

Биология внутренних вод, 2019, № 1, стр. 23-30
Структурные и количественные особенности сообществ фитоперифитона в водотоках бассейна реки Зея (Амурская область)
Л. А. Медведева 1, *, А. А. Семенченко 1, 2
1 Федеральный научный центр биоразнообразия наземной биоты Восточной Азии Дальневосточное отделение Российской академии наук
690022 Владивосток, просп. 100-летия Владивостока, д. 159, Россия
2 Дальневосточный федеральный университет
690950 Владивосток, ул. Суханова, д. 8, Россия
* E-mail: medvedeva@ibss.dvo.ru
Поступила в редакцию 25.10.2016
После доработки 20.06.2017
Принята к публикации 05.03.2017
Аннотация
Приведены результаты исследования водорослей перифитона р. Зея и девяти водотоков ее бассейна (Амурская обл.). Определен состав доминирующих видов, рассмотрены структурные особенности сообществ, получены количественные характеристики их отдельных компонентов. Общая численность фитоперифитона в р. Зея была 583 млрд кл./м2, биомасса – 102–1106 г/м2. В небольших водотоках численность колебалась от 16.2 до 100 млрд кл./м2, биомасса – от 2.0 до 18.3 г/м2. Основу биомассы составляли диатомовые водоросли (Encyonema silesiacum, Hannaea arcus, Ulnaria ulna, Gomphoneis olivaceum, крупноклеточные виды рода Gomphonema), основу численности – цианобактерии и диатомовые водоросли из родов Achnanthes, Fragilaria, Gomphonema и вид Encyonema minutum.
ВВЕДЕНИЕ
В лотических системах Дальнего Востока доминируют прикрепленные водоросли (фитоперифитон, эпилитон), способные выдерживать воздействие потока воды. Альгоценозы обрастаний характерны для малых рек, где наличие твердого субстрата и высокие скорости течения ограничивают развитие других экологических группировок водорослей [15]. Структура фитоперифитона определяет формирование и функционирование всех элементов водного биоценоза. Водорослевые обрастания образованы видами, различающимися по размеру (от нескольких микрон до нескольких десятков сантиметров) и морфологии. Сообщества чаще всего состоят из прикрепленных одноклеточных, преимущественно гетерополярных колониальных и нитчатых форм, приспособленных к условиям динамичного потока воды [15, 24, 31].
Изучению альгоценозов обрастаний пресноводных водоемов посвящено большое количество работ [4, 7, 9, 13–21, 23, 26, 29, 32, 35, 42–45], в которых в основном описана сукцессия видов или структура фитоперифитона на уровне отделов. Данные по биомассе и численности водорослей, а также их сезонной динамике приводятся реже [3, 34].
Первые сведения о водорослях р. Зея опубликованы Б.В. Скворцовым [28]. В торфяных болотах в верховьях реки были найдены десмидиевые и диатомовые водоросли, среди которых преобладал Cosmarium amurense Skvortzow. Проведенное авторами обследование ряда водотоков позволило значительно расширить сведения о пресноводных водорослях Амурской обл. [19].
Цель работы – исследование структуры сообществ прикрепленных водорослей водотоков бассейна р. Зея на уровне отделов, родов и видов и количественная оценка альгоценозов обрастаний.
МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЯ
Пробы перифитона собирали летом 2014 г. в р. Зея и девяти водотоках ее бассейна: Алленга, Тында, Малый Гармакан, Большой Гармакан, Широковская, Малая Макча, Большая Макча, Большая Эракингра, Гулик (рис. 1). Обработку материала проводили по общепринятым методикам [6]. Водорослевые обрастания с камней (эпилитон) смывали водой (объем 100 мл) и фиксировали 4%-ным формалином. Пробы просчитывали в счетной камере собственной конструкции [2] под микроскопом “Amplival” (увеличение ×400 и ×1000), при определении видов использовали работы [5, 8, 10–12, 22, 33, 36–39]. Идентификацию диатомовых водорослей проводили в постоянных препаратах, приготовленных перекисным методом. К доминирующим относили виды, численность или биомасса которых составляла ≥10% общей. Массу нитчатых водорослей Stigeoclonium tenue (C. Agardh) Kützing, 1843 определяли взвешиванием на торсионных весах материала, подсушенного при комнатной температуре.
Рис. 1.
Схема расположения станций отбора проб в водотоках бассейна р. Зея: 1 – Алленга, 2 – Тында, 3 – Малый Гармакан, 4 – Большой Гармакан, 5 – Широковская, 6 – Зея ниже плотины ГЭС, 7 – Зея у пос. Овсянка, 8 – Малая Макча, 9 – Большая Макча, 10 – Большая Эракингра, 11 – Гулик.
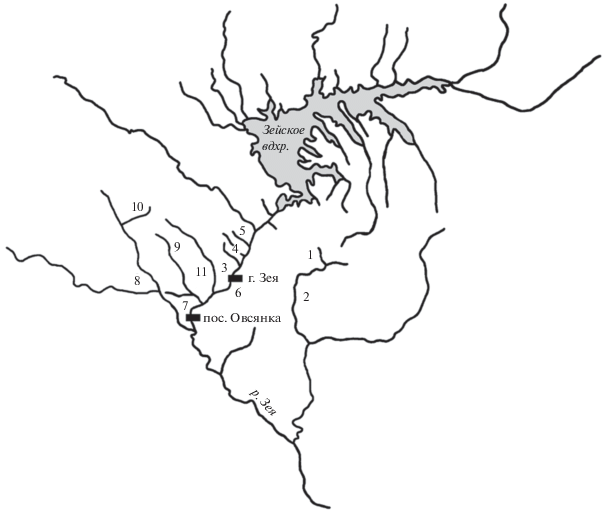
Река Зея – один из крупнейших левобережных притоков р. Амур. Исток реки находится на хребте Токинский Становик (55°44′10″ с.ш., 130°34′20″ в.д.), устье у г. Благовещенск (50°14′31″ с.ш., 127°35′52″ в.д.). Протяженность реки 1242 км, общая площадь водосбора 233 тыс. км2. Водосборный бассейн, который целиком располагается в пределах Амурской обл., характеризуется горным рельефом и сложным орографическим строением. Почти все притоки берут начало на отрогах горных хребтов, в верховьях они представляют собой бурные потоки, протекающие по узким ущельям. Южная часть бассейна охватывает территорию Зейско-Буреинской низменности, на которой водотоки обретают спокойное течение [25, 27]. На территории бассейна распространена многолетняя мерзлота, речные и грунтовые наледи сохраняются местами до летнего сезона. Муссонный характер климата определяет основные черты водного режима реки. Доля дождевого питания в среднем составляет 50–70% общего годового стока. С апреля по октябрь отмечаются 4–5 паводков, при которых уровень воды поднимается на 4–6 м, а скорость течения увеличивается до 3–4 м/с. Воды р. 3ея и ее притоков относятся к слабо минерализованным (минерализация <100 мг/дм3) ультрапресным [30], по классификации О.А. Алекина [1] – к гидрокарбонатно-кальциево-магниевым 2-го типа.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
В обрастаниях камней водотоков бассейна р. Зея обнаружено 128 видов водорослей и цианобактерий (131 с учетом внутривидовых таксонов) из 58 родов: Cyanobacteria – 7 видов, Euglenophyta – 2, Chrysophyta – 1, Bacillariophyta – 104 (107), Rhodophyta – 3, Chlorophyta – 11.
Наибольшее число видовых и внутривидовых таксонов (28–38) зафиксировано в реках Зея у пос. Овсянка, Большая и Малая Макча, наименьшее (14) – в реках Широковская и Большая Эракингра (табл. 1). В остальных водотоках отмечено 17 – 25 видов.
Таблица 1.
Данные по биомассе и численности основных отделов эпилитона в обследованных водотоках бассейна р. Зея
| Номер водотока | Река | Число видов | Циано- бактерии |
Золо- тистые |
Эвгле- новые |
Диато- мовые |
Крас- ные |
Зеле- ные |
Общая |
|---|---|---|---|---|---|---|---|---|---|
| 1 | Алленга | 17 | $\frac{{0.7}}{{0.0}}$ | – | – | $\frac{{15.4}}{{4.1}}$ | – | $\frac{{0.5}}{{0.2}}$ | $\frac{{16.6}}{{4.3}}$ |
| 2 | Тында | 18 | – | – | – | $\frac{{31.5}}{{11.8}}$ | – | $\frac{{0.3}}{{0.1}}$ | $\frac{{31.8}}{{11.9}}$ |
| 3 | Малый Гармакан | 21 | $\frac{{14.4}}{{0.4}}$ | – | – | $\frac{{2.0}}{{0.8}}$ | – | $\frac{{9.0}}{{3.1}}$ | $\frac{{25.4}}{{4.3}}$ |
| 4 | Большой Гармакан | 23 | $\frac{{0.9}}{{0.0}}$ | – | – | $\frac{{6.5}}{{3.6}}$ | $\frac{{1.3}}{{1.6}}$ | $\frac{{7.5}}{{4.5}}$ | $\frac{{16.2}}{{9.7}}$ |
| 5 | Широковская | 14 | $\frac{{95.9}}{{2.0}}$ | – | – | $\frac{{4.2}}{{1.1}}$ | – | – | $\frac{{100}}{{3.1}}$ |
| 6 | Зея ниже плотины ГЭС* | 25 | $\frac{{36.9}}{{10.3}}$ | – | – | $\frac{{27.8}}{{31.3}}$ | – | $\frac{{27.6}}{{1064}}$ | $\frac{{92.3}}{{1106}}$ |
| 7 | Зея у пос. Овсянка | 28 | $\frac{{386}}{{6.6}}$ | $\frac{{0.4}}{{0.4}}$ | $\frac{{0.4}}{{0.6}}$ | $\frac{{196}}{{94.0}}$ | – | – | $\frac{{583}}{{102}}$ |
| 8 | Малая Макча | 38 | – | – | – | $\frac{{4.5}}{{2.8}}$ | $\frac{{12.2}}{{15.3}}$ | $\frac{{0.5}}{{0.2}}$ | $\frac{{17.2}}{{18.3}}$ |
| 9 | Большая Макча | 29 | – | – | – | $\frac{{16.2}}{{8.2}}$ | – | – | $\frac{{16.2}}{{8.2}}$ |
| 10 | Большая Эракингра | 14 | $\frac{{20.0}}{{0.1}}$ | – | – | $\frac{{47.7}}{{15.3}}$ | – | $\frac{{1.2}}{{0.7}}$ | $\frac{{68.9}}{{16.1}}$ |
| 11 | Гулик | 23 | $\frac{{44.8}}{{0.9}}$ | – | – | $\frac{{2.9}}{{1.0}}$ | $\frac{{0.1}}{{0.1}}$ | – | $\frac{{47.8}}{{2.0}}$ |
Примечание. Здесь и в табл. 2 над чертой – численность, млрд кл./м2; под чертой – биомасса, г/м2 эпилитона. “–” – отсутствие группы. * Численность подсчитана без учета нитчаток.
В структуре сообщества р. Алленга по численности преобладали диатомовые водоросли Achnanthidium minutissimum (Kützing) Czarnecki, Gomphoneis olivaceum (Hornemann) Dawson ex Ross et Sims, Gomphonema angustatum (Kützing) Rabenhorst и G. angustum C. Agardh.
Видовой состав эпилитона в р. Тында сходен с таковым в р. Алленга. Здесь также доминировали Achnanthidium minutissimum, Gomphoneis olivaceum, Gomphonema angustatum и G. angustum, в качестве субдоминантов к ним прибавились Planothidium lanceolatum (Brébisson ex Kützing) Round et Bukhtiyarova и Reimeria sinuata (Gregory) Kociolek et Stoermer.
В р. Малый Гармакан развивались цианобактерии, диатомовые и один вид зеленых водорослей. В массе вегетировали Achnanthidium minutissimum, Cocconeis placentula Ehrenberg, Reimeria sinuata. В качестве субдоминанта отмечена Rhoicosphenia abbreviata (C. Agardh) Lange-Bertalot, не обнаруженная в других обследованных водотоках. Вид относится к ксено-олигосапробионтам, т.е. к показателям вод хорошего качества, предпочитает воды с низким содержанием солей.
В составе эпилитона в р. Большой Гармакан, кроме цианобактерий, диатомовых и зеленых водорослей, выявлен представитель отдела красных водорослей (Rhodophyta) Audouinella Bory. По численности доминировали виды рода Gomphonema: G. angustum и G. parvulum (Kützing) Kützing, а также Achnanthidium minutissimum, Cocconeis placentula и Reimeria sinuata.
В эпилитоне р. Широковская к доминирующим по численности видам относились Gomphonema parvulum и Gomphoneis olivaceum, к субдоминантам – Achnanthidium minutissimum и виды рода Encyonema.
В русле р. Зея ниже плотины Зейской ГЭС камни были покрыты мощными скоплениями зеленых нитчатых водорослей Stigeoclonium tenue. В состав доминантов входили также диатомовые Achnanthidium minutissimum, Diatoma mesodon (Ehrenberg) Kützing, Fragilaria capucina Desmazières, Gomphoneis olivaceum, Gomphonema parvulum, Tabellaria fenestrata (Lyngbye) Kützing, T. flocculosa (Roth) Kützing. Два последних вида характерны для стоячих водоемов и заводей рек. Ниже по течению р. Зея у пос. Овсянка массовые скопления Stigeoclonium tenue не отмечены. Комплекс доминирующих и по численности и биомассе видов диатомовых остался почти неизменным: Gomphonema parvulum, Achnanthidium minutissimum, Gomphoneis olivaceum, кроме них в массе развивалась цианобактерия Homoeothrix janthina (Bornet et Flahault) Starmach.
К особенностям эпилитона р. Малая Макча относилось преобладание красных водорослей из рода Audouinella. В массе вегетировали диатомовые Gomphonema angustatum, G. angustum, G. parvulum. В отличие от других рек, здесь обнаружено несколько видов рода Eunotia Ehrenberg (E. bilunaris (Ehrenberg) Mills, E. diadema Ehrenberg, E. diodon Ehrenberg, E. fallax A. Cleve-Euler, E. minor (Kützing) Grunow, E. praerupta Ehrenberg). Его представители чаще всего встречаются в заболоченных водах с пониженными значениями рН.
Эпилитон р. Большая Макча представлен исключительно диатомовыми водорослями. К массовым видам относились Achnanthidium minutissimum, Encyonema minutum, Fragilaria capucina, Cocconeis placentula и Gomphoneis olivaceum. Два последних вида доминировали и по биомассе.
Группировка водорослей в р. Большая Эракингра также в основном состояла из диатомовых с доминированием Achnanthidium minutissimum, Gomphoneis olivaceum, Gomphonema angustatum, G. angustum. Основная доля биомассы сообщества приходилась на виды родов Gomphoneis и Gomphonema.
В обрастаниях камней р. Гулик по численности преобладала мелкоклеточная цианобактерия Homoeothrix janthina, из диатомей наибольшая частота встречаемости отмечена у Reimeria sinuata.
Показатели развития эпилитона в небольших водотоках колебались в узких пределах (табл. 1): общая численность – 16.2–100 млрд кл./м2, биомасса – 2.0–18.3 г/м2. Наименьшая численность отмечена в реках Большая Макча, Большой Гармакан, Алленга; наименьшая биомасса – в реках Гулик, Широковская, Алленга. Эпилитон р. Зея от плотины ГЭС до пос. Овсянка характеризовался очень высокими количественными показателями, достигающими 583 млрд кл./м2 и 102–1106 г/м2. При этом ниже плотины Зейской ГЭС масса нитчатых водорослей Stigeoclonium tenue, от которых невозможно было отделить диатомеи, составляла 996 мг/м2.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Анализ структуры альгоценозов обрастаний в водотоках бассейна р. Зея позволил выделить основные типы сообществ на уровне отделов. В реках Малый Гармакан, Широковская, Гулик, на участке р. Зея у пос. Овсянка развиваются сообщества с численным преобладанием цианобактерий, в реках Алленга, Тында, Большая Макча – диатомовых водорослей. На обследованных участках рек Большой Гармакан и Зея ниже плотины ГЭС примерно в равных соотношениях численности представлены цианобактерии, диатомовые и зеленые водоросли. В р. Малая Макча выявлены группировки, в которых преобладают диатомовые или красные водоросли (рис. 2а).
Рис. 2.
Относительная численность (а) и биомасса (б) отделов водорослей обрастаний в водотоках бассейна р. Зея. 1 – цианобактерии, 2 – диатомовые, 3 – красные, 4 – зеленые. Обозначения 1–11 на оси абсцисс см. рис. 1.
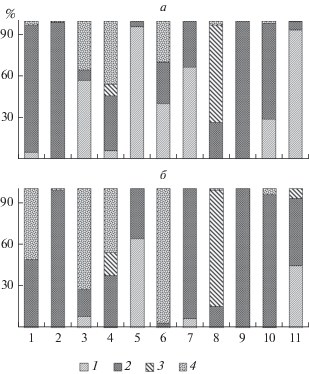
В сложении биомассы основная роль чаще принадлежит диатомовым водорослям, доминирующим в реках Тында, Зея у пос. Овсянка, Большая Макча, Большая Эракингра. Реже встречаются сообщества, в которых по биомассе превалируют зеленые водоросли (р. Зея ниже плотины Зейской ГЭС, реки Малый и Большой Гармакан), а в единичных случаях – красные водоросли (р. Малая Макча) и цианобактерии (р. Широковская) (рис. 2б).
У большинства рек Дальнего Востока сходная структура фитоперифитона с преобладанием диатомовых водорослей и цианобактерий [18–20, 41]. В целом наши данные сопоставимы с полученными при изучении водорослей перифитона водотоков различных рек данного региона [4, 15, 21, 32, 35, 42–45], лишь в отдельных случаях отмечено массовое развитие цианобактерий и зеленых водорослей [40, 46].
Высоким видовым разнообразием во всех обследованных водотоках характеризуются диатомовые водоросли, которые часто преобладают количественно, особенно в сложении биомассы сообществ (табл. 2). К наиболее распространенным относятся виды Achnanthidium minutissimum, Cocconeis placentula, Gomphoneis olivaceum, Gomphonema angustatum, G. parvulum, G. angustum, Reimeria sinuata и Ulnaria ulna (Nitzsch) Compère. Часто в состав доминантов входят представители маловидовых или монотипных родов Hannaea Patrick, Tabellaria Ehrenberg ex Kützing, Cocconeis Ehrenberg, Meridion C. Agardh.
Таблица 2.
Видовое разнообразие диатомовых водорослей эпилитона обследованных водотоков
| Вид | Водоток* | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | |
| Achnanthidium sp. | $\frac{{7.4}}{{0.9}}$ | $\frac{{5.8}}{{0.9}}$ | $\frac{{0.3}}{{0.1}}$ | $\frac{{0.4}}{{0.1}}$ | – | $\frac{{2.1}}{{0.3}}$ | $\frac{{83.8}}{{13.4}}$ | – | $\frac{{4.1}}{{0.7}}$ | $\frac{{21.3}}{{3.4}}$ | $\frac{{0.9}}{{0.1}}$ |
| Asterionella formosa | – | – | – | – | – | – | – | – | – | – | |
| Brebissonia boeckii | – | – | – | – | – | $\frac{{0.3}}{{1.1}}$ | – | – | – | – | – |
| Cocconeis placentula | – | – | $\frac{{0.2}}{{0.2}}$ | $\frac{{2.4}}{{2.4}}$ | – | – | – | – | $\frac{{0.5}}{{0.5}}$ | $\frac{{0.2}}{{0.2}}$ | $\frac{{0.4}}{{0.5}}$ |
| Diatoma mesodon | – | – | – | – | – | $\frac{{9.3}}{{10.9}}$ | $\frac{{5.6}}{{4.3}}$ | – | – | – | – |
| D. tenue | – | – | – | – | – | $\frac{{0.3}}{{0.1}}$ | $\frac{{4.0}}{{0.7}}$ | – | – | – | – |
| D. vulgare | – | – | – | – | – | – | $\frac{{5.6}}{{5.0}}$ | – | – | – | – |
| Encyonema minutum | $\frac{{0.2}}{{0.0}}$ | $\frac{{1.6}}{{0.2}}$ | – | $\frac{{1.7}}{{0.2}}$ | $\frac{{0.8}}{{0.1}}$ | – | $\frac{{4.0}}{{0.4}}$ | – | $\frac{{6.7}}{{0.7}}$ | $\frac{{0.5}}{{0.1}}$ | – |
| E. silesiacum | – | – | – | $\frac{{0.4}}{{0.3}}$ | $\frac{{0.5}}{{0.4}}$ | $\frac{{0.8}}{{0.6}}$ | $\frac{{5.2}}{{4.2}}$ | $\frac{{0.2}}{{0.2}}$ | $\frac{{2.0}}{{1.6}}$ | $\frac{{0.2}}{{0.1}}$ | – |
| Fragilaria capucina | – | – | – | – | – | $\frac{{1.1}}{{0.4}}$ | $\frac{{1.2}}{{0.7}}$ | $\frac{{0.2}}{{0.1}}$ | $\frac{{0.5}}{{0.4}}$ | – | – |
| F. sp. 1 | – | $\frac{{0.3}}{{0.1}}$ | $\frac{{0.1}}{{0.0}}$ | – | – | $\frac{{0.3}}{{0.1}}$ | $\frac{{5.2}}{{1.6}}$ | $\frac{{0.5}}{{0.1}}$ | $\frac{{2.0}}{{0.6}}$ | $\frac{{0.2}}{{0.1}}$ | $\frac{{0.2}}{{0.1}}$ |
| F. vaucheriae | – | $\frac{{2.0}}{{0.4}}$ | – | – | – | $\frac{{0.3}}{{0.1}}$ | – | – | – | – | – |
| Gomphonema sp. 1 (крупная) Gomphonema affine |
– | – | – | $\frac{{0.2}}{{0.3}}$ | – | $\frac{{0.5}}{{0.4}}$ | $\frac{{1.6}}{{2.6}}$ | – | $\frac{{1.0}}{{1.6}}$ | $\frac{{1.5}}{{2.4}}$ | – |
| Gomphonema sp. 2 (средняя) Gomphonema angustatum Gomphoneis olivaceum |
$\frac{{5.0}}{{2.2}}$ | $\frac{{7.9}}{{4.0}}$ | $\frac{{0.8}}{{0.4}}$ | $\frac{{0.3}}{{0.1}}$ | $\frac{{1.1}}{{0.5}}$ | $\frac{{11.2}}{{5.0}}$ | – | – | $\frac{{17.5}}{{7.9}}$ | – | |
| Gomphonema sp. 3 (мелкая) Gomphonema parvulum |
$\frac{{2.4}}{{0.4}}$ | $\frac{{9.9}}{{1.8}}$ | $\frac{{0.3}}{{0.1}}$ | $\frac{{0.2}}{{0.0}}$ | $\frac{{2.4}}{{0.4}}$ | $\frac{{0.3}}{{0.1}}$ | $\frac{{38.9}}{{7.0}}$ | $\frac{{0.2}}{{0.0}}$ | $\frac{{4.1}}{{0.7}}$ | $\frac{{6.0}}{{1.1}}$ | $\frac{{0.9}}{{0.2}}$ |
| Hannaea arcus | – | $\frac{{0.3}}{{0.3}}$ | $\frac{{0.1}}{{0.1}}$ | – | – | $\frac{{2.1}}{{1.5}}$ | $\frac{{9.8}}{{6.9}}$ | – | $\frac{{1.0}}{{1.1}}$ | $\frac{{0.1}}{{0.1}}$ | – |
| Meridion circulare | – | – | – | – | – | – | $\frac{{0.8}}{{0.2}}$ | – | $\frac{{0.5}}{{0.1}}$ | – | $\frac{{0.2}}{{0.1}}$ |
| Navicula sp. | – | – | – | – | – | – | $\frac{{4.0}}{{1.8}}$ | $\frac{{0.2}}{{0.1}}$ | – | – | $\frac{{0.2}}{{0.1}}$ |
| Nitzschia sp. | – | $\frac{{0.1}}{{0.1}}$ | – | – | – | $\frac{{1.3}}{{0.5}}$ | $\frac{{3.2}}{{0.5}}$ | – | – | – | – |
| Reimeria sinuata | $\frac{{0.2}}{{0.0}}$ | $\frac{{0.8}}{{0.2}}$ | – | $\frac{{0.6}}{{0.1}}$ | $\frac{{0.3}}{{0.1}}$ | – | – | – | – | – | – |
| Rhoicosphenia abbreviata | – | – | $\frac{{0.1}}{{0.1}}$ | – | – | – | – | – | – | – | – |
| Ulnaria ulna | $\frac{{0.1}}{{0.4}}$ | $\frac{{0.4}}{{2.5}}$ | – | – | – | $\frac{{0.9}}{{6.5}}$ | $\frac{{4.2}}{{29.5}}$ | – | – | – | – |
| Tabellaria fenestrata | – | – | – | – | – | $\frac{{4.0}}{{6.2}}$ | $\frac{{6.2}}{{9.6}}$ | – | – | – | – |
| T. flocculosa | – | – | – | – | – | $\frac{{1.3}}{{1.1}}$ | – | – | – | – | – |
* Номера водотоков соответствуют названиям, указанным в табл. 1.
Главная роль в формировании эпилитона принадлежит гетерополярным клеткам из родов Gomphonema Ehrenberg и Gomphoneis Cleve. При обработке количественных проб невозможно точно установить их видовую принадлежность, поэтому условно выделены три группы с разными размерами клеток. Просмотр постоянных препаратов позволил отнести крупные гомфонемы к Gomphonema affine Kützing и G. angustum, средние – к G. angustatum и Gomphoneis olivaceum, мелкие – к Gomphonema parvulum. Постоянные спутники гомфонем – виды рода Achnanthidium Kützing, Cocconeis placentula, Hannaea arcus (Ehrenberg) Patrick, обладающие способностью прикрепляться к субстрату. Для сообществ типичны также виды родов Encyonema Kützing (E. minutum, E. silesiacum) и Fragilaria Lyngbye. Крупноклеточная Ulnaria ulna обычно встречалась в небольших количествах, однако за счет своих размеров часто лидировала по биомассе.
Виды, которые доминируют по численности и биомассе или характеризуются максимальными значениями этих показателей, часто различны. Так, в перифитоне рек Восточной Фенноскандии численно преобладают диатомеи, а биомассу формируют, в основном, нитчатые зеленые и красные водоросли [15]. Причины этого связаны со структурой сообществ и морфологическими особенностями водорослей обрастаний, представленых видами с различными размерами.
В обследованных водотоках к видам, доминирующим по численности, относятся мелкоклеточные диатомовые Achnanthidium sp., Fragilaria sp. 1, Gomphonema sp. 3 (р. Зея в районе пос. Овсянка) и Encyonema minutum (р. Большая Макча). Основу биомассы чаще всего составляют крупноклеточные формы Gomphonema sp. 2 (cf. Gomphoneis olivaceum), Encyonema silesiacum, Hannaea arcus, Ulnaria ulna, Tabellaria fenestrata (р. Зея в районе пос. Овсянка). Gomphonema sp. 2 (cf. Gomphoneis olivaceum) доминирует также в р. Большая Эракингра, а Tаbellaria fenestrata – в р. Зея ниже плотины ГЭС (табл. 2). Иногда в сложении биомассы существенную роль играют мелкоклеточные виды с высокой численностью: Achnanthidium sp. и Gomphonema sp. 3 (р. Зея в районе пос. Овсянка).
В водорослевых обрастаниях р. Зея, в отличие от ее притоков, наряду с водорослями перифитона в значительных количествах обнаружены планктонные формы Asterionella formosa Hassall, Diatoma tenue C. Agardh, D. vulgare Bory, Fragilaria capucina, Nitzschia sp., Tabellaria fenestrate и T. flocculosa. Только ниже плотины Зейской ГЭС в состав доминантов входит галофил Brebissonia boeckii (Ehrenberg) Grunow, развивающийся в слабо солоноватых водах. Высокие показатели численности и биомассы эпилитона отмечаются на этом участке реки постоянно. Так, в 2010 г. камни в русле реки были покрыты мощным ковром водорослей толщиной до 2 см из нескольких нитчатых форм. Основную массу обрастаний на плесе формировали цианобактерии Phormidium corium (C. Agardh) Gomont, Phormidium sp., Symploca sp., Lyngbya sp., Aphanizomenon sp. и одноклеточный Synechococcus sp., а на перекате – нитчатые зеленые водоросли Ulothrix zonata (Weber et Mohr) Kützing и U. tenuissima Kützing. Биомасса водорослей на перекате достигала 1130 г/м2 [19], хотя температура воды на данном участке даже в летнее время не поднимается выше 5°С, высокое обилие эпилитона может быть связано со снижением скорости течения или с поступлением в русло реки вод из глубинных слоев водохранилища, богатых растворенными биогенными веществами.
Выводы. В водотоках бассейна р. Зея обнаружено 128 видов водорослей из шести отделов с преобладанием цианобактерий, диатомовых и зеленых. К доминирующим по численности видам относятся диатомовые водоросли Encyonema minutum и представители родов Achnanthidium, Fragilaria и Gomphonema. Основу биомассы составляют Encyonema silesiacum, Hannaea arcus, Ulnaria ulna, Gomphoneis olivaceum, крупноклеточные виды рода Gomphonema, реже – Tabellaria fenestrata. Максимальные численность и биомасса зарегистрированы в эпилитоне р. Зея.
Исследование выполнено при финансовой поддержке гранта Российского научного фонда (проект № 14-50-00034).
Список литературы
Алекин О.А. Основы гидрохимии. Л.: Гидрометеоиздат, 1970. 444 с.
Баринова С.С., Медведева Л.А. К методике количественного учета микрофитобентоса малых рек Дальнего Востока России // Альгология. 2004. Т. 14. № 1. С. 101–110.
Беляева П.Г. Структура фитоперифитонных сообществ в речных экосистемах (обзор) // Изв. Пензенск. гос. пед. ун-та им. В.Г. Белинского. 2011. Вып. 25. С. 484–492.
Беляева П.Г., Поздеев И.В. Донные сообщества р. Чусовая (бассейн Камы) // Вестн. Пермск. ун-та. Сер. Биол. 2005. Вып. 6. С. 103–107.
Виноградова К.Л., Голлербах М.М., Зауер Л.М., Сдобникова Н.В. Зеленые, красные и бурые водоросли. Определитель пресноводных водорослей СССР. Л.: Наука, 1980. Вып. 13. 248 с.
Водоросли. Справочник. Киев: Наук. думка, 1989. 608 с.
Глущенко Л.А., Дубовская О.П., Иванова Е.А. и др. Гидробиологический очерк некоторых озер горного хребта Ергаки (Западный Саян) // Журн. Сиб. фед. ун-та. Биология. 2009. Т. 2. № 3. С. 355–378.
Голлербах М.М., Косинская Е.К., Полянский В.И. Синезеленые водоросли. Определитель пресноводных водорослей СССР. М.: Сов. наука, 1953. Вып. 2. 652 с.
Девяткин В.Г. Динамика развития альгофлоры обрастаний в Рыбинском водохранилище // Тр. Ин-та биологии внутр. вод. АН СССР. 1979. Вып. 42(45). С. 78–108.
Диатомовые водоросли СССР (ископаемые и современные). Л.: Наука, 1988. Т. 2. Вып. 1. 116 с.
Диатомовые водоросли СССР (ископаемые и современные). СПб.: Наука, 1992. Т. 2. Вып. 2. 125 с.
Забелина М.М., Киселев И.А., Прошкина-Лавренко А.И., Шешукова В.С. Диатомовые водоросли. Определитель пресноводных водорослей СССР. М.: Сов. наука, 1951. Вып. 4. 619 с.
Игнатова Н.В., Помазкина Г.В. Биоценозы диатомовых водорослей и их роль в формировании поверхностного слоя донных отложений в южном Байкале // Проблемы экологии Прибайкалья: Тез. докл. 3 Всесоюз. науч. конф. Иркутск, 1988. Ч. 2. С. 96.
Качаева М.И. Количественный учет биомассы обрастаний водорослей р. Ингоды // Флора, растительность и растительные ресурсы Забайкалья и сопредельных областей. Вып. 4. Чита: Изд-во Забайкал. отд. Всесоюз. Ботан. об-ва АН СССР, 1972. С. 22–25.
Комулайнен С.Ф. Экология фитоперифитона малых рек Восточной Фенноскандии. Петрозаводск: Изд-во Карельск. науч. центра РАН, 2004. 182 с.
Левадная Г.Д. Микрофитобентос реки Енисей. Новосибирск: Наука, 1986. 286 с.
Левадная Г.Д., Чайковская Т.С. Водорослевая растительность Енисея и ее продукция // Круговорот вещества и энергии в водоемах. Элементы биотического круговорота. Лиственичное на Байкале, 1977. С. 96–99.
Медведева Л.А. Структурная характеристика сообществ водорослей перифитона водотоков бассейна реки Бурея (Хабаровский край, Российская Федерация) // Гидробиол. журн. 2006. Т. 42. № 6. С. 22–40.
Медведева Л.А. Альгологические исследования водотоков бассейна реки Зея и Зейского водохранилища // Гидробиологический мониторинг зоны влияния Зейского гидроузла. Хабаровск: Дальнаука, 2010. С. 45–92.
Медведева Л.А. Количественные характеристики сообществ перифитонных водорослей реки Кедровая (Приморский край) // Чтения памяти проф. В. Я. Леванидова. Вып. 6. Владивосток: Дальнаука, 2014. С. 443–452.
Метелёва Н.Ю. Структура и продуктивность фитоперифитона водоемов бассейна Верхней Волги: Автореф. дис. … канд. биол. наук. Институт билог. внутр. вод. Борок, 2013. 22 с.
Мошкова И.А., Голлербах М.М. Зеленые водоросли. Класс улотриксовые (1). Определитель пресноводных водорослей СССР. Л.: Наука, 1986. Вып. 10. 360 с.
Помазкина Г.В., Щербакова Т.А. Видовой состав Bacillariophyta литоральной зоны озера Байкал (Россия) // Альгология. 2010. Т. 20. № 4. С. 449–463.
Протасов А.А. Пресноводный перифитон. Киев: Наук. думка, 1994. 307 с.
Ресурсы поверхностных вод СССР. Дальний Восток. Т. 18: Верхний и Средний Амур. Л.: Гидрометеоиздат, 1966. Вып. 1. 781 с.
Рычкова М.А. Водоросли обрастаний озер Воже и Лача // Гидробиология озер Воже и Лача в связи с прогнозом качества вод, перебрасываемых на юг. Л.: Наука, 1978. С. 28–35.
Сиротский С.Е., Тесленко В.А. Физико-географическая характеристика бассейна реки Зея в районе исследований // Гидробиологический мониторинг зоны влияния Зейского гидроузла. Хабаровск: Дальнаука, 2010. С. 12–23.
Скворцов Б.В. Водоросли верховьев р. Зеи Амурской области. Материалы по флоре водорослей Азиатской России. 4. // Журн. рус. ботан. об-ва. 1917. Т. 2. С. 117–120.
Станиславская Е.В. Перифитон притоков Ладожского озера // Охрана и рациональное использование водных ресурсов Ладожского озера и других больших озер. СПб.: Лема, 2003. С. 249–252.
Шестеркина Н.М., Таловская В.С., Сиротский С.Е. и др. Условия формирования и качество воды реки Зея и ее притоков в пределах Зейско-Селемджинской и Амуро-Зейской возвышенных равнин // Гидробиологический мониторинг зоны влияния Зейского гидроузла. Хабаровск: Дальнаука, 2010. С. 23–35.
Algal ecology: freshwater benthic ecosystems. San Diego; N.Y.; Boston; L.; Sydney; Tokyo; Toronto: Acad. Press, 1996. 753 p.
Algarte V.M., Siqueira N.S., Rodrigues L. Desiccation and recovery of periphyton biomass and density in a subtropical lentic ecosystem // Acta Sci. Biol. Sci. 2013. V. 35. № 3. P. 311–318.
Hartley B., Barber H.G., Carter J.R. An Atlas of British Diatoms. Bristol: Biopress Ltd., 1996. 601 p.
Iwaniec D., Childers D.L., Rondeau D. et al. Effects of hydrologic and water quality drivers on periphyton dynamics in the southern Ever-glades // Hydrobiologia. 2006. V. 569. № 1. P. 223–235.
Izagirre O., Elosegi A. Environmental control of seasonal and inter-annual variations of periphytic biomass in a North Iberian stream // Ann. limnol. – Int. J. Lim. 2005. V. 41(1). P. 35–46.
Krammer K., Lange-Bertalot H. Bacillariophyceae: Naviculaceae. Süßwasserflora von Mitteleuropa. Bd 2, 1. Jena: Gustav Fisher Verlag, 1986. 876 S.
Krammer K., Lange-Bertalot H. Bacillariophyceae: Bacillariaceae, Epithemiaceae, Surirellaceae. Sußwasserflora von Mitteleuropa. Bd 2, 2. Stuttgart; N.Y.: Gustav Fisher Verlag, 1988. 596 S.
Krammer K., Lange-Bertalot H. Bacillariophyceae: Centrales, Fragilariaceae, Eunotiaceae. Sußwasserflora von Mitteleuropa. Bd 2, 3. Stuttgart; Jena: Gustav Fisher Verlag, 1991. 576 S.
Krammer K., Lange-Bertalot H. Bacillariophyceae: Achnanthaceae, Kritische Ergänzungen zu Navicula (Lineolatae) und Gomphonema Gesamtliteraturverzeichnis. Sußwasserflora von Mitteleuropa. Bd 2, 4. Stuttgart; Jena: Gustav Fisher Verlag, 1991. 437 S.
Lindstrøm E.A., Johansen S.W., Saloranta T. Periphyton in running waters – long-term studies of natural variation // Hydrobiologia. 2004. V. 521. № 1. P. 63–86.
Medvedeva L.A., Semenchenko A.A. Phytoperiphyton of the Samarga River Basin (Primorskii Krai) // Inland Water Biol. 2014. V. 7. № 2. P. 141–147. doi 10.1134/S1995082914010106
Nedbaliuc B., Chiriac E., Nedbaliuc R., Coadă V. Taxonomic structure and quantitative dinamics of the periphyton algoflora of the river Ichel (Republic of Moldova) // Analele Ştiinţifice ale Universitǎţii “Al. I. Cuza” Iaşi s. II a. Biologie vegetalǎ. 2013. V. 59. № 1. P. 27–33.
Pfeiffer T.Ž., Mihaljević M., Špoljarić D. et al. The disturbance-driven changes of periphytic algal communities in a Danubian floodplain lake // Knowl. Manag. Aquat. Ec. 2015. V. 416. № 2. P. 1–15.
Potapova M.G., Charles D.F. Benthic diatoms in USA rivers: distributions along spatial and environmental gradients // J. Biogeogr. 2002. V. 29. № 2. P. 167–187.
Potapova M.G., Charles D.F. Distribution of benthic diatoms in U.S. rivers in relation to conductivity and ionic composition // Freshwater Biol. 2003. V. 48. № 2. P. 1311–1328.
Villeneuve A., Montuelle B., Bouchez A. Effect of minor changes in light intensity, current velosity and turbulence on the structure and function of the periphyton // Aquat. Sci. 2010. V. 72. № 1. P. 33–44.
Дополнительные материалы отсутствуют.
Инструменты
Биология внутренних вод