

Биология моря, 2022, T. 48, № 4, стр. 262-272
Организация целомической системы лофофора у брахиоподы Coptothyris grayi (Davidson, 1852) (Brachiopoda: Terebratulida)
А. В. Ратновская 1, *, Т. В. Кузьмина 1, **
1 Московский государственный университет им. М.В. Ломоносова, Биологический факультет
119234 Москва, Россия
* E-mail: belka190199@gmail.com
** E-mail: kuzmina-t@yandex.ru
Поступила в редакцию 25.11.2021
После доработки 12.02.2022
Принята к публикации 24.03.2022
- EDN: CFFWPI
- DOI: 10.31857/S0134347522040088
Аннотация
Плектолофный лофофор – самый сложно организованный тип лофофора современных брахиопод, характерный для отряда Terebratulida. На примере Coptothyris grayi (Davidson, 1852) впервые построена 3D-реконструкция целомической системы для плектолофного лофофора. Показано, что его целомическая система состоит из трех изолированных друг от друга компартментов. Первый компартмент выполняет транспортную и выделительную функции и представлен периэзофагеальным целомом, малыми и щупальцевыми целомическими каналами. Второй компартмент образован большими каналами лофофора, играющими роль гидроскелета. Третий компартмент представлен брахиальными карманами, которые сообщаются с перивисцеральным целомом и несут в себе выросты пищеварительной железы, позволяющие эффективно использовать пространство мантийной полости.
Брахиоподы, или плеченогие, – реликтовая группа морских беспозвоночных, которая достигла наибольшего разнообразия в палеозое (Harper et al., 2017). Тело брахиопод заключено в минерализованную раковину с дорсальной и вентральной створками. Передняя стенка тела образует дорсальную и вентральную мантийные складки, выстилающие мантийную полость и секретирующие раковину. В мантийной полости находится лофофор – щупальцевый орган, создающий токи воды для дыхания и фильтрации пищевых частиц.
Лофофор является синапоморфией надтипового таксона Lophophorata, в состав которого входят брахиоподы, форониды и мшанки (Hyman, 1959). У всех представителей Lophophorata лофофор формирует брахиальная ось, которая представляет собой ряд щупалец. Вдоль брахиальной оси тянется складка стенки тела (брахиальная складка). Между щупальцами и брахиальной складкой располагается пищевой желобок (Rudwick, 1970; Kuzmina et al., 2021). В центре брахиальной оси внутри пищевого желобка всегда расположен рот. За ртом находится одинарный ряд околоротовых (оральных) щупалец. У большинства брахиопод с двух сторон ото рта ряд щупалец становится двойным, ближе к брахиальной складке располагаются внутренние гребенчатые щупальца, а дальше от нее – наружные желобчатые щупальца (Rudwick, 1970; Кузьмина, Малахов, 2007). На концах брахиальной оси расположена зона закладки новых щупалец лофофора (Atkins, 1959a, 1959b, 1959c; Kuzmina et al., 2021).
Было высказано предположение, что эволюция лофофора брахиопод связана с удлинением брахиальной оси (Williams et al., 1997; Williams, Carlson, 2007; Kuzmina, Temereva, 2019). У брахиопод выделено несколько типов щупальцевого аппарата (Beecher, 1897; Emig, 1992; Kuzmina, Temereva, 2019; Kuzmina et al., 2021). Наиболее просто устроены таксолофный, трохолофный и шизолофный типы лофофора. Они являются последовательными стадиями онтогенеза лофофора у брахиопод с более сложно организованными типами щупальцевого аппарата, таких как спиролофный, зиголофный и плектолофный (Rudwick, 1970; Emig, 1992).
Брахиоподы представлены подтипами Linguliformea, Craniiformea и Rhynchonelliformea (Williams et al., 1996). Отряд Terebratulida (Rhynchonelliformea: Rhynchonellata) – один из наиболее успешных отрядов современных плеченогих. У большинства его представителей самый сложно организованный тип щупальцевого аппарата среди современных брахиопод – плектолофный лофофор, состоящий из трех рук, свободно расположенных в мантийной полости. Плектолофный лофофор поддерживается известковым скелетом (брахидиумом), который может быть представлен длинной или короткой петлей (Rudwick, 1970).
Первые сведения об организации целомической системы у брахиопод получены в XIX веке на основе анатомирования животных (Hancock, 1858). Изучение лофофорального целома брахиопод продолжилось в XX−XXI веках с использованием гистологических методов. В разной степени подробности был описан целом лофофора у лингулиформных брахиопод Lingula anatina из семейства Lingulidae (см.: Pross, 1980) и Pelagodiscus atlanticus из семейства Discinidae (см.: Kuzmina, Temereva, 2019); у ринхонеллиформных брахиопод Pumilus antiquatus (см.: Atkins, 1958), Terebratalia transversa (см.: Atkins, 1959а), Platidia anomioides (см.: Atkins, 1959b), Macandrevia cranium (см.: Atkins, 1959c), Megathiris detruncata (см.: Atkins, 1960), Calloria inconspicua (см.: Atkins, 1961a) и Megerlia truncata (см.: Atkins, 1961b) из отряда Terebratulida, а также у Notosaria nigricans (см.: Hoverd, 1985) и Hemithiris psittacea (см.: Кузьмина и др., 2006) из отряда Rhynchonellida. С помощью методов трансмиссионной электронной микроскопии исследована ультраструктура целомического эпителия лофофора у лингулиды L. anatina (см.: Storch, Welsch, 1976; Temereva, 2017), ринхонеллиды H. psittacea (см.: Kuzmina, Malakhov, 2011; Kuzmina et al., 2018) и теребратулиды T. transversa (см.: Reed, Cloney, 1977). Относительно недавно для изучения целомической системы брахиопод стали применять метод 3D-реконструкции по серии гистологических срезов. Построены 3D-реконструкции лофофорального целома L. anatina (см.: Temereva et al., 2015; Temereva, 2017) и целомической системы кранииформной брахиоподы Novocrania anomala (см.: Plandin, Temereva, 2021).
Однако не все аспекты организации целома у брахиопод исследованы достаточно подробно. До сих пор отсутствуют сведения о строении периэзофагеального целома и его взаимосвязи с другими отделами целомической системы у теребратулид. В настоящей работе впервые построена 3D-модель целомических полостей плектолофного лофофора брахиоподы Coptothyris grayi (Rhynchonelliformea: Terebratulida) и проведен сравнительный анализ организации целомической системы у лофофорат.
МАТЕРИАЛ И МЕТОДИКА
Три взрослые особи C. grayi были собраны в июле 2019 г. на глубине 8–10 м в зал. Восток Японского моря в районе морской биологической станции “Восток” Национального научного центра морской биологии им. А.В. Жирмунского ДВО РАН. Животных фиксировали 4% формалином на морской воде.
Анатомию лофофора изучали с помощью метода ручного анатомирования крупных экземпляров (длина вентральной створки 19 мм) под бинокуляром Leica M165C со встроенной цифровой камерой Leica DFC420. Для изучения строения лофофора также был использован метод компьютерной микротомографии. Для этого целая брахиопода (длина вентральной створки 8 мм) была зафиксирована в формалине, отмыта в воде, обезвожена в спиртах восходящей концентрации, переведена в ацетон и высушена с использованием метода критической точки в CO2 на аппарате сушки Hitachi critical point dryer HCP-1. Из высушенной раковины был извлечен лофофор и отсканирован на микротомографе. Сканирование было проведено в Лаборатории природных ресурсов Геологического факультета МГУ им. М.В. Ломоносова на микротомографе SkyScan 1172 (Bruker) при разрешении 9 мкм с шагом вращения 0.3 градуса, без фильтра, при напряжении 40 кВ и силе тока 250 мА. Полученные данные были обработаны в программах NRecon, DataViewer и CTVox.
Для построения трехмерной реконструкции по стандартной гистологической методике (Валовая, Кавтарадзе, 1993) при помощи ротационного микротома Leica RM 2125 изготовили серию гистологических срезов толщиной 5 мкм в поперечной плоскости одного экземпляра (длина вентральной створки 15 мм). Срезы окрашивали гематоксилином Караччи и заключали в канадский бальзам, затем фотографировали при помощи слайд-сканера Olympus VS-ASW с объективом ×10 (срезы фотографировали с интервалом 20 мкм). Полученные фотографии обрезали и уменьшали в программе IrfanView, а затем выравнивали в программе Amira ver. 5.2.2; 3D-реконструкции получены в программе Imaris ver. 7.2.1 (Bitplane, Zurich, Switzerland).
РЕЗУЛЬТАТЫ
1. Анатомия лофофора.
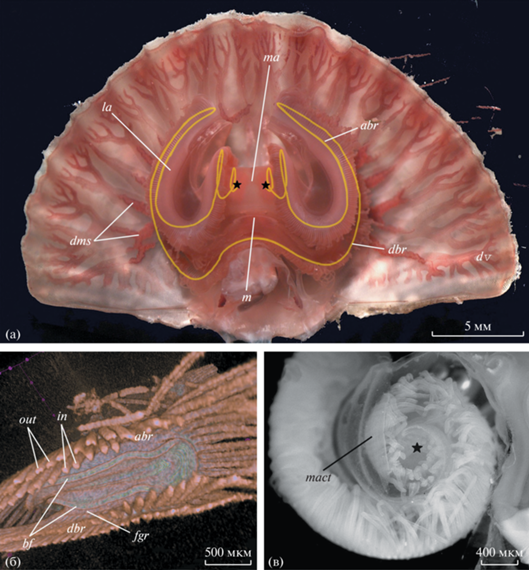
Мягкое тело C. grayi занимает заднюю часть объема между створками раковины брахиоподы, остальная часть пространства между створками представлена обширной мантийной полостью, в которой находится крупный лофофор (рис. 1а). Лофофор C. grayi образован сложно изогнутой брахиальной осью (рис. 1а). Почти на всем протяжении брахиальной оси щупальца образуют двойной ряд, в котором чередуются внутренние и наружные щупальца (рис. 1б). В центре брахиальной оси расположено щелевидное ротовое отверстие. С дорсальной стороны ото рта находится брахиальная складка, а с вентральной стороны расположен одинарный ряд околоротовых щупалец. В области рта брахиальная ось отрывается от передней стенки тела и формирует три свободно расположенные руки лофофора – две латеральные и одну медиальную. Слева и справа ото рта брахиальная ось направлена в сторону переднего края раковины, образуя нисходящую ветвь соответственно левой и правой латеральной руки (рис. 1а). На конце каждой латеральной руки брахиальная ось изгибается вверх и возвращается ко рту, образуя восходящую ветвь. Дистальные концы восходящих ветвей латеральных рук, расположенные симметрично слева и справа ото рта, объединяются соединительно-тканной диафрагмой и закручиваются в виде спирали в сагиттальной плоскости в сторону дорсальной створки, формируя медиальную руку (рис. 1а). Таким образом, каждая рука лофофора состоит из двух брахиальных осей. На концах брахиальной оси в дистальной части медиальной руки лофофора находится зона роста лофофора, в которой происходит закладка новых щупалец (рис. 1а, 1в).
Рис. 1.
Морфология лофофора Coptothyris grayi: а – вскрытая брахиопода с удаленной вентральной створкой (световая микроскопия), желтым цветом обозначена брахиальная ось; б – дистальный участок левой латеральной руки (микротомография); в – дистальная часть медиальной руки (световая микроскопия). Обозначения: abr – восходящая ветвь, bf – брахиальная складка, dbr – нисходящая ветвь, dms – дорсальные мантийные каналы, dv – дорсальная створка раковины, fgr – пищевой желобок, in – внутренние щупальца, la – латеральная рука, m – рот, ma – медиальная рука, mact – соединительно-тканная перегородка медиальной руки, out – наружные щупальца. Звездочками указана зона роста лофофора (место формирования новых щупалец).
2. Организация целомической системы лофофора.
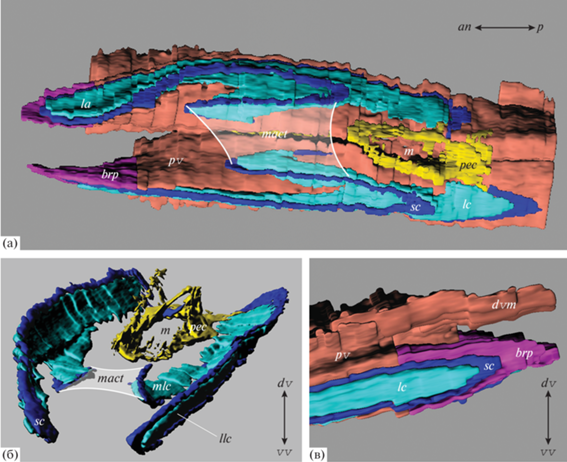
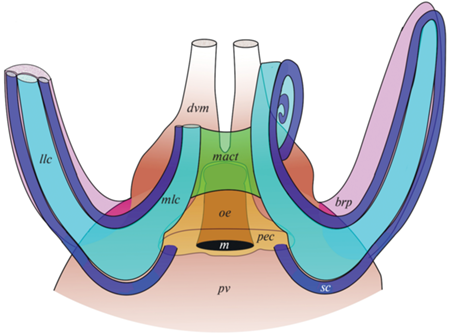
Целомическая система лофофора C. grayi образована периэзофагеальным целомом, малыми и большими целомическими каналами и брахиальными карманами (рис. 2а).
Рис. 2.
3D-реконструкция целомической системы лофофора Coptothyris grayi: а – общая 3D-реконструкция, вид с вентральной стороны; б – 3D-реконструкция больших и малых каналов лофофора; в – 3D-реконструкция целомической системы латеральной руки лофофора и связанного с ней перивисцерального целома. Обозначения: brp – брахиальный карман, dvm – дорсальный медиальный мантийный канал, la – латеральная рука, lc – большой канал, llc – большой канал латеральной руки, m – рот, mact – соединительно-тканная диафрагма медиальной руки, mlc – большой канал медиальной руки, pec – периэзофагеальный целом, pv – перивисцеральный целом, sc – малый канал. Двойные стрелки указывают направление к переднему (an) и заднему (p) концам тела, а также к дорсальной (dv) и вентральной (vv) створкам.
2.1. Периэзофагеальный целом.
Периэзофагеальный целом окружает рот и пищевод, наиболее обширен в околоротовой области. Затем он плавно сужается и в области перехода пищевода в желудок представлен узкими лакунами в соединительной ткани под передней стенкой тела (рис. 3а, 3г). Периэзофагеальный целом пересечен тонкими перегородками и дает многочисленные узкие ответвления (дивертикулы) в соединительную ткань, окружающую пищевод (рис. 3б). Данные 3D-реконструкции четко показали, что периэзофагеальный целом обособлен от туловищного целома (рис. 2а). За ртом от периэзофагеального целома берут начало целомические каналы околоротовых щупалец, которые слепо замыкаются на концах (рис. 3в, 3г). Кроме того, согласно данным 3D-реконструкции, с двух сторон ото рта от периэзофагеального целома берут начало малые каналы лофофора (рис. 3г, 4а–4г, 5).
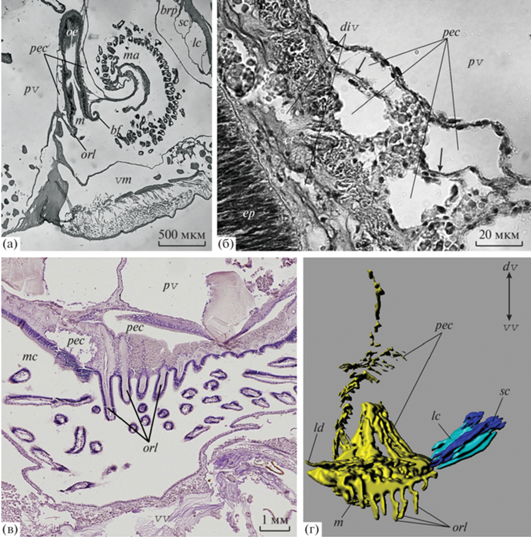
Рис. 3.
Периэзофагеальный целом Coptothyris grayi: a – сагиттальный срез через околоротовую область (световая микроскопия); б – срез через участок периэзофагеального целома (световая микроскопия); в – продольный срез через одинарный ряд щупалец за ртом (световая микроскопия); г – 3D-реконструкция периэзофагеального целома, вид сзади. Обозначения: bf – брахиальная складка, brp – брахиальный карман, ct – соединительная ткань, div – дивертикулы периэзофагеального целома, ep – эпителий пищевода, ld – боковое ответвление периэзофагеального целома, lc – большой канал, m – рот, ma – медиальная рука, mc – мантийная полость, oe – пищевод, orl – оральные щупальца, pec – периэзофагеальный целом, pv – перивисцеральный целом, sc – малый канал, vm – вентральная складка мантии. Стрелки указывают на перегородки периэзофагеального целома; двойные стрелки указывают направление к дорсальной (dv) и вентральной (vv) створкам.
2.2. Малые и щупальцевые целомические каналы.
Периэзофагеальный целом, с каждой стороны ото рта дает узкое ответвление, переходящее затем в левый и правый малые целомические каналы (рис. 3г, 4а). Малые целомические каналы на всем протяжении сопровождают брахиальную ось лофофора (рис. 2а, 2б, 4а, 5), формируя слепо замкнутые ответвления в каждое щупальце. Слева и справа ото рта малые каналы сначала заходят соответственно в левую и правую латеральные руки лофофора, тянутся вдоль нисходящей ветви брахиальной оси в сторону переднего края раковины, а затем делают поворот на конце руки и идут вдоль восходящей ветви брахиальной оси назад ко рту. Таким образом, каждая боковая рука содержит два малых целомических канала (рис. 2б, 4д, 5). Далее левый и правый малые каналы изгибаются вперед и сопровождают две брахиальные оси в медиальной руке лофофора, закручиваясь в спираль. На конце медиальной руки малые каналы слепо замыкаются. Таким образом, медиальная рука также содержит два малых целомических канала (рис. 4е, 5).
Рис. 4.
Малые и большие целомические каналы лофофора Coptothyris grayi: а – 3D-реконструкция участка соединения малых каналов с периэзофагеальным целомом, вид с вентральной стороны (щупальца не реконструированы); б–г – серия поперечных гистологических срезов с интервалом 25 мкм через левую часть лофофора в области впадения малого канала в периэзофагеальный целом; д – поперечный срез через левую латеральную руку лофофора; е – поперечный срез через медиальную руку лофофора. Обозначения: bf – брахиальная складка, brp – брахиальный карман, dm – дорсальная мантия, fgr – пищевой желобок, lc – большой канал, m – рот, mact – соединительно-тканная диафрагма медиальной руки, mc – мантийная полость, sc – малый канал, pec – периэзофагеальный целом, pv – перивисцеральный целом; i – плоскость среза, рис. 4б; ii – плоскость среза, рис. 4в; iii – плоскость среза, рис. 4г. Белые стрелки указывают на соединения периэзофагеального целома с малыми каналами, черные стрелки – на срезы выростов пищеварительной железы в брахиальном кармане. Двойные стрелки указывают направление к дорсальной (dv) и вентральной (vv) створкам.
2.3. Большие целомические каналы.
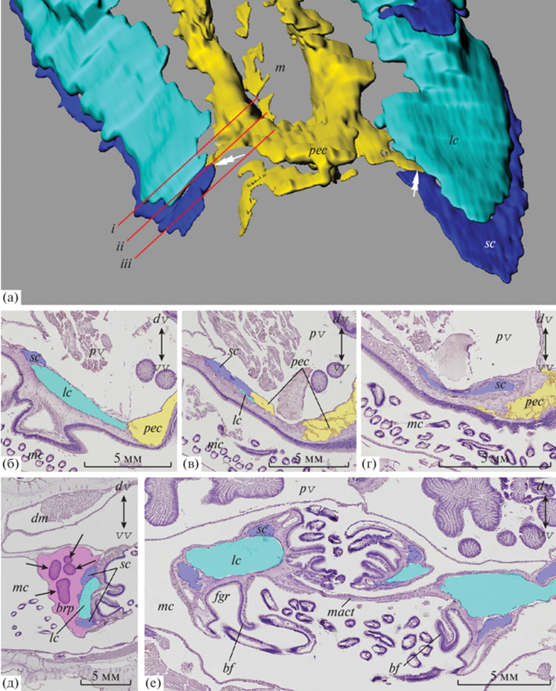
В околоротовой области слева и справа от периэзофагеального целома располагаются большие целомические каналы. В отличие от малых каналов они слепо замкнуты на проксимальном конце и не связаны с периэзофагеальным целомом, хотя очень близко к нему подходят (рис. 4а–в). В околоротовой области каждый большой канал разделяется на боковую и медиальную ветви. Боковая ветвь продолжается вперед в латеральную руку и слепо замыкается на ее конце; медиальная ветвь с каждой стороны заходит в медиальную руку лофофора, закручиваясь в спираль (рис. 2а, 2б). Таким образом, в каждой латеральной руке тянется по одному большому целомическому каналу (рис. 2б, 2в, 4д, 5), а в медиальной руке имеется два больших целомических канала (рис. 2а, 2б, 4е, 5).
2.4. Брахиальные карманы.
Рядом с большими целомическими каналами с внутренней стороны каждой латеральной руки лофофора тянется еще одна целомическая полость – брахиальный карман (рис. 2а, 2в, 4д). Данные 3D-реконструкции показали, что брахиальные карманы – это выросты дорсальной части перивисцерального целома (рис. 2в, 5). Перивисцеральный целом занимает большую часть тела брахиоподы и дает ответвления в мантийные складки, формируя разветвленную сеть мантийных каналов (рис. 1а).
С левой и правой стороны от сагиттальной плоскости дорсальная часть перивисцерального целома делится на две ветви: одна ветвь образует медиальный целомический канал мантии, который тянется к переднему краю мантии, а другая ветвь становится частью лофофора, формируя брахиальные карманы (рис. 2в, 5). Брахиальные карманы представляют собой широкие полости, которые постепенно сужаются и слепо замыкаются на концах латеральных рук. В брахиальные карманы заходят выросты пищеварительной железы (рис. 4д).
ОБСУЖДЕНИЕ
В настоящей работе показано, что у брахиоподы Coptothyris grayi брахиальная ось содержит большой и малый целомические каналы. Это характерно для всех типов лофофора, кроме таксолофного, который встречается лишь как онтогенетическая стадия, и трохолофного. Трохолофный лофофор у взрослых особей описан только у видов Gwynia capsula и Goniobrochus sp. из отряда Terebratulida (см.: Emig, 1992) и содержит всего один малый целомический канал, дающий ответвление в каждое щупальце. Малый канал всех типов лофофора несет лофофоральный кровеносный сосуд (Atkins, 1959b; Reed, Cloney, 1977; Kuzmina et al., 2018) и, вероятно, выполняет транспортную функцию, связанную с дыхательной функцией лофофора. При удлинении брахиальной оси возникает необходимость поддержки лофофора, поэтому, начиная с шизолофного лофофора, появляется большой целомический канал, который выполняет функцию гидроскелета.
Согласно нашим данным, большие каналы плектолофного лофофора C. grayi изолированы друг от друга в околоротовой области. Сходное строение больших каналов лофофора описано у теребратулиды P. antiquatus c шизолофным лофофором (Atkins, 1958), у лингулиды L. anatina (см.: Pross, 1980), кранииды N. anomala (см.: Plandin, Temereva, 2021) и ринхонеллиды H. psittacea (см.: Кузьмина и др., 2006) со спиролофными лофофорами, у дисциниды P. atlanticus с видоизмененным зиголофным лофофором (Kuzmina, Temereva, 2019), а также у теребратулид T. transversa, P. anomioides, C. inconspicua и M. truncata, обладающих плектолофным лофофором (Atkins, 1959a, 1959b, 1961a, 1961b). Такое строение позволяет большим каналам эффективнее контролировать давление целомической жидкости на стенки канала, тем самым увеличивая или уменьшая тургор лофофора. Гидростатическую функцию больших каналов лофофора подтверждает наличие мышц, образованных целомическим эпителием больших каналов. Они описаны для спиролофного лофофора лингулид (Temereva, 2017), краниид (Plandin, Temereva, 2021) и ринхонеллид (Kuzmina et al., 2018). У спиролофного лофофора N. anomala (см.: Plandin, Temereva, 2021) и H. psittacea (см.: Кузьмина и др., 2006) большие каналы имеют крупные дорсальные выросты в околоротовой области, которые, предположительно, могут хранить дополнительный объем жидкости для более эффективного функционирования гидроскелета больших целомических каналов (Plandin, Temereva, 2021). У C. grayi мы не обнаружили подобных выростов. Возможно, это связано с развитием у теребратулид скелетных поддерживающих структур, снижающих значимость гидроскелета для поддержки плектолофного лофофора.
В лофофоре C. grayi каждая рука образована двумя брахиальными осями. При этом в латеральных руках происходит слияние больших каналов двух брахиальных осей; в медиальной руке слияния целомических каналов не происходит, две брахиальные оси связаны лишь соединительно-тканной перегородкой. Такая организация брахиальных осей является апоморфным признаком для зиголофного и плектолофного лофофоров теребратулид (Atkins, 1959a, 1959b, 1959c; 1961a, 1961b). В модифицированном зиголофном лофофоре дисциниды P. atlanticus каждая латеральная рука также состоит из двух брахиальных осей, однако большие каналы не объединяются, а располагаются в общем соединительно-тканном матриксе (Kuzmina, Temereva, 2019). У теребратулид брахиальная ось значительно длиннее, чем у дисцинид, поэтому можно предположить, что слияние больших каналов двух брахиальных осей в латеральных руках приводит к дополнительному укреплению рук лофофора.
Пищевод C. grayi окружен периэзофагеальным целомом, который пересечен перегородками и образует многочисленные выросты в соединительную ткань вокруг пищевода. Сходная организация описана у периэзофагеального целома лингулиформных (Pross, 1980; Temereva, 2017), кранииформных (Blochmann, 1892; Robinson, 2014; Plandin, Temereva, 2021), а также ринхонеллиформных брахиопод из отрядов Rhynchonellida (Kuzmina, Malakhov, 2011) и Terebratulida (Atkins, 1958, 1959a, c). Исследования ультраструктуры периэзофагеального целома ринхонеллиформных (Kuzmina, Malakhov, 2011) и кранииформных (Plandin, Temereva, 2021) брахиопод выявили хорошо развитые мышечные клетки в составе эпителиальной выстилки выростов периэзофагеального целома в соединительную ткань, окружающую пищевод, что, вероятно, служит для усиления мышечной обкладки пищевода. Периэзофагеальный целом C. grayi также образует похожие выросты вокруг пищевода, однако для подтверждения аналогичной функции этих выростов требуются дальнейшие ультраструктурные исследования.
На примере C. grayi впервые для теребратулид показано, что периэзофагеальный целом плектолофного лофофора дает слепо замкнутые ответвления в каждое околоротовое щупальце. Сходные данные по организации периэзофагеального целома получены для спиролофного лофофора ринхонеллиды H. psittacea (см.: Кузьмина и др., 2006) и кранииды N. anomala (см.: Plandin, Temereva, 2021). Взаимосвязь целомов околоротовых щупалец и периэзофагеального целома, вероятно, является характерной чертой всех брахиопод.
Периэзофагеальный целом C. grayi сообщается с малыми целомическими каналами лофофора, что ранее было описано для спиролофного лофофора H. psittacea (см.: Kuzmina, Malakhov, 2011), N. anomala (см.: Plandin, Temereva, 2021) и L. anatina (см.: Pross, 1980), шизолофного лофофора P. antiquatus (см.: Atkins, 1958) и плектолофного лофофора M. cranium (см.: Atkins, 1959с). Известно, что в перегородках периэзофагеального целома H. psittacea проходит сеть кровеносных сосудов, стенки которых образованы подоцитоподобными клетками. Высказано предположение, что в периэзофагеальном целоме брахиопод происходит ультрафильтрация жидкости (Kuzmina, Malakhov, 2011). Взаимосвязь малых каналов лофофора с периэзофагельным целомом у всех изученных групп брахиопод, вероятно, обусловлена выделительной функцией малых каналов. Предполагается, что водорастворимые продукты обмена (в частности, аммоний) выводятся в окружающую среду диффузией сквозь стенку тела из кровеносных сосудов в щупальцах лофофора (Peck et al., 1997; Williams et al., 1997; Kuzmina, Malakhov, 2011).
В настоящей работе впервые для теребратулид показано, что периэзофагеальный целом C. grayi обособлен от перивисцерального целома. Это согласуется с данными, полученными ранее для лингулиформных (Temereva, 2017) и кранииформных (Plandin, Temereva, 2021) брахиопод, а также для ринхонеллиформных брахиопод из отряда Rhynchonellida (Kuzmina, Malakhov, 2011). Полученные результаты уточняют сведения об организации периэзофагеального целома у тереб-ратулид, для которых ранее не была показана изолированность периэзофагеальной полости от перивисцерального целома (Atkins, 1958; 1959a, 1959с).
У изученного вида C. grayi описаны брахиальные карманы, которые являются выростами перивисцерального целома и сопровождают латеральные руки лофофора. Наличие брахиальных карманов является апоморфным признаком плектолофного лофофора теребратулид (Atkins, 1959a, 1959b; 1961a, 1961b; Кузьмина, Малахов, 2007). Нами показано, что брахиальные карманы C. grayi содержат часть пищеварительной железы. Ранее отростки пищеварительной железы в брахиальных карманах были изображены на схемах поперечного среза через плектолофный лофофор у теребратулид C. inconspicua и M. truncata (см.: Atkins, 1961a, 1961b), хотя в тексте статей об этом не упоминается. Можно предположить, что расположение пищеварительной железы внутри латеральных рук лофофора позволяет более рационально использовать пространство мантийной полости.
Исходя из полученных результатов, у взрослых теребратулид можно выделить три изолированных компартмента целомической системы: целом больших каналов лофофора; периэзофагеальный целом, связанный с малыми и щупальцевыми каналами лофофора; обширный перивисцеральный целом, который дает выросты в брахиальные карманы, сопровождающие латеральные руки лофофора, и в мантию (рис. 5).
Рис. 5.
Схема строения целомической системы лофофора Coptothyris grayi. Обозначения: brp – брахиальный карман, dvm – дорсальный медиальный мантийный канал, llc – большой канал латеральной руки, m – рот, mact – соединительно-тканная диафрагма медиальной руки, mlc – большой канал медиальной руки, oe – пищевод, pec – периэзофагеальный целом, pv – перивисцеральный целом, sc – малый канал.
Ранее было показано, что в онтогенезе теребратулид целом лофофора формируется из единого целомического зачатка (Percival, 1960; Williams et al., 1997; Кузьмина и др., 2019), поэтому можно предположить, что большие, малые и щупальцевые каналы, а также периэзофагеальный целом являются одним отделом целомической системы и представляют собой лофофоральный целом. Таким образом, у теребратулид можно выделить два отдела целомической системы: лофофоральный и перивисцеральный. Сходным образом организована целомическая система у представителей другого отряда ринхонеллиформных брахиопод – Rhynchonellida (Кузьмина и др., 2006; Kuzmina, Malakhov, 2011).
У представителя кранииформных брахиопод N. anomala описана пентамерия целома и выделены следующие компартменты целомической системы: большие каналы; малые каналы, связанные с периэзофагеальным целомом; фронтальные камеры; перивисцеральный целом и камеры задних аддукторов (Plandin, Temereva, 2021). Поскольку у N. anomala целом лофофора формируется из одного целомического зачатка (Nielsen, 1991), малые целомические каналы, периэзофагеальный целом и большие целомические каналы лофофора можно отнести к лофофоральному целому. Фронтальные камеры и камеры задних аддукторов встречаются исключительно у кранииформных брахиопод, поэтому можно предположить, что они являются производными перивисцерального целома, внутри которых формируется мускулатура: внутри фронтальных камер располагаются фронтальные мышцы лофофора, а внутри задних камер формируются задние мышцы-замыкатели (Plandin, Temereva, 2021). Важно отметить, что мышцы лофофора и задние мышцы-замыкатели у представителей других подтипов брахиопод располагаются внутри перивисцерального целома (Williams et al., 1997).
У представителя лингулиформных брахиопод L. anatina помимо лофофорального и перивисцерального целомов описан преоральный целом, который располагается надо ртом в брахиальной складке (Temereva et al., 2015; Temereva, 2017). Таким образом, у брахиопод можно выделить тримерию целома (см. также: Темерева, Малахов, 2011). Преоральный целом, характерный только для лингулиформных брахиопод, представляет собой протоцель. Лофофоральный целом является мезоцелем, а перивисцеральный целом – метацелем.
Тримерия целомов рассматривается и для других представителей лофофорат – мшанок и форонид (Темерева, Малахов, 2006; Темерева, Малахов, 2011; Hyman, 1959; Mukai et al., 1997; Temereva, Mala-khov, 2011; Temereva, 2015), мезоцель которых также представлен лофофоральным целомом. Однако мезоцель у брахиопод имеет наиболее сложную морфологию и в большинстве случаев состоит из двух изолированных компартментов: большого канала, выполняющего функцию гидроскелета, а также связанного с щупальцевыми каналами и периэзофагеальным целомом малого канала, выполняющего циркуляторную функцию. Такое разделение функций целомической системы лофофора брахиопод позволило представителям этой группы максимально удлинить брахиальную ось и достигнуть наибольшего разнообразия типов лофофора по сравнению с другими лофофоратами.
Список литературы
Валовая М.А., Кавтарадзе Д.Н. Микротехника: правила, приемы, искусство, эксперимент. М.: Изд-во МГУ. 1993. 240 с.
Кузьмина Т.В., Малахов В.В., Темерева Е.Н. Анатомия целомической системы замковой брахиоподы Hemithyris psittacea (Brachiopoda, Articulata) // Зоол. журн. 2006. Т. 85. Вып. 9. С. 1118–1128.
Кузьмина Т.В., Малахов В.В. Организация лофофора брахиопод // Палеонт. журн. 2007. Т. 5. С. 46–63.
Кузьмина Т.В., Малахов В.В., Темерева Е.Н. Личиночное развитие брахиопод Coptothyris grayi (Davidson, 1852) (Terebratulida: Rhynchonelliformea) и эволюция жизненных циклов брахиопод // Зоол. беспозв. 2019. Т. 16. № 1. С. 27−40.
Темерева Е.Н., Малахов В.В. Ответ Томасу Бартоломеусу: “Личинка форониды Phoronopis harmeri Pixell, 1912 имеет тримерную организацию целома” // Зоол. беспозв. 2006. Т. 3. № 1. С. 1−21.
Темерева Е.Н., Малахов В.В. Свидетельство метамерии у взрослых брахиопод и форонид // Зоол. беспозв. 2011. Т. 8. № 2. С. 87−101. https://doi.org/10.15298/invertzool.08.2.01
Atkins D. A new species and genus of Kraussinidae (Brachiopoda) with a note on feeding // Proc. Zool. Soc. London. 1958. V. 131. P. 559–582.
Atkins D. The growth stages of the lophophore and loop of the brachiopod Terebratalia transversa (Sowerby) // J. Morphol. 1959a. V. 105. № 3. P. 401–426.
Atkins D. The growth stages of the lophophore of the brachiopods Platidia davidsoni (Eudes deslongchamps) and P. anomioides (Philippi), with notes on the feeding mechanism // J. Mar. biol. Assoc. U.K. 1959b. V. 38. P. 103–132.
Atkins D. The early growth stages and adult structure of the lophophore of Macandrevia cranium (Müller) (Brachiopoda, Dallinidae) // J. Mar. Biol. Assoc. U.K. 1959c. V. 38. P. 335–350.
Atkins D. The ciliary feeding mechanism of the Megathyridae (Brachiopoda), and the growth stages of the lophophore // J. Mar. Biol. Assoc. U.K. 1960. V. 39. P. 459–479.
Atkins D. A note on the growth stages and structure of the adult lophophore of the brachiopod Terebratella (Waltonia) inconspicua (G. B. Sowerby) // Proc. Zool. Soc. London. 1961a. V. 136. P. 255–271.
Atkins D. The growth stages and adult structure of the lopho-phore of the brachiopods Megerlia truncata (L.) and M. echinata (Fischer & Oehlert) // J. Mar. Biol. Assoc. U.K. 1961b. V. 41. P. 95–111.
Beecher C.E. Morphology of the Brachia // Bull. U.S. Geol. Surv. 1897. V. 87. P. 105–112.
Blochmann F. Untersuchungen über den Bau der Brachiopoden // Die Anatomie von Crania anomala (Müller). Jena: Gustav Fischer. 1892. Pt. 1. P. 1–65.
Emig C.C. Functional disposition of the lophophore in li-ving Brachiopoda // Lethaia. 1992. V. 25. P. 291–302.
Hancock A. On the organization of the Brachiopoda // Philos. Trans. R. Soc. London. 1858. V. 148. P. 791–869.
Harper D.A.T., Popov L.E., Holmer L.E. Brachiopods: origin and early history // Palaeontology. 2017. V. 60. № 5. P. 1–23.
Hoverd W.A. Histological and ultrastructural observations of the lophophore and larvae of the brachiopod, Notosaria nigricans (Sowerby 1846) // J. Nat. Hist. 1985. V. 19. № 5. P. 831–850.
Hyman L.H. The Invertebrates: Smaller coelomate groups. New York: McGraw Hill. 1959. P. 1–783.
Kuzmina T.V., Malakhov V.V. The periesophageal celom of the articulate brachiopod Hemithyris psittacea (Rhynchonelliformea, Brachiopoda) // J. Morphol. 2011. V. 272. P. 180–190.
Kuzmina T.V., Ratnovskaya A.A., Madison A.A. Lophophore evolution from the Cambrian to the present // Paleontol. J. 2021. V. 55. № 10. P. 1109–1140.
Kuzmina T.V., Temereva E.N., Malakhov V.V. Ultrastructure of the lophophoral coelomic lining in the brachiopod Hemithiris psittacea: functional and evolutionary significance // Zoomorphology. 2018. V. 137. № 2. P. 257–272.
Kuzmina T.V., Temereva E.N. Organization of the lophophore in the deep-sea brachiopod Pelagodiscus atlanticus and evolution of the lophophore in the Brachiozoa // Org. Diversity Evol. 2019. V.19. P. 31–39.
Mukai H., Terakado K., Reed C.G. Bryozoa // Microscopic anatomy of invertebrates. V. 13: Lophophorates, Entoprocta and Cycliophora. New York: Wiley-Liss. 1997. P. 45–206.
Nielsen C. The development of the brachiopod Crania (Neocrania) anomala (O. F. Müller) and its phylogenetic significance // Acta Zool. 1991. V. 72. № 1. P. 7–28.
Peck L.S., Rhodes M.L., Curry G.B. et al. Physiology // Treatise on invertebrate paleontology, Part H: Brachiopoda (revised). V. 1: Introduction. Lawrence, Kans.: Univ. of Kansas Press. 1997. P. 213–240.
Percival E. A contribution to the life-history of the brachiopod Tegulorhynchia nigricans // J. Cell Sci. 1960. V. s3-101. P. 439–457.
Plandin F.A., Temereva E.N. Anatomy of the coelomic system in Novocrania anomala (Brachiopoda, Craniiformea) and relationships within brachiopods // Zoolo-gy. 2021. V. 144. Art. ID 125884. https://doi.org/10.1016/j.zool.2020.125884
Pross A. Untersuchungen zur Gliederung von Lingula anatina (Brachiopoda). Archimerie bei Brachiopoden // Zool. Jahrb. Abt. Anat. Ontog. Tiere. 1980. V. 103. P. 250–263.
Reed C.G., Cloney R.A. Brachiopod tentacles: Ultrastructure and functional significance of the connective tissue and myoepithelial cells in Terebratalia // Cell Tissue Res. 1977. V. 185. P. 17–42.
Robinson J. The muscles, body wall and valve-opening mechanism of extant craniid (inarticulated) brachiopods // J. Nat. Hist. 2014. V. 48. P. 1231–1252.
Rudwick M.J.S. Living and fossil brachiopods. London: Hutchinson and Co. Ltd. 1970. 199 p.
Storch V., Welsch U. Elektronenmikroskopische und enzymhistochemische untersuchungen über lophophor und tentakel von Lingula unguis L. (Brachiopoda) // Zool. Jahrb., Abt. Anat. Ontog. Tiere. 1976. V. 96. P. 225–237.
Temereva E.N. Organization of the coelomic system in Phoronis australis (Lophotrochozoa: Phoronida) and consideration of the coelom in the lophophorates // J. Zool. 2015. V. 296. № 2. P. 79–94.
Temereva E.N. Ultrastructure of the coelom in the brachiopod Lingula anatina // J. Morphol. 2017. V. 278. P. 997–1011.
Temereva E.N., Gebruk A.A., Malakhov V.V. Demonstration of the preoral coelom in the brachiopod Lingula anatina with consideration of its phylogenetic significance // Zool. Anz. 2015. V. 256. P. 22–27.
Temereva E.N., Malakhov V.V. Organization of the epistome in Phoronopsis harmeri (Phoronida) and consideration of the coelomic organization in Phoronida // Zoomorphology. 2011. V. 130. Art. ID 121. https://doi.org/10.1007/s00435-011-0126-z
Williams A., Carlson S.J. Affinities of brachiopods and trends in their evolution // Treatise on Invertebrate Paleontology, Part H: Brachiopoda (revised). V. 6: Supplement. Lawrence, Kans.: Univ. of Kansas Press. 2007. P. 2822–2877.
Williams A., Carlson S.J., Brunton C.H.C. et al. A supra-ordinal classification of the Brachiopoda // Philos. Trans. R. Soc., B. 1996. V. 351. P. 1171–1193.
Williams A., James M.A., Emig C.C. et al. Anatomy // Treatise on Invertebrate Paleontology, Part H: Brachiopoda (revised). V. 1: Introduction. Lawrence, Kans.: Univ. of Kansas Press. 1997. P. 7–188.
Дополнительные материалы отсутствуют.