

Агрохимия, 2021, № 3, стр. 71-81
Исследование фитотоксичности свинца для растений редиса и салата при выращивании на разных типах почв
А. В. Дикарев 1, *, В. Г. Дикарев 1, Н. С. Дикарева 1
1 Всероссийский научно-исследовательский институт радиологии и агроэкологии
249032 Обнинск, Калужская обл., Киевское шоссе, 109 км, Россия
* E-mail: ar.djuna@yandex.ru
Поступила в редакцию 17.01.2020
После доработки 18.07.2020
Принята к публикации 10.12.2020
Аннотация
Растущее загрязнение окружающей среды техногенными поллютантами стало серьезной проблемой для аграрных экосистем. Одной из значимых групп таких загрязнителей являются тяжелые металлы, в частности, свинец. В связи с этим, оценка его воздействия на основные сельскохозяйственные культуры становится важной задачей для обеспечения продовольственной безопасности страны. В вегетационном опыте на разных типах почв изучали влияние свинца на морфофизиологические и биохимические показатели растений редиса и салата-латука. Анализировали площадь листьев, воздушно-сухую массу корнеплодов (редис) и растений в целом (салат), а также содержание свободного пролина и малонового диальдегида. С возрастанием дозы тяжелого металла (ТМ) происходило подавление роста и развития растений обеих культур, особенно салата, сокращалась площадь листьев и биомасса. Наблюдаемые эффекты зависели от типа почвы и были наиболее выражены на дерново-подзолистой почве, для которой отмечены резкие изменения содержания пролина и МДА, особенно в области низких концентраций ТМ. Данные результаты имеют важное фундаментальное значение для углубления понимания реакций организма на средовой стресс. Несомненно важно и их прикладное значение в деле решения проблемы охраны здоровья человека и домашних животных.
ВВЕДЕНИЕ
Изучение загрязнения агросферы тяжелыми металлами (ТМ) и его влияние на рост и развитие сельскохозяйственных растений, а также их продуктивность приобретает все большее значение. Например, >1 млн га сельскохозяйственных угодий России загрязнено особо токсичными элементами (I класс опасности) и 2.3 млн га – токсичными (II класс опасности). По данным ВОЗ, только свинца в атмосферу поступает ежегодно не менее 250 тыс. т. Городская пыль содержит ≈1% свинца, в дожде и снеге его количество достигает до 300 мг/дм3 [1].
В целом процессы поступления и метаболизма ТМ, а также механизмы устойчивости к ним являются общими для всех живых организмов, в том числе и для растений. Однако в случае с сельскохозяйственными культурами при исследовании воздействия ТМ следует учитывать не только видовые особенности растений, но и свойства почв, на которых они произрастают, такие как уровнень плодородия, гранулометрический состав и их физико-химические свойства. Именно эти особенности оказывают модифицирующее влияние на выраженность действия ТМ на сельскохозяйственные культуры.
Некоторые ТМ, такие как Cu, Zn, Co являются необходимыми для растений микроэлементами [2], без которых невозможно поддержание гомеостаза организма, хотя избыток этих элементов приводит к нарушениям его функций. В то же время в почве природных и аграрных экосистем присутствуют иные ТМ, биологическая роль которых изучена недостаточно. К ним относятся кадмий, свинец и ртуть, считающиеся высокотоксичными элементами.
Существенный интерес среди высокотоксичных ТМ представляет свинец. Механизмы токсического действия этого ТМ не вполне ясны [3–6]. Существует мнение [7], что свинец не приводит к значительному росту выработки свободных радикалов, однако вскоре после появления его ионов в среде усиливаются процессы перекисного окисления липидов в клеточных мембранах [8]. В то же время сообщают [9, 10], что введение в пораженный организм соединений-ловушек свободных радикалов и антиоксидантов ослабляет токсический стресс.
Проблема устойчивости растительных организмов к действию ТМ имеет не только теоретическое, но и прикладное значение. Вследствие негативного воздействия содержащихся в окружающей среде ТМ на сельскохозяйственные растения снижается их продуктивность. Это делает актуальными исследования, направленные на углубление понимания механизмов устойчивости к техногенному стрессу сельскохозяйственных культур. Помимо того, поступая в товарную часть растений, ТМ способны оказывать негативное воздействие на здоровье человека и домашних животных. Конечной целью такой работы является уточнение допустимых концентраций ТМ в различных типах почв и разработка методов, обеспечивающих получение качественной и безопасной сельскохозяйственной продукции на загрязненных территориях.
Главным путем, по которому ТМ попадают в организм растения, является поступление из почвенного раствора через корень, поэтому данный орган является первым барьером на пути миграции вредных ионов. Товарная часть редиса (Raphanus sativus L.) – это корнеплоды, которые непосредственно контактируют с почвой поэтому потенциально редис способен накапливать существенные количества ТМ. Ионы ТМ, преодолевшие корневой барьер, через проводящую систему попадают в надземную часть растения, которая является товарной частью салата (Lactuca sativa L.). Принимая во внимание указанные особенности этих овощных культур в отношении использования их товарной части для питания человека они представляют интересный объект для исследования воздействия ТМ на сельскохозяйственные растения, т.к. на их примере можно оценить особенности действия свинца на растения разных семейств их чувствительность к его действию на разных уровнях организации. Цель работы – изучение фитотоксичности ТМ на примере свинца для растений редиса и салата при выращивании их на разных типах почв с использованием биометрических, биохимических показателей и продуктивности.
МЕТОДИКА ИССЛЕДОВАНИЯ
Эксперимент по изучению влияния загрязнения почвы Pb на рост, развитие, биохимические показатели и продуктивность редиса (Raphanus sativus L., сорт Алешка 16-суточный) и салата-латука (Lactuca sativa L., сорт Московский парниковый) был заложен на 2-х типах почв: дерново-подзолистой супесчаной и черноземе мощном деградированном. Растения выращивали в пластиковых сосудах емкостью 5 кг по общепринятой методике [11]. Агрохимические характеристики почв [12, 13], приведены в табл. 1.
Таблица 1.
Физико-химические характеристики почв
| Показатель | Дерново-подзолистая супесчаная | Чернозем мощный деградированный |
|---|---|---|
| pHKCl | 5.27 ± 0.01 | 5.3 ± 0.02 |
| Гумус, % | 1.31 ± 0.02 | 5.07 ± 0.06 |
| Hг, мг-экв/100 г почвы | 1.82 ± 0.03 | 3.80 ± 0.02 |
| Сумма обменных оснований, мг-экв/100 г почвы | 6.05 ± 0.18 | 33.0 ± 0.01 |
| Обменный K2O, мг/кг почвы (по Масловой) | 77.6 ± 3.3 | 103 ± 2 |
| Подвижный P2O5, мг/кг почвы (по Кирсанову) | 65.0 ± 0.6 | 179 ± 4 |
Валовое содержание свинца в интактных почвах составляло в дерново-подзолистой почве и черноземе 8.6 и 10.9 мг/кг почвы соответственно. При закладке опыта почву тщательно перемешивали и вносили в нее питательные элементы в виде водных растворов солей NH4NO3, KCl и KH2PO4 из расчета N200P200K200. При внесении в почву питательных веществ учитывали и корректировали количество азота, поступающее с раствором соли ТМ. Свинец добавляли в дерново-подзолистую почву в виде водного раствора соли Pb(NО3)2 в 6-ти концентрациях (мг Pb/кг почвы): 50, 100, 250, 500, 1000, 1500; в чернозем – 500, 1000, 1500, 2000, 3000, 4000. Контролем служил вариант с NPK без внесения свинца. Опыт проводили в четырехкратной повторности. Перед посевом почву инкубировали в течение 14 сут при температуре 20–23°С и влажности 60% ПВ. Растения выращивали до товарной спелости при температуре 18–20°С в условиях постоянной влажности почвы (60% ПВ). Положение вегетационных сосудов меняли ежедневно по схеме, обеспечивающей однородные условия роста и развития растений. Поливали растения дистиллированной водой [14, 15].
Отбор растительных проб для анализа проводили перед уборкой через 40 сут после всходов. До урожая оставляли по 10 растений на сосуд. В течение всего вегетационного периода вели фенологические наблюдения. При уборке урожая через 38 сут после всходов в пробах определяли биометрические показатели растений: площадь листьев (методом высечек), определяли воздушно-сухую массу корнеплодов (редис) и растений в целом (салат). Биохимические показатели (содержание свободного пролина [16] и малонового диальдегида (МДА) [17]) определяли в отобранных через 38 сут после всходов отдельных пробах листьев редиса и салата. Статистическую обработку экспериментальных данных проводили стандартными методами с использованием MS Excel. На графиках и диаграммах приведены среднеарифметические показатели в расчете на одно растение и доверительные интервалы (в % к контролю) при 95%-ном уровне значимости.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Площадь листьев. Площадь листьев редиса (рис. 1а), выращенного на дерново-подзолистой почве, последовательно увеличивалась до 120% по отношению к контролю при увеличении концентрации свинца до 250 мг/кг почвы, а затем плавно снижалась до 70% при максимальной дозе ТМ 1500 мг/кг. Для площади листьев салата было характерно резкое, до 48% по отношению к контролю снижение площади листьев при действии минимальной концентрации свинца 50 мг/кг почвы. Дальнейшее повышение концентрации свинца вплоть до 1500 мг/кг вызывало небольшие изменения показателя на уровне от 50 до 40% по отношению к контролю.
Рис. 1.
Площадь листьев редиса и салата: (а) – на дерново-подзолистой почве, (б) – на черноземе (линейная аппроксимация дозовой зависимости).
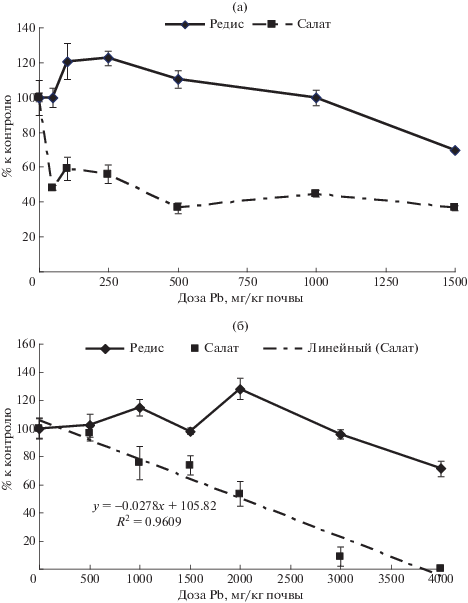
На черноземе у редиса в диапазоне доз 0–2000 мг Pb/кг отмечены слабо выраженные изменения площади листьев почти на уровне контроля, хотя наличествовала тенденция к росту этого показателя. Эта тенденция достигала максимума 128% по сравнению с контролем при концентрации свинца 2000 мг/кг (рис. 1б). При дальнейшем увеличении концентрации токсиканта площадь листьев постепенно снижалась до 70% по отношению к контролю. У салата на черноземе площадь листьев практически линейно снижалась до 13% от уровня контроля при концентрации свинца 3000 мг/кг почвы, что подтверждено высокой достоверностью аппроксимации этой дозовой зависимости линейной функцией: y = 0.0278x + 105.82 с коэффициентом R2 = 0.97 и r = 0.98. При максимальной концентрации свинца 4000 мг/кг наблюдали почти полное подавление развития площади листьев салата.
Биомасса. На дерново-подзолистой почве (рис. 2а) в диапазоне концентраций свинца от 0 до 100 мг/кг биомасса корнеплодов редиса возрастала, достигая уровня 133% от контроля. При повышении концентрации свинца до 250 мг/кг происходило резкое снижение величины показателя до уровня контроля. Далее наблюдали последовательное плавное уменьшение биомассы корнеплодов до 50% по отношению к контролю при дозе 1500 мг Pb/кг.
Рис. 2.
Биомасса салата и корнеплодов редиса: (а) – на дерново-подзолистой почве, (б) – на черноземе.
На черноземе биомасса корнеплодов достоверно снижалась до уровня 81% по отношению к контролю при концентрации свинца 1000 мг/кг почвы (рис. 2б). При дозе ТМ 1500 мг/кг отмечено небольшое недостоверное повышение показателя. Начиная с концентрации 2000 мг/кг и до 3000 мг/кг биомасса корнеплодов снижалась до 40% по отношению к контролю, оставаясь на том же уровне и при максимальной концентрации ТМ 4000 мг/кг. Интересно, что на черноземе практически линейной оказалась концентрационная зависимость для таких разных по физиологическим функциям органам, как корнеплоды редиса и листья салата, из которых у последнего практически полностью состоит биомасса, являющаяся одновременно и товарной частью урожая.
Биохимические параметры. Действие различных негативных факторов вызывает включение у живых организмов многочисленных негативных реакций на разных уровнях организации. Одним из важных типов повреждений является нарушение окислительно-восстановительного гомеостаза. Например, ТМ индуцируют образование перекисных и свободнорадикальных соединений. Реакции структурных и ферментных систем клеток с этими соединениями вызывают обширные повреждения в их структурах и нарушения ферментных циклов метаболизма, объединяемых под общим термином оксидативный стресс. Одним из классов соединений, которые участвуют в ответе организма на эти негативные воздействия, служит ряд аминокислот – γ-аминомасляная кислота, пролин, цистеин [18, 19], среди которых особый интерес вызывает пролин [18] благодаря своей многофункциональности. При стрессовых воздействиях на растения (в том числе и ТМ) в их тканях усиливается образование свободных радикалов, что приводит к интенсификации процессов перекисного окисления липидов (ПОЛ) мембран и нарушению структурно-функционального состояния клеток [18–20]. Конечным продуктом ПОЛ является малоновый диальдегид (МДА), который служит показателем интенсивности окислительных процессов в тканях растений.
Концентрационные зависимости содержания МДА в листьях редиса представлены на рис. 3. На черноземе содержание МДА во всем диапазоне концентраций ТМ достоверно не отличалось от контроля (рис. 3а). По этой причине можно предположить, что действие свинца не вызывало индукции синтеза МДА у редиса на этой почве. Однако достоверное увеличение содержания пролина до уровня 150% по отношению к контролю при концентрации свинца 500 мг/кг и такой же эффект при максимальной концентрации свинца 4000 мг/кг позволило предположить, что отсутствие реакции в содержании МДА вызвано подавлением синтеза последнего за счет индукции синтеза пролина как защитного соединения. В диапазоне концентраций ТМ 1000–3000 мг/кг почвы отсутствие изменений в концентрациях МДА и пролина вызвано равновесием противоположно направленных процессов синтеза повреждающего и защитного соединений.
Рис. 3.
Содержание МДА и пролина в листьях редиса: (а) – на черноземе, (б) – на дерново-подзолистой почве.
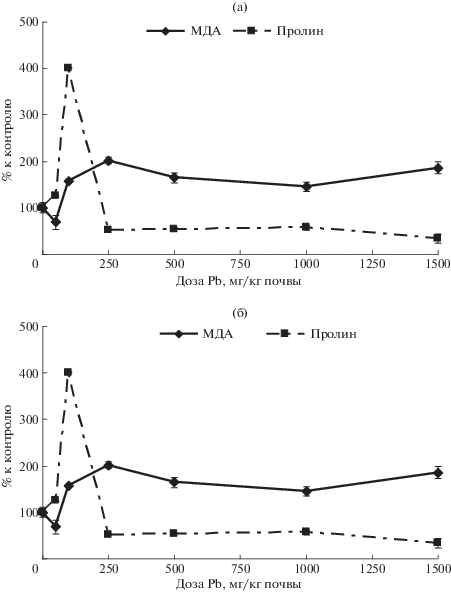
На дерново-подзолистой почве (рис. 3б) в листьях редиса в диапазоне концентраций от контроля до 250 мг/кг наблюдали резкое увеличение содержания МДА в 2 раза по сравнению с контролем. Этот факт свидетельствовал о том, что в диапазоне концентраций свинца 50–250 мг/кг действительно происходили существенные изменения метаболизма в надземных органах редиса в ответ на токсическое действие ТМ, которое вызывает еще более резкое увеличение содержания защитного соединения – свободного пролина, уровень которого повышался в 4 раза по сравнению с контролем. Пролин тормозит процесс перекисной деградации клеточных мембран, поэтому такое резкое усиление синтеза этого соединения способствовало ослаблению оксидативного стресса. Благодаря этому при дальнейшем повышении концентрации свинца до 1000 мг/кг происходило постепенное достоверное снижение содержания МДА до 146% по отношению к контролю. Однако такое повышение синтеза пролина, вероятно, вызвало истощение защитных возможностей организма, в результате чего концентрация пролина упала ниже контроля на 50% и еще больше снизилась при максимальной дозе свинца до 34%. По этой причине снижение уровня МДА сменилось достоверным повышением показателя до 186% при концентрации свинца 1500 мг/кг почвы. Следует отметить, что концентрационная кривая в данном случае напоминала по форме аналогичную кривую по содержанию пролина на этой же почве для салата.
На черноземе (рис. 4а) содержание МДА в листьях салата испытывало разнонаправленные изменения, достоверно не отличаясь от контроля. На дерново-подзолистой почве (рис. 4б) изменения содержания МДА в листьях салата были выражены менее отчетливо, чем в содержании свободного пролина. Вместе с тем формы концентрационных зависимостей в том и другом случае были сходными. Например, в диапазоне концентраций свинца от контроля до 100 мг/кг содержание МДА достоверно снижалось до 86% по отношению к контролю. Затем при концентрации 250 мг/кг оно слегка повышалось, достоверно оставаясь меньше контроля, и опять достоверно снижалось при концентрации ТМ 500 мг/кг до 64% по отношению к контролю. Далее следовал новый подъем содержания МДА до уровня ≈80% по отношению к контролю при концентрациях 1000 и 1500 мг Pb/кг. Такие различия в выраженности реакций различных видов растений по содержанию пролина и МДА, возможно, объясняются тем, что интенсификация синтеза пролина как защитного соединения стимулируется не только усилением перекисного окисления мембран, индикатором которого служит возрастание содержания МДА, но и другими нарушениями в клетках растений, вызванными токсическим действием свинца.
Рис. 4.
Содержание МДА и пролина в листьях салата: (а) – на черноземе, (б) – на дерново-подзолистой почве.

Экспериментальные данные по содержанию пролина в листьях редиса при действии свинца на черноземе представлены на рис. 3а. Концентрационная кривая этого показателя на черноземе демонстрирует практически полное отсутствие зависимости содержания пролина в листьях редиса от концентрации ТМ в диапазоне 1000–3000 мг/кг почвы. Аналогичная концентрационная зависимость имела место и в нашем эксперименте с конскими бобами [21].
На дерново-подзолистой почве обнаружено увеличение содержания свободного пролина в 4 раза по сравнению с контролем при концентрации свинца 100 мг/кг. Дальнейшее возрастание концентрации ТМ, наоборот, привело к резкому снижению содержания этой аминокислоты до уровня 50% по отношению к контролю, причем его содержание осталось практически на этом уровне, вплоть до максимальной концентрации свинца 1500 мг/кг почвы. В наших предыдущих экспериментах с кормовыми бобами [21] такое масштабное повышение содержания пролина в листьях отмечено на этой почве только при максимальных концентрациях ТМ. Обнаруженное явление, возможно, вызвано тем, что у скороспелых культур метаболизм происходит более интенсивно, обеспечивая более интенсивное поглощение ТМ и усиливая, таким образом, токсическое воздействие уже при действии малых его концентраций. Интенсивное поступление свинца в организм растения вызывает в свою очередь стимуляцию синтеза защитного соединения – свободного пролина. С другой стороны, редис и кормовые бобы относятся к растениям с разной стратегией защиты от действия ТМ [22, 23]. Возможно, у бобов имеются механизмы защиты, которые препятствуют поступлению свинца за счет барьера в корневой системе, в результате чего он попадает в надземные органы только тогда, когда этот барьер будет преодолен за счет повышения концентрации ТМ. У редиса этот барьер в корневой системе отсутствует или слабый, и реакция на действие свинца в надземных органах в виде увеличения содержания в листьях редиса свободного пролина возникает уже при малых концентрациях ТМ.
Аналогичное повышение содержания пролина было отмечено на дерново-подзолистой почве также и у салата (рис. 4б), однако этот эффект имел место только при концентрации свинца 250 мг/кг. При дальнейшем увеличении концентрации свинца содержание пролина резко уменьшалось при концентрации ТМ 500 мг/кг до 75% от уровня контроля. Но при концентрациях ТМ 1000 и 1500 мг/кг содержание пролина опять достаточно интенсивно росло, достигая уровня 120% по отношению к контролю. Это свидетельствовало, возможно, о проявлении эффекта 2-го барьера устойчивости, который преодолевался в диапазоне концентраций свинца 500–1000 мг/кг и за которым следовала новая индукция синтеза защитного соединения – пролина.
Содержание пролина в листьях салата на черноземе вплоть до концентрации свинца 3000 мг/кг достоверно не отличалось от контроля, хотя наблюдали пик в содержании МДА и пролина при концентрации свинца 1500 мг/кг. Следует отметить, что концентрационные кривые этих соединений почти сливались до концентрации Pb 3000 мг/кг почвы, что, возможно свидетельствовало о балансе в этом диапазоне процессов повреждения и защиты. Достоверное снижение содержания пролина до 80% по отношению к контролю отмечено только при максимальной концентрации ТМ 4000 мг/кг.
Полученные экспериментальные данные позволили выявить ряд закономерностей при действии разных доз свинца на растения салата и редиса при выращивании их на 2-х почвах, отличающихся разным уровнем плодородия. Показано, что угнетение развития обеих культур по показателям площадь листьев и урожай товарной части сильно зависел от типа почвы (табл. 2). Пороговой концентрацией ТМ для редиса на дерново-подзолистой почве по показателю площадь листьев являлась доза 1500 мг/кг почвы, для салата – 50 мг/кг. Для биомассы товарной части салата пороговая концентрация оказалась также равна 50 мг/кг, т.к. товарной частью салата является фактически масса листьев. Для биомассы корнеплодов редиса пороговой концентрацией оказалась доза 500 мг Pb/кг. На черноземе соответствующие пороговые концентрации составляли для салата 1500 и 1500 мг/кг, для редиса – 3000 и 2000 мг/кг соответственно. Хотя свинец в настоящее время не относится к микроэлементам, однако многие исследователи отмечали стимуляцию ряда показателей разных видов растений при действии определенных концентраций свинца [21]. В наших исследованиях с разными культурами это явление также отмечено [24]. В настоящей работе стимуляция площади листьев и биомассы корнеплодов имела место у редиса на дерново-подзолистой почве при дозах 100–250 мг Pb/кг почвы, стимуляция площади листьев – на черноземе при дозе 2000 мг Pb/кг. При этой же дозе на черноземе у салата имела место стимуляция площади листьев и биомассы.
Таблица 2.
Диапазоны стимулирующих концентраций и пороги фитотоксичности Pb в разных почвах для редиса и салата
| Показатель | Концентрация Pb в почве (мг/кг), при которой наблюдается | |||
|---|---|---|---|---|
| стимуляция растений | угнетение растений | |||
| дерново-подзолистая | чернозем | дерново-подзолистая | чернозем | |
| Редис | ||||
| Биомасса корнеплодов | 100 | нв | >500 | >2000 |
| Площадь листовой поверхности | 100–250 | 2000 | >1500 | >3000 |
| Салат | ||||
| Биомасса растений | нв | нв | >50 | >1500 |
| Площадь листовой поверхности | нв | нв | >50 | >1500 |
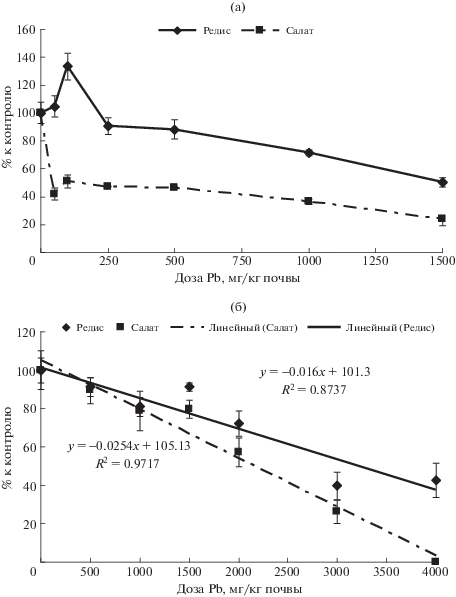
Анализ полученных экспериментальных данных показал, что салат являлся в целом более чувствительным к действию свинца, чем редис (рис. 1). Площадь листьев редиса и салата при действии увеличивающихся концентраций свинца в диапазоне от контроля до 2000–3000 мг Pb/кг на черноземе и торфе претерпевали значительные изменения как в сторону достоверного снижения до 80%, так и повышения до 120% по отношению к контролю. Достоверной пороговой концентрацией ТМ, после которой происходило кардинальное снижение этого показателя, была концентрация 3500 мг Pb/кг почвы (табл. 2). При концентрациях 2000–2500 мг Pb/кг у обеих культур происходило увеличение площади листьев до 120% по отношению к контролю. На дерново-подзолистой почве стимуляция площади листьев редиса до 120% отмечена при концентрациях свинца 100–250 мг Pb/кг. Пороговой, достоверно подавляющей развитие листового аппарата концентрацией Pb была концентрация 1500 мг/кг. Для салата достоверной пороговой концентрацией являлась доза 250 мг/кг, при которой площадь листьев уменьшалась до 56% по отношению к контролю.
Для биомассы корнеплодов редиса пороговой концентрацией свинца на черноземе было 2000 мг/кг, когда показатель снижался до 72% по отношению к контролю. На торфе пороговой концентрацией можно считать 3000 мг/кг, хотя такую концентрацию на этой почве не использовали и ее можно было определить только интерполяцией. На дерново-подзолистой почве пороговой концентрацией свинца, снижающей этот показатель до 71% по отношению к контролю, являлась концентрация 1000 мг Pb/кг. Однако следует отметить, что при концентрации 100 мг Pb/кг отмечено достоверное увеличение биомассы корнеплодов до 133% по отношению к контролю.
Биомасса растений салата на черноземе почти линейно снижалась с повышением концентрации свинца в почве, однако достоверное снижение по сравнению с контролем до 80% было отмечено, начиная с концентрации 1500 мг Pb/кг, которую можно считать пороговой.
На торфе пороговая концентрация свинца для биомассы салата была равна 2500 мг/кг, хотя при концентрации 2000 мг Pb/кг имело место достоверное увеличение биомассы до 130% по отношению к контролю.
Определение содержания МДА и свободного пролина в листьях редиса и салата оказалось наиболее информативным при выращивании этих культур на дерново-подзолистой почве. Для редиса характерна мощная стимуляция синтеза МДА и пролина уже при малых концентрациях свинца в 2.5 (МДА, 250 мг Pb/кг почвы) и 4 раза (пролин, 100 мг Pb/кг) по сравнению с контролем. При этом содержание МДА в листьях редиса при всех остальных концентрациях свинца (250–1500 мг/кг почвы) оставалось достоверно больше контроля, меняясь на уровне 180–250% по отношению к контролю. Содержание свободного пролина, наоборот, при концентрации 250 мг Pb/кг резко уменьшилось до уровня 50% по отношению к контролю и в диапазоне концентраций 250–1500 мг Pb/кг оставалось практически на том же уровне.
В листьях салата содержание МДА в диапазоне концентраций свинца от контроля до 500 мг/кг дерново-подзолистой почвы упало почти до 60% от контроля, затем несколько возросло до 80% при концентрации 1500 мг Pb/кг. Содержание свободного пролина скачкообразно изменялось, то снижаясь почти до 80% при концентрации ТМ 100 мг/кг, то повышаясь почти до 130% и вновь снижаясь до уровня 75% по отношению к контролю. В диапазоне концентраций свинца 500–1500 мг/кг содержание пролина опять повышалось до уровня 120% по отношении к контролю.
ЗАКЛЮЧЕНИЕ
По результатам проведенного исследования выявлено, что свинец оказывал существенное воздействие на жизненные процессы растений редиса и салата. Отмечено, что определенные дозы свинца (100 мг/кг для дерново-подзолистой почвы и 2000 мг/кг для чернозема) могли стимулировать развитие как надземной, так и подземной биомассы редиса. На дерново-подзолистой почве указанный эффект отмечен при более низких дозах поллютанта, чем на черноземе. Однако при высоких дозах ТМ происходило достоверное снижение данного показателя. У салата стимулирующего эффекта ТМ не было обнаружено, по мере роста концентрации свинца происходило практически линейное уменьшение биомассы.
Анализ биохимических параметров выявил следующие эффекты. Наиболее информативные данные удалось получить в эксперименте, проведенном на дерново-подзолистой почве. Для салата был характерен повышенный синтез МДА и пролина при действии свинца, при этом, если в случае с МДА высокое содержание данного метаболита наблюдали во всем диапазоне доз, то содержание пролина вначале резко возрастало (до 400% при 100 мг Pb/кг), но так же резко снижалось при росте концентрации поллютанта, а затем уже не менялось (50% от контроля при 250 мг Pb/кг и далее). У редиса содержание МДА существенно уменьшалось по мере роста концентрации (до 60% от контроля при 500 мг Pb/кг), но затем возрастало (до 77% от контроля при 1500 мг Pb/кг). Что касается пролина, то его содержание изменялось скачкообразно. При различных дозах ТМ отмечены и высокие уровни содержания в тканях растений редиса данной аминокислоты (до 125% при 250 мг Pb/кг и 120% при 1500 мг Pb/кг), и низкие (50% при 500 мг Pb/кг).
По результатам выполненной работы можно сделать вывод о большей чувствительности салата к действию свинца. Следует отметить, что концентрационная зависимость площади листьев салата на черноземе оказалась практически линейной и достоверно аппроксимировалась линейной функцией вида y = –0.0278x + 105.82 при величине достоверности аппроксимации R2 = 0.96 и r = = 0.98. Концентрационная кривая биомассы корнеплодов редиса удовлетворительно аппроксимировалась линейной функцией вида y = –0.016x + + 101.3 при величине достоверности аппроксимации R2 = 0.87, r = –0.94. Еще более надежно аппроксимировалась на этой почве биомасса салата линейной функцией вида y = –0.0254x + 105.13 при величине достоверности аппроксимации R2 = 0.97, и r = –0.99.
Таким образом, в ходе настоящего исследования была изучена зависимость ответа сельскохозяйственных культур на действие техногенных поллютантов от видовой принадлежности этих растений и типа используемого субстрата. Например, листовые культуры (салат) оказались более чувствительными, чем корневые (редис), а на дерново-подзолистой почве токсические эффекты были выражены сильнее, чем на черноземе. В целом зависимость доза–эффект для морфологических показателей растений носила линейный характер, что соответствовало общебиологическим закономерностям, выявленным при токсикологических исследованиях различных объектов. Воздействие техногенного поллютанта (свинца) на биохимические параметры носило несколько иной характер, что свидетельствовало о сложном и неоднозначном характере ответа организма на стресс, наблюдаемый на различных уровнях организации. В работе впервые изучены в широком диапазоне концентраций свинца дозовые зависимости действия этого ТМ на ряд биометрических и биохимических показателей и продуктивность растений салата и редиса при выращивании на 3-х типах почв. Полученные данные о пороговых концентрациях позволили уточнить имеющиеся сведения о ПДК и ПДУ для конкретных сельскохозяйственных культур на различных типах почв. Определены диапазоны стимулирующих концентраций и пороги фитотоксичности Pb в разных почвах для редиса и салата.
Список литературы
Алексахин Р.М., Фесенко С.В., Гераськин С.А. Методика оценки экологических последствий техногенного загрязнения агроэкосистем. М.: Изд-во МГУ, 2004. 206 с.
Ильин В.Б. Тяжелые металлы в системе почва–растение. Новосибирск: Наука, 1991. 150 с.
Гуральчук Ж.З. Механизмы устойчивости растений к действию тяжелых металлов // Физиол. и биохим. культ. раст. 1994. Т. 26. № 2. С. 107–117.
Феник С.И., Трофимяк Т.Б., Блюм Я.Б. Механизмы формирования устойчивости растений к тяжелым металлам // Усп. совр. биол. 1995. Т. 115. № 3. С. 261–276.
Серегин И.В., Иванов В.Б. Физиологические аспекты токсического действия кадмия и свинца на высшие растения // Физиология растений. 2001. Т. 48. № 4. С. 606–630.
Мурзаева С.В. Эффекты тяжелых металлов на проростки пшеницы; активация антиоксидантных энзимов // Прикл. биохим. и микробиол. 2004. Т. 40. № 1. С. 114–119.
Ochi T., Takahashi K., Ohsawa M. Indirect evidence for the induction of a peroxidant state of cadmium chloride in cultured mammal cells and a possible mechanism for the induction // Mutat. Res. 1987. V. 180. P. 257–266.
Muller L. Consequences of cadmium toxicity in rat hepathocytes: mitochondrial dysfunction and lipid peroxidation // Toxicology. 1986. V. 40. P. 285–292.
Farris M.W. Cadmium toxicity: unique cytoprotective properties of alpha tocopheryl succinate in hepathocytes // Toxicology. 1991. V. 69. P. 63–77.
Полесская О.Г. Растительная клетка и активные формы кислорода. М.: Университет, 2007. 139 с.
Журбицкий З.И. Теория и практика вегетационного метода. М.: Наука, 1968. 260 с.
Агрохимические методы исследования почв. М.: Наука, 1975. 656 с.
Аринушкина Е.В. Руководство по химическому анализу почв. М.: Изд-во МГУ, 1970. 488 с.
Методические указания по определению тяжелых металлов в почвах сельхозугодий и продукции растениеводства. Изд-е 2-е. М.: ЦИНАО, 1992. 62 с.
Доспехов Б.А. Методика полевого опыта (с основами статистической обработки результатов исследований). М.: Агропромиздат, 1985. 352 с.
Bates L.S., Waldern R.P., Teare I.D. Rapid determination of free proline for water–stress studies // Plant and Soil. 1973. V. 39. № 1. P. 205–207.
Heath R.L., Packer L. Photoperoxidation in isolated chloroplasts. I. Kinetics and stoichiometry of fatty acid peroxidation // Arch. Biochem. Biophys. 1968. V. 125. № 1. P. 189–198.
Бритиков Е.А. Биологическая роль пролина. М.: Наука, 1975. 124 с.
Baker A.J.M. Accumulators and excluders – strategies in the response of plants to heavy metals // J. Plant Nutr. 1981. V. 3. P. 93–111.
Baker A.J.M. Metal tolerance // New Phytol. 1987. V. 106. P. 93–111.
Арышева С.П., Дикарев В.Г., Гераськин С.А., Дикарева Н.С., Санжарова Н.И., Удалова А.А. Продуктивность, морфологические и физиолого-биохимические показатели бобов кормовых, выращенных на загрязненной свинцом почве // Агрохимия. 2013. № 2. С. 77–85.
Кузнецов В.В., Шевякова Н.И. Пролин при стрессе: биологическая роль, метаболизм, регуляция // Физиология растений. 1999. Т. 46. № 2. С. 321–336.
Барабой В.А. Механизмы стресса и перекисное окисление липидов // Усп. совр. биол. 1991. Т. 111 (6). С. 923–931.
Дикарев А.В., Дикарев В.Г., Дикарева Н.С. Сравнительный анализ частоты цитогенетических эффектов в апикальной меристеме корешков проростков сортов ярового ячменя (Hordeum vulgare L.), контрастных по устойчивости к свинцу // Тр. по прикл. бот., генет. и селекции. 2016. Т. 177. Вып. 1. С. 52–68.
Дополнительные материалы отсутствуют.