

Биология внутренних вод, 2019, № 1, стр. 79-85
Физиолого-биохимические показатели сеголетков аборигенного (Carassius auratus gibelio (Bloch, 1792)) и инвазионного (Perccottus glenii (Dybowski, 1877)) видов рыб в пойменном оз. Круглое (Саратовское водохранилище)
В. В. Халько 1, Е. В. Шемонаев 2, 3, *, Е. В. Кириленко 2, Н. А. Халько 1
1 Институт биологии внутренних вод им. И.Д. Папанина РАН
152742 Ярославская обл., Некоузский р-н, пос. Борок,, Россия
2 Институт экологии Волжского бассейна РАН
455003 Тольятти, ул. Комзина, д. 10, Россия
3 Тольяттинский государственный университет
445020 Тольятти, ул. Белорусская, 14, Россия
* E-mail: fosfolipid00@mail.ru
Поступила в редакцию 02.08.2017
После доработки 16.02.2018
Принята к публикации 26.01.2018
Аннотация
На примере оз. Круглое (Мордовинская пойма Саратовского водохранилища) приведен сравнительный анализ изменений физиолого-биохимических показателей сосуществующих в пределах общих биотопов сеголетков аборигенного (серебряный карась Carassius auratus gibelio) и инвазионного (ротан-головешка Perccottus glenii) видов. Выявлены межвидовые различия динамики массы тела, содержания белка, жира, воды и калорийности при увеличении длины особей в различные месяцы нагульного периода. Рассмотрены возможные причины и последствия их возникновения.
ВВЕДЕНИЕ
Озеро Круглое – водоем в Мордовинской пойме Саратовского водохранилища с площадью водной поверхности 0.005 км2, в ихтиофауне которого, как и в подобных ему озерах поймы, по численности доминируют два вида рыб, имеющих общий биотоп обитания: аборигенный – серебряный карась (Carassius auratus gibelio) (Bloch, 1792) и амурский вселенец – ротан-головешка (Perccottus glenii) (Dybowski, 1877). Впервые последний обнаружен в водоемах Мордовинской поймы Саратовского водохранилища в 1998 г. [1]. Он быстро заселил все пойменные водоемы и стал видом-доминантом [2]. Озеро Круглое представляет собой непроточный замкнутый водоем рельефного происхождения с илистым дном, его глубина ≤4 м, средняя – ≤2 м. По физико-химическому состоянию озеро характеризуется средней щелочной средой pH 8.57–9.15 [3]. В последней декаде июня в нем интенсивно развиваются макрофиты: телорез (Stratiotes aloides), элодея канадская (Elodea canadensis), роголистник темно-зеленый (Ceratophyllum demersum), многокоренник обыкновенный (Spirodela polyrrhiza), ряска маленькая (Lemna minor), с конца мая – рогоз (Typha angustifolia). В период половодья (последняя декада апреля – середина июня) озеро сообщается с Саратовским водохранилищем. С мая по сентябрь уровень воды в озере, как правило, снижается, и связь с водохранилищем прерывается [3]. Судьба двух сосуществующих в однотипных биотопах видов рыб – ротана-головешки и серебряного карася в водоемах Мордовинской поймы во многом зависит от физиолого-биохимического состояния разноразмерных ровесников в пополнении их популяций [6, 7]. Последнее обусловлено внутригенерационной разнокачественностью потомства и уровнем обеспеченности пищей быстро- и медленнорастущих особей генерации. Какие-либо сведения о физиолого-биохимическом состоянии сеголетков ротана-головешки и серебряного карася в водоемах Мордовинской поймы в литературе отсутствуют.
Цель работы – выявление межвидовых особенностей динамики физиолого-биохимических показателей сеголетков разноразмерных групп серебряного карася и ротана-головешки в различные месяцы нагульного периода в водоемах Мордовинской поймы Саратовского водохранилища.
МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЯ
Синхронный отлов сеголетков серебряного карася и ротана в оз. Круглое проводили с июля по октябрь 2013 г. с интервалом в 15 cут. Рыб отлавливали сачком и мальковой волокушей в прибрежной зоне озера. Из каждого улова отбирали мальков обоих видов рыб всех встречавшихся размеров. Длину тела рыб (от конца рыла до конца чешуйного покрова) измеряли с точностью до 1 мм, массу тела рыб – до 0.002 г. Всех пойманных рыб сгруппировали в размерные группы с интервалом в 5 мм – 10–14, 15–19, 20–24, 25–29, 30–34 и 35–39 мм. Пробы сеголетков каждой размерной группы были суммарными и включали целых особей. Количество одноразмерных рыб в каждую пробу подбирали, исходя из их средней массы, стараясь набрать суммарную навеску каждой пробы мальков ≥0.8 г, достаточную для анализа липидного состава рыб. По каждой размерной группе сравниваемых видов рыб отобрано по 3–25 проб, которые фиксировали этиловым спиртом после предварительного измельчения ножницами и хранили при температуре –10°С до начала биохимических анализов. Общее содержание липидов определяли по методу Фолча с соавт. [11] в модификации В.И. Лапина и Е.Г. Черновой [5]. Об общем содержании белка судили по количеству сухого обезжиренного остатка в теле рыб на >90% состоящего из белка [7–9]. Для этого обезжиренный и обезжиренный безбелковый (содержимое водно-метанольной фазы) остаток высушивали до постоянной массы при температуре 104°С и по сумме этих величин определяли количество сухого обезжиренного остатка. Содержание влаги оценивали по разности между сырой (начальной) и сухой (конечной) массами, полученной суммированием величин сухого обезжиренного остатка, липидов и экстрактивных веществ в водно-метанольной фазе после их высушивания до постоянной массы. Калорийность (К) рыб (кал/г сырого вещества) рассчитывали по их биохимическому составу, используя формулу:
где Б и Ж – соответственно содержание белка и липидов в сыром веществе, % [6].За весь период наблюдений определены длина и масса тела, содержание белка, липидов, воды и калорийность особей в 87 пробах сеголетков карася и в 190 пробах сеголетков ротана. Достоверность межвидовых различий средних значений проанализированных ростовых и биохимических параметров молоди рыб определяли по критерию Стьюдента при р = 0.05. Статистический анализ результатов исследований проводили по работе [4].
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
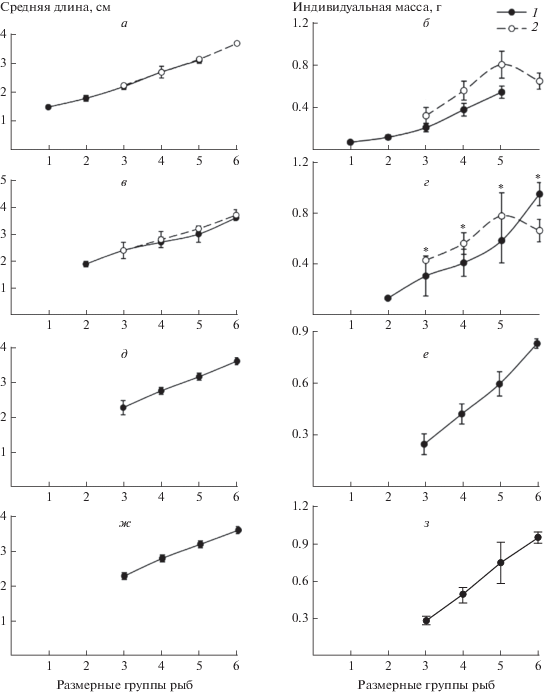
Проведенные исследования показали, что средняя длина тела у сеголетков ротана и серебряного карася одноразмерных групп была фактически одинаковой (p > 0.05) и возрастала на протяжении нагульного периода у обоих видов рыб. При межвидовом сравнении динамики средней массы тела у одноразмерных особей, таковая у сеголетков ротана устойчиво возрастала с увеличением длины рыб, что отчетливо проявлялось ежемесячно с июля по октябрь. У молоди же серебряного карася подобная закономерность сохранялась в июле–августе только у рыб размером до 30–34 мм. У более крупных особей (35–39 мм) средняя масса тела существенно (р < 0.05) снижалась в оба месяца наблюдений и в августе оказывалась достоверно (р < 0.05) меньше, чем у ротана. При отсутствии существенных различий (р > 0.05) средней длины тела между молодью карася и ротана одинаковых размерных групп – 20–24, 25–29 и 30–34 мм в июле–августе сеголетки серебряного карася по массе тела значительно (р < 0.05) превосходили своих одноразмерных ровесников ротана (рис. 1).
Рис. 1.
Средняя длина (a, в, д, ж) и индивидуальная масса (б, г, е, з) сеголетков ротана (1) и серебряного карася (2) различных размерных групп в июле (а, б), августе (в, г), сентябре (д, е) и октябре (ж, з) 2013 г. в оз. Круглое. Размерные группы рыб: 1 – 10–14 мм; 2 – 15–19 мм; 3 – 20–24 мм; 4 – 25–29 мм; 5 – 30–34 мм; 6 – 35–39 мм. Межвидовые различия достоверны при р = 0.05; приведены значения средней арифметической и ее среднеквадратических отклонений.
Межвидовые различия средней массы тела одноразмерных особей и характера ее ростовой динамики отразились и на физиолого-биохимических показателях сравниваемых видов рыб. В качестве общей закономерности для молоди серебряного карася и ротана можно отметить ежемесячно отмечавшееся на протяжении всего периода наблюдений возрастание содержания белка в их организме при увеличении длины тела. При этом межвидовые различия в содержании белка у одноразмерных особей оказались наиболее выраженными (р < 0.05) в июле, когда сеголетки ротана достоверно превосходили своих ровесников серебряного карася по величине этого биохимического показателя (рис. 2). Однофакторный дисперсионный анализ показал, что жирность сеголетков ротана (р > 0.05) незначительно изменялась при увеличении длины тела в каждый месяц наблюдений и в большинстве случаев не отличалась (р > > 0.05) от жирности одноразмерных особей серебряного карася. Лишь в августе содержание общих липидов в теле у ротана размерной группы 35–39 мм было достоверно (р < 0.05) выше, чем у молоди серебряного карася (рис. 2). Иной характер изменений жирности тела в зависимости от размера особи наблюдался у сеголетков серебряного карася, у которых величина этого биохимического показателя в июле достоверно (р < 0.05) снижалась при увеличении длины тела рыб и оставалась фактически неизменной (р > 0.05) – в августе (рис. 2). Содержание воды в организме сеголетков сравниваемых видов рыб изменялось противонаправленно изменению их жирности; достоверные различия в величине этого показателя между одноразмерными особями карася и ротана проявлялись только в июле (рис. 3).
Рис. 2.
Содержание белка (а, в, д, ж) и жира (б, г, е, з) у сеголетков ротана и серебряного карася различных размерных групп в июле (а, б), августе (в, г), сентябре (д, е) и октябре (ж, з) 2013 г. в оз. Круглое. Остальные обозначения, как на рис. 1.
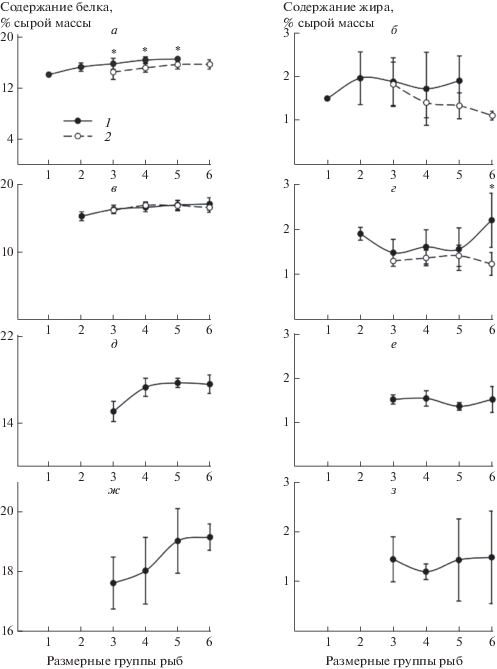
Рис. 3.
Содержание воды (а, в, д. ж) в теле и калорийность (б, г, е, з) сеголетков ротана и серебряного карася различных размерных групп в июле (а, б), августе (в, г), сентябре (д, е) и октябре (ж, з) 2013 г. в оз. Круглое. Остальные обозначения, как на рис. 1.
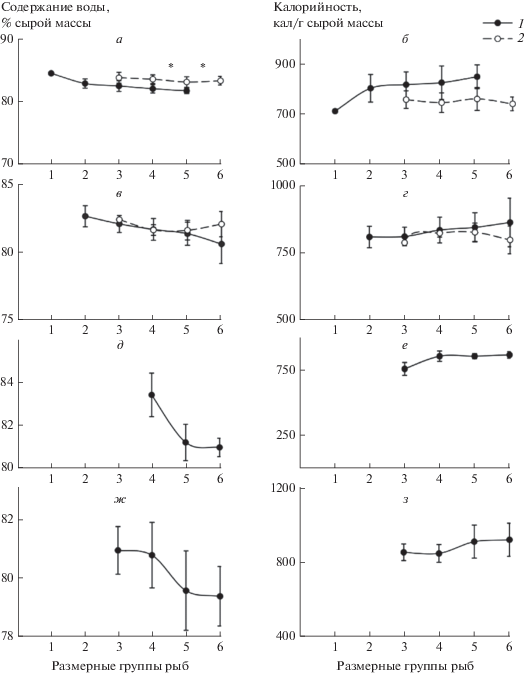
Межвидовые различия были выявлены и в калорийности одноразмерных особей. Калорийность сеголетков ротана возрастала при увеличении длины тела рыб ежемесячно с июля по октябрь. Калорийность же молоди карася в летние месяцы снижалась по мере увеличения длины тела особей, и в июле была существенно ниже, чем у одноразмерных ровесников ротана (рис. 3).
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
У молоди многих видов рыб разных экологических групп при отсутствии энергетических затрат на генеративный обмен основная часть энергии ассимилированной пищи расходуется на энергетический и пластический обмен [6, 7, 10]. В большинстве случаев это выражается в интенсивном увеличении линейных и массовых характеристик тела, сопровождающихся возрастанием содержания белка, снижением оводненности и повышением калорийности тела особи. Такая закономерность наиболее отчетливо проявляется у бореальных видов рыб в сезонном плане, когда устойчивое осеннее понижение температуры воды служит сигнальным фактором для интенсификации процесса жиронакопления, замедления процесса белкового роста, уменьшения оводненности организма и увеличения его калорийности. Одно из проявлений внутригенерационной разнокачественности потомства рыб – наличие среди ровесников особей с разным темпом роста. В результате этого в каждой генерации рыб присутствуют разноразмерные особи, что служит одним из адаптивных свойств вида, обеспечивающих его выживание в разных трофических ситуациях. Уровень обеспеченности пищей рыб зависит от ее экологической, физиологической и морфологической доступности [6]. В реальной экологической ситуации трофические условия для разноразмерных особей генерации обычно неодинаковы. Как правило, с увеличением размеров тела сеголетки переходят на более крупные, энергетически ”выигрышные” объекты питания [10]. Однако в зависимости от экологической обстановки, преимущество в питании, как правило, получают не все особи размерного ряда ровесников, а лишь те, для которых уровень обеспеченности пищей достаточен для реализации генетически заложенной интенсивности протекания обменных процессов в организме. В этом, по-видимому, и заключается один из механизмов естественного отбора в пополнении популяций рыб. В случаях, когда обеспеченность пищей (ее обилие, экологическая, морфологическая и физиологическая доступность) достаточна для молоди всех размерных групп, содержание белка и жира в теле рыб обычно возрастает, а содержание воды – снижается при увеличении длины тела особей. Одновременно с этим повышается и калорийность рыб. Отклонения от этой закономерности, как правило, свидетельствуют о неблагополучии условий нагула для особей отдельных размерных групп – возможных “кандидатов” на естественный отход при необратимости сложившейся трофической ситуации.
Подобная картина выявлена у сеголетков серебряного карася в июле–августе, когда с увеличением длины тела снижалась масса, жирность и калорийность его особей. Уменьшение жирности сеголетков серебряного карася по мере увеличения их размеров сопровождалось повышением содержания воды в организме (наиболее выражено в августе). Из этого следует, что уровень обеспеченности пищей сеголетков серебряного карася в июле–августе, вероятно, снижался по мере увеличения длины их тела. Условия же нагула для одноразмерных с карасем сеголетков ротана были лучше, так как показатели массового роста, содержание жира и калорийность у них возрастали, а содержание воды снижалось с увеличением длины тела сеголетков.
Принимая во внимание, что сеголетки серебряного карася и ротана могут служить пищей для хищных видов рыб, наиболее “выигрышная” в энергетическом плане для последних (при прочих равных условиях) молодь ротана.
Отмеченные авторами особенности изменений ростовых и биохимических показателей у сеголетков серебряного карася позволяют предположить, что условия их нагула в оз. Круглое и в подобных ему водоемах Мордовинской поймы Саратовского водохранилища в летние месяцы хуже, чем для сеголетков инвазионного вида – ротана. При этом ухудшение условий нагула для молоди серебряного карася возрастает по мере увеличения размеров сеголетков. Это позволяет предположить, что к концу нагульного периода в пополнении популяции карася останутся мелкие, наименее подготовленные к зимовке особи – первоочередные “кандидаты” на естественный отход в подледный период. Соответственно, в ближайшие годы можно ожидать снижение численности пополнения популяции серебряного карася в водоемах Мордовинской поймы Саратовского водохранилища, подобных оз. Круглое.
Выводы. Сравнительный анализ изменения ростовых и физиолого-биохимических показателей сосуществующих в едином биотопе сеголетков аборигенного вида – серебряного карася Carassius auratus gibelio (Bloch, 1972) и инвазионного – ротана-головешки Perccottus glenii (Dybowski, 1877) в оз. Круглое – одном из водоемов Мордовинской поймы Саратовского водохранилища, выявил межвидовые различия в их ростовых и физиолого-биохимических показателях. Снижение массы тела, жирности и калорийности сеголетков серебряного карася по мере увеличения длины тела в летние месяцы свидетельствует о снижающемся уровне обеспеченности пищей потомства по мере его роста. Повышение аналогичных ростовых и биохимических показателей сеголетков ротана указывает на лучшую, по сравнению с молодью серебряного карася, обеспеченность пищей особей всех размерных групп в генерации. На этом основании можно предположить, что преимуществом в выживании обладает потомство ротана, и численность его популяций в водоемах Мордовинской поймы Саратовского водохранилища в ближайшие годы, по-видимому, будет возрастать. Одновременно с этим следует ожидать снижение численности пополнения популяций серебряного карася и, соответственно, изменения популяционной структуры этого аборигенного вида рыб.
Работа выполнена в рамках государственного задания № АААА-А18-118012690123-4, АААА-А17-117112040040-3.
Список литературы
Евланов И.А., Козловский С.В., Антонов П.И. Кадастр рыб Самарской области. Тольятти: Изд-во Института экологии волжск. бассейна РАН, 1998. 222 с.
Кириленко Е.В., Шемонаев Е.В. Данные о морфологии и биологии ротана-головешки Perccottus glenii Dybowski, 1877 из озера Круглое Мордовинской поймы Саратовского водохранилища // Изв. Самар. науч. центра РАН. 2011. Т. 13. № 1. С. 207–210.
Кириленко Е.В., Шемонаев Е.В. Актуальные проблемы экологии и охраны окружающей среды // Матер. X Междунар. науч.-практ. конф. “Татищевские чтения: актуальные проблемы науки и практики”. Тольятти: Волжск. ун-т им. В.Н. Татищева, 2013. С. 60–66.
Лакин Г.Ф. Биометрия. М.: Высш. шк., 1990. 352 с.
Лапин В.И., Чернова Е.Г. О методике экстракции жира из сырых тканей рыб // Вопр. ихтиологии. 1970. Т. 10. Вып. 4. С. 753–756.
Шатуновский М.И. Экологические закономерности обмена веществ морских рыб. М.: Наука, 1980. 281 с.
Шульман Г.Е. Физиолого-биохимические особенности годовых циклов рыб. М.: Пищ. пром-сть, 1972. 368 с.
Шульман Г.Е., Кокоз Л.M. Особенности белкового роста и жиронакопления у черноморских рыб // Биология моря. 1968. Вып. 15. С. 159–203.
Шульман Г.Е., Кокоз Л.М. Содержание сухого обезжиренного вещества в теле некоторых черноморских рыб // Вопр. ихтиологии. 1971. Т. 11. Вып. 2. С. 339−344.
Халько В.В. Закономерности формирования продукционных показателей молоди рыб разных экологических групп: Автореф. дис. … канд. биол. наук. М., 1983. 24 с.
Folch J., Lees M., Stonley A. A simple method for isolation and purification of total lipids from animal tissues // J. Biol. Chem. 1957. V. 226. № 1. P. 497–509.
Дополнительные материалы отсутствуют.
Инструменты
Биология внутренних вод