

Биология моря, 2020, T. 46, № 6, стр. 420-432
Инвентаризация бентосной флоры литорали и верхней сублиторали бухты Троицы (залив Петра Великого, Японское море)
О. С. Белоус 1, 2, *, Т. В. Титлянова 1, Э. А. Титлянов 1
1 Национальный научный центр морской биологии им. А.В. Жирмунского ДВО РАН
690041 Владивосток, Россия
2 Тихоокеанский институт биоорганической химии им. Г.Б. Елякова ДВО РАН
690022 Владивосток, Россия
* E-mail: ksu_bio@mail.ru
Поступила в редакцию 09.07.2019
После доработки 19.12.2019
Принята к публикации 19.03.2020
Аннотация
В 2004–2018 гг. с июня по сентябрь исследовали видовой состав макрофитов в б. Троицы (зал. Петра Великого, Японское море). Найдено 111 видов макрофитов, из них 28 видов – эпифиты. Наиболее широко был представлен отдел Rhodophyta – 58 видов (54% от общего числа обнаруженных видов), найдено 35 видов (32%) из отдела Ochrophyta (класс Phaeophyceae), а также 15 видов (14%) из отдела Chlorophyta; отмечено 3 вида (3%) морских трав. Видовое разнообразие водорослей увеличивалось от литорали к сублиторали. На литорали и в верхней сублиторали б. Троицы выделено 23 растительных сообщества.
Опубликовано большое количество работ, посвященных изучению макробентоса юго-западной части зал. Петра Великого (Перестенко, 1968, 1971а, 1971б, 1980, 1994; Паймеева, 1972; Вышкварцев, Пешеходько, 1982; Кафанов, Жуков, 1986, 1993; Некрасов и др., 2000), однако данные о флоре б. Троицы немногочисленны (Галышева, Христофорова, 2008; Белоус и др., 2013; Белоус, Дроздов, 2015) и ограничены сведениями только о видовом составе макрофитобентоса. Так, в сублиторали б. Троицы найдено 38 видов макрофитов (Галышева, Христофорова, 2008). Показано, что на открытых участках побережья в основном встречались бурые и красные водоросли, а на защищённых от волн участках – зеленые водоросли и морские травы. Ранее на литорали и в верхней сублиторали б. Троицы нами найдено 102 вида макрофитов, из них 48% – красные водоросли, 34% – бурые, 15% – зеленые и 3% – морские травы (Белоус и др., 2013). В результате изучения растительных сообществ в зал. Посьета (Перестенко, 1980; Кафанов, Жуков, 1993; Лебедев и др., 2004), в который входит б. Троицы, обнаружена 21 литоральная ассоциация водорослей (Перестенко, 1980), а в соседней б. Витязь выделено 5 фитоценозов (Кафанов, Жуков, 1993). В б. Троицы сообщества макрофитобентоса ранее не были исследованы.
Цели настоящей работы – уточнить таксономический состав макрофитов и выделить фитоценозы в пределах литоральной и верхней сублиторальной зон б. Троицы.
МАТЕРИАЛ И МЕТОДИКА
Место и время исследования
Б. Троицы (42°38′55″ с.ш., 131°06′17″ в.д.) (рис. 1) – акватория в северо-восточной части зал. Посьета (зал. Петра Великого, Японское море). Общая площадь бухты составляет 16.8 км2, максимальная глубина – 30 м (Лоция Японского моря, 1972).

Район расположен в умеренном климатическом поясе муссонного типа. Самые теплые месяцы – июль и август; температура воздуха в это время колеблется от 20 до 25°С. Среднее значение температуры в поверхностном слое воды в августе составляет 23°С, а в полузакрытых бухточках достигает 27°С. Движение воды в б. Троицы осуществляется под влиянием как постоянных (например, антициклоническая циркуляция воды в центральной части бухты), так и временных (нагонно-сгонных) течений. От Корейского полуострова до зал. Посьета и б. Троицы доходит ветвь теплого Цусимского течения. С севера от Татарского пролива вдоль берегов Приморья проходит холодное Приморское течение. Соленость поверхностного слоя воды в бухте в сухой период года колеблется от 30 до 32‰, в дождливый период вода может сильно опресняться (Атлас океанов, 1974). Средняя величина прилива составляет 0.2 м с максимальной амплитудой до 0.5 м (Григорьева и др., 2001).
Водоросли собирали на восьми участках (точки 1–8) побережья (рис. 2) с июня по сентябрь в 2004–2018 гг. в литоральной и верхней сублиторальной зонах на глубине 0–2.5 м. Степень прибойности определяли по: Гурьянова и др., 1930.
Рис. 2.
Места сбора водорослей. а – мыс Андреева (точка 1), б – водолазная станция (точки 2 и 3), в – мыс Слычкова (точки 4 и 5), г – мыс Стенина (точка 6), д – б. Идол (точка 7), е – б. Рисовая (точка 8).

Точка 1 – восточное побережье бухты у мыса Андреева, от верхней литорали до верхней сублиторали; характеризуется скалисто-каменистым дном и прибойностью II степени (рис. 2а).
Точка 2 – восточное побережье бухты вблизи водолазной станции; характеризуется песчаным грунтом в верхней литорали и песчано-илистым с камнями грунтом в верхней сублиторали. Прибойность IV степени (рис. 2б).
Точка 3 – восточное побережье бухты (водолазная станция); верхняя литораль и верхняя сублитораль представлены скалисто-каменистым грунтом. Прибойность IV степени.
Точки 4 и 5 – район у мыса Слычкова (юго-западный входной мыс бухты). Берега мыса возвышенные и обрывистые; дно скалистое в верхней литорали, скалисто-каменистое с галькой и валунами в сублиторали. Дно открытой части побережья покрыто крупными валунами. Прибойность II степени (рис. 2в).
Точка 6 – акватория у мыса Стенина (восточный входной мыс бухты) со скалистым обрывистым берегом; грунт в верхней литорали скалисто-каменистый, в сублиторали песчано-каменистый. Прибойность II степени (рис. 2г).
Точка 7 – восточное побережье (б. Идол); характеризуется песчано-галечно-валунным грунтом с выходами коренных пород. Прибойность IV степени (рис. 2д).
Точка 8 – восточное побережье (б. Рисовая); дно песчаное с отдельно встречающимися камнями и выходами коренных пород. Прибойность III степени (рис. 2е).
Методика сбора образцов; оценка встречаемости водорослей, проективного покрытия и доминирования отдельных видов
Водоросли собирали при отливе на литорали и в верхней сублиторали. В каждой зоне водорослевые сообщества выделяли визуально и фотографировали. Проективное покрытие дна водорослями определяли в процентном соотношении визуально и по фотографиям. Виды с проективным покрытием дна менее 10% считали редкими (+), с проективным покрытием 10–50% – обычными (++) и более 50% – массовыми (+++).
Тип сообщества определяли по количеству доминирующих видов (Титлянов и др., 2018). К монодоминатным относили сообщества, в которых доминирующий вид покрывал более 50% поверхности дна, к бидоминантным – сообщества, в которых два вида водорослей покрывали более чем 50% поверхности дна. В полидоминантных сообществах более 50% дна покрывали несколько доминантных видов. Названия фитоценозам давали без учета ярусности. Термины “фитоценоз” и “сообщество” употребляли как синонимы.
Водоросли, собранные из разных сообществ, помещали в отдельные пластиковые пакеты, которые хранили в холодильнике. При идентификации материала использовали определители (Виноградова, 1979; Перестенко, 1980, 1994; Титлянов и др., 2016). Названия таксонов приведены согласно www.algaebase.org (Guiry, Guiry, 2019). Иерархическая классификация отдела Rhodophyta соответствует предложенной ранее (см.: Saunders, Hommersand, 2004). Система классификации отделов Chlorophyta и Ochrophyta дана по: Tsuda, 2003.
Степень видового сходства в разных точках устанавливали по коэффициенту Серенсена–Чекановского (Песенко, 1982). Дендрограммы строили по методу группового среднего с использованием коэффициента сходства Серенсена–Чекановского, рассчитанного на основе данных по встречаемости видов. Для анализа сходства точек по видовому составу использовали пакет статистических программ PRIMER v.6 (Clarke, Gorley, 2006).
Гербарий водорослей хранится в Лаборатории автотрофных организмов Национального научного центра морской биологии им. А.В. Жирмунского ДВО РАН.
РЕЗУЛЬТАТЫ
Видовое разнообразие макрофитов б. Троицы
В б. Троицы обнаружено 108 видов макроводорослей и 3 вида морских трав (табл. 1, 2). В собранной коллекции отдел Rhodophyta содержит 58 видов (54% от всей коллекции), принадлежащих к трем классам (Bangiophyceae, Compsopogonophyceae, Florideophyceae), 12 порядкам, 27 семействам и 45 родам; отдел Ochrophyta представлен 35 видами (32% коллекции) из класса Phaeophyceae, принадлежащими к семи порядкам, 14 семействам и 30 родам; отдел Chlorophyta содержит 15 видов (14%) из класса Ulvophyceae, принадлежащих к четырем порядкам, шести семействам и восьми родам. Среди красных водорослей наибольшее число видов принадлежит к семействам Rhodomelaceae (12 видов), Ceramiaceae (5 видов) и Gigartinaceae (4 вида); среди бурых водорослей – к семействам Chordariaceae (9 видов) и Sargassaceae (4 вида); среди зеленых – к семействам Ulvaceae (6 видов) и Cladophoraceae (4 вида). Отдел морские травы (Tracheophyta) представлен тремя видами, принадлежащими к классу Monocots, порядку Alismatales, семейству Zosteraceae. Это виды родов Phyllospadix (1 вид) и Zostera (2 вида). Места сбора по мере снижения их видового разнообразия (убывания числа видов) располагаются следующим образом: б. Рисовая (67 видов) > б. Идол (61 вид) > мыс Андреева (49 видов) > водолазная станция (43 вида) > мыс Слычкова (25 видов) > > мыс Стенина (24 вида) (табл. 1, 2).
Таблица 1.
Растительные сообщества, обнаруженные в б. Троицы, и количество представленных в них видов
| Точки сбора | Обнаруженные сообщества | Количество видов в сообществе |
|---|---|---|
| 1 | МС – Nemalion vermiculare | 1 |
| МС – Corallina pilulifera | 1 | |
| БС – Corallina pilulifera + Neorhodomela aculeata | 5 | |
| МС – Mastocarpus pacificus | 6 | |
| ПС – доминанты:Dictyota dichotoma, Chondrus armatus, Chordaria flagelliformis, Neorhodomella aculeata | 49 | |
| 2 | МС – Gloiopeltis furcata | 1 |
| БС – Polysiphonia morrowii + Desmarestia viridis | 6 | |
| 3 | ПС – доминанты:Ulva spp., Leathesia marina, Punctaria plantaginea | 43 |
| БС – Ulva linza + Cladophora stimpsonii | 2 | |
| БС – Punctaria plantaginea + Ulva lactuca | 8 | |
| МС – Zostera marina | 5 | |
| 4, 5, 6 | МС – Ralfsia verrucosa | 3 |
| БС – Corallina pillulifera + Chordaria flagelliformis | 4 | |
| ПС – Costaria costata, Undaria pinnatifida, Scytosiphon lomentaria, Melanothamnus japonicus, Chordaria flagelliformis и др. | 25 (точки 4, 5) 24 (точка 6) |
|
| МС – Laurencia nipponica | 2 | |
| МС – Saccharina angustata | 5 (точка 6) | |
| 7 | МС – Gloiopeltis furcata | 1 |
| МС – Ralfsia verrucosa | 1 | |
| БС – Leathesia marina + Nemalion vermiculare | 2 | |
| ПС – доминанты:Sargassum miyabei, Dictyota dichotoma, Dictyopteris divaricata | 61 | |
| БС – Sargassum miyabei + Chorda filum | 15 | |
| 8 | МС – Gloiopeltis furcata | 1 |
| МС – Analipus japonicus | 14 | |
| МС – Chaetomorpha moniligera | 4 | |
| ПС – доминанты:Melanothamnus japonicus, Analipus japonicus, Chordaria flagelliformis | 67 | |
| МС – Sargassum miyabei | 13 |
Таблица 2.
Инвентаризационный список водорослей, найденных в б. Троицы в 2004–2018 гг.
| Видовой состав | Зона произрастания | Обилие | Экологическая группировка | Место произрастания |
|---|---|---|---|---|
| Отдел Rhodophyta | ||||
| Класс Bangiophyceae Порядок Bangiales Семейство Bangiaceae |
||||
| Porphyra inaequicrassa L.P. Perestenko | вл | ед | эпп, эф (Chorda fillum) | Ид |
| Pyropia yezoensis (Ueda) M.S. Hwang & H.G. Choi | вл | + | эл | Ид |
| Класс Compsopogonophyceae Порядок Erythropeltales Семейство Erythrotrichiaceae |
||||
| Erythrotrichia carnea (Dillwyn) J. Agardh | нл, вс | + | эф | П |
| Класс Florideophyceae Порядок Nemaliales Семейство Nemaliaceae |
||||
| Nemalionvermiculare Suringar | вл | ++ | эл | П |
| Порядок Gelidiales Семейство Gelidiaceae |
||||
| Gelidium amansii (J.V. Lamouroux) J.V. Lamouroux | вс | + | эл | Ид |
| Gelidium vagum Okamura | вс | ед | эл | Ид |
| Порядок Corallinales Семейство Corallinaceae |
||||
| Pachyarthroncretaceum (Postels et Ruprecht) Manza | вл, лл, сл, нл, вс | ++ | эл | П |
| Corallina pilulifera Postels et Ruprecht | вл, лл, сл, нл, вс | ++ | эл | П |
| Pneophyllum zostericola (Foslie) D. Fujita | нл, вс | + | эф | Вод, Рис |
| Семейство Hydrolithaceae | ||||
| Hydrolithon farinosum (J.V. Lamouroux) Penrose & Y.M. Chamberlain | c | + | эф | Ан |
| Семейство Lithothamniaceae | ||||
| Lithothamnion phymatodeum Foslie | вл, нл, вс, сл | + | эл | П |
| Порядок Gigartinales Семейство Dumontiaceae |
||||
| Masudaphycusirregularis (Yamada) Lindstrom | вс | ед | эл | Рис |
| Семейство Phyllophoraceae | ||||
| Mastocarpuspacificus (Kjellman) L.P. Perestenko | вл, лл, нл | + | эл | Ан, Вод, Сл, Ст |
| Ahnfeltiopsisflabelliformis (Harvey) Masuda | вл, лл, нл, сл | + | эл | Ан |
| Coccotylus orientalis (A.D. Zinova & Makienko) Perestenko | c | + | эл | Вод |
| Семейство Gigartinaceae | ||||
| Chondrus armatus (Harvey) Okamura | нл, вс | + | эл | Ан, Вод |
| Chondruspinnulatus (Harvey) Okamura | вл, лл, сл, нл, вс | + | эл | Ан |
| Chondrusyendoi Yamada et Mikami | вл, нл, вс, сл | + | эл | Рис |
| Mazzaellajaponica (Mikami) Hommersand | вл, лл, сл, нл, вс | + | эл | Рис |
| Семейство Gloiosiphoniaceae | ||||
| Gloiosiphonia capillaris (Hudson) Carmichael | нл, вс | + | эл | Ид |
| Семейство Tichocarpaceae | ||||
| Tichocarpuscrinitus (S.G. Gmelin) Ruprecht | нл, вс | + | эл | Ан, Рис |
| Семейство Endocladiaceae | ||||
| Gloiopeltisfurcata (Postels et Ruprecht) J. Agardh | вл | ++ | эл | П |
| Семейство Furcellariaceae | ||||
| Opuntiella parva Perestenko | c | + | эпп,эл | Вод |
| Семейство Kallymeniaceae | ||||
| Callophyllisrhynchocarpa Ruprecht | вс | + | эл, эпп | Вод |
| Порядок Halymeniales Семейство Halymeniacea |
||||
| Grateloupia acuminata Holmes | нл, вс | + | эл | Сл, Ан |
| Grateloupiadivaricata Okamura | вл, нл, сл, лл, вс | + | эл, эпп | Ид, Рис, Ан, Ст |
| Grateloupiaturuturu Yamada | нл, вс | + | эл | П |
| Порядок Ahnfeltiales Семейство Ahnfeltiaceae |
||||
| Ahnfeltiatobuchiensis (Kanno et Matsubara) Makienko | с | ++ | нф | Вод |
| Порядок Gracilariales Семейство Gracilariaceae |
||||
| Gracilariopsislongissima (S.G. Gmelin) M. Steentoft, L.M. Irvine & W.F. Farnham | нл, вс | + | эл, эпп | Ид |
| Порядок Palmariales Семейство Palmariaceae |
||||
| Palmariastenogona Perestenko | нл, вс | + | эл | Ид, Рис, Вод |
| Halosaccionglandiforme (S.G. Gmelin) Ruprecht | нл, вс | ед | эл | Рис |
| Порядок Ceramiales Семейство Callithamniaceae |
||||
| Ptilotafilicina J. Agardh | нл, вс | + | эл, эпп | Ид, Рис |
| Семейство Ceramiaceae | ||||
| Campylaephora crassa (Okamura) Nakamura | нл, вс | + | эл, эпп, эф | Ид, Рис |
| Campylaephorahypnaeoides J. Agardh | вл, нл, вс, сл | + | эл, эпп, эф | Ид |
| Ceramium cimbricum H.E. Petersen | нл, вс | + | эпп, эф | П |
| Ceramiumjaponicum Okamura | нл, вс | + | эл, эпп | Рис |
| Ceramiumkondoi Yendo | нл, вс | ++ | эл, эпп | П |
| Семейство Dasyaceae | ||||
| Dasyasessilis Yamada | нл, вс | + | эл, эпп | Ид, Вод, Рис |
| Dasysiphonia japonica (Yendo) H.-S. Kim | нл, с | + | эл, эпп | Ан |
| Семейство Delesseriaceae | ||||
| Nienburgia angusta A.D. Zinova | c | + | эф, эл | Вод |
| Cumathamnion serrulatum (Harvey) M.J. Wynne & G.W. Saunders | нл, вс | + | эл, эпп | Ид |
| Phycodrys riggii N.L. Gardner | с | ед | эл, эпп | Ид |
| Семейство Rhodomelaceae | ||||
| Chondria dasyphylla (Woodward) C. Agardh | нл, лл | + | эл, эф | Рис |
| Chondriadecipiens Kylin | нл, вс | + | эл, эпп | Ид, Вод, Рис |
| Laurencianipponica Yamada | нл, вс | ++ | эл | П |
| Laurencia pinnata Yamada | нл, вс | + | эл | Рис |
| Janczewskia morimotoi Tokida | нл, вс | + | пп (L. nipponica) | Сл |
| Neorhodomelaaculeata (L.P. Perestenko) Masuda | нл, вс | ++ | эл, эпп | П |
| Neorhodomelamunita (L.P. Perestenko) Masuda | нл, вс | + | эл, эпп | Рис |
| Melanothamnusjaponicus (Harvey) Diaz-Тapia & Maggs | вл, нл, вс, cл | +++ | эл, эпп, эф | П |
| Polysiphoniamorrowii Harvey | вл, нл, лл, вс, сл | ++ | эл, эпп | Вод, Рис, Ан |
| Polysiphonia yendoi Segi | нл, вс | + | эл | Рис |
| Symphyocladialatiuscula (Harvey) Yamada | нл, с | + | эл, эпп | Ид |
| Symphyocladiamarchantioides (Harvey) Falkenberg | нл, вс | + | эл, эф | П |
| Порядок Rhodymeniales Семейство Champiaceae |
||||
| Champia parvula (C. Agardh) Harvey | нл | ед | эл, эф | Ид |
| Семейство Lomentariaceae | ||||
| Lomentariahakodatensis Yendo | нл, вс | + | эл | Ид, Рис, Ан |
| Семейство Rhodymeniaceae | ||||
| Chrysymeniawrightii (Harvey) Yamada | нл, вс | ед | эл, эпп | Рис |
| Sparlingiapertusa (Postels et Ruprecht) G.W. Saunders, I.M. Strachan et Kraft | вс | ед | эл, эпп | Рис |
| Отдел Ochrophyta | ||||
| Класс Phaeophyceae Порядок Ectocarpales Семейство Ectocarpaceae |
||||
| Ectocarpus siliculosus (Dillwyn) Lyngbye | нл, вс | ед | эл, эпп, эф | Ид |
| Семейство Chordariaceae | ||||
| Acrothrix pacifica Okamura et Yamada | нл | ед | эл, эф | Ид |
| Chordariaflagelliformis (O.F. Müller) C. Agardh | нл, вс | ++ | эл | Рис, Ан, Сл, Ст |
| Coilodesme japonica Yamada | вс | + | эф (S. crassipes) | Рис |
| Sphaerotrichia divaricata (C. Agardh) Kylin | нл, вс | ++ | эл, эпп | Вод, Ид |
| Dictyosiphonchordaria Areschoug | нл, вс | + | эл, эпп | Вод |
| Halothrix lumbricalis (Kützing) Reinke | нл | + | эл, эпп, эф | Ид |
| Punctariaplantaginea (Roth) Greville | лл, вл, сл, нл, вс | ++ | эл, эпп, эф | Вод, Рис, Ан |
| Leathesiamarina (Lyngbye) Decaisne | вл, лл | ++ | эл, эф | П |
| Saundersella simplex (De A. Saunders) Kylin | вл, нл, вс | + | эф (Chordariа) | Рис, Ан |
| Семейство Scytosiphonaceae | ||||
| Melanosiphon intestinalis (De A. Saunders) M.S. Wynne | вл | + | эл | Ид, Рис |
| Colpomeniaperegrina Sauvageau | лл, вл, сл, нл, вс | + | эл, эф | Рис |
| Scytosiphonlomentaria (Lyngbye) Link | вл, нл, сл, лл, вс | ++ | эл, эпп | Рис, Ан, Вод, Ст, Сл |
| Порядок Ralfsiales Семейство Ralfsiaceae |
||||
| Analipusjaponicus (Harvey) M.J. Wynne | нл, вс | ++ | эл | Ид, Рис, Ан |
| Ralfsiafungiformis (Gunnerus) Setchell et N.L. Gardner | нл, вс | ++ | эл | Ид, Вод, Рис |
| Ralfsia verrucosa (Areschoug) Areschoug | вл, сл | ++ | эл | Ид, Ст, Сл |
| Порядок Desmarestiales Семейство Desmarestiaceae |
||||
| Desmarestialigulata (Stackhouse) J.V. Lamouroux | вс | + | эл | Рис, Ан |
| Desmarestiaviridis (O.F. Müller) J.V. Lamouroux | нл, вс | +++ | эл, эпп | П |
| Порядок Laminariales Семейство Chordaceae |
||||
| Chorda filum (Linnaeus) Stackhouse | нл, вс | ++ | эл, эпп | Ид, Рис, Ан, Вод |
| Семейство Pseudochordaceae | ||||
| Pseudochorda nagaii (Tokida) Inagaki | нл | ед | эл | Ид |
| Семейство Agaraceae | ||||
| Agarumclathratum Dumortier | вс | + | эл, эпп | Рис |
| Costariacostata (C. Agardh) De A. Saunders | вл, нл, сл, лл, вс | + | эл, эпп | П |
| Семейство Laminariaceae | ||||
| Saccharinaangustata (Kjellman) C.E. Lane, C. Mayes, Druehl & G.W. Saunders | нл, вс | + | эл | Ст |
| Saccharinacichorioides (Miyabe) C.E. Lane, C. Mayes, Druehl et G.W. Saunders | с | ++ | эл, эпп | Рис, Ан, Вод |
| Saccharinajaponica (Areschoug) C.E. Lane, C. Mayes, Druehl & G.W. Saunders | с | + | эл | Ид, Вод |
| Семейство Alariaceae | ||||
| Undariapinnatifida (Harvey) Suringar | нл, с | + | эл | Ан, Сл |
| Порядок Sphacelariales Семейство Sphacelariaceae |
||||
| Sphacelariarigidula Kützing | вл, нл, лл, сл | + | эл, эпп, эф | Ид, Рис |
| Порядок Dictyotales Семейство Dictyotaceae |
||||
| Dictyotadichotoma (Hudson) J.V. Lamouroux | нл, вс | ++ | эл | Ид, Рис, Ан |
| Dictyopterisdivaricata (Okamura) Okamura | нл | + | эл | Ид, Ан, Рис |
| Порядок Fucales Семейство Sargassaceae |
||||
| Stephanocystis crassipes (Mertens ex Turner) Draisma, Ballesteros, F. Rousseau et T. Thibaut | нл, вс | + | эл | Рис, Ан |
| Sargassummiyabei Yendo | вс | +++ | эл, эпп | П |
| Sargassumpallidum (Turner) C. Agardh | нл, вс | + | эл, эпп | Рис, Ан |
| Coccophoralangsdorfii (Turner) Greville | нл, с | + | эл, эпп | Рис |
| Семейство Fucaceae | ||||
| Fucusevanescens C. Agardh | вл, лл | + | эл | Ан |
| Silvetiababingtonii (Harvey) E.A. Serrão, T.O. Cho, S.M. Boo & Brawley | вл, лл | + | эл | Ан |
| Отдел Chlorophyta | ||||
| Класс Ulvophyceae Порядок Ulotrichales Семейство Ulotrichaceae |
||||
| Urospora penicilliformis (Roth) Areschoug | вл | + | эл, эф | Ид |
| Семейство Gomontiaceae | ||||
| Monostromagrevillei (Thuret) Wittrock | нл, вс | + | эл, эф | Ид |
| Порядок Ulvales Семейство Ulvaceae |
||||
| Ulva clathrata (Roth) C. Agardh | нл, вс | + | эл | Рис |
| Ulvaflexuosa Wulfen | нл, вс | + | эл, эф | Рис |
| Ulvalactuca Linnaeus | вс, лл, сл, нл, вс | +++ | эл, эпп, эф | П |
| Ulva linza Linnaeus | нл, вс | ++ | эл, эпп | Ид, Рис, Вод |
| Ulvaprolifera O.F. Müller | вл, лл, сл, нл | + | эл, эпп, эф | Ид, Рис, Вод |
| Ulvariasplendens (Ruprecht) Vinogradova | нл, вс | + | эл, эпп | Ид |
| Порядок Cladophorales Семейство Cladophoraceae |
||||
| Cladophora opaca Sakai | лл, вл, сл, нл | + | эл, эф | Рис |
| Cladophorastimpsonii Harvey | нл, вс | ++ | эл, эпп, эф | Ид, Рис, Вод |
| Chaetomorphalinum (O.F. Müller) Kützing | лл | ед | эл, эпп | Ид |
| Chaetomorpha moniligera Kjellman | вл, лл, нл, сл | ++ | эл | Ан, Вод, Рис, Ид |
| Порядок Bryopsidales Семейство Bryopsidaceae |
||||
| Bryopsisplumosa (Hudson) C. Agardh | нл, вс | + | эл, эпп | Ан |
| Семейство Codiaceae | ||||
| Codiumfragile (Suringar) Hariot | нл, вс | + | эл, эпп, эф | Ид |
| Codiumyezoense (Tokida) K.L. Vinogradova | нл, вс | + | эл, эпп | Ид |
| Отдел Tracheophyta | ||||
| Класс Monocots
Порядок Alismatales Семейство Zosteraceae |
||||
| Zostera marina Linnaeus | нл, с | ++ | эпп | Ан, Вод, Ид, Рис |
| Zostera asiatica Miki | нл, с | + | эпп | Ан |
| Phyllospadix iwatensis Makino | с | + | эл | Ан, Рис |
Примечание. Зона произрастания: вл – верхняя литораль, сл – средняя литораль, нл – нижняя литораль, вс – верхняя сублитораль, с – сублитораль, лл – литоральные лужи; обилие видов: (+) – редкие, (++) – обычные, (+++) – массовые, ед – единичные; экологическая группировка: эф – эпифит, эл – эпилит, эпп – эпипелит, пп – полупаразит, нф – неприкрепленная форма; место произрастания: Ид – б. Идол, Рис – б. Рисовая, Ан – мыс Андреева, Вод – водолазная станция, Сл – мыс Слычкова, Ст – мыс Стенина, П – повсеместно.
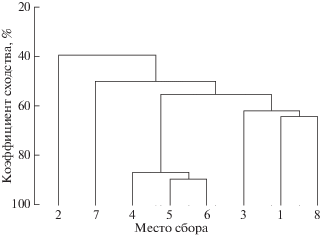
Сходство видового состава, определенное с помощью коэффициента Серенсена–Чекановкого, наблюдалось между мысами Слычкова (точки 4 и 5) и Стенина (точка 6) и составляло 87–89%. Результаты иерархической кластеризации показали, что на уровне 55% сходства все исследованные участки сгруппировались в 4 кластера (рис. 3). Самый большой кластер на уровне 87–89% сходства видового состава объединил точки 4, 5 и 6. Точки 1, 3 и 8 также образовали один кластер на уровне сходства 62–64%. Точки 2 и 7 – это отдельные кластеры на уровне сходства 24%.
Рис. 3.
Дендрограмма сходства видового состава макрофитов б. Троицы по коэффициенту Серенсена–Чекановского. 1–8 – точки.
В исследованном районе доминировали макрофиты, растущие на твердом субстрате (эпилиты), – 100 видов (90% от всех собранных). Водоросли этой группы росли на скалах, крупных валунах, а также на мелких камнях и створках моллюсков, замытых песком. На мягких грунтах (эпипелиты) отмечены морские травы Zostera marina и Z. asiatica. В точке 4 (мыс Слычкова) найдена полупаразитическая водоросль Janczewskia morimotoi, живущая на красной водоросли Laurencia nipponica. В центральной части бухты в районе антициклонического течения на илистом грунте располагалось поле неприкрепленной к грунту красной водоросли Ahnfeltia tobuchiensis. На твердом грунте и эпифитно обитали 23 вида; только эпифитно встречались 5 видов: Erythrotrichia carnea, Hydrolithon farinosum, Pneophyllum zostericola, Saundersella simplex и Coilodesme japonica (табл. 2).
Распространение водорослей и их сообществ от верхней литорали до верхней сублиторали
Наибольшее число видов (90) найдено в нижней литорали и верхней сублиторали (неосушной зоне), 32 вида обнаружено в верхней и средней литорали. Во всех приливо-отливных зонах присутствовали красные водоросли Pachyarthron cretaceum, Corallina pillulifera, Chondrus pinnulatus, Mazzaella japonica, Polysiphonia morrowii, Grateloupia divaricata, бурые водоросли Punctaria plantaginea, Colpomenia peregrina, Costaria costata, Scytosiphon lomentaria и зеленая водоросль Ulva lactuca (табл. 2).
На основе полученных данных выделено 23 растительных сообщества (табл. 1).
Точка 1, мыс Андреева (табл. 1, рис. 2а). В верхней литорали (глубина 0–0.5 м при полном приливе) отвесные скалы были заселены монодоминантным сообществом Nemalion vermiculare, ниже располагался пояс монодоминантного сообщества C. pillulifera (рис. 4а), сменявшийся бидоминантным сообществом C. pillulifera + Neorhodomela aculeata и/или монодоминантным сообществом Mastocarpus pacificus с сопутствующим видом Ahnfeltiopsis flabelliformis. В нижней литорали и в верхней сублиторали твердый субстрат был занят мозаичным полидоминантным сообществом с доминированием Dictyota dichotoma, Chondrus armatus, Chordaria flagelliformis и N. aculeata.
Рис. 4.
Примеры водорослевых сообществ, выделенных на литорали и в верхней сублиторали б. Троицы. а – монодоминантное сообщество Corallina pillulifera (мыс Андреева); б – бидоминантное сообщество Leathesia marina (Lm) + Nemalion vermiculare (Nv) (б. Идол); в – полидоминантное сообщество (водолазная станция); г – монодоминантное сообщество Ralfsia verrucosa (Rv) и бидоминантное сообщество Corallina pilulifera (Cp) + Chordaria flagelliformis (Cf) (мысы Слычкова и Стенина).

Точки 2 и 3, водолазная станция (табл. 1, рис. 2б). В точке 2 в верхней литорали скалисто-каменистый грунт был занят монодоминантным сообществом красной водоросли Gloiopeltis furcata, а в средней и нижней литорали – бидоминантным сообществом P. morrowii + Desmarestia viridis с сопровождающими видами. В точке 3 в верхней литорали выделено полидоминантное сообщество с доминированием U. lactuca, Leathesia marina и P. plantaginea (рис. 4в). В наиболее защищенных местах верхней литорали (точки 2 и 3) каменистый грунт был занят бидоминантным сообществом Ulva linza + Cladophora stimpsonii; в верхней сублиторали на каменистом грунте преобладало бидоминантное сообщество P. plantaginea + U. lactuca, а на песчаном грунте – монодоминантное сообщество морской травы Z. marina, среди которой встречались Sargassum myabei, Chorda filum и др.
Точки 4, 5 (мыс Слычкова) и точка 6 (мыс Стенина) (табл. 1, рис. 2в) характеризовались сходной картиной распределения макрофитов. На литорали обоих мысов прослеживалась четкая поясность. На скалисто-каменистом субстрате самый верхний пояс (верхняя литораль) занимало монодоминантное сообщество, сформированное Ralfsia verrucosa (Rv) (рис. 4г), с вкраплениями L. marina и Nemalion vermiculare. В средней и нижней литорали располагалось бидоминантное сообщество Corallina pillulifera (Cp) + Chordaria flagelliformis (Cf) (рис. 4г) с мозаичным включением N. vermiculare и Scytosiphon lomentaria. Cамый нижний пояс литорали был представлен монодоминантным сообществом L. nipponica с сопутствующим видом Melanothamnus japonica. В верхней сублиторали (точка 5) на отдельных валунах отмечено полидоминантное сообщество с мозаичным доминированием C. costata, Undaria pinnatifida, S. lomentaria, M. japonicus и др. В сублиторали макрофиты были представлены бедно; встречались отдельно растущие экземпляры U. pinnatifida, C. costata, D. viridis и др. На мысе Стенина (точка 6) в нижней литорали на отвесных скалах располагалось монодоминантное сообщество Saccharina angustata.
Точка 7, б. Идол (табл. 1, рис. 2г). Верхняя литораль бухты была занята монодоминантными сообществами G. furcata и R. verrucosa, а также бидоминантным сообществом L. marina (Lm) + + N. vermiculare (Nv) (рис. 4б). В средней и нижней литорали располагалось полидоминантное сообщество с доминированием S. miyabei, D. dichotoma, Dictyopteris divaricata и сопровождающими видами. В верхней сублиторали преобладало бидоминантное сообщество S. miyabei + Chorda filum с сопровождающими видами Lithotamnion phymatodeum, D. dichotoma, D. divaricata и L. nipponica.
Точка 8, б. Рисовая (табл. 1, рис. 2д). В верхней литорали на валунах располагалось монодоминантное сообщество G. furcatа, а ниже – монодоминантное сообщество Analipus japonicus. В верхней и средней литорали встречалось монодоминантное сообщество Chaetomorpha moniligera с сопутствующими видами Ulva lactuca, U. сlathrata и U. flexuosa, а в средней и нижней литорали – полидоминантное сообщество с доминированием M. japonicus, A. japonicus, Sargassum spp., Chondrus spp., N. aculeata, Ulva spp., Ch. flagelliformis и др. На границе нижней литорали и верхней сублиторали доминировало монодоминантное сообщество S. miyabei, в котором часто встречались Ulva spp., N. aculeatа и другие виды.
ОБСУЖДЕНИЕ
С мая по сентябрь в 2004–2018 гг. на литорали и в верхней сублиторали б. Троицы обнаружено 108 видов макроводорослей (54% красных, 32% бурых, 14% зеленых) и 3 вида морских трав. Собранные макрофиты составили 34% от всех известных для зал. Петра Великого видов (сборы разных авторов с 1965 по 2018 г.) (Скрипцова, 2019). По богатству видов флора б. Троицы близка к соседней б. Витязь (101 вид) (Кафанов, Жуков, 1993), также относящейся к зал. Посьета.
По видовому составу и богатству флора исследованной акватории сходна с флорой других акваторий Японского моря, расположенных на этой же широте. Так, на о-ве Хоккайдо в б. Учиура-ван (Uchiura-wan) в районе Института альгологических исследований (Institute of Algological Research) в 1980-х годах зарегистрировано 194 вида водорослей, из которых красные водоросли составляли 57%, бурые – 30% и зеленые – 13% (Sakai, 1986). Семьдесят один вид водорослей (66% нашей коллекции) – общие с коллекцией водорослей из б. Учиура-ван, что указывает не только на значительное сходство флор, но и на бóльшее богатство флоры западного побережья о-ва Хоккайдо по сравнению с таковым южных бухт зал. Петра Великого.
Сравнительный анализ видового состава водорослей в разных точках б. Троицы показал, что близкие биотопы имеют сходный набор водорослей. Так, сходство видового состава водорослей на входных мысах Стенина и Слычкова (характеризующихся высоким водообменном и скальным грунтом) составило около 90%. Наибольшее число видов найдено вблизи внутренних мысов бухты (точка 1 – мыс Андреева и точка 2 – камни у водолазной станции), а также на выходах коренных пород в б. Рисовая (точка 8). Схожесть флоры в этих точках составила более 60%. Наиболее бедной флора была на песчаных грунтах с редким выходом коренных пород или присутствием камней. На литорали и в верхней сублиторали бухты выделено 11 моно-, 7 би- и 5 полидоминатных растительных сообществ.
На литорали входных мысов (точки 4 и 6) в распределении монодоминантных и бидоминантных сообществ наблюдалась поясность, отсутствующая на других участках литорали и сублиторали. В центральной части бухты располагалось поле Ahnfeltia tobuchiensis, которое было отмечено ранее (Некрасов и др., 2000; Галышева, Христофорова, 2008).
Литоральные сообщества б. Троицы были характерны для зал. Посьета, однако в этой бухте отсутствовали такие монодоминантные сообщества, как M. japonicus [= Polysiphonia japonica], Ulva clathrata [Enteromorpha clathrata], Palmaria stenogona, Neorhodomela aculeata [= Rhodomela larix], и бидоминантные сообщества Leathesia difformis + + C. pillulifera, L. nipponica + C. pillulifera, указанные ранее для этого залива (Перестенко, 1980). В то же время в зал. Посьета не обнаружены выделенные нами в б. Троицы монодоминантные сообщества S. angustata, Ch. moniligera, L. nipponica, C. pillulifera и бидоминантные сообщества C. pillulifera + N. aculeata, P. morrowii + D. viridis, Ulva linza + + C. stimpsonii, P. plantaginea + U. lactuca.
Bидовой состав водорослей в б. Витязь незначительно отличался от такового в б. Троицы; здесь выделено 5 фитоценозов: A. japonicus, Ch. flagelliformis + Coccophora langsdorfii, C. langsdorfii + + P. stenogona, Chondria decipiens + C. langsdorfii и Neorhodomela larix (Кафанов, Жуков, 1993). Небольшое количество выделенных фитоценозов можно объяснить тем, что исследованы лишь три точки в вершинной части б. Витязь. Эти фитоценозы по доминирующим видам совпадают с выделенными нами моно- и бидоминантными сообществами в точке 8 (б. Рисовая).
В заключение следует отметить, что бентосная флора б. Троицы по богатству и видовому составу водорослей-макрофитов близка к таковой соседних южных акваторий зал. Петра Великого и по видовому составу более чем на 60% сходна с флорой бухт западного побережья о-ва Хоккайдо, расположенных на этой же широте. Богатство видового состава водорослей и разнообразие условий для их вегетации предопределяют формирование более чем 20 моно-, би- и полидоминантных сообществ.
Список литературы
Атлас океанов. Тихий океан. М.; Л.: Гл. упр. навигации и океаногр. МО РФ. 1974. 302 с.
Белоус О.С., Дроздов А.Л. Бухта Троицы (залив Петра Великого, Японское море): физико-географическая характеристика, макробентос // Биота и среда заповедников Дальнего Востока. 2015. № 1. С. 3–12.
Белоус О.С., Титлянова Т.В., Титлянов Э.А. Морские растения бухты Троицы и смежных акваторий (залив Петра Великого, Японское море). Владивосток: Дальнаука. 2013. 264 с.
Виноградова К.Л. Определитель водорослей дальневосточных морей СССР. Зеленые водоросли. Л.: Наука. 1979. 146 с.
Вышкварцев Д.И., Пешеходько В.М. Картирование доминирующих видов водной растительности и анализ их роли в экосистеме мелководных бухт залива Посьета Японского моря // Подводные гидробиологические исследования. Владивосток: Изд-во ДВНЦ АН СССР. 1982. С. 120–130.
Галышева Ю.А., Христофорова Н.К. Состав и количественное распределение макробентоса сублиторали бухты Троицы залива Посьета // Материалы науч. конф., посвящ. 70-летию С.М. Коновалова “Современное состояние водных биоресурсов”. 24–26 марта 2008 г., г. Владивосток. Владивосток: ТИНРО. 2008. С. 45–51.
Григорьева Н.И., Федосеев В.Я., Кучерявенко А.В. Абиотические условия среды в местах размещения плантаций марикультуры залива Посьета (залив Петра Великого, Японское море) // Изв. ТИНРО. 2001. Т. 128. С. 501–514.
Гурьянова Е.Ф., Закс И.Г., Ушаков П.В. Литораль Кольского залива. Ч. III // Тр. Ленингр. о-ва естествоиспыт. 1930. Т. LX. Вып. 2. С. 17–107.
Кафанов А.И., Жуков В.Е. Водоросли-макрофиты бухты Витязь (Японское море): видовое обилие и разнообразие в гетерогенной среде. Препринт № 16 / Институт биологии моря ДВНЦ АН СССР. Владивосток: ДВНЦ АН СССР. 1986. 20 с.
Кафанов А.И., Жуков В.Е. Прибрежное сообщество водорослей-макрофитов залива Посьета (Японское море). Владивосток: Дальнаука. 1993. 156 с.
Лебедев Е.Б., Левенец И.Р., Вышкварцев Д.И. Донные сообщества бухты Миноносок залива Посьет (Японское море) // Изв. ТИНРО. 2004. Т. 137. С. 378–392.
Лоция Японского моря. Северо-западная часть моря от реки Тюмень-Ула до мыса Белкина. Ч. I. 1972. 287 с.
Некрасов Д.А., Чербаджи И.И., Попова Л.И. Продукционные показатели популяции анфельции тобучинской в бухте Баклан залива Петра Великого Японского моря // Биол. моря. 2000. Т. 26. № 4. С. 259–264.
Паймеева Л.Г. Характеристика зарослей и состояние запасов зостеры в юго-западной части залива Петра Великого от бухты Бойсмана до бухты Сивучей // Изв. ТИНРО. 1972. Т. 93. С. 153–161.
Перестенко Л.П. Водоросли залива Посьета (Японское море). 1 // Новости системат. низш. раст. 1968. Т. 5. С. 48–53.
Перестенко Л.П. Водоросли залива Посьета, новые для флоры Южного Приморья и советских берегов Японского моря // Исслед. фауны морей. 1971а. Т. VIII (XVI). С. 7–21.
Перестенко Л.П. Список флоры и фауны залива Посьета Японского моря. Отделы Chlorophyta, Phaeophyta, Rhodophyta, Embryophyta-Siphonogama // Исслед. фауны морей. 1971б. Т. VIII (XVI). С. 303–305.
Перестенко Л.П. Водоросли залива Петра Великого. Л.: Наука. 1980. 232 с.
Перестенко Л.П. Красные водоросли дальневосточных морей России. СПб.: Ольга. 1994. 330 с.
Песенко В.Ф. Принципы и методы количественного анализа в фаунистических исследованиях. М.: Наука. 1982. 287 с.
Скрипцова А.В. Водоросли-макрофиты залива Петра Великого Японского моря // Биота и среда заповедных территорий. 2019. № 3. С. 14–57.
Титлянов Э.А., Титлянова Т.В., Белоус О.С. Полезные морские растения стран Азиатско-Тихоокеанского региона. Владивосток: Дальнаука. 2016. 360 с.
Титлянов Э.А., Титлянова Т.В., Ли Ш., Хуанг Х. Инвентаризация морских бентосных макроводорослей острова Хайнань (Китай) // Биол. моря. 2018. Т. 44. № 3. С. 145–154.
Clarke K.R., Gorley R.N. PRIMER version 6: User manual/tutorial. Plymouth, U.K: PRIMER-E. 2006. 192 p.
Guiry M.D., Guiry G.M. AlgaeBase. World-wide electronic publication. National University of Ireland, Galway. 2019. http://www.algaebase.org. Accessed January 15, 2019.
Sakai Y. A list of marine algae from the vicinity of the institute of algological research of Hokkaido University, Muroran, Japan // Sci. Pap. Inst. Algol. Res., Fac. Sci., Hokkaido Univ. 1986. V. 8. № 1. P. 1–30.
Saunders G.W., Hommersand M.H. Assessing red algal supraordinal diversity and taxonomy in the context of contemporary systematic data // Am. J. Bot. 2004. V. 91. № 10. P. 1494–1507.
Tsuda R.T. Checklist and bibliography of the marine benthic algae from the Mariana Islands (Guam and CNMI), University of Guam Marine Laboratory Technical Reptorts. № 107. Mangilao, Guam: Univ. of Guam Mar. Lab. 2003. P. 1–37.
Дополнительные материалы отсутствуют.
Инструменты
Биология моря