

Биология внутренних вод, 2019, № 1, стр. 3-11
Закономерности организации речных экосистем: ретроспектива становления современных концепций (Обзор)
Н. С. Батурина *
Новосибирский государственный университет
630090 Новосибирск, ул. Пирогова, 2, Россия
* E-mail: Natalya.s.baturina@gmail.com
Поступила в редакцию 01.11.2017
После доработки 16.02.2018
Принята к публикации 19.12.2017
Аннотация
Представлен обзор развития базовых концепций, описывающих организацию речных экосистем. Рассмотрены первые зональные модели описания рек, переход к континуальным концепциям, их последующая критика и развитие дисконтинуальных моделей. Описаны основные дискретные концепции как главные альтернативные подходы к описанию устройства рек. Отражены основные направления развития идей об иерархичности организации речных систем и их трансформирование в современные комплексные концепции.
ВВЕДЕНИЕ
За последние четыре десятилетия предпринято немало попыток объяснить устройство речных экосистем. Создание концепции, предсказывающей изменчивость структуры речной системы в пространстве и времени, представляется наиболее важной задачей для исследователей водотоков. Задача становится еще более увлекательной, если учесть, что реки – это открытые, крайне изменчивые в пространстве экосистемы, в которых преобладает снос вещества по продольному градиенту [15].
Первые разработанные концепции создали базу для дальнейшего развития представлений о комплексности речных экосистем. Среди них концепции речного континуума [36], концепция спиралинга питательных веществ [38], последовательной дисконтинуальности [37], пульсирующих наводнений [17], придонного коридора [31]. Последующее развитие представлений об устройстве речных экосистем отошло от описания линейных моделей – реки стали рассматриваться как дискретные иерархические системы. Такой подход отражен в концепциях динамики пятен [35], динамики речных систем [7], приоритета геоморфологических процессов [20], иерархичности структуры классификации местообитаний [13]. Последним этапом в формировании представлений об организации речных систем стало развитие комплексных и синтетических подходов, объединяющих ранее разработанные модели. В результате были созданы концепции синтеза речных экосистем [32], лотических гидросистем [24] и реобиома [3].
В работе приводится ретроспектива развития представлений об устройстве речных экосистем, критика наиболее распространенных концепций и современные представления, появившиеся в результате комбинирования и синтетического обобщения разных концепций и подходов.
Зональный подход к описанию речных систем
Первые попытки описания структурной организации речных экосистем появились в конце XIX в. Они сводились к выделению речных зон на основе распространения определенных видов рыб [22]. Последующим этапом стало выявление абиотических параметров, характеризующих эти зоны, и описание сообществ беспозвоночных. Уже в начале XX в. опубликована первая работа, детально описывающая шесть речных зон, характерных для центральной Европы [34]. Однако применение этой классификации для других регионов было затруднительно, и спровоцировало развитие универсальных подходов к описанию рек. Изначально выделяли только верхнюю зону – быстротекущую (эрозионную) и нижнюю – медленнотекущую (аккумулятивную) [29]. Позднее была проведена более детальная классификация, представленная в концепции речных зон [16], выделено три основных зоны: креналь (собственно исток реки), ритраль и потамаль, нижележащие участки ритрали разделены на зоны эпиритрали, метаритрали и гипоритрали. В качестве параметров среды значимых при определении границ между зонами были использованы факторы, лимитирующие распространение рыб – содержание кислорода, характер грунта, амплитуда среднемесячных температур воды, угол уклона русла, линейная скорость течения [27].
На сегодняшний день подход к описанию речных систем как ряда последовательно расположенных зон, отличающихся по определенным характеристикам среды, во многом уже утратил свою популярность. Одной из наиболее выдающихся в рамках этого подхода можно считать работу Герберта Хоукса “Классификации и зонирование рек” с классификацией рек Великобритании, основанную на физико-химических характеристиках водотоков и видовом составе беспозвоночных [14].
Континуальные модели речных систем
В основу континуальных подходов положена идея о непрерывном, последовательном изменении физических параметров реки и распределении вещества в пределах системы, что отражается на структуре биотических сообществ.
Концепция спиралинга питательных веществ (КСПВ). Необходимость в формировании универсальной концепции подтолкнула многих исследователей к рассмотрению речной системы с точки зрения синтеза, транспорта и накопления энергии и вещества [8–11, 19, 21]. Одна из таких моделей описана в концепции спирального оборота, или спиралинга, питательных веществ – КСПВ [21, 38]. Согласно КСПВ, биогенные вещества в речной системе перемещаются свободно в воде до момента их потребления живыми организмами. После отмирания организмов биогенные элементы снова попадают в поток и могут быть повторно использованы биотой [39]. Поскольку в реках преобладает направленное движение водных масс, то наблюдается постоянное смещение органического вещества в системе, что приводит к образованию частично открытых циклов оборота вещества в виде спиралей [12]. Частота расположения оборотов спирали зависит от мощности потока, с одной стороны, интенсивности ассимиляции и объема накопления органического вещества биотой – с другой. При стабильности гидрологического режима потока, чем короче расстояние между петлями спирали, тем интенсивнее используют вещество сообщества. По частоте оборота спирали, мощности и диаметру выделяют типы речных сообществ [19], каждый из которых соответствует определенной степени выносливости/устойчивости экосистемы.
Концепция речного континуума (КРК). Многочисленные исследования медленно текущих равнинных и равнинно-низкогорных рек лесных регионов воплотились в модель, описанную в одной из самых популярных концепций XX в. – КРК [36].
КРК подразумевает, что целостность и устойчивость речной системы детерминируется непрерывностью потока вещества и энергии, характер которого на всем протяжении водотока определяется влиянием континуума физических параметров среды [36]. Следовательно, процессы накопления, транспорта, утилизации и синтеза органического вещества происходят непрерывно вдоль русла реки [33, 36]. В каждой конкретной точке водотока органическое вещество, необходимое для формирования биологической продукции консументов, поступает из трех основных источников: локальные смывы органического вещества из прибрежных систем (аллохтонная продукция), первичная продукция водных растений (автохтонная продукция) и транспорт органического вещества с вышерасположенных участков. Соотношение величин этих трех типов продукции меняется на протяжении русла. КРК рассматривает бентосные сообщества как один из основных компонентов аккумуляции органического вещества. Структура бентосных сообществ зависит от соотношения типов органического вещества на каждом конкретном участке русла. Таким образом, количественные и качественные характеристики потока органического вещества детерминируют структуру биотических сообществ.
Критика КРК и дисконтинуальные модели
Публикация первой универсальной модели – КРК, стало отправной точкой для создания альтернативных моделей речных систем, построенных на критике КРК [18, 23, 30].
Концепция последовательной дисконтинуальности (КПД). При рассмотрении гидрологического режима потока было отмечено, что для многих рек наблюдается антропогенное изменение режима стока. Возведение дамб, плотин, каналов нарушает континуальность речных систем. Следовательно, КРК не может быть применима для описания организации таких рек. Разработана КПД [37], где подчеркивается, что реки становятся дисконтинуальными при постройке гидротехнических сооружений. Однако согласно КПД, КРК верно описывает изменчивость биотических сообществ в пределах нетронутых участков. Еще одна модель, в которой основное значение придается гидрологическому режиму, представлена в парадигме природного режима стока [25]. В ее рамках стабильность естественного режима стока считается основным фактором поддержания разнообразия лотических экосистем.
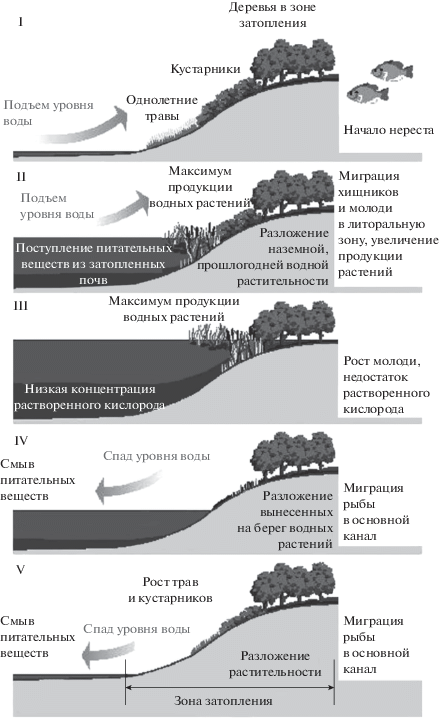
Концепция пульсирующих наводнений (КПН). В отличие от КРК, сформированной на исследованиях медленно текущих равнинных и низкогорных рек, КПН разработана на основе исследований водотоков тропического пояса, где огромное влияние на устройство экосистемы оказывают сезонные наводнения [17]. Согласно КПН, при увеличении размеров поймы реки снижается частота наводнений и увеличивается их прогнозируемость. Следовательно, крупные реки имеют более стабильный режим стока. Согласно КПН, обмен органическим веществом между рекой и поймой при стабильном режиме наводнений в значительной степени влияет на структуру сообществ. Зоны затопления рассматриваются как транзитные участки между наземными и водными экосистемами, где происходит активный обмен веществом (рис. 1). Таким образом, в роли фактора, определяющего структуру речной системы, выступает годовой режим стока.
Концепция динамики речных сетей (КДРС). Один из существенных моментов в КРК, КПН и КПД – рассмотрение рек как линейных систем (редко встречается в природе). Реки, как правило, представляют собой целую сеть водотоков, формирующих речной водосборный бассейн. В КДРС предпринята попытка отразить именно сетевой характер речных систем [7]. Согласно КДРС, в организации речной сети огромную роль играют боковые притоки. После впадения притока меняется гидрологический режим основного русла, также существенно изменяется энергетический баланс системы. Воды притока несут органическое вещество с вышерасположенных участков, где активно происходит синтез автохтонной и накопление аллохтонной продукции. В местах впадения боковых притоков наблюдается формирование “горячих точек” видового разнообразия, что также не соответствует КРК.
Концепция придонного коридора (КПК). КПК описывает модель устройства речной системы, где основная роль отводится придонным зонам русла и подземным водам. Придонные зоны русла (hyporheic zones) – это пространственные зоны, образованные элементами донного субстрата, между водным потоком и породами, слагающими речное русло [31]. Вода может поступать туда как из русла реки, так и из подземных источников. Данная зона является местообитанием для многочисленных водных беспозвоночных, бактериальных и водорослевых сообществ. Кроме того, в придонной зоне происходит циркуляция газов, накопление, транспорт и минерализация питательных веществ, пополнение водоносного прируслового слоя, обмен с прибрежными системами. Следовательно, глубина придонной зоны и тип образующего ее субстрата во многом определяет организацию речной биоты.
Дискретные модели речных систем
Дискретные модели описывают реки как “экологическую мозаику”, элементы которой представлены биотическими сообществами. Причем, структура каждого сообщества определяется локальными геоморфологическими характеристиками русла, так и параметрами выше- и нижерасположенных участков [26].
Концепция иерархической структуры классификации местообитаний (КИСКМ). Если рассматривать речные системы как мозаику из сообществ с разной структурой, то неизменно встает вопрос о масштабе рассмотрения системы. КИСКМ предлагает модель речной системы, основанную на пространственных и временных изменениях среды [13]. На каждом уровне рассмотрения системы выделяются сообщества, структура которых строго определена локальными условиями среды. При изменении масштаба рассмотрения системы проявляется эффект иерархичности данной модели. Участки “экологической мозаики” одного масштаба оказываются “вложенными” в “мозаику” другого масштаба (рис. 2). Такая иерархическая модель позволяет учитывать влияние множества переменных и дает возможность использовать данные из разных источников.
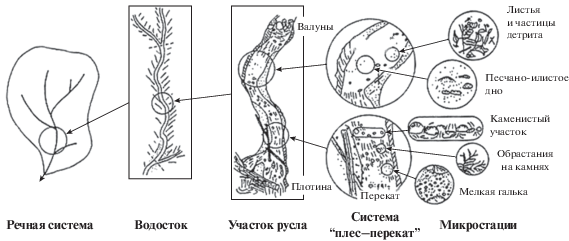
Концепция динамики пятен (КДП). В качестве основной альтернативы КРК среди дискретных концепций долгое время выступала КДП [35, 40]. Модель данной концепции отражает пространственную гетерогенность и временнyю изменчивость участков речной сети. В рамках КДП получило развитие представление о рефугиумах – убежищах гидробионтов, потенциально значимых в случаях реколонизации разрушенных/нарушенных сообществ [28]. Согласно КДП, большое разнообразие гидробионтов обеспечивается постоянными изменениями условий среды в пространстве водотока. Рефугиумы рассматриваются как “островки безопасности” при техногенной или природной трансформации русла рек. Следовательно, формирование рефугиумов – один из путей поддержания устойчивости экосистемы через сохранение разнообразия.
Концепция приоритета геоморфологических процессов (КПГП). Еще одна иерархическая модель организации речных систем, описанная Монтгомери в КПГП, указывает на значимость местных геоморфологических параметров и высотного положения участков русла реки при формировании биотических сообществ [20]. В этой работе подчеркивается, что при изменении абсолютной высоты происходит изменение геоморфологических характеристик русла, что определяет формирование особых “зон геоморфологического приоритета”, где характер локальных физических параметров среды детерминирует структуру сообществ в каждой точке водотока. КПГП указывает, что КРК применима только для низкорельефных водоразделов с относительно постоянным климатом и простым геоморфологическим устройством. Однако для регионов с высокогорным рельефом, изменчивым климатом, сложным геологическим строением речных долин необходимо применять КПГП.
Современные концепции речных систем
Большинство вышеперечисленных концепций постулирует, что физические параметры среды определяют биологическую структуру речных сообществ. При этом в каждой точке потока достигается динамическое равновесие между биотическими и абиотическими компонентами системы. Однако такой подход является слишком упрощенным для создания реалистичной, прогнозируемой модели речной системы, применимой повсеместно. Безусловно, разработанные концепции оказались применимы в частных случаях, но потребность в создании универсальной комплексной модели оставалась актуальной.
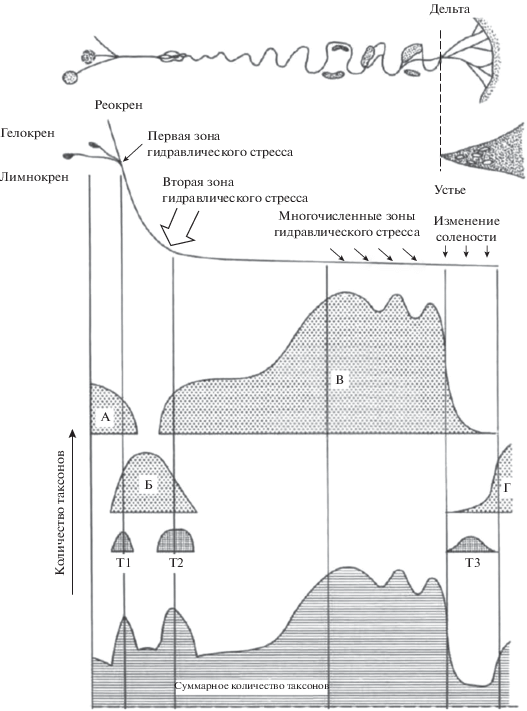
Концепция лотических гидросистем (КЛГ). Одна из наиболее популярных моделей представлена в КЛГ, где реки рассматриваются как трехмерные пространственные системы (рис. 3), изменяющиеся во времени. Структура таких систем определяется продольными, латеральными и вертикальными потоками вещества и энергии [24]. Таким образом, учитываются обмен с грунтовыми водами (КПК) – вертикальное измерение; снос вещества и изменчивость физических параметров потока вдоль русла реки (КРК, КПД, КСПВ) – продольное измерение и обмен с прибрежными экосистемами (КПН) – латеральное измерение. Следовательно, в рамках данной концепции гидрологический режим и геоморфологические характеристики русла реки определяют разнообразие типовых местообитаний, а также интенсивность, продолжительность и частоту взаимодействий между населяющими их сообществами. Кроме того, данный подход рассматривает речную систему как иерархическую модель, состоящую из множества речных подсистем, где на каждом уровне организации значимы конкретные физические процессы, осуществляемые в водотоке. Например, при мелком масштабе рассмотрения системы, структуру сообществ определяет распределение в потоке органического вещества вдоль продольного и поперечного профиля русла, что согласуется с КРК и КПН. При переходе на средний масштаб рассматриваются сообщества в системе заводей и примыкающих перекатов (КПК), а при крупном масштабе рассмотрения – распределение конкретных видов в соответствии с давлением потока воды в пределах переката. Следовательно, выбор масштаба рассмотрения речной системы определяет уровень детальности описания ее структуры. Кроме того, необходимо помнить, что факторы, детерминирующие распространение видов в водотоке, могут отличаться для разных участков русла (например, для верховий и среднего течения) и разных речных систем.
Рис. 3.
Общая модель продольной организации речной системы согласно КЛГ (со стандартными характеристиками потока) ([24], с модификациями). А – виды, населяющие ручьи и кренальную зону рек; Б – виды, обитающие на участках с большим уклоном русла; В – виды метаритрали; Г – морские виды; Т1, Т2, Т3 – виды, обитающие в зонах гидравлического удара. Стрелка – изменение количества таксонов в различных зонах реки.
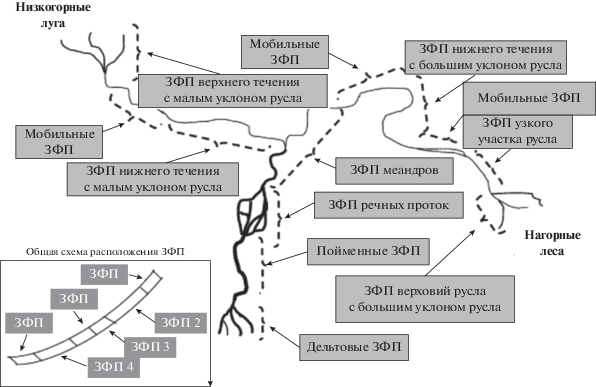
Концепция синтеза речных экосистем (КСРЭ). В КСРЭ [32] предпринята попытка объединить представления, сформировавшиеся с момента публикации КРК. В данной концепции биокомплексность – это структурный и функциональный феномен, появившийся благодаря динамическим взаимодействиям между биотическими компонентами экосистемы и физико-химическим параметрами среды. Реки рассматриваются как биокомплексные “массивы” или сети, состоящие из множества геоморфологически неоднородных участков [32]. Такие участки обозначаются как “зоны функциональных процессов” (ЗФП). Идея о ЗФП восходит к представлениям о “зонах приоритета геоморфологических процессов”. Однако, в отличие от КПГП, которая противоречит континуальным представлениям о речных системах КРК, КСРЭ их не отрицает. Согласно КСРЭ, реки – это открытые системы, структура которых в каждой точке во многом зависит от сноса веществ с вышерасположенных участков русла. Следовательно, границы ЗФП определяются не только согласно изменению ландшафта и высотному положению участка русла (как в КПГП), но и характером гидрологического режима, типом пойменных и наземных экосистем, соотношением автохтонной и аллохтонной продукции [32]. Гидрогеоморфологическая неоднородность русла – это та матрица, на основе которой ЗФП могут быть визуализированы. Однако ЗПФ сами по себе описывают участки русла, где формируются функционально различные сообщества, структура которых определяется физическими параметрами среды. Остается открытым вопрос, как ЗФП располагаются вдоль русла реки, есть ли какая-то строгая система их пространственного и порядкового расположения. Возможно, КСРЭ имеет гораздо больше общего с КРК, чем кажется на первый взгляд. Характеристики речного потока (гидрологический режим, геоморфологическое строение русла) могут изменяться последовательно при изменении высоты расположения участков русла, но не непрерывно. Следовательно, КСРЭ указывает на то, что КРК может достоверно предсказать лишь некоторые особенности в организации ЗФП.
На рис. 4 показаны различные ЗФП, сформированные геоморфологической неоднородностью русла от верховий водотока (лесные и луговые экосистемы) до дельты реки [33]. Одна и та же ЗФП может быть представлена в разных местах вдоль русла водотока. Если рассматривать особенности оборота вещества в ЗФП, то для верховий характерно меньшее время удержания органического вещества, слабо выраженные аккумулятивные процессы, в отличие от нижнего течения. Время удержания органического вещества – весьма изменчивый показатель, во многом зависящий от геоморфологической неоднородности русла [33], что согласуется с некоторыми тезисами концепции спиралинга питательных веществ.
Рис. 4.
Схематическое изображение сложной речной системы ([32], с модификациями). ЗФП ‒ зоны функциональных процессов. На выноске – порядок расположения вдоль русла реки одинаковых ЗФП и различных ЗФП (2–4).
Таким образом, КСРЭ представляет собой универсальную модель, которую с легкостью можно применить для исследования как горных, так и равнинных рек, расположенных в разных климатических зонах.
Концепция реобиома. Многочисленные исследования водотоков Дальнего Востока позволили разработать комплексную модель организации речных систем, описанную в комбинированной концепции (КК) [2, 3]. Согласно КК, многие альтернативные концепции организации речных систем (КДРС, КСПВ, КДП, КПН) не противоречат КРК, а дополняют ее [4]. С получением новых данных по исследованию рек европейской и центральной части России [5], дополнивших КК, сложились предпосылки для создания концепции реобиома (КР). Под реобиомом понимается надсистемная форма организации природного речного комплекса – континуума речных экосистем [1].
При изучении крупных и средних рек, чей водосборный бассейн расположен в нескольких физико-географических и климатических зонах, стало очевидно, что организация такой речной биосистемы зависит от континуальности гидрологических процессов [6]. Безусловно, рассматривать всю речную систему как последовательные и непрерывные градиенты изменений физических параметров потока невозможно. Однако в условиях неизменности геоморфологического устройства русла, можно наблюдать процессы последовательного и непрерывного перемещения органического вещества в системе и однотипность механизмов его фиксации в биоценозах. Следовательно, крупные и средние реки следует рассматривать как реобиомы – совокупность речных подсистем, образующих континуальную надсистемную структуру.
Изменение структуры сообществ вдоль русла может иметь как плавный, так и скачкообразный характер, что зависит от степени изменчивости абиотических параметров вдоль русла реки. В формировании континуальности структуры реобиома участвуют все входящие в него экосистемы, но не напрямую, а опосредовано, через механизмы транспорта питательных веществ и формирования рефугиумов в следующих друг за другом локальных биотических сообществах [3]. Следовательно, в речных биомах обнаруживается структура связей, обеспечивающих взаимодействие между разными подсистемами реобиома (речными экосистемами). Такие подсистемы не автономны, поскольку их связывают дрифтовые процессы, слабо влияют на структуру соседних подсистем, но обладают сильными внутренними двусторонними связями между биотической и абиотической компонентой [3]. Такая организация надсистемных природных комплексов позволяет сохранять высокую способность к выживанию всей системы даже при ликвидации одной из подсистем.
ЗАКЛЮЧЕНИЕ
Исследования экологии речных систем в бóльшей мере сфокусированы на обсуждении процессов, определяющих структурную организацию биоты, и на разработке концепций, описывающих механизм функционирования речных экосистем.
Рассмотренные в данной работе базовые концепции представляют континуальные, дискретные и иерархические подходы к описанию организации речных сообществ. Первоначальный вектор к описанию рек, как линейных моделей, вызвал много критики со стороны исследователей, что было толчком к формированию дискретных моделей описания речных систем. Следующий шаг – создание более сложных трехмерных моделей с иерархической структурой. Концепции, описывающие такие модели, объединяют идеи, разработанные в КРК, КСПВ, КДП, КПН, КПД, КПК, КПГП, и рассматривают реки как надсистемные природные комплексы, состоящие из разного количества подсистем, в зависимости от масштаба рассмотрения. В пределах одной речной подсистемы геоморфологические параметры, процессы синтеза и накопления органического вещества определяют структуру сообществ, а целостность подсистемы поддерживается сетью прямых и обратных биотических связей внутри сообществ.
Список литературы
Богатов В.В. Экология речных сообществ Российского Дальнего Востока. Владивосток: Дальнаука, 1994. 218 с.
Богатов В.В. Комбинированная концепция функционирования речных экосистем // Вестн. Дальневосточ. отд. РАН. 1995. № 3. С. 51–61.
Богатов В.В. О закономерностях функционирования речных экосистем в свете базовых научных концепций // Вестн. Северо-Восточного центра ДВО РАН. Гидробиология. 2013. № 4. С. 90–99.
Богатов В.В., Никулина Т.В., Вшивкова Т.С. Соотношение биоразнообразия фито- и зообентоса в континууме модельной горной реки Комаровки (Приморский край, Россия) // Экология. 2010. № 2. С. 134–140.
Чертопруд М.В. Разнообразие и классификация реофильных сообществ макробентоса средней полосы европейской России // Журн. общ. биологии. 2011. Т. 72. № 1. С. 51–73.
Bayley P.B. Understanding large river – flood plain ecosystems // BioScience. 1995. V. 45. P. 153–158.
Benda L., Poff L.R., Miller D. et al. Network dynamics hypothesis: How Channel Networks Structure Riverine Habitats // BioScience. 2004. № 54. P. 413–427.
Cummins K.W. Predicting variations in energy flow through a semicontrolled lotic ecosystem // Institute of Water Research, Michigan State University. Technical Report. 1972. № 19. P. 1–21.
Cummins K.W. Macroinvertebrates // River Ecology. Oxford: Blackwell Sci., 1975. P. 170–198.
Cummins K.W., Klug M.J. Feeding ecology of stream invertebrates // Ann. Rev. Ecol. Syst. 1978. № 10. P. 147–172.
Cummins K.W. Riparian – stream linkage paradigm // Verh. Int. Ver. theor. und angew. Limnol. 2002. V. 28. P. 49–58.
Elwood J.W., Newbold J.D., O’Neill R.V., Van Winkle W. Resource spiraling: an operational paradigm for analyzing lotic ecosystems // Dynamics of lotic ecosystems. Ann Arbor: Ann Arbor Sci. Publ. M. I., 1983. P. 3–27.
Frissell C.A., Liss W.J., Warren C.E., Hurley M.D. A hierarchical framework for stream habitat classification: viewing streams in a watershed context // Environ. Management. 1986. V. 10(2). P. 199–214.
Hawkes H.A. River zonation and classification // River Ecology. Oxford: UK. Blackwell Sci. Publ., 1975. P. 312–374.
Hynes H.B.N. The ecology of running waters. Liverpool: University of Liverpool Press., 1970. 555 p.
Illies J., Botosaneanu L. Problemes et methodes de la zonation ecologique des eaux corantes, considerees sur tout du point de vue faunistice // Mitteilungen, Internat. Vereinigung fur Theoretische und Angevandte Limnol. 1963. V. 12. P. 1–57.
Junk W.J., Bayley P.B., Sparks R.E. The flood pulse concept in river– floodplain systems // Canadian Special Publication of Fisheries and Aquatic Sciences. 1989. № 106. P. 110–127.
Maiolini B., Bruno M.C. The River Continuum Concept revisited: lessons from the Alps // The Water Balance of the Alps. Alpine space – man & environment. Innsbruck: University Press., 2007. V. 3. P. 21–30.
Minshall G.W., Peterson R.C., Cummins K.W. et al. Interbiome comparison of stream ecosystems dynamics // Ecology. Monographs. 1983. V. 53. P. 1–25.
Montgomery D.R. Process domains and the river continuum // J. Am. Water Res. Assoc. 1999. V. 35 (2). P. 397–410.
Newblod J.D., Elwood J.W., O’Neill R.V., Winkle W. Nutrient spiraling in streams: The concept and its field measurement // Can. J. Fish. Aquat. Sci. 1981. V. 38. P. 860–863.
Nowicki M. Report of the Phisiography Commission (cited in Nowicki M.) Fishes of raier system of Wisla, Styr, Dniestr and Prut in Galicja. Krakow: Wydz. Krajowy. Poland., 1889. 54 p.
Perry J.A., Schaeffer D.J. The longitudinal distributions of or riverine benthos: a river discontinuum? // Hydrobiologia. 1987. V. 148. P. 257–268.
Petts G.E., Amoros C. Fluvial Hydrosystems. N.Y.: US. Chapman & Hall., 1996. 278 p.
Poff N.L., Allan J.D., Bain M.B. et al. The natural flow regime: a paradigm for river conservation and restoration // BioScience. 1997. V. 47. P. 796–784.
Pool G.C. Fluvial landscape ecology addressing uniqueness within the river discontinuum // Freshwater Biol. 2002. V. 47. P. 641–660.
Ricker W.E. An ecological classification of central Ontario Streams // Univ. Toronto Studies, Biological Series. 1934. V. 37. 114 p.
Sedell J.R., G.H. Reeves F.R., Hauer J.A. et al. Role of refugia in recovery from disturbances: modern fragmented and disconnected river systems // Environ. Management. 1990. № 14. P. 711–724.
Shelford V., Samuel E. Methods for the study of stream communities // Ecology. 1929. № 10. P. 382–391.
Statzner B., Higler B. Questions and comments on the river continuum concept // Can. J. Fish. Aquat. Sci. 1985. V. 42. P. 1038–1044.
Stanford J.A., Ward J.V. An ecosystem perspective of alluvial rivers: connectivity and the hyporheic corridor // J. North Am. Benthol. Soc. 1993. V. 12. P. 48–60.
Thorp J.H., Thorns M.C., Delong M.D. The riverine ecosystem synthesis: Biocomplexity in river networks across space and time // River Res. Appl. 2006. № 22. P. 123–147.
Thorp J.H., Thoms M.C., Delong M.D. The riverine ecosystem synthesis: Towards conceptual cohesiveness in river science. San Diego, CA: Elsevier Acad. Press, 2008. 208 p.
Thienemann A. Der Bergbach des Sauerlandes // Int. Rev. gesamt. Hydrobiol. Hydrogr., Biol. Suppl. 1912. V. 4. 125 p.
Townsend C.R. The patch dynamics concept of stream community ecology // J. North Am. Benthol. Soc. 1989. V. 8. № 1. P. 36–50.
Vannote R.L., Minshall G.W., Cummins K.W. et al. The River Continuum Concept // Can. J. Fish. Aquat. Sci. 1980. V. 37. № 1. P. 370–377.
Ward J.V., Stanford J.A. The serial discontinuity concept of lotic ecosystems // Dynamics of Lotic Systems. California. US: Ann Arbor Sci., 1983. P. 29–42.
Webster J.R. Analysis of potassium and calcium dynamics in stream ecosystems on three southern Appalachian watersheds of contrasting vegetation // Dissertation. University of Georgia, Athens, Georgia, USA. 1975. 238 p.
Webster J.R., Patten B.C. Effect of watershed perturbation on streams potassium and calcium dynamics // Ecol. Monogr. 1979. V. 49. P. 57–72.
Winemiller K.O., Flecker A.S., Hoeinghaus D.J. Patch dynamics and environmental heterogeneity in lotic ecosystems // J. North Am. Benthol. Soc. 2010. № 29. P. 84–99.
Дополнительные материалы отсутствуют.
Инструменты
Биология внутренних вод