

Биология внутренних вод, 2019, № 1, стр. 42-50
Распределение рыб в среднем течении большой равнинной реки под влиянием гидрофизических факторов
Ю. В. Герасимов 1, *, Э. С. Борисенко 2, М. И. Базаров 1, И. А. Столбунов 1, А. И. Цветков 1
1 Институт биологии внутренних вод им. И.Д. Папанина РАН
152742 Ярославская обл., Некоузский р-н, пос. Борок, Россия
2 Институт проблем экологии и эволюции им. А.Н. Северцова РАН
119071 Ленинский просп., д. 33, Москва, Россия
* E-mail: gu@ibiw.yaroslavl.ru
Поступила в редакцию 09.11.2016
После доработки 12.05.2017
Принята к публикации 02.03.2017
Аннотация
Исследовано пространственное распределение рыб на двух участках р. Хопер (каждый длиной 9 км) в Хоперском заповеднике и вне его с использованием эхолота горизонтального зондирования. Изучены плотности рыб на отрезках русла реки с разной степенью меандрирования и глубиной, а также влияние на распределение рыб островов и скоплений древесных остатков. В период высокого уровня воды (конец весны) все рассмотренные факторы оказывали влияние на пространственное распределение рыб. Осенью, при низком уровне воды, разница в плотности скоплений рыб на участках с разной степенью извилистости, с островами и без них уменьшалась, на участках с разной глубиной – возрастала. Плотность рыб на участке реки в пределах заповедника была всегда выше, чем за его пределами.
ВВЕДЕНИЕ
Абиотический фактор водотоков, в основном определяющий распределение рыб в водоеме – стоковое течение со специфическими гидродинамическими характеристиками [10, 13, 14, 16, 19, 22]. Параметры речного потока (скорость, глубина и т.д.) – ключевые факторы, влияющие на распространение или биомассу рыб в пределах сообщества [13, 19–22, 24, 25].
Стремление к минимизации энергетических трат для противодействия сносу вынуждает рыб избирать местообитания с менее выраженным воздействием течения. К ним относятся участки за различными препятствиями (острова, остатки древесной растительности, камни и т.д.) или, наоборот, плесы и ямы, за которыми или в которых образуется гидравлическая тень – область отсутствия или пониженных скоростей течения [5, 6]. Кроме того, пространственная структура распределения рыб в потоке может определяться реоградиентным фактором [7, 8]. Высокая изменчивость гидродинамических показателей потока обусловливает пространственные и временные изменения в распределении речных рыб [17, 18, 23].
Биологическая трактовка причин и закономерностей формирования скоплений рыб в русле реки имеет важное теоретическое и практическое значение. Оценка численности и пространственного распределения рыб в реках – сложнейшая задача, для решения которой в наибольшей степени подходят методы с использованием гидроакустической аппаратуры, но и они имеют определенные ограничения. Для водотоков характерны заросли макрофитов и прибрежных кустарников, обширные отмели и перекаты, резкие перепады глубин, присутствие в толще воды значительных концентраций воздушных пузырьков и различных взвесей, которые затрудняют регистрацию рыб. Часто из-за малых глубин в реках невозможно использовать гидроакустическую аппаратуру вертикальной локации, что обусловливает необходимость применения гидроакустических комплексов горизонтального зондирования водной среды.
Цель работы – оценить численность и распределение рыб на типичном для среднего течения р. Хопер участке с использованием гидроакустической аппаратуры.
МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЯ
Исследования проводили в конце мая и конце сентября 2014 г. в среднем течении р. Хопер в пределах Воронежской обл. Общая длина реки 1008 км, площадь бассейна 61 120 км2. Это самый крупный приток р. Дон. Русло р. Хопер на изученном участке сильно извилистое, шириной 60–100 м. Глубина на перекатах в межень ≤0.3–0.5 м, на некоторых плесах она достигает 5–10 м. Пойма образована мягкими (суглинистыми и супесчаными) грунтами, из-за их подмыва образуются многочисленные отмели. В результате оползневых процессов и сползания многолетних деревьев в реку русло сильно закоряжено. Высокая скорость течения вызывает постоянное движение песчаных грунтов в русле реки, поэтому закономерное чередование перекатов и плесов отсутствует. С этим связано и слабое развитие водной растительности в реке. На исследованном участке в р. Хопер впадают реки Ворона, Карачан и Савала. Негативное влияние на санитарное состояние вод здесь оказывают стоки городов Борисоглебск и Новохоперск [3].
Изучение механизмов формирования структуры речных рыбных сообществ, требует разномасштабного подхода, который отражает иерархию факторов, определяющих распределение гидробионтов в естественных речных системах [10, 14, 16]. Одним из факторов, определяющих выбор участка реки, было наличие на одной из его частей режима особо охраняемой территории (государственный природный Хоперский заповедник).
Участки р. Хопер в заповеднике и за его пределами различались по морфометрии. Участок реки в пределах заповедника был сильно меандрирован и однороден (коэффициент извилистости (к.и.) равен 1.87 по всей длине данного участка). Участок реки вне заповедника был менее извилист (к.и. = 1.36), но неоднороден. Его верхняя часть (5.8 из 9.1 км) имела сходную степень извилистости с участком в заповеднике (к.и. = 1.86). Средняя часть (1.9 км) представляла собой фактически прямой участок (к.и. = 1.02). В нижней части (1.6 км, к.и. = 1.15) находились яма глубиной >5 м и место впадения в р. Хопер ее правого притока – р. Савалы. По глубине и размерам яма отличалась от плесов, которые встречались на исследованном участке реки.
На обоих участках были сходные скорости течения (средняя скорость по руслу 0.28 м/с, на стрежне до 0.8 м/с, ближе к берегу ≤0.15 м/с) и форма донного рельефа (перекаты перемежались с плесами). При этом глубина варьировала от 0.3 до 5.5 м, ширина – от 56 до 120 м. Характер зарастания ‒ вдольбереговой, степень зарастания 1%, грунт – песок.
Учитывая, что русло реки имеет развитое меандрирование, исследовали различия в распределении рыб на поворотах русла и прямых участках. Поскольку основная площадь реки занята перекатами и отмелями, перемежающимися плесами, распределение рыб в зависимости от глубины оценивали по двум категориям участков: мелководные (глубина 1–2 м) и глубоководные (>2 м). На исследуемом участке находятся две группы островов, а также многочисленные вдольбереговые завалы из упавших в реку деревьев, за которыми образуется гидравлическая тень. Плотности скопления рыб в зонах гидродинамического влияния указанных препятствий сравнивали со средней плотностью рыб на остальной акватории участка.
Значимым был и сезонный фактор. Исследования на выделенных участках проводили весной и осенью (при снижении уровня воды в реке на 0.7 м).
Скорость течения по поперечному сечению реки измеряли с помощью доплеровского профилографа Sontek MiniADP, буксируемого на плавучей платформе. Максимальная глубина профилирования 6.2 м, разрешение ячейки профилирования 0.15–0.25 см.
При качественном анализе ихтиофауны использовали сетные орудия лова. Молодь отлавливали мальковой волокушей (длина 9 м, высота 1.5 м, ячея в крыльях и кутке 4 мм), рыб старших возрастных групп – ставными жаберными сетями (ячея 45 мм, 2 × 70 м – 1 шт.; 55 мм, 1.8 × 35 м – 1 шт.) и закидным неводом (длина 24 м, ячея в крыльях 12 мм, ячея в кутке 6 мм). После промеров рыб старших возрастных групп отпускали в реку. Раннюю молодь фиксировали для видовой идентификации в лабораторных условиях.
Гидроакустические исследования проводили на участках в пределах Хоперского заповедника – от с. Варварино (51°11.757′ с.ш., 41°43.116′ в.д.) 9 км вниз по течению (51°10.049′ с.ш., 41°38.473′ в.д.) – и за его пределами – от г. Новохоперска (51°6.175′ с.ш., 41°2.168′ в.д.) 9 км вниз по течению реки – до устья р. Савалы (51°6.782′ с.ш., 41°40.663′ в.д.) при помощи малогабаритного научно-исследовательского комплекса горизонтального зондирования “PanCor” (ООО “Промгидроакустика”, Россия) [2], установленного на надувной лодке. Рабочая частота гидролокатора 455 кГц, направленность одного луча антенны на уровне 6 дБ – 10°, ширина многолучевого сектора в плоскости веера лучей – 60°, максимальная дистанция регистрации одиночной рыбы – 20 м. Гидроакустические съемки вели, двигаясь вниз по течению реки на постоянной скорости (~2–2.5 м/с) моторной лодки с малошумным электромотором, равномерными пилообразными галсами, двигаясь. Акваторию зондировали в горизонтальном режиме, в плоскости, перпендикулярной направлению движения в сторону увеличения глубины [2, 11, 12]. Сетку галсов съемок определяли с учетом характеристик рельефа дна и изгибов русла. Из-за наличия обширных отмелей и сильной закоряженности реки эхосъемки проводили только в светлое время суток. Различия в плотности скоплений рыб анализировали с использованием непараметрического критерия Манна–Уитни (W), поскольку распределение исходных данных в большинстве случаев существенно отличалось от нормального.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
При исследовании выделенного участка реки с использованием сетных орудий лова обнаружено 13 видов рыб. В уловах мальковым неводом на русле зарегистрирована молодь 10 видов рыб. Основу уловов составляли карповые. По численности доминировали лещ Abramis brama (L.) (32%), плотва Rutilus rutilus (L.) (20) и густера Blicca bjoerkna (L.) (14). Уклейка Alburnus alburnus (L.) и елец Leuciscus leuciscus (L.) присутствовали в русле и в затонах (7 и 8% соответственно). В затонах преобладала молодь плотвы (78%). Из других семейств в русле реки присутствовала только молодь окуня Perca fluviatilis L. (7%), в затонах – щуки Esox lucius L. (4%). Из рыб старших возрастов в уловах ставными сетями в русле реки отмеченo шесть видов: лещ (33%), густера (20%), окунь (20%), щука (13%), плотва (7%), в том числе и подуст Chondrostoma nasus (L.) (6%), молодь которого отсутствовала в уловах малькового невода.
Ранее в 2005 г. при обследовании участка р. Хопер в пределах Воронежской обл. в уловах зарегистрировано 29 видов, преобладали уклейка (40.6%), лещ (17.2%) и плотва (14.6%) [3]. Расхождение в количестве обнаруженных в 2014 г. видов с таковым в 2005 г. обусловлено гораздо меньшей протяженностью исследованного участка и ограниченным количеством притонений невода и постановок сетей. Тем не менее, все виды, доля которых в уловах 2005 г. была >2%, отмечены и в 2014 г., отсутствуют только относительно редкие (<0.9%).
Сравнительный анализ распределения рыб в заповеднике и за его пределами
В мае плотность рыб на участке в заповеднике была в восемь раз выше, чем на сходной по морфометрии верхней части участка вне заповедника (0.84 ± 0.2 и 0.12 ± ± 0.04 экз./м2; W = 18150, p < 0.01). При этом на сравниваемых участках оставались сходными коэффициенты вариации плотности (233 и 296% соответственно) и асимметрии (32% на обоих участках), коэффициент эксцесса был выше вне заповедника (102 против 87%). В осенний период средняя плотность на участке вне заповедника была ниже в 48 раз, чем на территории заповедника (0.005 ± 0.002 и 0.24 ± 0.07 экз./м2; W = 10 987, p < 0.01). Относительным сходством на обоих участках отличались коэффициенты вариации плотности (КВП) (305 и 325% соответственно), значения коэффициентов асимметрии (КА) (42 и 32% соответственно) и коэффициент эксцесса (КЭ) (127 и 103% соответственно).
Рис. 1.
Размерный состав (а, в, %) и плотность (б, г, экз./м2) рыб различных размерных групп по результатам гидроакустических съемок на участках р. Хопер в заповеднике (а, б) и вне его (в, г): 1 – 29–30 мая 2014 г., 2 – 22–23 сент. 2014 г.
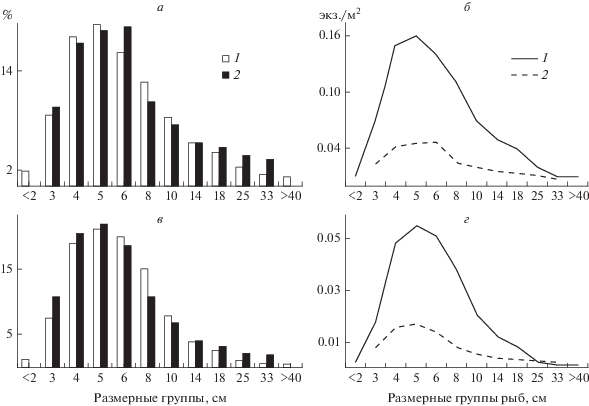
Сравнение плотности рыб в районе упавших в реку деревьев с таковой на прилегающей к нему территории не показало достоверных различий. Единственное обнаруженное отличие в мае – на закоряженных участках (в пределах заповедника) отмечена тенденция к снижению доли рыб с размерами <8 см и повышению таковой с размерами >14 см.
Распределение рыб на участке в пределах Хоперского заповедника
Русло реки в пределах Хоперского заповедника сильно меандрировано, крутые повороты реки чередуются с относительно короткими прямыми участками. Весной средние плотности скоплений на поворотах русла достигали 1.07 ± 0.32 экз./м2, на прямых участках – 0.43 ± ± 0.14 экз./м2 (W = 17 226, p < 0.05). На прямых участках и поворотах были сходными КВП (208 и 216% соответственно), КА (20 и 21% соответственно) и КЭ (47 и 48% соответственно). Зарегистрировано почти двукратное достоверное превышение плотности рыб на глубоководных участках по сравнению с мелководными (1.14 ± 0.38 и 0.62 ± 0.20 экз./м2 соответственно; W = 13 916, p < 0.01), при этом КВП (235 и 216% соответственно), КА (22 и 19%) и КЭ (41 и 43%) были высокими и сходными.
Осенью при снижении уровня воды в реке характер распределения рыб изменился. Плотность скопления рыб значительно уменьшилась, по сравнению с таковой в весенний период (0.84 ± ± 0.20 и 0.24 ± 0.07 экз./м2 соответственно; W = 35 ± ± 251, p < 0.01), увеличились КВП (до 305%), КА (до 42%) и КЭ (до 127%).
Кроме того, возросла разница между средней плотностью скоплений на поворотах русла реки (0.47 ± 0.16 экз./м2) и на прямых участках (0.055 ± ± 0.018 экз./м2) (W = 25516, p<0.01). КВП фактически не изменились (248 и 220% соответственно) на этих участках, но значения КА на прямых участках превысили таковые на поворотах почти в два раза (30 и 18% соответственно), а КА – в три раза (96 и 33% соответственно).
Сохранилось двукратное достоверное превышение плотности рыб на глубоководных участках по сравнению с мелководными (0.33 ± 0.13 и 0.15 ± ± 0.07 экз./м2 соответственно; W = 21 573, p < 0.01). КВП рыб на мелководных участках превысил таковой на глубоководных (361 и 262% соответственно), КА на мелководных участках стал почти в два раза больше, чем на глубоководных (44 и 22% соответственно), а КА – в четыре раза (207 и 49% соответственно).
На одном из прямых участков реки по течению располагались два острова, один за другим на расстоянии 22 м. В мае плотность рыб в проливе между островами (0.61 ± 0.18 экз./м2) и на расстоянии 100 м от второго острова (0.85 ± 0.24 экз./м2) достоверно превышала среднюю плотность рыб на прямых участках реки (W = 15 648, p < 0.05; W = 14 315, p < 0.05 соответственно). Осенью с понижением уровня воды в реке, скопления рыб за островами исчезли, и их плотность уже не отличалась от средней на этом участке (W = 18 126, p = = 0.43).
Распределение рыб на участке за пределами заповедника
Анализ распределения рыб на участке реки вне заповедника проводили отдельно для каждой из трех выделенных частей.
Весной плотность рыб на сильно меандрированной верхней части участка почти не отличалась от таковой на относительно прямом среднем участке (0.11 ± 0.04 и 0.13 ± 0.05 экз./м2; W = 12 297, p < < 0.05 соответственно). КВП был больше в средней части (297 и 182% соответственно), а КА и КЭ – в верхней части (32 и 17%, 101 и 48% соответственно). Плотности рыбы в верхней и нижней частях участка различались в 3 раза (0.11 ± 0.04 и 0.31 ± ± 0.11 экз./м2; W = 14 134, p < 0.01 соответственно). КВП (297 и 164% соответственно), КА (32 и 16%) и КЭ (101 и 47%) были больше в верхней части. Плотности рыбы в средней и нижней частях участка также отличались почти в три раза (0.13 ± ± 0.05 и 0.31 ± 0.11 экз./м2; W = 5 738, p < 0.01 соответственно), но КВП (182 и 164%), КА (17 и 16%) и КЭ (48 и 47%) были почти одинаковыми.
Плотность скоплений осенью значительно уменьшилась по сравнению с таковой в мае. Так, в верхней части участка средняя плотность снизилась более чем в 20 раз (до 0.005 ± 0.002 экз./м2; W = 1664, p < 0.01), при этом КВП увеличился (до 325%), а КА и КЭ остались на прежнем уровне (32 и 103% соответственно). В средней части участка плотность уменьшилась более чем в 40 раз (до 0.003 ± 0.001 экз./м2; W = 54, p < 0.01), а КВП (152%), а КА (15%) и КЭ (45%) остались почти одинаковыми. Только в нижней части участка снижение плотности было недостоверным (до 0.26 ± 0.10 экз./м2; W = 2 610, p < 0.21), но КВП увеличился (203%), а КА (13%) и КЭ (29%) уменьшились. Плотности рыб на верхнем и среднем участках осенью также достоверно не различались между собой (0.005 ± 0.002 и 0.003 ± 0.001 экз./м2; W = 7647, p < 0.01 соответственно), а в нижней части участка она увеличилась в ~50 раз (0.26 ± 0.10 экз./м2; W = 5069, p < 0.01).
Весной на поворотах плотность рыб была значительно выше (0.35 ± 0.22 экз./м2), чем на прямых участках (0.09 ± 0.02 экз./м2) (W = 6858, p < 0.05). По сравнению с прямыми участками на поворотах были выше КВП (136 и 455% соответственно), КА (58 и 16%) и КЭ (318 и 30%). Осенью достоверных различий между плотностью рыб на поворотах русла (0.07 ± 0.04 экз./м2) и прямых участках (0.06 ± 0.04 экз./м2) не наблюдалось (W = 12 013, p = 0.13). КВП стали сходными (437 и 399% соответственно), уменьшились и различия между КА (39 и 29% соответственно) и КЭ (161 и 101% соответственно).
Весной плотность рыб на глубоководных участках была достоверно выше, чем на мелководных (0.18 ± 0.05 и 0.07 ± 0.02 экз./м2; W = 2042, p < 0.01 соответственно). При этом значения КВП (222 и 178% соответственно), КА (29 и 26%) и КЭ (95 и 100%) были сходными. Достоверные различия плотности рыб на глубоководных участках и в яме, расположенной при впадении р. Савалы, отсутствовали (0.18 ± 0.05 и 0.17 ± 0.06 экз./м2; W = = 12 045, p < 0.11 соответственно). Однако КВП (222 и 102% соответственно), КА (29 и 5%) и КЭ (95 и 8%) на глубоководных участках значительно превышали таковые на мелководных. На мелководных участках плотность рыб была достоверно ниже, чем в яме (0.07 ± 0.02 и 0.17 ± 0.06 экз./м2; W = 2945, p < 0.01 соответственно), но КВП (178 и 102%), КА (26 и 5%) и КЭ (100 и 8%) были значительно выше.
Осенью после понижения уровня в реке, плотности рыб на глубоководных (0.03 ± 0.02 экз./м2) и мелководных участках (0.01 ± 0.005 экз./м2) достоверно не различались (W = 10 002, p = 0.59). При этом плотность рыб в яме достоверно превышала таковую на глубоководных участках (0.37 ± ± 0.19 экз./м2; W = 6288, p = 0.01). В ряду мелководные, глубоководные участки и яма зарегистрировано снижение КВП (443, 345 и 176% соответственно), КА (40, 22 и 8%) и КЭ (188, 55 и 13%). На средней части участка располагались три острова, за которыми весной образовались скопления рыб со значительно более высокой средней плотностью (1.19 ± 0.15 экз./м2) и со сравнительно низким КВП (96.6%), чем на всей остальной акватории р. Хопер. Осенью скопления за островами исчезли, и плотность рыб за ними уже не отличалась от средней на этом участке.
Размерный состав рыб
По результатам гидроакустической съемки выявлен размерный состав рыб на исследованных участках (рис. 1а и 1в). В заповеднике и за его пределами соотношение размерных групп было сходным: доминировали группы 4–6 см, следующие по численности группы были особи длиной 3, 8 и 10 см (весной – ранняя молодь и годовики, осенью – сеголетки и двухлетки). Сезонные изменения на обоих участках заключались в отсутствии осенью особей из размерных групп <2 и >40 см (рис. 1). В размерную группу <2 см входила ранняя молодь от нереста весенне-нерестующих рыб в данном году, к осени подросшая молодь перешла в более крупную размерную группу. Размерная группа >40 см, по-видимому, была представлена производителями, зашедшими в р. Хопер для нереста и скатившимися после его окончания.
Плотность рыб всех выделенных размерных групп на участке в пределах заповедника (рис. 1б) значительно превосходила плотность таковых на участке вне заповедника (рис. 1г). Осенью снижение плотности наблюдалось на обоих участках и для всех размерных групп: у особей размером 4–10 см в среднем в 4 раза, >14 см – в 2 раза.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Сравнение численности рыб показало, что при приблизительно сходной площади водной поверхности участков общая численность рыб весной и осенью в Хоперском заповеднике была значительно выше таковой за его пределами (табл. 1). Подобное соотношение сохранилось и осенью после снижения плотности рыб всех размерных групп на обоих участках, обусловленного скатом части молоди и производителей после окончания нерестового периода. В отличие от показателей плотности характер распределения рыб на обоих участках был сходным, на что указывают относительно небольшие различия между участками в весенних и осенних значениях КВП, КА и КЭ.
Таблица 1.
Площадь (м2), средняя плотность (экз./м2), численность (экз.) рыб на исследованных участках р. Хопер в 2014 г.
| Показатель | Участок реки | |
|---|---|---|
| в заповеднике | вне заповедника | |
| Площадь | $\frac{{1{\kern 1pt} {\kern 1pt} 179{\kern 1pt} {\kern 1pt} 084}}{{1{\kern 1pt} {\kern 1pt} 179{\kern 1pt} {\kern 1pt} 000}}$ | $\frac{{1{\kern 1pt} {\kern 1pt} 470{\kern 1pt} {\kern 1pt} 900}}{{1{\kern 1pt} {\kern 1pt} 470{\kern 1pt} {\kern 1pt} 911}}$ |
| Средняя плотность | $\frac{{0.84}}{{0.24}}$ | $\frac{{0.12}}{{0.005}}$ |
| Численность | $\frac{{990{\kern 1pt} {\kern 1pt} 430}}{{282{\kern 1pt} {\kern 1pt} 960}}$ | $\frac{{176{\kern 1pt} {\kern 1pt} 509}}{{7355}}$ |
Более высокие плотности рыб в пределах заповедника объясняются более ограниченным негативным антропогенным воздействием на рыбное население реки – в первую очередь отсутствием легальной и нелегальной добычи рыбы. Возможно, что определенное влияние оказывает и санитарное состояние вод. Крупный населенный пункт, стоки которого могли бы влиять на качество среды на участке в пределах заповедника (г. Борисоглебск), расположен в 51 км выше по течению. Участок же, находящийся вне заповедника, начинается непосредственно в черте г. Новохоперска.
Сезонные изменения на участке в пределах заповедника сопровождались более чем трехкратным снижением плотности рыб. При этом увеличился КВП и значения КА и КЭ, что свидетельствует о возрастании доли скоплений рыб с низкой плотностью. На участке вне заповедника осеннее снижение плотности было более значительным, но не сопровождалось изменениями в распределении рыб. Исключением стала его нижняя часть с ямой, в которой на фоне обмеления реки не было отмечено достоверного снижения плотности, а уменьшение значений КА и КЭ указывало на увеличение плотности отдельных скоплений рыб в яме.
Положительную связь плотности рыб с глубиной речного русла подтверждало и сравнение распределения рыб на перекатах и плесах в пределах заповедника, где весной и осенью плотность рыб на глубоководных участках была больше, чем на мелких. Но если весной КВП, КА и КЭ на мелководных и глубоководных участках были сходными, то осенью после понижения уровня воды в реке, они значительно повысились на мелководье, что указывает на возрастание варьирования плотности и увеличение доминирования скоплений с низкой плотностью.
На участке вне заповедника плотность рыб на плесах и в яме в весенний период была сходной, но достоверно отличалась от плотности на мелководных участках. При этом КВП, КА и КЭ на глубоководных и мелководных участках были фактически равными, но значительно превышали соответствующие показатели в яме. Осенью достоверной разницы между плотностью рыб на глубоководных и мелководных участках не наблюдалось. Плотность рыб в яме вдвое увеличилась относительно весенней и достоверно отличалась от плотности рыб на других участках реки. КВП, КА и КЭ на глубоководных и мелководных участках значительно увеличились, а в яме остались на весеннем уровне.
Следовательно, осенью, в связи со значительным снижением уровня воды в реке, рыбы концентрировались в основном на глубоких участках русла, создавая относительно плотные скопления с более низким КВП и низкими положительными значениями КА и КЭ. На обширных мелководьях скопления рыб характеризовались большим коэффициентом вариации. Высокие и положительные значения КА и КЭ указывали на преобладание здесь скоплений рыб с низкой плотностью.
Более глубокие участки реки в условиях значительных колебаний уровня воды представляют собой местообитания с более стабильными условиями окружающей среды, чем мелкие [4, 19]. Образующаяся здесь гидравлическая тень (низкая скорость течения и отсутствие реоградиента) способствует минимизации энергетических трат на сопротивление течению [5, 6]. На прямых участках русла с переменной глубиной высокие скорости наблюдаются только на перекате, а в углублениях дна до и после переката скорости течения в 2–3 раза меньше (рис. 2в). Кроме того, в большинстве рек с быстрым течением планктон почти отсутствует и основным источником пищи для многих рыб служат донные беспозвоночные. При этом большое значение имеет дрифт бентосных организмов, которые перемещаются речным потоком вниз по течению [1]. При отборе проб дрифта на плесах среди дрейфующих организмов отмечена значительная доля беспозвоночных, обычно обитающих на перекатах [1], т.е. плесы играют роль ловушек для организмов, сносимых с участков с быстрым течением. Минимизации энергетических трат на сопротивление течению и более высокое обилие кормовых организмов способствуют образованию скоплений рыб на плесах.
Рис. 2.
Распределение скоростей (cм/c) течения по поперечному сечению реки р. Хопер на разрезах по данным профилографа: а – на повороте русла, б – через плес на прямом участке, в – через перекат и плес, г – перекат в условиях низкого уровня. По оси ординат – глубина (м).
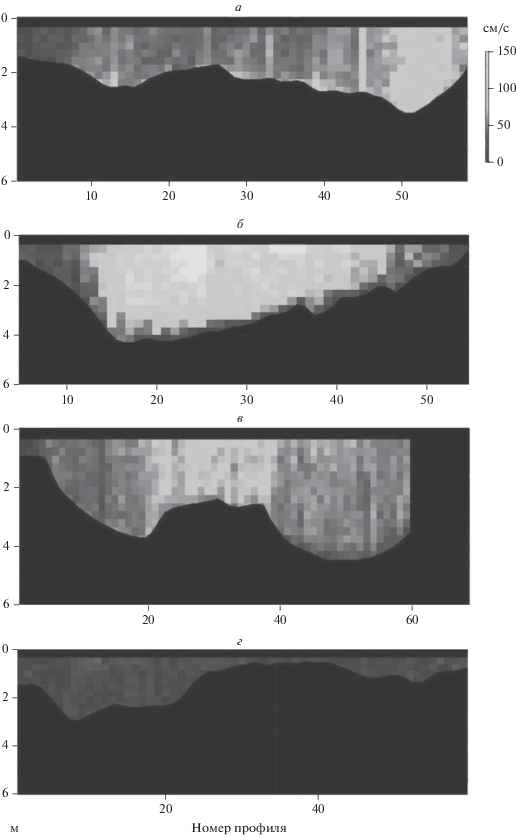
Пространственная структура распределения рыб в потоке определяется реоградиентным фактором. Рыбы в основном приурочены к участкам пониженных скоростей на периферии оси стокового течения (<15 см/с для рео-лимнофильных рыб) [8]. Высокие градиенты скорости (зоны сгущения изотах) способствуют сжатию скоплений, а размытые градиенты скорости (зоны разрежения изотах) – диффузии скоплений [8]. Особенно выражены реоградиентные условия на поворотах русла, что обусловлено смещением области максимальных скоростей течения воды относительно геометрической оси русла реки [7] в сторону внешнего берега излучины (см. рис. 2а).
Весной в пределах заповедника и вне его средние плотности скоплений на поворотах русла достоверно превышали таковые на прямых участках. КВП, КА и КЭ были близкими по значению. Вне заповедника эти показатели существенно превышали таковые на поворотах, где чаще встречались скопления с высокой плотностью. На поворотах образующийся реоградиент (рис. 2а) способствовал распределению рыб в соответствии с избираемыми ими скоростями течения, это приводило к уплотнению скоплений, как и наиболее интенсивный дрифт на участках рек, где скорость потока выше [1, 4]. Следовательно, рыбы должны “оптимизировать” свое положение в потоке в соответствии со скоростью потока и обилием дрифта. На обширных песчаных перекатах и плесах, занимающих на прямых участках всю ширину русла (рис. 2б и 2г) происходит размытие реоградиента, т.е. скорости течения по всей ширине русла одинаковые. Это приводит к диффузии скоплений и, соответственно, к снижению диапазона колебаний их плотности.
Осенью в пределах заповедника на фоне снижения общей плотности рыбных скоплений увеличилась разница между средней плотностью скоплений на поворотах и на прямых участках. Вне заповедника, где общее снижение плотности было более значительным, достоверная разница между плотностями отсутствовала. При этом КВП, КА и КЭ на прямых участках приблизились к значениям этих показателей на поворотах русла или превысили их. Выявленные изменения обусловлены снижением уровня реки из-за уменьшения стока, что привело к снижению скорости потока и сглаживанию реоградиента на поворотах русла (рис. 2г) и, как следствие, к диффузии скоплений.
Наличие в русле реки островов также способствовало образованию реоградиента от нулевых скоростей в их гидравлической тени и до максимума в обтекающем потоке, что привело к перераспределению скоплений рыб в потоке в соответствии с избираемыми скоростями течения и плотностью организмов дрифта. Весной при более высоких уровне и скоростях течения плотность скоплений рыб за островами была достоверно выше и со сравнительно низким КВП (96.6%), чем на остальной акватории реки. Осенью со снижением уровня и скорости течения скопления рыб за островами исчезли – плотность рыб за ними не отличалась от средней плотности на остальной прилегающей акватории, что было обусловлено диффузией скоплений в результате размытия реоградиента.
Упавшие в реку деревья в условиях быстрого течения также создавали гидравлическую тень, однако, достоверной разницы в плотности рыб в районе упавших в реку деревьев с таковой на прилегающей акватории не обнаружено. Поскольку наряду с абиотическим факторами, биотические взаимодействия также могут оказывать заметное влияние на структуру речных рыбных сообществ [15], причина может заключаться в отпугивающем влиянии хищников. Так, хищники-засадчики [9] щука и окунь (13.3 и 20% численности рыб в сетных уловах в русле р. Хопер соответственно) в условиях крайне низкой степени зарастания русла вынуждены использовать в качестве укрытий стволы упавших в воду деревьев. Это отпугивает многочисленную молодь рыб от таких биотопов, препятствуя образованию здесь плотных скоплений. Подтверждением может служить то, что доля рыб с размерами <8 см на закоряженных участках ниже, чем в среднем по реке, а с размером >14 см – выше, т.е. эти участки выбирают хищники и крупные мирные рыбы из-за своего размера недоступные хищникам, численность которых в реке меньше, чем рыб с размерами <8 см.
Выводы. Плотность рыб на участке реки в пределах заповедника была всегда выше, чем за его пределами, что объясняется отсутствием легальной и менее интенсивной нелегальной добычи рыбы. Все исследованные гидрофизические факторы оказывали влияние на пространственное распределение рыб. На всем протяжении исследованного участка реки средние плотности скоплений на поворотах русла и на участках с островами были достоверно выше, чем на прямых участках и участках без островов, что определялось особенностями реоградиентных условий. Осенние изменения в распределении рыб сопровождались более чем трехкратным снижением их плотности, связанным со скатом рыб. При осеннем снижении уровня воды разница в плотности скоплений рыб на участках русла с разной степенью меандрирования увеличилась, но на большинстве прямых участков скопления рыб исчезли полностью. Осенью со снижением уровня и скорости течения не стало плотных скоплений рыб за островами – в этот период плотность не отличалась от средней на остальной прилегающей акватории, что было обусловлено диффузией скоплений в результате размытия реоградиента. Возросла разница в плотности на участках с разной глубиной. Положительная связь плотности рыб с глубиной речного русла объясняется тем, что более глубокие участки реки в условиях значительных колебаний уровня воды представляют собой местообитания с более стабильными и благоприятными условиями окружающей среды, чем мелкие места.
Работа выполнена в рамках темы государственного задания № г/р АААА-А18-118012690102-9.
Список литературы
Богатов В.В. Значение бентостока в процессах биологического продуцирования в реках // Экология. 1984. № 3. С. 51–60.
Борисенко Э.С., Мочек А.Д., Павлов Д.С., Чемагин А.А. Распределение рыб в речной системе Нижнего Иртыша // Вопр. ихтиологии. 2013. Т. 53. № 1. С. 31–43.
Гладких К.К., Делицына Л.Ф., Карпов Н.А. К познанию ихтиофауны бассейна реки Хопер // Тр. Хоперск. гос. заповедника. Воронеж: Изд-во Воронеж. гос. ун-та, 2012. Вып. 7. С. 3–19.
Ефремов Д.А. Распределение и поведение реофильных видов рыб в реках Восточной Фенноскандии: Автореф. дис. … канд. биол. наук. Петрозаводск, 2012. 24 с.
Мартинсен Ю.В. Влияние течений на поведение рыб // Рыб. хоз-во. 1940. № 12. С. 23–27.
Мартинсен Ю.В. Движение рыб в реке // Рыб. хоз-во. 1937. № 2. С. 27–30.
Павлов Д.С., Барекян А.Ш., Рипинский И.И. и др. Экологический способ защиты рыб на повороте струей открытого потока. М.: Наука, 1982. 112 с.
Поддубный С.А., Герасимов Ю.В., Новиков Д.А. Структура течений и распределение рыб в речных плесах верхневолжских водохранилищ // Биология внутр. вод. 2003. № 1. С. 89– 97.
Фортунатова К.Р., Попова О.А. Питание и пищевые взаимоотношения хищных рыб в дельте Волги. М.: Наука, 1973. 299 с.
Biggs B.J.F., Nikora V.I., Snelder T.H. Linking scales of variability to lotic ecosystem structure and function // River Res. Appl. 2005. V. 21. P. 283–298.
Borisenko E.S., Gusar A.G., Goncharov S.M. The target strength dependence of some freshwater species on their length-weiht characteristics // Proceedings of the Institute of Acoustics. England: Lowestoft, 1989. V. 11. Pt. 3. P. 27–34.
Borisenko E.S., Degtev A.I., Mochek A.D., Pavlov D.S. Hydroacoustic characteristics of mass fishes of Ob-Irtish basin // J. Ichthyol. 2006. V. 46. Suppl. 2. P. 227–234.
Bunn S.E., Arthington A.H. Basic principles and ecological consequences of altered flow regimes for aquatic biodiversity // Environ. Manag. 2002. V. 30. P. 492–507.
Fausch K.D., Torgersen C.E., Baxter C.V., Li H.W. Landscapes to river scapes: bridging the gap between research and conservation of stream fishes // BioScience. 2002. V. 52. P. 483–498.
Jackson D.A., Peres-Neto P.R., Olden J.D. What controls who is where in freshwater fish communities: the roles of biotic, abiotic and spatial factors? // Can. J. Fish. Aquat. Sci. 2001. V. 58. P. 157–170.
Lowe W.H., Likens G.E., Power M.E. Linking scales in stream ecology // BioScience. 2006. V. 56. P. 591–597.
Hynes H.B.N. The ecology of running waters. Liverpool, England: Liverpool University Press, 1970. 555 p.
Kennard M.J., Pusey B.J., Arthington A.H. et al. Utility of a multivariate modelling method for prediction of freshwater fish assemblages and evaluation of river health // Hydrobiologia. 2006. V. 572. P. 33–57.
Kennard M.J., Olden J.D., Arthington A.H. et al. Multiscale effects of flow regime and habitat and their interaction on fish assemblage structure in eastern Australia // Can. J. Fish. Aquat. Sci. 2007. V. 64. P. 1346–1359.
Milner N.J., Wyatt R.J., Scott M.D. Variability in the distribution and abundance of stream salmonids, and the associated use of habitat models // J. Fish Biol. 1993. V. 43. Suppl. A. P. 103–119.
Poff N.L., Allan J.D. Functional organization of stream fish assemblages in relation to hydrologic variability // Ecology. 1995. V. 76. P. 606–627.
Poff N.L., Allan J.D., Bain M.B. et al. The natural flow regime: a paradigm for riverine conservation and restoration // BioScience. 1997. V. 47. P. 769–784.
Pusey B.J., Kennard M.J., Arthington A.H. Discharge variability and the development of predictive models relating stream fish assemblage structure to habitat in north-eastern Australia // Ecol. Freshwater Fish. 2000. V. 9. P. 30–50.
Rahel F.J. The hierarchical nature of community persistence: a problem of scale // Amer. Natur. 1990. V. 136. P. 328–344.
Strange E.M., Moyle P.B., Foin T.C. Interactions between stochastic and deterministic processes in stream fish community assembly // Environ. Biol. Fishes. 1992. V. 36. P. 1–15.
Дополнительные материалы отсутствуют.
Инструменты
Биология внутренних вод