229.
Niinemets U., Kull O., Tenhunen J.D. Variability in leaf and chemical composition as a function of canopy light environment in coexisting deciduous trees // Inter. J. Plant Sci. 1999. V. 160. P. 837–848.
Дополнительные материалы отсутствуют.