

Биология моря, 2021, T. 47, № 4, стр. 228-234
Смертность икринок минтая Gadus chalcogrammus Pallas, 1814 в течение эмбриогенеза в водах западного побережья Камчатки
А. В. Буслов a, *, А. В. Смирнов a
a Тихоокеанский филиал ФГБНУ “ВНИРО” (ТИНРО)
690091 Владивосток, Россия
* E-mail: aleksandr.buslov@tinro-center.ru
Поступила в редакцию 03.06.2020
После доработки 01.02.2021
Принята к публикации 10.02.2021
Аннотация
В апреле–мае 2019 г. в период массового икрометания минтая Gadus chalcogrammus на нерестилище у западного побережья Камчатки было выполнено пять последовательных ихтиопланктонных съемок. На основе оценок численности икры минтая получены величины ее суточной продукции для каждой из съемок. Рассчитаны общая за нерестовый сезон продукция и смертность икры в процессе эмбриогенеза. Максимальные коэффициенты смертности определены для икринок на I и III стадиях развития. Общая убыль составила 86.8% от числа выметанных икринок. Полученные результаты позволяют существенно улучшить качество оценок нерестового запаса охотоморского минтая.
В водах шельфа западного побережья Камчатки функционирует одно из крупнейших в бассейне Охотского моря нерестилищ минтая Gadus chalcogrammus (Pallas, 1814) (Фадеев, 1987; Фадеев, Смирнов, 1987; Шунтов и др., 1993). В этом районе ежегодно проводятся ихтиопланктонные съемки по учету развивающейся икры с целью оценки численности производителей (Фадеев, 1999). Одним из важных параметров, который следует использовать при интерпретации результатов ихтиопланктонных съемок у западной Камчатки, является оценка убыли икринок, выметанных до начала учетов. Предполагается, что игнорирование этого показателя может быть причиной значительного занижения численности нерестового запаса (Кузнецов и др., 2008). Проблема смертности икры минтая в процессе эмбриогенеза до настоящего времени недостаточно изучена. Существует мнение, что в водах западной Камчатки на начальной стадии развития (первые 5–8 сут) погибает 74–85% выметанных икринок минтая, а неучтенная убыль может приводить к недооценке общего количества выметанной икры в 2–10 раз (Лисовенко, 2000; Привалихин и др., 2001). При этом авторы отмечают, что приведенные цифры весьма приблизительны, поскольку для более точной оценки выживаемости икринок необходимо проведение повторных ихтиопланктонных съемок с разницей во времени, близкой к продолжительности одной стадии развития, чего не обеспечивали имевшиеся у них материалы. Цель настоящего исследования – оценка смертности развивающихся икринок минтая как на отдельных стадиях развития, так и за весь период эмбриогенеза у западного побережья Камчатки.
МАТЕРИАЛ И МЕТОДИКА
В апреле–мае 2019 г. у западного побережья Камчатки на научно-исследовательском судне “Дмитрий Песков” было выполнено пять ихтиопланктонных съемок по стандартной сетке станций (67 станций), охватывающей площадь 53 тыс. км2 (табл. 1). Полигон был ограничен координатами 51°00′–57°00′ с.ш., изобатой 20 м с востока и 154°30′ в.д. с запада. Исследования проводили по широтным разрезам с юга на север. При завершении съемки в крайней северной точке судно возвращалось на южный разрез и приступало к следующей съемке. Вертикальные обловы ихтиопланктона выполняли сетью ИКС-80 в слое дно – поверхность (над изобатами менее 200 м) или 200 м – поверхность (над изобатами глубже 200 м). Для учета икринок от возможного глубоководного нереста минтая в южной части полигона (южнее 52° с.ш.) облавливали слой 500–0 м (Буслов, Тепнин, 2002). Температуру воды в слое облова измеряли через 1 м гидрологическим зондом “SBE-19”.
Таблица 1.
Сроки проведения ихтиопланктонных съемок у западного побережья Камчатки в 2019 г.
| № съемки | Начало | Окончание | Средняя дата съемки |
|---|---|---|---|
| 1 | 1 апреля | 5 апреля | 3 апреля |
| 2 | 8 апреля | 13 апреля | 11 апреля |
| 3 | 15 апреля | 23 апреля | 19 апреля |
| 4 | 24 апреля | 30 апреля | 27 апреля |
| 5 | 1 мая | 7 мая | 4 мая |
Икринки минтая выбирали из улова и подсчитывали, определяли стадии их развития и фиксировали в 4% растворе формальдегида. Стадии развития икринок определяли с помощью бинокуляра “Olimpus SZ30” по четырехбалльной шкале Расса, адаптированной для минтая (Расс, Казанова, 1966; Буслов, Сергеева, 2013). С этой целью отбирали 200 икринок, в случае меньших уловов просматривали все икринки.
Численность икринок на акватории съемок рассчитывали с помощью метода сплайн-аппроксимации, реализованного в геоинформационной системе “КАРТМАСТЕР” (Бизиков, Поляков, 2004).
Продолжительность стадий развития икринок в зависимости от температуры воды определяли по уравнениям, подобранным для каждой стадии (Буслов, Сергеева, 2013). Среднюю температуру воды, при которой развивались икринки в период съемок, определяли для каждой стадии. При расчете возраста икринок и даты начала нереста приняли, что в момент поимки икринки находились в середине соответствующей стадии развития.
Для каждой съемки рассчитывали суточную продукцию икринок (P) по стадиям развития как отношение учтенного количества икринок (N) i-й стадии на всей акватории съемки к продолжительности этой стадии (Лисовенко, 2000). При моделировании кривых суточной продукции допустили, что полученные величины приходятся на среднюю дату проведения съемки.
Поскольку динамика суточной продукции икринок минтая в течение нерестового периода пропорциональна плотности нормального распределения (Антонов, Золотов, 1987; Picquelle, Megrey, 1993; Буслов и др., 2004), для построения кривой суточной продукции икринок использовали функцию нормального распределения Гаусса: y = bexp(h(x – a)2), где y – суточная продукция икры (×1012 экз.); х – дни от начала нереста (сут); a – продолжительность нерестового периода до пика (сут); b – высота пика, выражающаяся через общую продукцию (×1012 экз.); h – коэффициент, определяющий изгиб кривой. Коэффициент рассчитан по методу избранных точек посредством минимизации суммы квадратов отклонений модельных оценок суточной продукции от наблюденных (Зайцев, 1984).
Абсолютную величину общей убыли определяли как разницу между суммарной продукцией икринок предыдущей и последующей стадий (Лисовенко, 2000). При этом на продукционной кривой предыдущей стадии отсекали последний временной отрезок, который необходим икринкам для перехода с середины одной стадии до середины следующей стадии.
При расчетах мгновенных (суточных) коэффициентов убыли (Z) исходили из условия, что смертность за весь период имеет экспоненциальный характер. Суточную выживаемость икринок на i-й стадии развития определяли как корень в степени Ni – 1 из выживаемости за период прохождения i-й стадии (Ni – длительность i-й стадии при зарегистрированной температуре воды). Численное представление смертности (А) через выживание (S) выражали отношением: Nt/N0= S = 1 – A, где N0 – начальная численность икринок, а Nt – численность икринок через период времени t (Рикер, 1979).
Информация о средней плодовитости самок и средней массе производителей минтая в нерестовый период у западного побережья Камчатки в 2019 г. любезно предоставлена заведующим Лабораторией минтая и сельди Тихоокеанского филиала ФГБНУ “ВНИРО” (“ТИНРО”) Е.Е. Овсянниковым.
РЕЗУЛЬТАТЫ
Средняя температура воды в период проведения съемок изменялась в узких пределах. Развитие икринок проходило преимущественно при положительных значениях температуры около 1°С, за исключением первых трех съемок, когда икринки на I стадии облавливались при слабоотрицательной температуре воды (табл. 2). Продолжительность стадий развития варьировала незначительно, размах колебаний между разными съемками, как правило, не превышал 1.5 сут. Поимка икринок на IV стадии развития во время 1-й съемки дает основание считать, что нерест минтая в 2019 г. начался не позднее 1 марта. Даты появления в ихтиопланктоне икринок на II, III и IV стадиях развития, исходя из их продолжительности, пришлись на 10, 18 и 29 марта.
Таблица 2.
Средняя температура воды (t, °С) и длительность (T, сут) стадий развития икринок минтая (I−IV) у западного побережья Камчатки в 2019 г.
| № съемки | I | II | III | IV | ||||
|---|---|---|---|---|---|---|---|---|
| t | T | t | T | t | T | t | T | |
| 1 | –0.29 | 9.4 | 0.32 | 7.9 | 0.61 | 10.9 | 0.60 | 10.0 |
| 2 | –0.21 | 9.2 | 0.25 | 8.0 | 0.47 | 11.2 | 0.58 | 10.1 |
| 3 | –0.04 | 8.9 | 0.09 | 8.3 | 0.43 | 11.3 | 1.49 | 7.3 |
| 4 | 0.40 | 8.0 | 0.18 | 8.1 | 0.10 | 12.1 | 0.46 | 10.5 |
| 5 | 0.55 | 7.9 | 0.53 | 7.6 | 0.49 | 11.1 | 0.75 | 9.6 |
Численность икринок на I стадии развития, характеризующей интенсивность нереста, увеличивалась в первой половине апреля и снижалась во второй половине месяца (табл. 3). Значения суточной продукции икры на этой стадии свидетельствуют о том, что исследования включали период нарастания массового икрометания, его разгар и снижение. Пик нереста пришелся на третью съемку, т.е. на 19 апреля. Численность и суточная продукция икринок на II–IV стадиях увеличивались от первой к последней съемке.
Таблица 3.
Численность (N, ×1012 экз.) и суточная продукция (Р, ×1012 экз.) икринок минтая разных стадий развития (I−IV) у западного побережья Камчатки в 2019 г.
| № съемки | I | II | III | IV | ||||
|---|---|---|---|---|---|---|---|---|
| N | P | N | P | N | P | N | P | |
| 1 | 73.7 | 7.8 | 11.8 | 1.5 | 2.0 | 0.2 | 0.02 | 0.002 |
| 2 | 95.8 | 10.4 | 27.0 | 3.4 | 7.7 | 0.7 | 0.1 | 0.01 |
| 3 | 96.0 | 10.8 | 54.1 | 6.5 | 30.9 | 2.7 | 4.0 | 0.6 |
| 4 | 64.6 | 8.1 | 60.3 | 7.4 | 41.8 | 3.5 | 8.3 | 0.8 |
| 5 | 55.9 | 7.1 | 66.4 | 8.7 | 49.9 | 4.5 | 12.6 | 1.3 |
Моделирование динамики продукции икры показало снижение значений от I к IV стадии со смещением по оси абсцисс, что характеризует последовательную убыль икринок в процессе эмбриогенеза (рис. 1, табл. 4). Численность икринок I, II, III и IV стадий развития, продуцированных с момента появления в ихтиопланктоне и до 4 мая (средняя дата последней съемки), оценена в 397.3, 186.7, 70.9 и 15.2 × 1012 экз. соответственно. Разница между суммарной продукцией икринок на смежных стадиях с учетом их продолжительности равна величине общей смертности, которая в разные периоды эмбриогенеза продолжительностью 9–11 сут колебалась от 30 до 50% (табл. 5). Полученные величины характеризуют темп убыли икринок с середины I стадии развития до середины IV стадии. В наблюдавшихся температурных условиях этот период составлял 30 сут или 81% всего периода эмбриогенеза. Если допустить, что в начале I стадии и в конце IV стадии значения мгновенных коэффициентов убыли (Z) близки к расчетным (0.069 и 0.064 соответственно), то кривая посуточной убыли икры для всего периода эмбриогенеза, а также для отдельных стадий (рис. 2, табл. 6) свидетельствует о том, что общая смертность икринок от вымета до выклева составила 86.8%, а более половины погибшей икры приходилось на I стадию развития.
Рис. 1.
Эмпирическая (точки) и расчетная (линии) динамика суточной продукции икринок минтая на разных стадиях развития (I–IV) у западного побережья Камчатки в 2019 г.
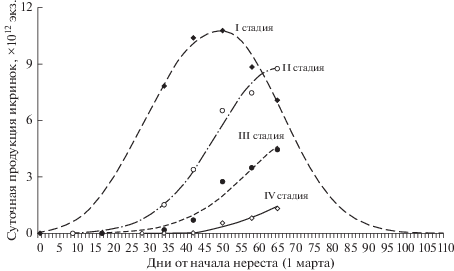
Таблица 4.
Коэффициенты уравнения кривой Гаусса для суточной продукции икринок минтая по стадиям развития (I−IV)
| Стадия развития | Коэффициенты функции Гаусса | SE | ||
|---|---|---|---|---|
| a | b | h | ||
| I | 50 | 10.79 | –0.0015 | 0.148 |
| II | 65 | 8.74 | –0.0018 | 0.178 |
| III | 75 | 5.43 | –0.0016 | 0.166 |
| IV | 82 | 2.50 | –0.0021 | 0.044 |
Таблица 5.
Смертность икринок минтая в отдельные периоды эмбриогенеза в водах западного побережья Камчатки в 2019 г.
| Показатель | Период эмбриогенеза | ||
|---|---|---|---|
| от середины I до середины II стадии |
от середины II до середины III стадии |
от середины III до середины IV стадии |
|
| Длительность периода, сут | 9 | 10 | 11 |
| Убыль икры, ×1012 экз. | 144.7 | 32.7 | 17.1 |
| Коэффициент общей смертности (А) | 0.437 | 0.321 | 0.484 |
| Мгновенный коэффициент (Z) | 0.069 | 0.038 | 0.064 |

ОБСУЖДЕНИЕ
В работе Балыкина (1993) приведена ссылка на неопубликованные данные О.Г. Золотова, согласно которым у западного побережья Камчатки выживаемость икринок стадии IV относительно стадии I составляла 18.6%. В статье отсутствует информация о том, к какому периоду стадий приведены расчеты (начальный, конечный или его середина), однако величина убыли близка к полученным нами результатам. По другим районам обитания минтая данные о величине смертности икринок за период эмбрионального развития также немногочисленны. Для минтая, нерестящегося на шельфе юго-восточного побережья Камчатки, величина убыли на IV стадии относительно стадии I оценена в 89.3% (Антонов, 1991), а для глубоководных каньонов этого же района она составляет 95.4% (Буслов и др., 2004). Аналогичный показатель для нерестилища в юго-западной части Берингова моря – 95.4–97.5% (Балыкин, 1993). Однако в последнем случае автор считает его завышенным из-за недооценки икринок поздних стадий. В прол. Шелихова западной части зал. Аляска оценки общей убыли икринок, полученные по расчетам через продукцию за нерестовый сезон, колебались в течение ряда лет от 62 до 98% (Picquelle, Megrey, 1993). По-видимому, на шельфовых нерестилищах в разных частях ареала уровень общей смертности икринок минтая относительно близок и варьирует от 70 до 90%.
Убыль икры в процессе эмбриогенеза изменяется неравномерно. Наибольшие коэффициенты убыли характерны для I и III стадий развития, а минимальные значения приходятся на II стадию (табл. 6). Очевидно, что значения коэффициентов в первую очередь связаны с продолжительностью стадий, которая в нашем исследовании в среднем составляла 9, 8, 11 и 9 сут (в соответствии с их последовательностью). Однако максимальную смертность икры на I стадии можно объяснить не ее продолжительностью, а критическим периодом в развитии, приходящимся на данный этап эмбриогенеза. Ранее отмечали, что основная убыль икринок наблюдается в период между морулой и началом гаструляции, т.е. приурочена к процессу дифференциации клеток бластодиска и приходится примерно на середину I стадии (Буслов, Сергеева, 2013). Японские исследователи в экспериментах по инкубированию икры минтая также выделяли морулу как критический период эмбриогенеза (Nakatani, Maeda, 1984). Причины этого неизвестны, однако данные о гибели в течение первых 4–6 сут развития около половины выметанной минтаем икры подтверждают и другие авторы (Brodeur et al., 1996).
В ходе дальнейшего развития смертность икринок снижается приблизительно на 50% и остается такой до середины III стадии, затем вновь увеличивается (рис. 2). Последнее обстоятельство может быть связано с процессом роста эмбриона и началом пигментации глаз и головы (Буслов, Сергеева, 2013). В данный период икринка становится заметнее в толще воды, вследствие этого увеличивается вероятность ее гибели от хищников, из которых основными являются гиперииды, эвфаузииды и амфиподы (Brodeur, Merati, 1993). Известно также о каннибализме минтая по отношению к развивающейся икре (Балыкин, 1997; Волков и др., 2003). Однако исследования показали, что масштабы выедания икринок минтая представителями нектона и медузами невелики и составляют не более 1–2% сезонной продукции икринок (Brodeur et al., 1996; Буслов и др., 2004; Горбатенко и др., 2012).
Оценки убыли, полученные в нашем исследовании, позволяют вернуться к вопросу о масштабах занижения нерестового запаса при расчетах, не учитывающих смертность развивающихся икринок. Согласно кривой, построенной по результатам пяти съемок, суммарная продукция икринок на I стадии развития за нерестовый сезон 2019 г. составила 467.2 × 1012 экз. (рис. 1). Поскольку при расчетах исходили из того, что на момент поимки все икринки в среднем достигли середины стадии, то их возраст может быть оценен в 4.0–4.7 сут (табл. 2). Следовательно, в расчетах не были учтены икринки I стадии развития, погибшие за это время. Исходя из посуточной кривой убыли (рис. 2), можно восстановить численность икринок через коэффициенты выживания учтенной икры, которые в зависимости от продолжительности стадии изменялись в пределах 0.725–0.751. Новые данные о величине суточной продукции аппроксимировали с помощью кривой Гаусса по рассчитанным ранее коэффициентам (рис. 3). В итоге суммарная продукция икринок на I стадии развития, оцененная с учетом смертности, составила 642.3 × 1012 экз., что на 37.5% больше первоначальной величины.
Рис. 3.
Расчетная динамика суточной продукции икринок минтая I стадии развития у западного побережья Камчатки в 2019 г. без учета (1) и с учетом (2) смертности.

Предположения о том, что общее количество выметанной икры, рассчитанное без учета ее убыли, занижается в 2–10 раз, значительно преувеличены (Лисовенко, 2000; Привалихин и др., 2001). Однако выявленное различие, на наш взгляд, весьма существенно, если оперировать терминами численности и биомассы нерестового запаса промыслового объекта. Продемонстрируем это на обобщенных расчетах. Зная среднюю плодовитость самок (153.9 тыс. икринок) и массу производителей в период икрометания (0.463 кг), а также приняв соотношение полов равным 1 : 1, можно рассчитать численность и биомассу нерестового запаса минтая в водах западного побережья Камчатки в 2019 г. Для двух вариантов оценок общего количества выметанной икры эти значения будут следующими: 6.1 млрд экз. или 2.8 млн т без учета убыли и 8.3 млрд экз. или 3.9 млн т, если смертность икринок учитывать.
Таким образом, при введении в расчеты поправки на смертность икры увеличение нерестового запаса минтая составило более 1 млн т. Представляется, что такой подход позволяет дать объективную оценку состояния его запасов по результатам ихтиопланктонных съемок.
Список литературы
Антонов Н.П. Биология и динамика численности восточнокамчатского минтая: Автореф. дис… канд. биол. наук. Владивосток: ТИНРО. 1991. 23 с.
Антонов Н.П., Золотов О.Г. Особенности размножения восточнокамчатского минтая // Популяционная структура, динамика численности и экология минтая. Владивосток: ТИНРО. 1987. С. 123–131.
Балыкин П.А. Изменчивость сроков нереста и смертность развивающейся икры у западноберинговоморского минтая (Theragra chalcogramma (Pallas)) // Исследования биологии и динамики численности промысловых рыб камчатского шельфа. Петропавловск-Камчатский: КоТИНРО. 1993. Вып. 2. С. 166–176.
Балыкин П.А. Некоторые особенности экологии размножения минтая Theragra chalcogramma // Вопр. ихтиологии. 1997. Т. 37. Вып. 2. С. 265–269.
Бизиков В.А., Поляков А.В. Географическая информационная система “КАРТМАСТЕР”: новые возможности и перспективы для рыбохозяйственных исследований // Математическое моделирование и информационные технологии в исследованиях биоресурсов Мирового океана: Тез. докл. Материалов отраслевого семинара. Владивосток: ТИНРО-центр. 2004. С. 89–91.
Буслов А.В., Сергеева Н.П. Эмбриональное развитие и ранний постэмбриогенез тресковых дальневосточных морей // Исследования водных биологических ресурсов Камчатки и северо-западной части Тихого океана: Сб. науч. тр. Камчат. НИИ рыб. хоз-ва и океаногр. 2013. Вып. 29. С. 5–69.
Буслов А.В., Тепнин О.Б. Условия нереста и эмбриогенеза минтая Theragra chalcogramma (Gadidae) в глубоководных каньонах тихоокеанского побережья Камчатки // Вопр. ихтиологии. 2002. Т. 42. Вып. 5. С. 617–625.
Буслов А.В., Тепнин О.Б., Дубинина А.Ю. Некоторые особенности экологии нереста и эмбриогенеза восточнокамчатского минтая // Изв. ТИНРО. 2004. Т. 138. С. 282–298.
Волков А.Ф., Горбатенко К.М., Мерзляков А.Ю. Планктон, состояние кормовой базы и питание массовых рыб эпи- и мезопелагиали Охотского моря в зимне-весенний период // Изв. ТИНРО. 2003. Т. 133. С. 169–235.
Горбатенко К.М., Овсянников Е.Е., Лаженцев А.Е., Шейбак А.Ю. Оценка выедания икры минтая нектоном и медузами в северной части Охотского моря в весенний период // Изв. ТИНРО. 2012. Т. 169. С. 12–20.
Зайцев Г.Н. Математическая статистика в экспериментальной ботанике. М.: Наука. 1984. 423 с.
Кузнецов В.В., Котенев Б.Н., Кузнецова Е.Н. Проблемы оценки численности и допустимого изъятия североохотоморского минтая Theragra chalcogramma // Вопр. рыболовства. 2008. Т. 9. № 2 (34). С. 276–293.
Лисовенко Л.А. Размножение рыб с прерывистым оогенезом и порционным нерестом на примере минтая западной Камчатки. М.: ВНИРО. 2000. 112 с.
Привалихин А.М., Варкентин А.И., Норвилло Г.В. Некоторые методологические подходы к оценке выживаемости минтая Theragra chalcogramma (Pallas) западной Камчатки в 1996–1998 годах // Вопр. рыболовства. 2001. Приложение 1. С. 230–236.
Расс Т.С., Казанова И.И. Методическое руководство по сбору икринок, личинок и мальков рыб. М.: Пищ. пром-ть. 1966. 42 с.
Рикер У.Е. Методы оценки и интерпретации биологических показателей популяций рыб. М.: Пищ. пром-сть. 1979. 408 с.
Фадеев Н.С. Нерестилища и сроки размножения минтая северной части Охотского моря // Популяционная структура, динамика численности и экология минтая. Владивосток: ТИНРО. 1987. С. 5–23.
Фадеев Н.С. Методика оценки запасов минтая по численности икры и размерно-возрастному составу // Биол. моря. 1999. Т. 25. № 3. С. 246–249.
Фадеев Н.С., Смирнов А.В. Оценка численности икры и производителей минтая в северной части Охотского моря // Биол. моря. 1987. № 4. С. 19–25.
Шунтов В.П., Волков А.Ф., Темных О.С., Дулепова Е.П. Минтай в экосистемах дальневосточных морей. Владивосток: ТИНРО. 1993. 426 с.
Brodeur R.D., Merati N. Predation on walleye pollock (Theragra chalcogramma) eggs in the western Gulf of Alaska: the roles of vertebrate and invertebrate predators // Mar. Biol. 1993. V. 117. P. 483–493.
Brodeur R.D., Picquelle S.J., Blood D.M., Merati N. Walleye pollock eggs distribution and mortality in the western Gulf of Alaska // Fish. Oceanogr. 1996. V. 5. № s1. P. 92–111.
Nakatani T., Maeda T. Thermal effect on the development of walleye pollock eggs and their upward speed to the surface // Bull. Jpn. Soc. Sci. Fish. 1984. V. 50. P. 937–942.
Picquelle S.J., Megrey B.A. A preliminary spawning biomass estimate of walleye pollock, Theragra chalcogramma, in the Shelikof Strait, Alaska, based on the annual egg production method // Bull. Mar. Sci. 1993. V. 53. P. 728–749.
Дополнительные материалы отсутствуют.