

Биология моря, 2022, T. 48, № 4, стр. 242-250
Крупная фракция светящегося планктона в пиках биолюминесценции в Черном море
А. В. Темных 1, *, М. И. Силаков 1, А. В. Мельник 1
1 Институт биологии южных морей имени А.О. Ковалевского РАН
299011 Севастополь, Россия
* E-mail: atemnykh@yandex.ru
Поступила в редакцию 27.07.2021
После доработки 01.03.2022
Принята к публикации 24.03.2022
- EDN: DJNJDW
- DOI: 10.31857/S0134347522040118
Аннотация
В Черном море в вертикальном профиле биолюминесценции летом наблюдали, как правило, один пик, который в ночное время располагался на глубине 5–30 м, а в дневное – на глубине 50–82 м. На двух из 14 станций в утренние и вечерние часы отмечены два пика, соответствующие этим же горизонтам. В верхнем пике зарегистрирована сильная положительная связь суммарной интенсивности биолюминесценции с обилием трех доминирующих видов: коэффициент корреляции R составил по +0.79 для видов Noctiluca scintillans и Oikopleura dioica, +0.70 для вида Oithona similis и +0.82 для общей численности светящегося планктона. В верхнем пике отмечена значимая отрицательная зависимость численности указанных видов от температуры; у N. scintillans выявлена значимая отрицательная связь с изменением солености (R = −0.72). Практически на всех станциях по численности доминировала динофлагеллята N. scintillans. В нижних пиках значимой корреляции между биолюминесценцией и численностью светящихся видов не обнаружено. Возможно, вклад в поле биолюминесценции вносят и другие зоопланктонные организмы, способность которых к свечению еще не исследована.
Биолюминесценция – проявление жизнедеятельности организмов в виде электромагнитного излучения в видимой области спектра. Способность к свечению обнаружена у таких представителей Мирового океана, как бактерии, динофлагелляты, простейшие, кишечнополостные, черви, моллюски, ракообразные, иглокожие и рыбы (Гительзон и др., 1992; Токарев, 2006; Haddock et al., 2010; Martini, Haddock, 2017).
Суммарный световой эффект, создаваемый совокупностью морских биолюминесцентов в толще воды, называется полем биолюминесценции (ПБ) (Токарев и др., 2000). Пространственной структуре этого поля присуща вертикальная стратификация, определяемая параметрами пелагического сообщества (видовым составом, трофической структурой и т.д.). Данные параметры, в свою очередь, зависят от гидрологического режима и гидродинамических характеристик водных масс (Токарев, 2006).
Исследованию мелкой фракции планктонных биолюминесцентов, в первую очередь одноклеточных водорослей, посвящены многие работы (Евстигнеев и др., 1993; Черепанов и др., 2007; Серикова и др., 2010, 2017; Брянцева и др., 2014; Полонский и др., 2018, и др.), в которых описаны виды одноклеточных водорослей, вносящие вклад в ПБ, и показана связь их распределения с пространственной структурой ПБ Черного моря. Однако в Черном море способность к свечению выявлена и у более крупных организмов, к ним относятся представители зоопланктона и динофлагеллята Noctiluca scintillans (Macartney) Kofoid & Swezy, 1921 (Dinophyceae: Noctilucales). Судя по энергетическим параметрам биолюминесцентного сигнала отдельных организмов (Токарев, 2006), этот вклад должен быть достаточно большим.
В списках светящихся крупных планктонных видов Черного моря имеются расхождения, так как одни таксоны были включены в список биолюминесцентов на основании собственных экспериментальных данных (Евстигнеев, Битюков, 1990; Евстигнеев и др., 1993), а другие – в результате анализа литературного материала по всему Мировому океану (Токарев, 2006).
В соответствии с данными Токарева (2006), к светящимся организмам мы относили ктенофор Pleurobrachia pileus (O.F. Müller, 1776), Mnemiopsis leidyi A. Agassiz, 1865 и Beroe ovata Bruguière, 1789, динофлагелляту N. scintillans, циклопоидную копеподу Oithona similis Claus, 1866, кладоцеру Evadne spinifera P.E. Müller, 1867 и аппендикулярию Oikopleura (Vexillaria) dioica Fol, 1872.
Цель настоящей работы – исследование состава и количественных характеристик крупной фракции планктонных биолюминесцентов в пиках поля биолюминесценции в Черном море в летние месяцы.
МАТЕРИАЛ И МЕТОДИКА
Материал собирали в северной части Черного моря на 14 станциях, выполненных в научном рейсе № 108 НИС “Профессор Водяницкий” в июле–начале августа 2019 г. (рис. 1). Для учета крупной фракции светящегося планктона в слоях, соответствующих пикам интенсивности биолюминесценции, использовали ловы сетью Джеди с ячеей сита 115 мкм и входным отверстием 0.1 м2. Это позволило исправить недостаток методов учета биолюминесцентов, приведенных в работе Левина с соавторами (1975), когда пробу планктона брали батометром с одного горизонта и соотносили с интенсивностью биолюминесценции в слое определенной толщины, иногда достигающей нескольких десятков метров.
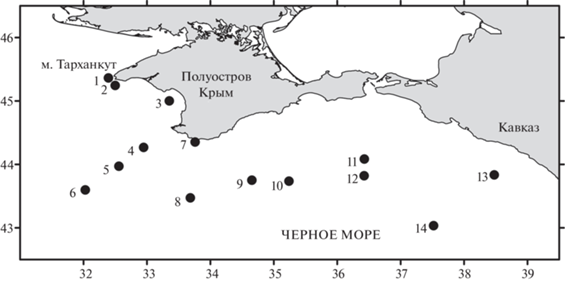
Таксономический состав и количественные характеристики биологического материала определяли по методике Яшнова (1934). К крупной фракции светящегося планктона относили биолюминесцентные организмы размером более 0.2 мм.
Данные о биолюминесценции, температуре и солености получили с помощью гидробиофизического комплекса “Сальпа-М” методом многократного зондирования (Гительзон и др., 1992; Токарев, 2006) от поверхности до глубины 120 м. Для уменьшения влияния астрономического фона в погружной части комплекса была встроена подвижная светоловушка, внутри которой располагалась теневая камера; датчик сигнала находился внутри рабочей камеры. При погружении комплекса поток воды проходил сквозь прибор. При минимальном сопротивлении набегающему потоку водной среды это обеспечивало ослабление астрономического фона в 2 × 107 раза и механическую стимуляцию светящихся гидробионтов. Дискретность синхронных измерений комплексом всех параметров при зондировании вниз со скоростью 1.2 м/с составляла 0.25 м, программное обеспечение интегрировало ее до 1 м. В результате обработки сигналов датчиков получены вертикальные профили биолюминесценции, температуры и солености; по этим параметрам была рассчитана условная плотность воды. Методика сбора и обработки информации с использованием комплекса “Сальпа-М” подробно описана ранее (Токарев и др., 2009). Данные по интенсивности биолюминесценции в пиках суммировали по вертикали, и исследовали взаимосвязь суммарного обилия планктонеров в этих пиках с кумулятивной интенсивностью биолюминесценции, а также с показателями температуры и солености в данных слоях.
РЕЗУЛЬТАТЫ
Слои, в которых отмечены максимальные величины интегральной биолюминесценции, значительно превышающие величины в ниже- или вышележащих слоях, принято называть пиками биолюминесценции. В ночное время на исследованных станциях зарегистрирован один пик, располагавшийся в слое термоклина на глубине 5–30 м (верхний пик). В дневное время выявлен пик биолюминесценции на глубине 50–82 м (нижний пик). В утреннее и вечернее время на станциях 4 и 8 зарегистрировано по два пика биолюминесценции (верхние, обозначенные как 4.1 и 8.1, и нижние – 4.2 и 8.2). Толщина пиков по вертикали варьировала от 5 до 15 м. Значения температуры и солености в пиках биолюминесценции на станциях представлены в табл. 1.
Таблица 1.
Параметры температуры и солености в пиках биолюминесценции
| № станции | Расположение слоя | Глубина слоя, м | Температура, °С | Соленость, ‰ | ||
|---|---|---|---|---|---|---|
| средняя | максимальная | средняя | максимальная | |||
| Ночь | ||||||
| 1 | Верхний | 12–26 | 15.72 | 22.81 | 17.61 | 18.10 |
| 2 | Верхний | 5–20 | 15.49 | 23.43 | 17.65 | 18.09 |
| 3 | Верхний | 20–30 | 14.90 | 16.62 | 17.88 | 17.99 |
| 6 | Верхний | 5–15 | 18.29 | 22.85 | 17.56 | 17.80 |
| 7 | Верхний | 8.5–18 | 16.35 | 19.06 | 17.90 | 18.05 |
| 11 | Верхний | 10–20 | 20.29 | 25.15 | 17.75 | 18.31 |
| 14 | Верхний | 7–12 | 22.13 | 26.33 | 17.29 | 18.12 |
| День | ||||||
| 4.1 | Верхний | 12–18 | 16.95 | 23.21 | 17.38 | 17.89 |
| 4.2 | Нижний | 76–80 | 9.14 | 9.16 | 20.19 | 20.27 |
| 5 | Нижний | 72–82 | 9.04 | 9.07 | 19.96 | 20.10 |
| 8.1 | Верхний | 21–25 | 14.45 | 18.21 | 17.23 | 17.65 |
| 8.2 | Нижний | 68–75 | 9.04 | 9.07 | 19.83 | 19.99 |
| 9 | Нижний | 50–60 | 9.13 | 9.17 | 19.71 | 19.96 |
| 10 | Нижний | 50–55 | 9.16 | 9.19 | 19.71 | 19.80 |
| 12 | Нижний | 71–80 | 9.00 | 9.04 | 20.12 | 20.23 |
| 13 | Нижний | 68–75 | 9.04 | 9.06 | 19.99 | 20.08 |
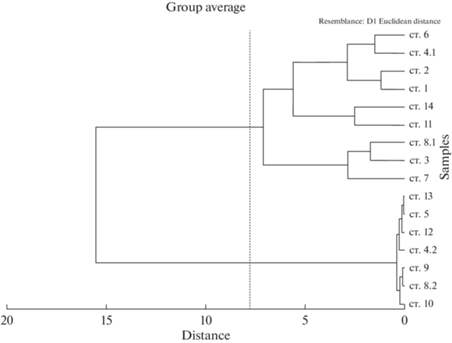
На основании построенной по данным температуры и солености дендрограммы сходства (мера расстояния – Эвклидово расстояние) в слое пика биолюминесценции выявлено несколько групп станций с высокой степенью сходства (рис. 2). В первой группе станций разброс значений температуры и солености был крайне мал: средняя температура составляла 9.00–9.16°С, соленость − 19.71–20.19‰ (табл. 1). В эту группу попали станции с нижним пиком биолюминесценции. Вторая группа − это станции, между которыми в слоях пиков биолюминесценции наблюдался гораздо больший размах данных по средней температуре и солености: температура изменялась от 16.62 до 26.33°С, соленость – от 17.23 до 17.90‰. В эту группу объединились станции с верхним пиком биолюминесценции (рис. 2); станции 3, 7 и верхний пик станции 8 образовали подгруппу, которая характеризовалась максимальными значениями суммарной биолюминесценции по слою. Отмечено, что на этих трех станциях максимальная температура в слое пика биолюминесценции не превышала 19.06°С, в то время как на глубоководных станциях в восточной части Черного моря (станции 11 и 14) вода прогревалась до 25.15–26.33°С, соленость достигала 18.12–18.31‰ (табл. 1).
Рис. 2.
Дендрограмма сходства станций по Эвклидову расстоянию на основании данных по температуре и солености.
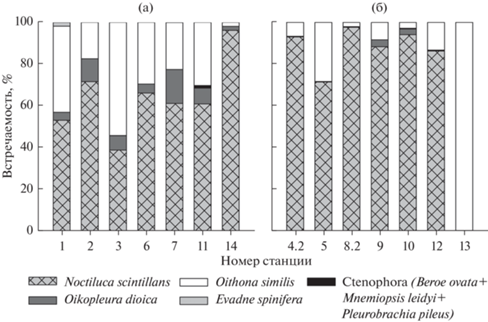
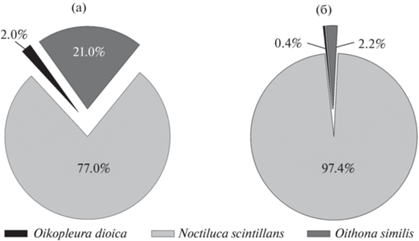
Суммарная численность светящегося планктона в пиках биолюминесценции колебалась от 45 экз./м3, что соответствовало кумулятивной интенсивности биолюминесценции 1251/1012 Вт/(см2 × л), до 10 457 экз./м3 для максимального зарегистрированного пика, кумулятивная интенсивность свечения в котором достигала 53 146/1012 Вт/(см2 × л). Биолюминесцентные виды составляли от 8 до 75% общей численности планктона. Основой крупной фракции светящегося планктона Черного моря в порядке убывания численности являлись Noctiluca scintillans, Oithona similis и Oikopleura dioica; остальные организмы были малочисленны (рис. 3).
Рис. 3.
Вклад отдельных видов в численность крупной фракции светящегося планктона Черного моря в верхнем ночном (а) и нижнем дневном (б) пиках биолюминесценции.
Значения зарегистрированных ночью пиков интенсивности биолюминесценции были существенно выше показателей, полученных в дневное время. Это обычное явление, как и значительное различие в количестве светящихся планктонных организмов в слоях биолюминесценции в разное время суток. Максимальное обилие светящихся планктонных организмов было отмечено в ночное время, когда пики биолюминесценции наблюдались в верхних слоях воды на глубине 5–30 м. Максимальный пик интенсивности биолюминесценции 53 146/1012 Вт/(см2 × л) отмечен на станции 3 в слое толщиной 10 м на глубине 20–30 м. На этой станции были зарегистрированы максимумы численности O. similis, N. scintillans, O. dioica и Pleurobrachia pileus.
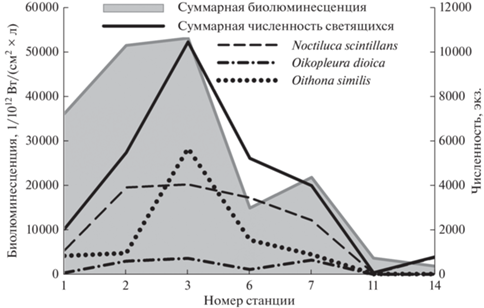
Для верхнего пика была характерна сильная положительная связь суммарной интенсивности биолюминесценции с обилием трех доминирующих видов: для N. scintillans и O. dioica R = +0.79, для O. similis R = +0.70 и для общей численности светящихся R = +0.82. По численности доминировала динофлагеллята N. scintillans, за исключением станции 3 в Каламитском заливе, где преобладала копепода O. similis (рис. 3а). Количество N. scintillans в верхних пиках достигало 4 тыс. экз./м3; в нижних пиках численность составляла 200–300 экз./м3, а зарегистрированный максимум не превышал 900 экз./м3 (рис. 4). Значимая отрицательная зависимость численности N. scintillans от средней температуры воды и максимальных значений солености отмечена в ночном верхнем пике, она составляла –0.68 и –0.72 соответственно.
Рис. 4.
Суммарное значение биолюминесцентного сигнала и численность светящихся видов в верхнем пике биолюминесценции.
Копепода O. similis была наиболее многочисленной в верхних пиках с максимумом 5675 экз./м3 на станции 3 у западного побережья Крыма напротив мыса Тарханкут; средняя по станциям численность O. similis в слое пика биолюминесценции составляла 676 экз./м3. В популяции преобладали самки, их доля на некоторых станциях достигала 60%, и пятые копеподитные стадии (до 48%). Заметная, но не тесная, степень положительной связи численности с интенсивностью биолюминесценции (+0.66) может свидетельствовать о наличии биолюминесцентной реакции не у всех стадий копеподы, однако для однозначного вывода необходимы дальнейшие лабораторные исследования. Отмечена высокая степень отрицательной связи численности вида с максимальной температурой в пиках (R = –0.83).
Средняя по станциям численность O. dioica в слое пика биолюминесценции составляла 145 экз./м3. В верхнем пике обилие аппендикулярии было на 2 порядка выше, чем в нижнем. Как и для копеподы O. similis, для O. dioica отмечена высокая степень отрицательной связи численности с максимальной температурой в пиках (R = –0.83).
Только 6 экз. гребневика P. pileus зарегистрированы на станции 3, на остальных станциях этот вид был обнаружен в единичных экземплярах или отсутствовал. Из других представителей ктенофор однажды встречен Mnemiopsis leidyi, гребневик B. ovata в пробах отсутствовал. Кладоцера E. spinifera в небольшом количестве обнаружена на трех станциях.
На всех станциях в нижнем и верхнем пиках с долей 72–97% в общей численности светящихся доминировала N. scintillans. Исключением была станция 13 у кавказского побережья, где светящиеся виды были представлены только O. similis (рис. 3б).
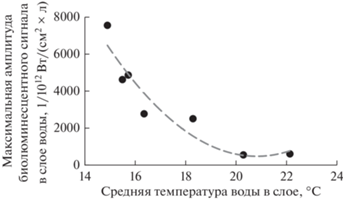
Выявлена сильная отрицательная зависимость между максимальными значениями амплитуды биолюминесценции в верхних пиках и средними значениями температуры в соответствующих слоях (R = –0.91). С повышением температуры величина максимальной интенсивности биолюминесценции уменьшалась. Для слоя нижнего пика выявлена высокая положительная связь амплитуды биолюминесценции с абиотическими параметрами; коэффициент корреляции интенсивности биолюминесценции с максимальными температурой и соленостью в этом пике составил соответственно +0.81 и +0.83. Значимой корреляции в нижних пиках между биолюминесценций и численностью биолюминесцентных видов не обнаружено.
На станциях 4 и 8 в вечернее и утреннее время отмечено два биолюминесцентных пика (рис. 5). Интенсивность биолюминесценции в верхнем пике была значительно выше, чем на горизонтах, расположенных ниже. Вертикальный профиль нижнего пика более пологий в отличие от островершинного верхнего пика, поэтому кумулятивная интенсивность биолюминесценции нижнего пика может превосходить таковую верхнего пика и вносить больший вклад в общую интенсивность биолюминесценции по всему столбу воды, как это наблюдалось на станции 8.
Рис. 5.
Вертикальные профили биолюминесценции на станциях 4 (а) и 8 (б) с двумя пиками поля биолюминесценции.
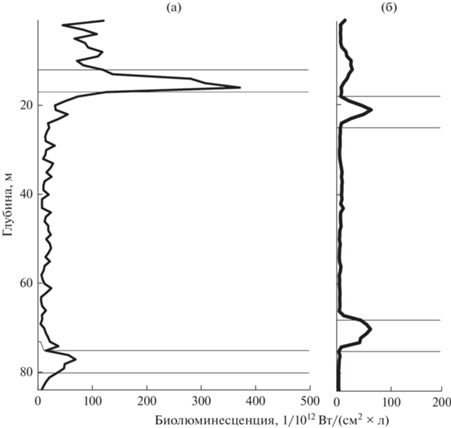
На станциях с двумя пиками интенсивности биолюминесценции сохранялось доминирование N. scintillans. Вклад O. similis по численности в верхнем пике был на порядок больше, чем в нижнем, и составлял 21%. Численность O. dioica не превышала 2% (рис. 6).
ОБСУЖДЕНИЕ
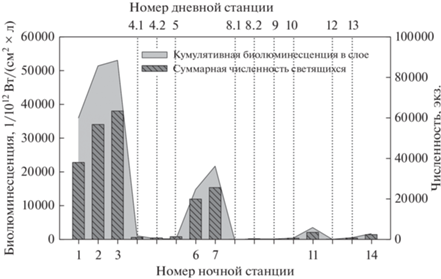
Максимальные величины кумулятивной биолюминесценции наблюдались в ночное время в верхних пиках биолюминесценции на мелководных (глубина 30–90 м) станциях 1–3 и 7, расположенных вдоль юго-западного прибрежья п-ва Крым (рис. 1, 4). Данные величины на два порядка превышали значения, отмеченные для глубоководных районов в нижних пиках биолюминесценции в дневное время. Существенных значений кумулятивная биолюминесценция достигала также на станции 6, находившейся в западной халистазе. На исследуемых горизонтах на этих станциях наблюдалось наибольшее обилие представителей крупной фракции биолюминесцентного планктона (рис. 7).
Рис. 7.
Кумулятивный биолюминесцентный сигнал и численность светящихся видов крупой фракции биолюминесцентов в дневное и ночное время.
На большинстве станций в течение суток по численности доминировала динофлагеллята Noctiluca scintillans (рис. 3), уровень биолюминесцентного сигнала которой может достигать 1/103 мкВт/см2 (Токарев и др., 2016). Очевидно, этот вид вносил значительный вклад в поле биолюминесценции крупной фракции планктонных видов. На доминирующую роль N. scintillans в свечении морской воды указывали и ранее (Токарев и др., 2000; Заворуев, 2002).
Биолюминесценция копеподы Oithona similis оценена в 4.6/102 мкВт/см2 (Евстигнеев, Битюков, 1990; Токарев и др., 2016), что на порядок больше, чем у N. scintillans. Следовательно, вклад O. similis в кумулятивную биолюминесценцию также может достигать больших величин. Согласно нашим данным, свечение зарегистрировано на станции 13, где крупная фракция светящегося планктона была представлена только O. similis. Однако на станциях 2 и 3, на которых величина кумулятивной биолюминесценции была высокой и почти одинаковой (рис. 7), численность этой копеподы различалась в 7 раз (839 и 5675 экз./м3 соответственно). Прогрев воды на исследуемых горизонтах значительно различался (табл. 1), что также могло иметь значение для параметров свечения.
Аппендикулярия Oikopleura dioica в массовом количестве присутствовала в верхних пиках биолюминесценции ночью на прибрежных станциях 2, 3 и 7, где ее численность достигала 600–725 экз./м3; на глубоководной станции 6 обилие этого вида составляло 225 экз./м3, в остальных районах не превышало 15 экз./м3. В литературе значения биолюминесцентного сигнала для O. dioica не приводятся, поэтому сложно судить о ее вкладе в кумулятивный биолюминесцентный сигнал.
Ктенофоры, индивидуальная интенсивность биолюминесценции которых превышает таковую у других планктонных организмов и должна существенно влиять на суммарный сигнал, встречались лишь в единичных экземплярах; они отмечены на станциях с максимальной интенсивностью биолюминесценции и на станциях, где свечение было крайне мало.
Значимой корреляции кумулятивной биолюминесценции с температурой и соленостью воды не обнаружено, однако выявлена сильная обратная зависимость между максимальными значениями амплитуды интенсивности поля биолюминесценции в верхних пиках в ночное время и средними значениями температуры в соответствующем слое. Максимум амплитуды биолюминесцентного сигнала приходится на слои с температурой воды 15°С; при прогреве свыше 20°С значения амплитуды минимальны (рис. 8).
Рис. 8.
Зависимость максимальных значений интенсивности поля биолюминесценции в верхнем пике от средней температуры воды в этом слое.
В настоящее время отсутствуют сведения о влиянии глубины, на которой были выловлены гидробионты, на долю высветившихся (т.е. проявивших реакцию в виде вспышки) в эксперименте особей разных видов. Однако не исключено, что этот параметр также может влиять на энергию свечения. Возможно, свой вклад в поле биолюминесценции вносят и другие зоопланктонные виды, способность к свечению которых еще не изучена. На современном этапе исследований сложно определить, какой вид вносит наибольший вклад в общую интенсивность биолюминесценции.
На основании полученных нами данных можно сделать следующие выводы:
1. В летнее время в Черном море крупная фракция биолюминесцентных планктонных организмов в пиках биолюминесценции чаще всего представлена динофлагеллятой N. scintillans и копеподой O. similis, причем в большинстве случаев доминировала N. scintillans, в остальных – O. similis.
2. Наибольшая численность светящихся видов обнаружена в прибрежных районах Крыма.
3. Доля крупной фракции планктонных биолюминесцентов составляла 8−75% от общей численности планктонных видов.
4. В верхнем пике биолюминесценции в ночное время отмечено большое количество светящихся видов и зарегистрированы высокие значения кумулятивной биолюминесценции. Обнаружена сильная корреляционная связь максимальной амплитуды биолюминесценции с температурой.
5. Для верхних пиков биолюминесценции в ночное время найдена значимая связь между температурой в слое пика и численностью трех видов крупной фракции биолюминесцентов: N. scintillans, O. similis и O. dioica.
6. Различие между величинами биолюминесценции в пиках обусловлено не только таксономическим составом светящихся видов, но и циркадным ритмом свечения.
Список литературы
Брянцева Ю.В., Серикова И.М., Суслин В.В. Межгодовая изменчивость разнообразия динофлагеллят и поля биолюминесценции у берегов Севастополя // Экосистемы, их оптимизация и охрана. 2014. Вып. 11. С. 158–164.
Гительзон И.И., Левин Л.А., Утюшев Р.Н. и др. Биолюминесценция океана. СПб.: Гидрометеоиздат. 1992. 283 с.
Евстигнеев П.В., Битюков Э.П. Биолюминесценция морских копепод. Киев: Наукова думка. 1990. 144 с.
Евстигнеев П.В., Битюков Э.П., Околодков Ж.Е. Видовой состав и специфичность биолюминесценции водорослей Dynophyceae // Ботан. журн. 1993. Т. 78. С. 1−15.
Заворуев В.В. Суточные изменения биолюминесценции планктона Черного и Эгейского морей и корреляция свечения с концентрацией хлорофилла // Исследовано в России (электронный ресурс). МФТИ. http://zhurnal.mipt.rssi.ru. 2002. С. 2204–2214.
Левин Л.А., Утюшев Р.Н., Артемкин А.С. Распределение интенсивности биолюминесцентного поля в экваториальной части Тихого океана // Тр. Ин-та океанол. им. П.П. Ширшова. 1975. Т. 102. С. 94–101.
Полонский А.Б., Мельникова Е.Б., Серебренников А.Н. и др. Региональные особенности интенсивности свечения гидробионтов и концентрации хлорофилла а в водах Черного моря // Оптика атмосферы и океана. 2018. Т. 31. Вып. 4. С. 275–281.
Серикова И.М., Брянцева Ю.В., Токарев Ю.Н. и др. Особенности сезонной динамики поля биолюминесценции и биомассы светящихся динофлагеллят у Севастополя (2008–2009 гг.) // Наук. зап. Терноп. нац. пед. ун-ту. Сер. Біол. 2010. № 3(44). С. 230–234.
Серикова И.М., Токарев Ю.Н., Евстигнеев В.П., Суслин В.В. Поле биолюминесценции как показатель агрегированности динофитовых водорослей в Черном море и его сезонная и межгодовая динамика // Оптика атмосферы и океана. Физика атмосферы. Материалы XXIII Международ. симпоз. Конференция C. Исследование атмосферы и океана оптическими методами. https://symp.iao.ru/ru/aoo/23/proceedings. 2017. C. С455–С458.
Токарев Ю.Н. Основы биофизической экологии гидробионтов. Севастополь: ЭКОСИ-Гидрофизика. 2006. 342 с.
Токарев Ю.Н., Битюков Э.П., Василенко В.И. и др. Поле биолюминесценции – характерный показатель структуры планктонного сообщества Черного моря // Экол. моря. 2000. Вып. 53. С. 20–25.
Токарев Ю.Н., Василенко В.И., Жук В.Ф. Новый гидробиофизический комплекс для экспрессной оценки состояния прибрежных экосистем // Современные методы и средства океанологических исследований. Материалы XI Международ. научно-техн. конф. Ч. 3. 25–27 ноября 2009. М.: Изд-во РАН. 2009. С. 23–27.
Токарев Ю.Н., Евстигнеев П.В., Машукова О.В. Планктонные биолюминесценты Мирового океана: видовое разнообразие, характеристики светоизлучения в норме и при антропогенном воздействии. Симферополь: Нижняя Орианда. 2016. 340 с.
Черепанов О.А., Левин Л.А., Утюшев Р.Н. Связь биолюминесценции с биомассой и численностью светящегося и всего планктона. 2. Черное море // Мор. экол. журн. 2007. Т. 6. Вып. 3. С. 84–89.
Яшнов В.А. Инструкция по сбору и обработке планктона. М.: ВНИРО. 1934. 40 с.
Haddock S.H.D., Moline M.A., Case J.F. Bioluminescence in the sea // Annu. Rev. Mar. Sci. 2010 V. 2. № 1. P. 443−493.
Martini S., Haddock S.H.D. Quantification of bioluminescence from the surface to the deep sea demonstrates its predominance as an ecological trait // Sci. Rep. 2017. V. 7. 45750. https://doi.org/10.1038/srep45750
Дополнительные материалы отсутствуют.