

Биология моря, 2019, T. 45, № 1, стр. 3-7
Динамика численности сообщества настоящих тюленей семейства Phocidae в заливе Пильтун (остров Сахалин) в неледовый период 1999 и 2014–2017 годов
А. М. Трухин 1, *, П. А. Пермяков 1
1 Тихоокеанский океанологический институт им. В.И. Ильичева ДВО РАН
690041 Владивосток, Россия
* E-mail: marian1312@mail.ru
Поступила в редакцию 12.03.2018
После доработки 06.06.2018
Принята к публикации 18.10.2018
Аннотация
Исследовано береговое лежбище настоящих тюленей, расположенное в устье зал. Пильтун (северо-восток о-ва Сахалин), и определена численность тюленей на нем в течение неледового периода. На сегодняшний день это крупнейшее поливидовое лежбище настоящих тюленей на северо-восточном побережье о-ва Сахалин. В летне–осенний период здесь в сопредельной акватории одновременно нагуливаются три вида пагофильных тюленей: морской заяц (лахтак), кольчатая нерпа (акиба) и пятнистая нерпа (ларга). В 1999 г. и с 2014 по 2017 гг. общее время наблюдений за лежбищем составило 630 сут, выполнен 3361 учет численности тюленей. Показано, что сезонная динамика обилия животных на лежбище характеризуется тремя ежегодными последовательными периодами увеличения их численности (июнь–июль, август–сентябрь, октябрь), каждый из которых совпадает с миграциями через устье залива проходных видов рыб. В 1999 г. динамика численности тюленей статистически значимо отличалась от таковой в 2014–2017 гг., а в 2014 г. – от таковой в 2016 г. В 1999 г. медиана общей численности была значительно ниже, чем в любой из сезонов 2014–2017 гг.
На протяжении последних 20 лет происходит активное промышленное освоение северо-востока Сахалинского шельфа. Введение в середине 1990-х годов в эксплуатацию Пильтун-Астохского нефтегазового месторождения вызвало закономерные опасения научных и общественных организаций, связанные с возможным негативным воздействием нефтегазоразработок на биоту зал. Пильтун. Ситуация осложнялась недостаточной изученностью местной фауны. Хорошим индикатором состояния любой экосистемы служат виды, занимающие наиболее высокие трофические уровни в пищевых цепях (Одум, 1975; Коли, 1979). На акватории северо-восточного шельфа о-ва Сахалин роль консумента высшего порядка играют морские млекопитающие. По этой причине в качестве объекта исследования были выбраны настоящие тюлени (Phocidae), обитающие в устье зал. Пильтун. В неледовый период здесь встречаются три вида пагофильных безухих тюленей: морской заяц (лахтак) Erignathus barbatus (Erxleben, 1777), кольчатая нерпа (акиба) Pusa hispida Schreber, 1775 и пятнистая нерпа (ларга) Phoca largha Pallas, 1811, которые используют лежбище, являющееся в настоящее время крупнейшим на о-ве Сахалин (Трухин, Блохин, 2003). Ядро берегового сообщества составляют тюлени-ихтиофаги ларга и акиба, с явным преобладанием первого вида. Численность бентосоядного лахтака исчисляется несколькими десятками.
Об использовании зал. Пильтун ластоногими в летне–осеннее время упоминалось и ранее (Амброз, 1931; Косыгин и др., 1986; Лагерев, 1988), однако подробно пильтунское лежбище было описано только в 1999 г. (Трухин, 2000). Было показано наличие сложной хорологической структуры лежбища, описано пространственно-временное распределение тюленей по видам и возрастным категориям на береговых участках и в заливе (Трухин, Блохин, 2003; Соболевский, 2004), а также получены первые нередко противоречивые сведения о суточной и сезонной динамике численности тюленей (Соболевский, 2000; Трухин, Блохин, 2003; Bradford, Weller, 2005). Максимальная численность пильтунского сообщества тюленей была оценена в 1–2 тыс. особей (Трухин, Блохин, 2003; Соболевский, 2004). Данный этап исследований ластоногих был завершен к началу 2000-х годов, а в 2013 г. нефтедобывающими компаниями было принято решение о строительстве берегового объекта нефтедобычи “Одопту” недалеко от лежбища тюленей, что вызвало обеспокоенность его будущим. В связи с этим в 2014 г. исследования тюленей были возобновлены в виде мониторинговых работ по нескольким направлениям.
Цель данного исследования – изучение характера изменения численности поливидового скопления тюленей на лежбище в устье зал. Пильтун в 1999 г., 2014–2017 годах. Для достижения цели были определены основные задачи: выявить особенности сезонной динамики численности сообщества тюленей на лежбище в зал. Пильтун и определить многолетнюю тенденцию динамики численности сообщества с 1999 по 2017 г.
МАТЕРИАЛ И МЕТОДИКА
Исследование было выполнено в неледовые периоды 2014–2017 гг. в устье зал. Пильтун. Ежегодно работы начинали после исчезновения в прибрежной акватории ледового покрова, а их окончание было связано с появлением в прибрежье ледовых заберегов, препятствующих выходу животных на сушу. В общий массив данных включены материалы, собранные здесь же в летне-осенний период 1999 г. (Трухин, Блохин, 2003). Во все сезоны наблюдения проводили со смотровой площадки, расположенной в башне Пильтунского маяка в 1 км от лежбища (напротив устья залива). Основным методическим приемом был прямой визуальный учет численности тюленей. Дифференциацию учетных данных по видам тюленей не проводили, считая их единым поливидовым скоплением (сообществом). Для контроля численности ластоногих использовали оптические приборы фирмы “Nikon” (бинокли 8 × 40 и 20–60-кратную подзорную трубу). В течение 630 сут (от 114 до 136 сут за сезон) был выполнен 3361 учет численности (от 487 до 801 учета за сезон) (табл. 1).
Таблица 1.
Результаты наблюдений за лежбищем настоящих тюленей в устье зал. Пильтун
| Показатель | 1999 г. | 2014 г. | 2015 г. | 2016 г. | 2017 г. |
|---|---|---|---|---|---|
| Начало наблюдений | 30.05 | 3.07 | 18.06 | 7.06 | 6.06 |
| Окончание наблюдений | 12.11 | 30.10 | 26.10 | 25.10 | 26.10 |
| Продолжительность наблюдений, сут | 136 | 114 | 124 | 128 | 126 |
| Количество учетов численности | 619 | 487 | 728 | 726 | 801 |
| Медиана общей численности, особи | 172 | 502 | 611 | 650 | 556 |
| Максимум сезонной численности, особи | 1715 | 2620 | 2473 | 2042 | 2596 |
| Дата максимума сезонной численности | 6.10 | 17.09 | 26.09 | 25.09 | 24.08 |
Сравнивали массивы данных, в основу которых были положены максимальные значения численности за каждые сутки. Массивы данных не проходили тестов на нормальность (Шапиро–Вилк, p < 0.05), поэтому в качестве центральной тенденции использовали медиану численности. Доверительные интервалы (95%) для оценки медианы рассчитывали методом перцентильного бутстрепа (BCI; R = 1000).
Групповое сравнение сезонов выполнено по критерию Крускала–Уоллиса; попарное сравнение сезонов – по критерию Манна–Уитни (Гланц, 1999; Zar, 2010). При сравнении выборок с сильно различающимися распределениями (2016 г.) попарные сравнения дополняли двухвыборочными перестановочными тестами (Efron, Tibshirani, 1993). В качестве тестовой статистики (Θ) перестановочного теста приняли разницу между медианами в субсетах данных (R = 1000). Накопленный уровень значимости (ASL) определили как долю случаев, в которых разница между медианами в субсетах псевдовыборок была по модулю больше, чем разница между медианами сравниваемых выборок. Уровни значимости тестовых статистик (p, ASL) для всех методов приняли равными 0.05.
Статистическая обработка данных выполнена с помощью программы MSExcel и статистического пакета GraphPad Prism.
РЕЗУЛЬТАТЫ
Сезонная динамика
Сезонная динамика численности тюленей на лежбище в зал. Пильтун ежегодно характеризовалась тремя хорошо различимыми периодами увеличения численности, когда скользящее 5-суточное среднее оказывалось выше 300–370 особей, и ее снижения (рис. 1). Впервые тюлени появлялись на лежбище, как правило, после исчезновения из прибрежий льда, обычно в начале июня. Численность тюленей в течение этого месяца стабильно увеличивалась и достигала максимальных для этого периода значений к концу июня – в первой декаде июля, а к концу июля снижалась (рис. 1а). Медиана численности Me с начала сезона до 25 июля составляла 387 особей (IQR = 314–553 особи). Характерно возникновение непродолжительной паузы между первым и вторым периодами увеличения и спада численности тюленей (третья декада июля). В течение этого короткого периода медиана численности составляла 324 особи (IQR = 314–331 особь). Второй период был более продолжительным, он начинался в августе и длился до конца сентября (рис. 1б). В это время Me = 744 особи (IQR = 577–832 особи). Для данного периода можно выделить несколько коротких и нерегулярных всплесков численности, следовавших один за другим. Ежегодный максимум сезонной численности приходился на вторую половину сентября – начало октября (табл. 1), и лишь в 2017 г. он был зарегистрирован в третьей декаде августа. Третье повышение численности происходило в октябре (рис. 1в). В это время медиана численности составляла 399 особей (IQR = 229–535 особей).
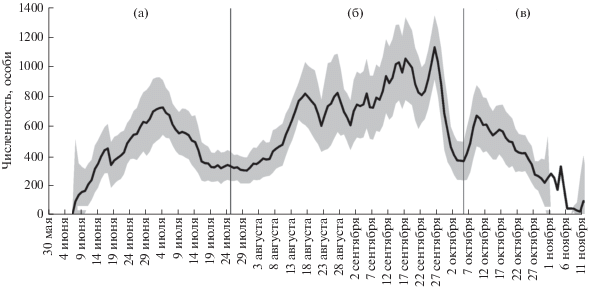
Межгодовая динамика
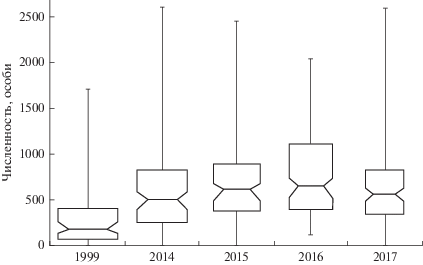
При групповом сравнении сезоны разных лет статистически значимо различались (Крускал–Уоллис, p < 0.01). Попарные сравнения показывают различия между 1999 г. и сезонами 2014–2017 гг. (Манн–Уитни, p < 0.05 во всех случаях). Отмечено также существование значимых различий между сезонами 2014 и 2016 гг. (Манн–Уитни, p < 0.01; перестановочный тест ASL = 0.04). Сезонная динамика численности для всего неледового периода 2014, 2015 и 2017 гг. статистически не различалась (Крускал–Уоллис, p = 0.25). Наблюдаемые различия определяются общим уровнем численности, характерным для группировки в разные годы. В 1999 г. медиана сезонной численности была чрезвычайно низкой (172 особи; BCI = 126–256 особей) по сравнению с таковой в 2014 г. и позже (рис. 2). К 2014 г. произошел рост медианы численности (до 502 особей; BCI = 384–824 особи). Наибольшего значения медиана численности достигла в 2016 г. (650 особей; BCI = 523–738 особей). Наблюдаемое снижение медианы численности к 2017 г. не было статистически достоверным, так как границы доверительных бутстреп-интервалов пересекаются (рис. 2).
ОБСУЖДЕНИЕ
На расположенном в зал. Пильтун лежбище тюлени не размножаются, поэтому жесткая сезонность в его использовании тюленями представляется, на первый взгляд, маловероятной. Тем не менее нельзя утверждать, что сезонная динамика численности пильтунского берегового сообщества тюленей носит случайный характер. Из года в год сезонная численность животных изменяется по сходному сценарию, воспроизводя одну и ту же картину с тремя пиками, конфигурация которых впервые была определена еще в конце прошлого века (Трухин, Блохин, 2003). Ранее предполагалось, что сезонные всплески численности тюленей связаны с периодическим изменением состава корма (Трухин, Блохин, 2003). По представлениям Соболевского (2004), характер сезонных изменений численности тюленей на пильтунском лежбище обусловлен изменением локального обилия проходных лососевых. Веские основания для подтверждения того, что все три периода увеличения и снижения численности тюленей совпадают с периодами рунного хода анадромных лососей, отсутствуют. Однако предположение об обусловленности сезонной динамики численности именно трофическими факторами среды представляется нам наиболее реалистичным.
Стабильный рост численности тюленей в июне (рис. 1а) по срокам совпадает с нерестовыми миграциями в районе лежбища сельди Clupea pallasii Valenciennes, 1847; малоротой корюшки Hypomesus olidus (Pallas, 1814); крупночешуйной красноперки Tribolodon hakonensis (Günther, 1877); амурского сига Coregonus ussuriensis Berg, 1906; мойвы Mallotus villosus (Müller, 1776); трехиглой колюшки Gasterosteus aculeatus Linnaeus, 1758; кунжди Salvelinus leucomaenis (Pallas, 1814) и горбуши Oncorhynchus gorbuscha (Walbaum, 1792) (см.: Гриценко, 2002; Земнухов, 2008; наши данные).
С июля по первую декаду августа численность тюленей на лежбище имеет сравнительно низкие значения. Эта пауза может быть связана с летним прогреванием акватории зал. Пильтун, из-за которого в лимане присутствует преимущественно фоновая ихтиофауна, образованная эвритермными видами. Второе повышение численности тюленей (рис. 1б), по-видимому, связано с нагульно-нерестовыми миграциями лососевых: кеты Oncorhynchus keta (Walbaum, 1792), кижуча Oncorhynchus kisutch (Walbaum, 1792) и кунджи, а также с миграциями красноперки и малоротой корюшки. Третье повышение численности тюленей на лежбище (рис. 1в) соотносится с нагульными миграциями сельди, а также с нерестовыми миграциями кижуча и зимовальными миграциями красноперки и азиатской корюшки Osmerus mordax (Mitchill, 1814). Каждое повышение численности тюленей на лежбище происходило в периоды увеличения объемов уловов местной рыбодобывающей артели, ведущей промысел в лагуне и на сопредельной акватории.
Численность бентосоядного лахтака на лежбище на протяжении всего сезона не превышала нескольких десятков особей. Этот вид не оказывает заметного влияния на общую численность сообщества тюленей в зал. Пильтун.
В разное время в заливе обитает большая группа видов рыб небольшого размера. Эти некрупные рыбы (среди которых молодь кунджи, наваги и других видов), во-первых, доступны всем тюленям-ихтиофагам (независимо от размера или возраста последних), во-вторых, образуют обильную и стабильную кормовую базу. Следовательно, флуктуация численности тюленей в течение их нагульного сезона связана как с нерестовым ходом собственно лососевых, так и с миграцией через устье зал. Пильтун других видов проходных рыб, особенно в периоды прекращения рунного хода того или иного вида лососей.
Сравнительно высокая численность тюленей на данном лежбище в течение неледовых периодов – следствие наличия здесь подходящих участков суши, обеспечивающих высокую степень защищенности, и богатой кормовыми ресурсами акватории. Общий характер динамики численности всего лежбищного скопления тюленей в значительной мере определяется сезонными и межгодовыми изменениями численности доминирующей в составе сообщества ларги. С 1999 г. численность тюленей в местном сообществе выросла, достигнув максимального значения к 2015–2016 гг. Снижение обилия тюленей в 2017 г. не было статистически значимым, что не дает оснований говорить об уменьшении их численности на лежбище.
Таким образом, ежегодная динамика численности тюленей на лежбище в устье зал. Пильтун во время неледового периода воспроизводит одну и ту же картину с тремя хорошо выраженными эпизодами увеличения и снижения численности и определяется локальным обилием ихтиофауны, которое, в свою очередь, связано с нагульными и нерестовыми миграциями через устье залива проходных видов рыб. В 2014–2017 гг. каких-либо негативных изменений численности сообщества ластоногих в устье зал. Пильтун, связанных с деятельностью в этом районе нефтедобывающих компаний, не отмечено.
Авторы признательны сотрудникам “Эксон Нефтегаз Лимитед” М. Свиндоллу (M. Swindoll), Э.Н. Калинину, В.В. Ефремову, С.П. Стародымову, Е.В. Вяткиной, работа которых в значительной степени помогла успешному выполнению данного исследования. Г.В. Думенко способствовала решению задач, связанных с логистикой. Отдельная благодарность командованию войсковой части № 13178, предоставившему возможность проживания научной группы на Пильтунском маяке, и начальнику маяка Д.А. Рожно за помощь в организации быта. Местные рыболовецкие бригады сообщали о рыбопромысловой обстановке в заливе и на сопредельной морской акватории. Дополнительную полезную информацию предоставляли нам в разное время А.В. Бобков, В.В. Вертянкин, В.В. Черницын, С.В. Фомин, А.А. Шестак, П. ван дер Волф (P. van der Wolf), а также А.Е. Волков, О.А. Сыченко и участники руководимой ими группы IFAW. В разные годы в состав полевых групп помимо авторов данной статьи входили В.Г. Кавозг, П.Г. Маметьев, Н.В. Сутягин и А.И. Чесноков. В обсуждении работы принимали участие П.М. Жадан и С.Д. Рязанов. Авторы признательны рецензентам за ценные замечания.
Исследование выполнено на средства и при организационной поддержке компании “Эксон Нефтегаз Лимитед”.
Список литературы
Амброз А.И. Некоторые наблюдения над морскими млекопитающими восточного Сахалина // Рыб. хоз-во Дальнего Востока. 1931. Т. 3–4. С. 46–48.
Гланц С. Медико-биологическая статистика. 4. М.: Практика. 1999. 459 с.
Гриценко О.Ф. Проходные рыбы острова Сахалин (систематика, экология, промысел). М.: Изд-во ВНИРО. 2002. 248 с.
Земнухов В.В. Ихтиофауна залива Пильтун (северо-восточный Сахалин): состав, экология, происхождение: Автореф. дис. … канд. биол. наук. 03.00.10. Владивосток: ИБМ ДВО РАН. 2008. 23 с.
Коли Г. Анализ популяций позвоночных. М.: Мир. 1979. 365 с.
Косыгин Г.М., Трухин А.М., Бурканов В.Н., Махнырь А.И. Лежбища ларги на берегах Охотского моря // Научно-исследовательские работы по морским млекопитающим северной части Тихого океана в 1984–1985 гг. М.: ВНИРО. 1986. С. 60–70.
Лагерев С.А. Результаты авиационного обследования береговых лежбищ тюленей Охотского моря в 1986 г. // Научно-исследовательские работы по морским млекопитающим северной части Тихого океана в 1986–1987 гг. М.: ВНИРО. 1988. С. 80–89.
Одум Ю. Основы экологии. М.: Мир. 1975. 741 с.
Соболевский Е.И. Распределение и численность тюленей у северо-восточного Сахалина // Морские млекопитающие Голарктики: Материалы конф. Архангельск, 21–23 сентября 2000 г. М.: КМК. 2000. С. 347–349.
Соболевский Е.И. Распределение и численность тюленей в заливе Пильтун (северо-восточный Сахалин) в летне-осенний период // Биол. моря. 2004. Т. 30. № 4. С. 312–315.
Трухин А.М. Кольчатая нерпа на восточном побережье острова Сахалин // Морские млекопитающие Голарктики: Тез. докл. Архангельск, 21–23 сентября. Архангельск. 2000. С. 394–396.
Трухин A.M., Блохин С.А. Особенности функционирования поливидового лежбища настоящих тюленей (Phocidae) в районе добычи углеводородного сырья на шельфе острова Сахалин // Экология. 2003. № 4. С. 316–322.
Bradford A.L., Weller D.W. Spotted seal haul-out patterns in a costal lagoon on Sakhalin Island, Russia // Mammal Study. 2005. V. 30. P. 145–149. http://dx.doi.org/. doi 10.3106/1348-6160(2005)30[145:SSHPIA]2.0.CO;2
Efron B., Tibshirani R.J. An Introduction to the Bootstrap. London: Chapman & Hall. 1993. 456 p.
Zar J.H. Biostatistical analysis, 5th ed. N.J.: Prentice Hall. 2010. 944 p.
Дополнительные материалы отсутствуют.
Инструменты
Биология моря