

Акустический журнал, 2019, T. 65, № 1, стр. 82-92
Акустика дельфинов и рыб (обзор)
Е. В. Романенко *
a Институт проблем экологии и эволюции им. А.Н. Северцова РАН
119071 Москва, Ленинский просп. 33, Россия
* E-mail: evromanenko33@mail.ru
Поступила в редакцию 12.03.2018
После доработки 28.08.2018
Принята к публикации 06.07.2018
Аннотация
Обзор посвящен анализу исследований и разработок, связанных с акустикой дельфинов и рыб. Основное внимание уделено структуре эхолокационных и коммуникационных звуков дельфинов в условиях воздействия интенсивных шумовых помех. Звуки регистрировали и излучали непосредственно на голове дельфина в точках предположительного входа акустической информации. Исследованы звуки внутри носового прохода с целью более точной локализации источников излучения. Звуки рыб регистрировали в водоеме, в котором они находились. Анализ показал, что дельфины способны целенаправленно изменять структуру излучаемых звуковых сигналов под воздействием предельно интенсивных шумовых помех с целью повышения их помехоустойчивости. Высказаны предположения о механизме излучения звуков рыбами.
ВВЕДЕНИЕ
Механизмы излучения и приема звуков дельфинами представляют собой наиболее трудные проблемы акустики китообразных. При решении проблемы излучения исследователи пока вынуждены пользоваться лишь косвенными данными, получаемыми из анализа звуковых сигналов дельфинов, зарегистрированных в дальнем звуковом поле, и в результате морфологических исследований. О природе источника звука в этом случае и месте его расположения можно судить только предположительно и неоднозначно.
В относительно небольшом обзоре [1] проведен анализ ряда экспериментальных исследований и показано, что:
– интенсивность эхолокационного сигнала играет доминирующую роль в определении его характеристик;
– сигналы свободно плавающего животного могут отличаться от сигнала неподвижного животного;
– на характеристики направленности сигнала в большей степени влияют челюсть и воздушные мешки, в меньшей степени влияет мелон;
– излучение импульсных звуков исходит из правого носового прохода, а свисты излучаются левым проходом – последний результат согласуется с опубликованными ранее данными работы [2].
В работе [3] экспериментально определен частотный диапазон дельфина (Tursiops truncatus), используемый им для распознавания металлических мишеней сферической формы (стальных и латунных диаметром 7.6 см). Показано, что для целей распознавания таких мишеней дельфин использует частоты между 29 и 42 кГц.
В работе [4] экспериментально показано распределение эхолокационного поля в окрестности головы дельфина. Измерения проведены с помощью нескольких миниатюрных гидрофонов, закрепленных непосредственно на голове животного с помощью присосок. Определена зона максимальной амплитуды сигнала. Эксперимент проведен на двух животных. Полученные результаты хорошо согласуются. Обнаружена асимметрия распределения амплитуд, которая объясняется асимметрией структур, ответственных за процесс излучения звука. Асимметрия неодинакова на правой и левой стороне головы, она заметнее на правой стороне.
В работе [5] проведены экспериментальные исследования по определению диаграмм направленности звукового излучения дельфина (Tursiops truncatus). Измерения проведены в вертикальной и горизонтальной плоскостях. Расстояние от головы дельфина до решетки гидрофонов около 1.5 метров. Все гидрофоны были расположены на глубине 1 м при измерениях в горизонтальной плоскости. При измерениях в вертикальной плоскости на глубине 1 м располагали центральный гидрофон (под углом 0° к горизонтальной оси животного).
Следует подчеркнуть, что большинство проанализированных работ в значительной степени повторяют исследования, проведенные в России много лет назад [6]. Слабую информированность зарубежных исследователей о работах, проводившихся в России, следует отнести прежде всего на счет языкового барьера.
Опубликовано несколько крупных обзорных работ по акустике китообразных [7–11]. Однако все эти работы грешат одним существенным недостатком. Ни в одной из них не ставилась задача исследования акустики дельфинов в экстремальных условиях, когда животное вынуждено было бы мобилизовать все свои способности для решения эхолокационной задачи. Акустические способности дельфинов изучали в спокойной обстановке и, как правило, в дальней зоне эхолокации. В результате зачастую делались неверные выводы. Например, в работе [7] автор делает следующий вывод: “В результате мы пришли к мнению, что у дельфинов в условиях бассейна отсутствует целенаправленная подстройка спектрального состава эхолокационного импульса”. Как будет показано далее, это совсем не так.
Места входа акустической информации на голове дельфина также до сих пор являются предметом спора. Помимо очевидного предположения о том, что акустическая информация поступает к внутреннему уху дельфина через наружные слуховые проходы, существуют и другие.
В работе [12] выдвинуто предположение о том, что у китообразных звук к улитке поступает через нижнюю челюсть. В работе [13] нижняя челюсть рассматривается как катеноидальный рупор, заполненный жировым тяжем, в передней части которого находятся отверстия, служащие входом для отраженных от подводных целей эхолокационных сигналов. В результате анализа работы рупора Рябовым делается вывод о несостоятельности существующих гипотез о важной роли наружных слуховых проходов и “акустического окна” нижней челюсти. Вывод о несостоятельности гипотезы о роли слуховых проходов следует считать неубедительным, так как существуют экспериментальные данные [14], подтверждающие важную роль слуховых проходов в проведении звука к внутреннему уху дельфина. В работе [15] показано наличие приемной точки (акустического окна) на уровне слухового прохода, хотя точную локализацию места входа акустической информации установить не удалось. В работе [15] допускается существование по крайней мере двух входов – нижняя челюсть и слуховой проход. В работе [16] предполагается, что важную роль в проведении звука к улитке играет лобный выступ.
Более достоверные сведения о механизме излучения и приема звука могли бы быть получены при инструментальном проникновении приемника звука в дыхательную систему дельфина. Однако это исключительно трудная задача, требующая разработки специальных измерительных приборов и отработки соответствующей методики. Тем не менее, такие попытки делаются. Первые успешные попытки приблизиться к источнику звука дельфина предприняты в работах [6, 17] и заключаются в регистрации звука непосредственно на голове животного. Этот метод оказался весьма плодотворным и уже позволил установить ряд интересных особенностей излучения звука дельфином. В работе [18] проведено экспериментальное исследование структур, ответственных за генерацию эхолокационных импульсов и свистовых сигналов. Исследование проводилось с помощью двух видеокамер, снабженных тонкими щупами (стекловолокно), и миниатюрных гидрофонов. Щупы видеокамер и гидрофоны вводили в носовые проходы и гортань. С их помощью наблюдали вибрации носовых пробок в процессе эхолокации и регистрировали эхолокационные и свистовые сигналы. Сделан вывод о способности дельфинов излучать одновременно разные сигналы от двух звуковых источников, которые могут быть синхронными и несинхронными.
Однако возможности метода оказались значительно шире в результате создания специального комплекса приборов, закрепляемого на дельфине и позволяющего всесторонне изучить его акустическую систему [6, 19]. В состав комплекса входят следующие приборы: трехканальный широкополосный магнитофон, три миниатюрных гидрофона, два независимых генератора шума, несколько излучателей шума, система дистанционного управления работой магнитофона и генераторов шума, трассер, позволяющий визуально прослеживать движение дельфина в полной темноте. Такая аппаратура позволяет изучать эхолокационное поведение дельфина в ситуациях, осложненных наличием искусственно создаваемой помехи. В настоящей работе будут изложены некоторые результаты исследования акустической системы черноморской афалины (Tursiops truncatus) с помощью такого комплекса приборов.
ВЛИЯНИЕ ШИРОКОПОЛОСНОГО КОРРЕЛИРОВАННОГО АКУСТИЧЕСКОГО ШУМА НА ЭХОЛОКАЦИОННОЕ ПОВЕДЕНИЕ ДЕЛЬФИНА
В экспериментах [20, 21] исследовано влияние интенсивного широкополосного коррелированного акустического шума на эхолокационную активность в процессе решения дельфином задачи по обнаружению предъявляемой ему рыбы в бассейне в полной темноте. Шум предъявляли взрослому дельфину локально на различных участках поверхности головы (у предполагаемых акустических входов слуховой системы: у наружных слуховых проходов, на нижней челюсти и лобном выступе). Коррелированный шум – это шум, излучаемый двумя или бóльшим количеством излучателей, питаемых от одного и того же генератора. В основу экспериментов была положена идея, согласно которой возможности эхолокационного аппарата у дельфинов должны наиболее полно проявляться лишь в исключительно сложных условиях, когда локация затруднена, но является единственным средством обнаружения. В работе [20] эксперимент состоял из двух этапов.
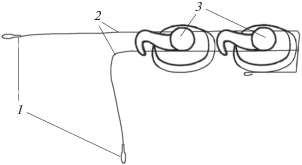
На первом этапе два излучателя коррелированного шума (сферы из пьезокерамики цирконата-титаната свинца ЦТС диаметром 30 мм) были закреплены у правого и левого наружных слуховых проходов (0 на рис. 1а и 1б). Три гидрофона размещены следующим образом. Гидрофон 1, представляющий собой сферу из пьезокерамики диаметром 15 мм, закреплен рядом с излучателем 0 в области правого наружного слухового прохода. Гидрофон 2, представляющий собой пьезокерамический цилиндр диаметром 2 мм и высотой 3 мм, закреплен на правой части лобного выступа, приблизительно на одинаковом расстоянии от дыхала и переднего конца лобного выступа. Гидрофон 3, точно такой же, как гидрофон 2, расположен на переднем конце лобного выступа в том месте, где он переходит в рострум. Излучатели подключены к генератору шума, а гидрофоны – ко входам трехканального магнитофона. Генератор шума и магнитофон закреплены на спинном плавнике дельфина. Управление включением генератора шума и магнитофона осуществляли по подводному радиоканалу с помощью командных радиосигналов длительностью около 1 с.
Рис. 1.
Расположение излучателей шума 0 и гидрофонов 1–3 на голове дельфина: (а) и (б) разные экспериментальные варианты.
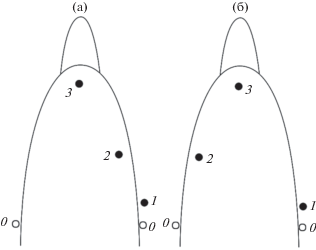
На втором этапе эксперимента два излучателя коррелированного шума были расположены на нижней челюсти животного слева и справа, третий излучатель – на лобном выступе, точно посередине между дыхалом и рострумом. Излучатели идентичны тем, которые были использованы на первом этапе эксперимента. Гидрофоны расположены так, как показано на рис. 1, и также идентичны использованным на первом этапе эксперимента. В процессе эксперимента дельфин находился в бассейне размером 12.5 × 6 м при глубине 1.2 м. Стартовая позиция дельфина находилась у одной короткой стороны бассейна. У другой короткой стенки бассейна (в одном из углов) предъявляли рыбу на тонкой нити с громким плеском, который служил стартовым сигналом. С этого момента дельфин начинал движение к рыбе, которую в это время бесшумно передвигали на 2.5–3 м в сторону вдоль короткой стенки бассейна. Так как эксперимент проводили в полной темноте, дельфин вынужден был практически непрерывно пользоваться эхолокацией. Расстояние между стартовой позицией дельфина и местом предъявления рыбы составляло 9–10 м. Приблизительно на половине пути дельфина между стартовой позицией и местом предъявления рыбы включали широкополосный акустический шум, излучаемый упоминавшимися выше излучателями. Спектр шума, постоянный по уровню в диапазоне частот от 5 до 30 кГц, на более высоких частотах спадал по закону 6–7 дБ на октаву. Уровень шума по давлению в общей полосе частот составлял около 120 ± 6 дБ относительно 1 Па. Реакцию дельфина на включение шума определяли двумя способами: во-первых, визуально, по изменению траектории светящегося трассера, во-вторых, по изменению эхолокационной активности. На первом этапе эксперимента, когда шум предъявляли в области наружных слуховых проходов, наблюдалась четкая двигательная реакция в виде резкого изменения траектории трассера. Дельфин вздрагивал при включении шума. Иногда такие эксперименты проводили в дневное время (при хорошем освещении) и было отчетливо видно, как при включении шума дельфин в первые секунды резко двигал головой, пытаясь освободиться от мешающего действия шума, но затем быстро успокаивался. На втором этапе эксперимента, когда шум предъявляли в области нижней челюсти и лобного выступа, никакой двигательной реакции отмечено не было.
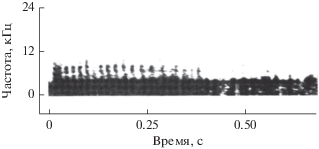
Аналогично обстояло дело и с эхолокационной активностью. В отсутствие шума дельфин пользовался для обнаружения подводных предметов, в том числе и рыбы, главным образом стереотипными импульсами, подобными изображенным на рис. 2а.
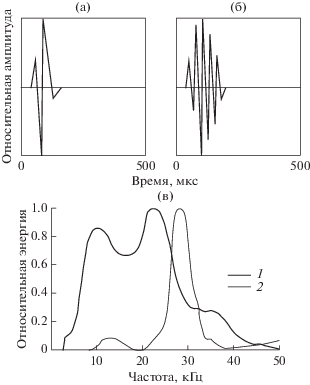
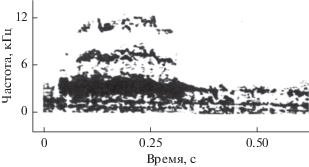
Форма импульсов может варьировать в некоторых пределах [20], но в среднем все импульсы близки по форме и имеют широкий спектр. Энергетический спектр этих импульсов представлен на рис. 2в (кривая 1). В литературе такая форма импульса считается очень устойчивой по отношению к меняющимся условиям эксперимента, в частности к появлению помехи, излучаемой отдаленными источниками [7]. Эксперименты с предъявлением коррелированного шума в область наружных слуховых проходов дали совершенно неожиданный результат. В момент включения шума дельфин осуществляет в течение весьма короткого времени (200–300 мс) значительную перестройку спектра импульсов в плане значительного его сужения. Импульс преобразуется из стереотипного или близкого к таковому в колебательный (рис. 2б), а его спектр становится узкополосным, как показано на рис. 2в (кривая 2). Такая перестройка систематически регистрируется гидрофоном 3, установленным вблизи рострума, при предъявлении шума в области слуховых проходов.
Еще более неожиданным оказался тот факт, что импульсы, регистрируемые гидрофоном 2 (см. рис. 1), остаются стереотипными, несмотря на то, что идут синхронно с импульсами, регистрируемыми гидрофоном 3. Однако стереотипные импульсы, регистрируемые гидрофоном 2, при включении шума значительно возрастают по амплитуде (в 2.5–3 раза), тогда как импульсы, регистрируемые гидрофоном 3, изменяясь по спектру, остаются неизменными по амплитуде. Иногда гидрофон 2, закрепленный сбоку от лобного выступа дельфина, также регистрировал колебательные импульсы. В частности, была зарегистрирована очень интересная эхолокационная серия, состоящая из чередующихся стереотипных и колебательных импульсов, разделенных интервалом времени в несколько миллисекунд.
Эту серию можно рассматривать как результат наложения двух серий, излученных одновременно и состоящих одна из стереотипных импульсов, а другая – из колебательных. Отметим, что на осциллограммах импульсов, зарегистрированных при наличии шума, сам шум не просматривается, поскольку его излучение производилось вблизи наружных слуховых проходов, а регистрация эхолокационных сигналов – на лобном выступе. При этом вблизи точки регистрации отношение амплитуды сигналов к среднеквадратичному значению шума было не менее 40 дБ. Еще один интересный факт заключается в том, что импульсы, излучаемые дельфином при наличии шума, иногда имеют “предвестники”, отмечавшиеся ранее [7, 22]. Правда, достоверно установлено пока наличие перед импульсами только по одному предвестнику. Величина предвестника изменяется в широких пределах и иногда может достигать величины основного импульса: импульс становится двойным. Однако длительность предвестника при этом остается несколько меньше длительности основного импульса. Осциллограммы одного и того же импульса с предвестником, зарегистрированные гидрофонами 2 и 3 в присутствии шума, показаны на рис. 3а и 3б. Их энергетические спектры изображены на рис. 3в. Отличительной особенностью этих спектров является их изрезанность. Функциональное назначение предвестников и изрезанность спектра, соответствующая их наличию, пока неясны. Гидрофон 1, расположенный у слухового прохода, никогда не регистрировал излучения колебательных импульсов. Вместо них он в те же моменты времени регистрировал импульсы, близкие по форме к стереотипным. Колебательные импульсы приходят к гидрофону у слухового прохода лишь в виде отраженных от расположенных впереди объектов.
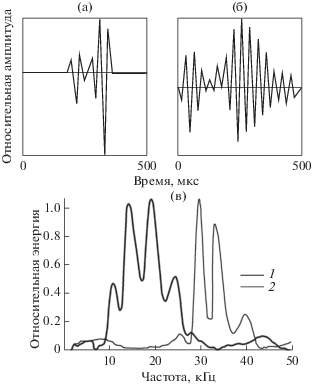
На втором этапе эксперимента, когда излучатели шума были закреплены на нижней челюсти и лобном выступе дельфина, практически никаких значительных изменений в эхолокационной активности отмечено не было. Импульсы как до включения шума, так и после мало отличались от стереотипных (в точках расположения гидрофонов 2 и 3). Можно лишь констатировать, что дельфин слышал включаемый шум: отмечалось незначительное увеличение амплитуды импульсов в точках расположения всех трех гидрофонов. При этом заметных изменений формы и частоты следования импульсов не наблюдалось.
ВЛИЯНИЕ ШИРОКОПОЛОСНОГО НЕКОРРЕЛИРОВАННОГО АКУСТИЧЕСКОГО ШУМА НА ЭХОЛОКАЦИОННОЕ ПОВЕДЕНИЕ ДЕЛЬФИНА
В этом эксперименте использовался другой, но тоже взрослый дельфин. На этот раз задача дельфина по обнаружению рыбы была более сложной. Для того чтобы максимально затруднить эхолокационное обнаружение, вблизи каждого слухового прохода при помощи отдельных шумовых генераторов излучалось интенсивное поле некоррелированного широкополосного акустического шума с уровнем около 130 дБ относительно 1 Па в общей полосе частот. Некоррелированный шум – это шум, излучаемый несколькими излучателями, питаемыми от разных генераторов. В этом случае уровень шума был всего на 6–9 дБ ниже предельного уровня, при котором дельфин отказывался лоцировать рыбу. Спектр шума был равномерным в диапазоне частот от 8 до 50 кГц и затем возрастал на 6–8 дБ на октаву вплоть до частоты 180 кГц. Схема размещения гидрофонов на голове дельфина показана на рис. 1б. Основные результаты можно сформулировать следующим образом: в эксперименте с некоррелированным шумом не отмечена такая четкая зависимость частотно-временных характеристик эхолокационных импульсов дельфина от наличия или отсутствия шума, как в эксперименте с коррелированным шумом. Дельфин в большинстве случаев предпочитал пользоваться колебательными импульсами. Создается впечатление, что, будучи в первый раз (при первом включении) напуган интенсивным некоррелированным шумом, дельфин даже в промежутках времени между включениями шума продолжал пользоваться колебательными импульсами [20]. И лишь время от времени дельфин перемежал колебательные импульсы стереотипными даже в пределах одной и той же серии. Другой характерной особенностью акустического поведения дельфина оказалось то, что колебательные импульсы в этом случае содержали, как правило, 5–10 периодов. Часто можно наблюдать плавный переход от излучения стереотипных импульсов к излучению колебательных. На рис. 4а–4д показан такой переход. Приведенные импульсы зарегистрированы гидрофоном вблизи рострума. На рис. 4е–4к приведены энергетические спектры показанных импульсов.
Рис. 4.
Переход от стереотипных импульсов к колебательным в пределах одной серии: (а–д) импульсы серии, (е–к) их частотные спектры.
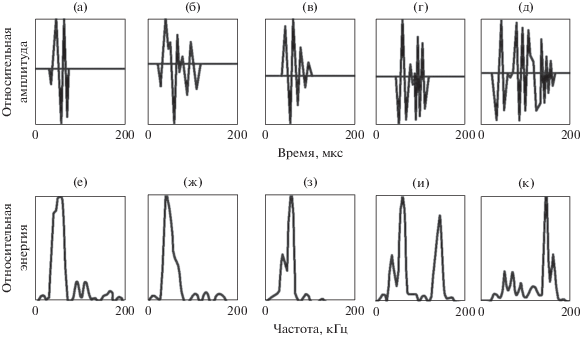
Рассмотрим более подробно импульс, показанный на рис. 4г. В этом импульсе особенно четко заметны две особенности. Первая состоит в том, что импульс начинается с фрагмента (предвестника), очень напоминающего стереотипный импульс. Анализ большого числа импульсов показал, что практически все они начинаются с такого фрагмента (такой же фрагмент виден и в импульсе на рис. 4д, хотя здесь он выражен менее отчетливо). Вторая особенность импульса состоит в том, что его более высокочастотная часть заметно разделена на две секции. Такое разделение, по-видимому, не случайно, так как встречаются серии из нескольких десятков импульсов, имеющих низкочастотный фрагмент (предвестник) и разделенных на секции, которых может быть две или три. Значительно реже встречаются импульсы, не разделенные на секции. Так же как и в эксперименте с коррелированным шумом, гидрофон, расположенный у слухового прохода, никогда не регистрировал излучения колебательных импульсов. Вместо них он в те же моменты времени регистрировал импульсы, близкие по форме к стереотипным. Колебательные импульсы приходят к гидрофону у слухового прохода лишь в виде отраженных от расположенных впереди объектов. Гидрофон, расположенный сбоку лобного выступа, в эксперименте с некоррелированным шумом практически всегда регистрировал колебательные импульсы одновременно с гидрофоном, расположенным вблизи рострума, тогда как в эксперименте с коррелированным шумом такие случаи бывали очень редко – боковой гидрофон регистрировал почти всегда стереотипные импульсы. Приведенные результаты позволяют сделать следующие выводы.
1. Дельфины афалины способны к целенаправленному изменению частотно-временных характеристик эхолокационных импульсов в условиях предъявления им весьма интенсивных широкополосных шумов в области наружных слуховых проходов.
2. Не исключено, что у дельфинов существуют различные источники излучения стереотипных и колебательных импульсов.
3. Акустическая информация поступает во внутреннее ухо слуховой системы дельфина через наружные слуховые проходы.
4. Система излучения колебательных импульсов имеет низкую добротность и работает в вынужденном режиме.
ЗВУКИ ВНУТРИ ДЫХАТЕЛЬНОЙ СИСТЕМЫ ЧЕРНОМОРСКОЙ АФАЛИНЫ
Описанный выше комплекс приборов позволяет проникнуть внутрь дыхательной системы дельфина и зарегистрировать звуки, сопровождающие процесс излучения эхолокационных импульсов. Такая информация весьма полезна при оценке гипотезы о пневматической природе излучаемых сигналов дельфина [23].
Первые опыты по введению регистрирующих устройств в дыхательную систему дельфинов были осуществлены в 1988 году [24]. Авторы вводили в носовой проход (через дыхало) дельфина Delphinapterus leucas три инструмента: два полых катетера (диаметром 3 и 1 мм) для измерения давления воздуха и катетер с приемником звука. Инструменты вводили в дыхало на глубину 20 см.
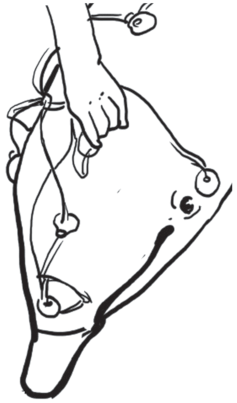
В экспериментах [25] для регистрации звука внутри дыхательной системы дельфина (Tursiops truncatus) были использованы миниатюрные широкополосные гидрофоны (описанные выше гидрофоны 2 и 3), закрепленные на специальном держателе, показанном на рис. 5. Гидрофоны 1 фиксируются в необходимом положении с помощью жестких держателей 2 из проволоки (нержавеющая сталь) диаметром 1 мм. Держатели в свою очередь закреплены на двух присосках 3, предназначенных для фиксации всего устройства на голове дельфина. Устройство, показанное на рис. 5, предназначено для введения в дыхало дельфина только одного гидрофона (его держатель изогнут под прямым углом). Другой гидрофон при этом располагается над поверхностью головы животного впереди дыхала.
В описываемом эксперименте гидрофон вводили в дыхало дельфина на глубину 6 см. Процесс введения гидрофона в дыхало, по-видимому, не вызывал у дельфина значительных неприятных ощущений. Во всяком случае, после двух–трех попыток вытолкнуть гидрофон из дыхала потоком воздуха дельфин смирялся и начинал охотно брать рыбу и лоцировать ее. На рис. 6 схематически показан процесс введения гидрофона в дыхало дельфина. На том же рисунке видны излучатели шума и другие гидрофоны, закрепленные на голове животного [20, 25].
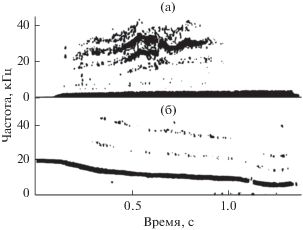
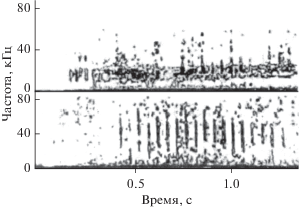
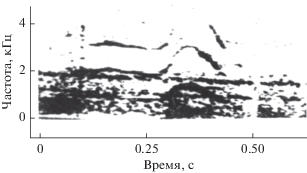
Миниатюрные гидрофоны вводили попеременно в правый и левый носовой проходы. В периоды активной эхолокации звуковые импульсы внутри носового прохода либо отсутствуют, либо слышны существенно слабее, чем снаружи. Из этого можно сделать вывод, что гортань, по-видимому, не является источником эхолокационных сигналов. Кроме того, в правом носовом проходе зарегистрированы звуки, не слышные снаружи и сопровождающие процесс эхолокации. Эти звуки имеют характер свистов (или мяуканья) и по своей структуре значительно отличаются от коммуникационных свистов. Это, как правило, узкополосные звуки, иногда с четко выраженной амплитудной модуляцией, характеризующейся боковыми частотами, не кратными основной частоте (рис. 7а). В коммуникационных свистах обычно присутствуют кратные гармоники (рис. 7б).
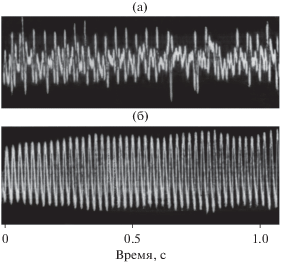
На рис. 8а и 8б показаны осциллограммы внутреннего звука и коммуникационного свиста. Видно, что коммуникационный свист имеет в значительной степени гармонический характер, тогда как внутренний звук выглядит последовательностью одно- и двухпериодных импульсов, которую можно трактовать как сигнал с амплитудной перемодуляцией [13, 18, 19].
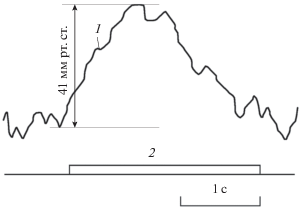
На рис. 9а и 9б приведены сонограммы соответственно внутреннего звука на фоне слабозаметной серии импульсов (внутри дыхала) и той же серии эхолокационных импульсов, зарегистрированной на голове дельфина вблизи рострума. Следует еще раз подчеркнуть, что все описанные выше внутренние звуки зарегистрированы в правом носовом проходе. В левом носовом проходе никаких звуков не обнаружено. Немногочисленные имеющиеся данные позволяют сделать лишь предварительный вывод о том, что область левого носового прохода ниже мускульной пробки не имеет отношения к эхолокации, тогда как правый носовой проход, несомненно, имеет. Практически все внутренние звуки по времени совпадают с процессом эхолокации. Лишь в двух случаях из 17 внутренние звуки не сопровождались эхолокацией (возможно, она была очень слабой и не прослушивалась). Еще в двух случаях из тех же 17 во время эхолокации не прослушивались внутренние звуки (возможно, они были очень слабыми). Длительность внутренних звуков не совпадает с длительностью эхолокационных серий – она, как правило, меньше. Не исключено, что внутренние звуки имеют аэродинамическую природу и сопровождают процесс передувания воздуха в структурах, ответственных за генерацию эхолокационных звуков. Возможно и другое объяснение. Внутренние звуки могут свидетельствовать о значительной разнице давления по разные стороны мускульной пробки и генерируются, когда воздух травится. Это объяснение подтверждается измерениями давления воздуха в дыхательной системе дельфинов [18], как показано на рис. 10. На этом рисунке давление воздуха 1 в носовом канале между внутренней мускульной пробкой и наружным клапаном возрастает в процессе эхолокации 2 на 30–50 мм рт. ст. Дальнейшие исследования должны пролить свет на природу внутренних звуков. В настоящее же время наличие этих звуков свидетельствует в пользу гипотезы о пневматическом механизме генерации звуков дельфинами.
АКУСТИКА РЫБ
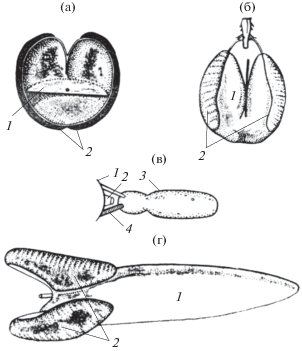
Исследования проводились на морских и пресноводных рыбах. Регистрацию звуков осуществляли в диапазоне частот от 50 до 10 000 Гц. Полное описание звуков исследованных рыб дано в работах [26–28, 30, 31] а также в Атласе звуков рыб [29]. Подробный анализ сигнального значения звуков рыб и некоторых механизмов их излучения дан в монографии [30]. Значительная часть звуков излучается рыбами с помощью плавательного пузыря. Плавательный пузырь, представляющий собой воздушную полость той или иной конфигурации, возбуждается с помощью специальных мышц. При этом плавательный пузырь работает как резонатор. На рис. 11 показаны плавательные пузыри нескольких видов рыб. Видно, что форма пузыря может быть довольно причудливой. Однако форма практически не влияет на его резонансную частоту, а определяет в основном только его добротность. Резонансная же частота определяется его объемом.
Рис. 11.
Плавательные пузыри некоторых рыб. (а) Opsanus tau: 1 – диафрагма с центральным отверстием, 2 – барабанные мускулы; (б) Macrurus berglax: 1 − плавательный пузырь, 2 – барабанные мускулы; (в) Theraponth (схема): 1 – череп, 2 – первый позвонок, 3 – плавательный пузырь, 4 – мышцы; (г) Роrichtys notatus: 1 − плавательный пузырь, 2 – барабанные мускулы.
Если деформированный пузырь характеризовать радиусом R сферы эквивалентного объема, то частота основного резонанса может быть оценена по формуле
Наличие высокочастотных компонент в спектрах звуков рыб можно объяснить, если вспомнить, что воздушную полость в воде можно рассматривать не только как систему с сосредоточенными параметрами, но и как систему с распределенными параметрами, т.е. необходимо учесть резонансные колебания воздушного объема внутри пузыря, как в жестких границах. При этом необходимо учитывать, что плавательный пузырь белуги имеет форму, близкую к цилиндрической.
Вероятнее всего, что свистовой и импульсный сигналы с резонансом на 4 кГц излучаются в результате радиальных колебаний плавательного пузыря. Об этом свидетельствует соотношение обертонов и основного тона, которое с точностью 2% совпадает с расчетным. Вычисления показывают, что радиус цилиндрического плавательного пузыря белуги должен иметь величину около 2.4 см. Свистовой сигнал с резонансом на частоте 1 кГц излучается, вероятнее всего, в результате продольных колебаний воздуха в плавательном пузыре. Об этом свидетельствует соотношение обертонов, которое представляет собой гармонический ряд. Оценка показывает, что при этом длина плавательного пузыря должна быть около 17–20 см (размер экземпляра белуги, звуки которой приведены на рисунках, около 80 см).
В заключение следует сказать несколько слов о некоторых особенностях регистрации звуков рыб в связи с их электрическими свойствами. В середине прошлого века широкое развитие получили исследования электрических свойств рыб. При этом обнаружено [31], что импульсам электролокации, как правило, сопутствуют акустические импульсы. Форма и спектры электрических и акустических импульсов могут существенно отличаться друг от друга, в то время как раздельный прием этих импульсов представляет трудную задачу. Это обстоятельство нужно иметь в виду при изучении биоакустики рыб. Не исключено, что в том обширном фактическом материале, который уже накопился в мировой литературе по звукам рыб, значительную долю составляют сведения об электрических сигналах рыб, принимавшихся за акустические. О необходимости внимательного анализа регистрируемых акустических сигналов рыб именно с точки зрения учета возможности одновременной записи и электрических сигналов напоминал В.Р. Протасов [31], один из крупнейших исследователей биоакустики и электрических свойств рыб.
ЗАКЛЮЧЕНИЕ
Как показал обзор исследований акустики дельфинов, большинство работ не отражает всех их способностей в эхолокации подводных объектов. Было показано, что дельфины способны к целенаправленному изменению спектра эхолокационных импульсов, если воздействующая на орган слуха шумовая помеха по уровню близка к предельной, при которой дельфин отказывается лоцировать подводный объект. Это свидетельствует о том, что эхолокационный аппарат дельфина еще до конца не изучен и нуждается в дальнейшем исследовании.
Что же касается исследований звуков рыб, то необходимо принимать некоторые меры в тех случаях, когда есть опасность одновременной регистрации с помощью гидрофонов акустических и электрических сигналов рыб. Прежде всего, это тщательная электрическая экранировка гидрофона и регистрирующей аппаратуры. Однако такая мера не всегда может быть эффективной (в частности, при слабых акустических и сильных электрических излучениях). В таких случаях можно рекомендовать прием сигналов рыб одновременно на два приемника – акустический и электрический, максимально удаленные от источника сигналов. Это позволит, во-первых, разделить по времени принимаемые акустический и электрический сигналы и, во-вторых, сравнить по форме и спектру сигналы, принятые гидрофоном и электроприемником. Такое сравнение позволит сделать вывод о существовании одновременно акустического и электрического сигналов или только одного из них.
Список литературы
Au Whitlow W.L., Chong Wei A review of new understanding of the role of individual structure within the head of dolphins in formation of biosonar signal and beam // Oceanography & Fisheries. 2017. V. 2. № 1. P. 1–8.
Romanenko E.V. Sounds inside the respiration system of Tursiops truncates // In: Sensory Systems of Aquatic Mammals. Ed.: Kastelein R.A., Thomas J.A., Nachtigall P.E. De Spil Publishers, Woerden, The Netherlands, 1995. P. 157–162.
Stuart D.I., Au W.W.L., Paul E.N., Breese M. Functional bandwidth of an echolocating Atlantic bottlenose dolphin (Tursiops truncates) // J. Acoust. Soc. Am. 2009. V. 125. № 2. P. 1214–1221.
Au W.W.L., Houser D.S., Finneran J.J., Lee Wu-Jung, Talmadge L.A., Moore P.W. The acoustic field on the forehead of echolocating Atlantic bottlenose dolphins (Tursiops truncates) // J. Acoust. Soc. Am. 2010. V. 128. № 3. P. 1426–1434.
Au W.W.L., Branstetter B., Moore P.W., Finneran J.J. The biosonar field around an Atlantic bottlenose dolphin (Tursiops truncatus) // J. Acoust. Soc. Am. 2012. V. 131. № 1. P. 569–576.
Романенко Е.В. Физические основы биоакустики. М.: Наука, 1974. 180 с.
Дубровский Н.А. Эхолокация у дельфинов: (Обзор). JI.: ЦНИИ “Румб”, 1975. 78 с.
Animal Sonar. Processes and Performance / Eds. Nachtigall P.E. and Moore P.W. NATO ASi Series. Series A: Life sciences, 1988. V. 156. 862 p.
Sensory Abilities of Cetaceans. Laboratory and Field Evidence / Eds. Thomas J.A. and Kastelein R.A. NATO ASi Series. Series A: Life Sciences, 1990. V. 196. 710 p.
Kamminga C. Research on dolphin sounds. Delft: Delft University of Technology, Faculty of Electrical Engineering, 1994. 208 p.
Амазонский дельфин / Под ред. Соколова В.Е. М.: Наука, 1996. 296 с.
Norris K.S. Some problems of echolocations in Cetacea // Oxford: Marine bioacoustics, 1964. P. 317–336.
Рябов В.А. Механизмы проведения эхолокационного слуха дельфина / Карадагский природный заповедник. Национальная академия наук Украины, Феодосия, Украина. 2009.
Попов В.В. Особенности слуха китообразных: электрофизиологический подход. Автореферат дисс. на соискание ученой степени доктора биол. наук. М., 2005. 48 с.
Сысуева Е.В. Звукопроведение в слуховой системе зубатых китообразных. Автореферат дисс. на соискание ученой степени канд. биол. наук. М., 2012. 26 с.
McCormick J.G., Wever E.G., Palin J., Ridgway S.H. Sound conduction in the dolphin ear // J. Acoust. Soc. Am. 1970. V. 48. № 6 (2). P. 1418–1428.
Diercks K.J., Trochta R.T., Greenlow C.F., Evans W.E. Recording and analisis of dolphin echolocation signals // J. Acoust. Soc. Am. 1971. V. 49. № 6 (1). P. 1729.
Cranford T.W., Elsberry W.R., Van Borin W.G., Jeffress J.A., Chaplin M.S., Blackwood D.J., Carder D.A., Kamolnick T., Todd M.A., Ridjway S.H. Observation and analysis of sonar signal generation in the bottlenose dolphin (Tursiops truncatus): Evidence for two sonar sources // J. Experimental Marine Biology and Ecology. 2007. V. 407. P. 81–96.
Романенко Е.В., Чикалкин В.А. Магнитофон для подводной записи // Морфология, физиология и акустика морских млекопитающих. М.: Наука, 1974. С. 165–173.
Романенко Е.В. Некоторые результаты исследования акустики черноморских афалин / В кн.: Черноморская афалина. Изд. Наука, 1997. С. 609–620.
Romanenko E.V. Purposeful changes in the Structure of ecolocation pulses in Tursiops truncates / In: Sensory Abilites of Cetaceans. Laboratory and Field Evidence. Eds. Thomas J.A. and Kastelein R.A. NATO ASi Series, Series A: Life Science. Plenum Press. New York. 1990. V. 196. P. 317–319.
Заславский Г.Л. Исследование локационных сигналов дельфинов при помощи двухканальной системы регистрации // Биофизика. 1972. Т. 17. № 4. С. 717–720.
Айрапетьянц Е.Ш., Константинов А.И. Эхолокация в природе. Л.: Наука, 1974. 512 с.
Ridgway S.H., Carder D.A. Nasal pressure and sound production in an echolocation white whale Delphinopterus leucas / In.: Animal Sonar. Processes and Performance. Eds. Nachtigall P.E., Moore P.W.B. NATO ASi Series. Series A: Life Science. 1988. V. 156. P. 53–66.
Даргольц В.Г., Романенко Е.В., Юматов Е.А., Янов В.Г. Методика телеметрической регистрации динамики давления крови и дыхания у свободно плавающих дельфинов // Физиол. журн. 1981. Т. 67. № 11. С. 1744–1748.
Протасов В.Р., Романенко Е.В. О характере звуков, издаваемых некоторыми рыбами Черного моря // Докл. АН СССР. 1961. Т. 139. № 3. С. 725–728.
Протасов В.Р., Романенко Е.В. Звуки, издаваемые некоторыми рыбами, и их сигнальное значение // Зоологический журн. 1962. Т. 41. № 10. С. 1516–1527.
Протасов В.Р., Романенко Е.В., Подлипалин Ю.Д. О биологическом значении звуков некоторых рыб // Вопросы ихтиологии. 1965. Т. 5. № 3/36. С. 532–539.
Никольский И.Д., Протасов В.Р., Романенко Е.В., Шишкова Е.В. Звуки рыб. Атлас. М.: Наука, 1968. 93 с.
Протасов В.Р. Биоакустика рыб. М.: Наука, 1965. 206 с.
Протасов В.Р. Биоэлектрические поля в жизни рыб. М.: ЦНИИТЭИРХ, 1971. 228 с.
Дополнительные материалы отсутствуют.
Инструменты
Акустический журнал