

Биология внутренних вод, 2019, № 1, стр. 12-22
Центрохелидные солнечники разнотипных пресноводных местообитаний Южной Патагонии и Огненной Земли (Чили)
К. И. Прокина 1, *, А. П. Мыльников 1
1 Институт биологии внутренних вод им. И.Д. Папанина РАН
152742 Ярославская обл., Некоузский р-н, пос. Борок, Россия
* E-mail: kristin892@mail.ru
Поступила в редакцию 27.12.2016
После доработки 20.10.2017
Принята к публикации 12.05.2017
Аннотация
Впервые изучен видовой состав центрохелидных солнечников разнотипных пресноводных местообитаний юга Чили: сфагнового болота, временных и постоянных водоемов и водотоков. Выявлено 14 видов: Acanthocystis nichollsi, A. pectinata, A. penardi, Pseudoraphidiophrys discoidea, P. formosa, P. veliformis, Raphidiophrys ovalis, Choanocystis perpusilla, Pterocystis foliacea truncata, P. pyriformis, P. quadrata, Raineriophrys erinaceoides, R. fortesca и R. pteromorphos. Большинство видов впервые обнаружены в южной части страны, два (Acanthocystis nichollsi и Pterocystis quadrata) – новые для страны. Методами электронной микроскопии изучена внешняя морфология покровных неорганических чешуек клеток, имеющих важное диагностическое значение. Приведенные морфологические описания и фотографии исследованных чешуек расширяют представление о внутривидовой изменчивости и дополняют диагнозы видов.
ВВЕДЕНИЕ
Солнечники – полифилетическая группа протистов, включающая в себя организмы с округлой формой тела и схожими структурами клетки (псевдоподиями или аксоподиями с экструсомами; наружным скелетом в виде кремниевых или органических чешуек, реже – органических капсул), а также схожим типом питания (в большинстве случаев – гетеротрофы, поедающие других одноклеточных и микроскопических многоклеточных организмов) [4]. Центрохелиды – монофилетическая группа солнечников. Это облигатные гетеротрофы, имеющие кремниевый или органический наружный скелет в виде тангентально и радиально ориентированных чешуек [8]. Большая часть центрохелидных солнечников обнаружена в пресноводных континентальных местообитаниях, около половины из них найдена также в морских и соленых континентальных водах [3, 6, 7, 9, 27, 28]. Среди континентальных пресноводных объектов сфагновые болота отмечены как наиболее богатые по видовому составу центрохелидных солнечников [4, 8, 28].
Чили располагается в юго-западной части Южной Америки между западным побережьем Тихого океана и горной цепью Анд. Его климат меняется от тропического на севере до умеренного океанического на юге [1], как и состав фаунистических комплексов. Фауна солнечников ранее была хорошо изучена в центральной части этой страны [10, 15–17, 26], характеризующейся субтропическим климатом. Также есть немногочисленные данные о солнечниках Северной Патагонии [31]. Однако юг Чили, представляющий собой обширную территорию с массой разнотипных континентальных водных объектов, остается малоизученным.
Цель работы – изучение видового состава и деталей морфологии чешуек центрохелидных солнечников в разнотипных пресных биотопах Южной части Чили (Южная Патагония и Огненная Земля) методами трансмиссионной и сканирующей электронной микроскопии.
МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЯ
В октябре–ноябре 2015 г. исследованы сфагновое болото и разнотипные временные и постоянные водоемы и водотоки, располагающиеся в чилийской части Южной Патагонии и Огненной Земли. В континентальной части Южной Америки материал собирали в окрестностях г. Пунта-Аренас и в пределах Национального парка Торрес дель Пайне (Torres del Paine), на архипелаге Огненная Земля – в окрестностях оз. Бланко. Пробы отбирали из 14 водных объектов: сфагнового болота (мочажина, озерко и ручей – пробы № 1, 2, 3); пяти временных весенних водоемов (№ 4, 6, 8, 10, 11); оз. Бланко (№ 12), двух прудов на малой реке, заселенных бобрами Castor canadensis Kuhl, 1820 (№ 9, 14); трех постоянных водотоков (№ 5, 7, 13). Географические координаты водных объектов, их гидрофизические и гидрохимические данные на период сбора материала представлены в табл. 1. В работе приведены сведения только о тех водных объектах, в которых найдены солнечники. В еще 32 обследованных водных объектах солнечники не обнаружены, в том числе ни в одном водоеме с соленостью >2‰.
Таблица 1.
Физико-химическая характеристика водных объектов в 2015 г.
| № пробы |
Водный объект | Дата сбора |
Координаты, ю.ш., з.д. | Высота н. у. м, м | T, °C | pH | EC, мкСм | TDS, мг/л |
|---|---|---|---|---|---|---|---|---|
| Южная Патагония (South Patagonia), окрестности г. Пунта-Аренас (Punta Arenas) | ||||||||
| 1 | Сфагновое болото, мочажина | 26.Х | 53°24′01.2″, 71°13′32.2″ | 5 | 10.2 | 5.45 | 52 | 26 |
| 2 | озерко | 27.Х | 53°24′01.2″, 71°13′32.2″ | 5 | 11.8 | 4.43 | 45 | 29 |
| 3 | ручей | 27.Х | 53°24′01.2″, 71°13′32.2″ | 5 | 8.5 | 4.54 | 34 | 17 |
| 4 | Временный водоем | 9.ХI | 52°38′44.7″, 71°07′76.7″ | 3 | 9.8 | 8.84 | 794 | 398 |
| 5 | Ручей в окрестностях форта Бульнес | 18.ХI | 53°37′13.9″, 70°57′18.1″ | 10 | 9.4 | 8.75 | 111 | 55 |
| Национальный парк Торрес дель Пайне (Torres del Paine) | ||||||||
| 6 | Временное мелководное озеро | 01.ХI | 50°93′35.6″, 72°76′55.0″ | 200 | 17.5 | 8.06 | 382 | 191 |
| 7 | Ручей на склоне холма | 01.ХI | 50°59′15.1″, 72°47′47.7″ | 226 | 12.1 | 7.71 | 507 | 253 |
| 8 | Временный водоем между холмами | 01.ХI | 50°59′45.8″, 72°48′32.4″ | 309 | 12.7 | 8.13 | 526 | 262 |
| Огненная Земля (Tierra del Fuego), окрестности оз. Бланко (Blanko Lake) | ||||||||
| 9 | Жилой бобровый пруд на ручье | 04.ХI | 54°01′35.1″, 68°55′62.9″ | 125 | 8.6 | 7.16 | 115 | 57 |
| 10 | Временный водоем на краю леса | 04.ХI | 54°02′03″, 68°54′50.6″ | 28 | 19.6 | 7.79 | 229 | 114 |
| 11 | Временный водоем на лугу | 04.ХI | 54°01′89.7″, 68°54′78.3″ | 124 | 16.5 | 6.62 | 141 | 72 |
| 12 | оз. Бланко | 04.ХI | 54°01′62.2″, 68°56′16.9″ | 115 | 9.3 | 6.50 | 85 | 43 |
| 13 | Горная река | 04.ХI | 54°04′23.9″, 68°52′71.7″ | 158 | 13.5 | 7.06 | 105 | 54 |
| 14 | Нежилой бобровый пруд на горной реке (№ 13) | 04.ХI | 54°04′23.9″, 68°52′71.7″ | 158 | 11.3 | 7.44 | 296 | 146 |
Пробы воды с придонным грунтом помещали в 15-миллилитровые пластиковые сосуды и транспортировали в лабораторию при температуре 4°С. Там их помещали в чашки Петри и инкубировали в термостате при температуре 22°С в темноте. В виде корма солнечников использовали культуру жгутиконосцев Parabodo caudatus (Dujardin, 1841) Moreira et al., 2004. Для прижизненных наблюдений использовали световой микроскоп “AxioScope A1” (Carl Zeiss, Германия), оснащенный фазовым и интерференционным контрастами, объективами водной иммерсии (общее увеличение ×1120), цифровой фотокамерой “AxioCam ERc 5s” и аналоговой видеокамерой “AVT HORN MC1009/S”. Препараты солнечников для электронной микроскопии подготавливали согласно общепринятым методикам [8, 24] и просматривали в трансмиссионном (ТЭМ) “JEM-1011” и сканирующем (СЭМ) “JSM–6510 LV” электронных микроскопах (Joel, Япония). Для СЭМ живые клетки в капле воды с помощью микропипетки пересаживали на покровное стекло 18 × 18 мм и высушивали в термостате при температуре 60°С. Затем стекла промывали дистиллированной водой, снова высушивали, наклеивали на столик и напыляли золотом. Для ТЭМ клетки солнечников с каплей воды наносили на бленды, высушивали при комнатной температуре, промывали дистиллированной водой и снова высушивали, затем напыляли вольфрамом.
Идентификацию солнечников проводили по работам [5, 6, 8, 10, 14–19, 24, 25, 27, 28], используя современную систему [11, 37].
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
Обнаружено 14 видов центрохелидных солнечников из шести родов и четырех семейств. Данные о видовом составе найденных солнечников в водных объектах приведены в табл. 2. Морфологическая характеристика обнаруженных солнечников представлена ниже. Звездочками отмечены новые для фауны Чили виды.
Таблица 2.
Видовой состав центрохелидных солнечников в исследованных водных объектах
| Таксон | Водный объект* | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | |
| Cем. Acanthocystidae | ||||||||||||||
| Acanthocystis nichollsi | – | – | – | – | – | – | – | – | – | – | – | – | + | – |
| A. pectinata | – | – | – | + | + | – | – | – | – | – | – | – | – | – |
| A. penardi | + | – | – | + | + | – | – | – | – | – | – | + | – | + |
| Сем. Raphidiophryidae | ||||||||||||||
| Pseudoraphidiophrys discoidea | – | – | + | – | – | – | – | – | – | – | – | – | – | – |
| P. formosa | + | – | – | – | – | + | – | – | – | – | – | – | – | – |
| P. veliformis | + | – | – | – | – | – | – | – | – | – | – | – | – | – |
| Raphidiophrys ovalis | + | + | – | – | – | + | – | – | – | – | – | – | – | – |
| Сем. Choanocystidae | ||||||||||||||
| Choanocystis perpusilla | – | – | + | – | – | – | + | – | – | – | + | – | – | – |
| Сем. Pterocystidae | ||||||||||||||
| Pterocystis foliacea truncata | – | – | – | – | – | + | + | – | + | – | – | – | – | – |
| P. pyriformis | – | – | + | – | – | – | – | + | – | – | – | – | – | – |
| P. quadrata | – | – | – | – | – | – | – | – | – | + | + | – | – | – |
| Raineriophrys fortesca | – | – | – | – | – | – | – | – | – | – | + | – | – | – |
| R. pteromorphos | – | – | – | – | – | – | – | – | – | – | + | – | – | – |
| R. erinaceoides | – | – | – | – | – | – | – | + | – | – | + | – | – | – |
| Всего видов | 4 | 1 | 3 | 2 | 2 | 3 | 2 | 2 | 1 | 1 | 5 | 1 | 1 | 1 |
Класс Centrohelea Kühn, 1926 ex Cavalier-Smith, 1993;
Отряд Acanthocystida Cavalier-Smith in Yabuki et al., 2012;
Подотряд Chalarothoracina Hertwig and Lesser, 1874 emend Yabuki et al., 2012;
Семейство Acanthocystidae Claus, 1874 emend. Cavalier-Smith et von der Heyden, 2007.
Acanthocystis nichollsi* Siemensma et Roijackers, 1988 (рис. 1а–1г).
Обнаружены чешуйки одной клетки. Радиальные чешуйки двух типов. Первый тип – короткие, длиной 2.0–3.5 мкм, диаметр ствола 0.3 мкм, основание имеет небольшую базальную пластинку. Дистальная часть чешуйки расширена, разделяется на четыре–шесть фурок, которые соединяются тонкой мембраной. Диаметр расширенной дистальной части коротких радиальных чешуек 1.0–1.5 мкм. Второй тип – длинные радиальные чешуйки, 6.5–9.0 мкм в длину, отличаются от первого типа меньшим расширением верхушки и слабой выраженностью мембраны между фурками. Пластинчатые чешуйки овальные с небольшим сужением по бокам, 2.5–3.0 × 1.5–2.2 мкм. На поверхности пластинчатых чешуек находится аксиальный гребень 0.3–0.6 мкм длиной, а также радиальные слабовыраженные ребра, отсутствующие у некоторых чешуек.
Рис. 1.
Морфология чешуек солнечников сем. Acanthocystidae: а–г – Acanthocystis nichollsi, д–з – A. pectinata, и–м – A. penardi; а.г – аксиальный гребень, б.п – базальная пластинка радиальной чешуйки, п.ч – пластинчатая чешуйка, р.ч – радиальная чешуйка, ф – фурка. Использовали СЭМ для а–ж, и–м, ТЭМ – для з.
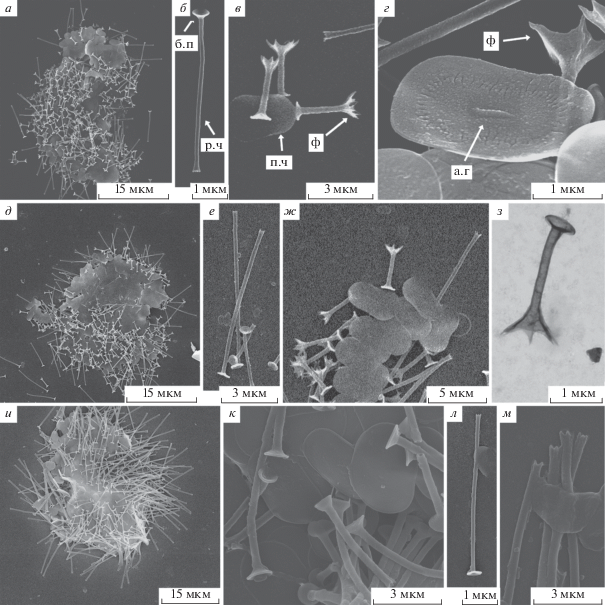
Морфология изученных чешуек соответствует предыдущим описаниям, однако радиальные ребра пластинчатых чешуек были слабо развиты или отсутствовали на некоторых чешуйках. Подобная изменчивость ранее отмечена у представителей этого вида [18]. Некоторые исследователи описывали чешуйки большего размера [5, 18, 20].
Распространение: пресные воды европейской части России [5, 19, 20], Нидерландов, Швеции [29], Великобритании [11], Индии [36], Монголии [18], Канады, как A. pectinata [26], США, как A. pectinata [33, 34]; соленые континентальные воды европейской части России [3].
Acanthocystis pectinata Penard, 1889 emend. Nicholls, 1983 [syn.: A. radiosa Roskin, 1929] (рис. 1д–1з).
Обнаружены чешуйки двух клеток. Радиальные чешуйки двух типов, схожи с чешуйками A. nichollsi. Первый тип – короткие, 2–3 мкм в длину, ствол 0.3 мкм в диаметре, основание немного расширено. Их верхняя часть чашеобразная, разделяется на четыре–шесть фурок, между которыми располагается тонкая мембрана. Диаметр дистальной расширенной части коротких чешуек – 1–2 мкм. Второй тип – длинные радиальные чешуйки, 7–10 мкм в длину, с небольшим расширением основания, дистальная часть несет две–три фурки. Пластинчатые чешуйки овальные с небольшим сужением по бокам, 2.0–2.5 × 1.0–1.5 мкм. Полоса беспорядочно расположенных мелких гранул лежит вдоль края пластинки.
Морфология изученных чешуек соответствует предыдущим описаниям, однако некоторые исследователи указывали большие размеры чешуек [5, 20, 25].
Распространение: пресные воды европейской части России [20, 22], Украины [2], Эстонии [23], Нидерландов, Швеции [29], Канады [25], Австралии [12], Чили, Новой Зеландии, Малайзии [15]; соленые континентальные воды европейской части России [3, 27].
Acanthocystis penardi Wailes, 1925 [syn.: A. spinifera Greef, 1869 sensu Penard, 1904 nec. A. spinifera Greef, 1869 sensu Siemensma et Roijackers, 1988; A. heterospina Nicholls, 1983; A. dresscheri Siemensma et Roijackers, 1988] (рис. 1и–1м).
Обнаружены чешуйки шести клеток. Радиальные чешуйки 6–14 мкм в длину, ствол 0.25–0.35 мкм в диаметре. Основание несет базальную пластинку 0.9–1.8 мкм в диаметре, дистальная часть разделяется на три–семь слабовыраженных фурок. Пластинчатые чешуйки овальной формы, 3.0–5.0 × 2.0–3.5 мкм, с хорошо выраженным утолщением края пластинки.
Морфология изученных клеток соответствует предыдущим описаниям, однако в работах [15, 18] указан меньший размер чешуек.
Распространение: пресные воды eвропейской части России [5, 19, 20, 22], Украины [2], Эстонии [23], Монголии [18], Канады, как A. heterospina [25], Малайзии, заболоченные водоемы Чили [10, 15].
Семейство Raphidiophryidae Mikrjukov, 1996 emend. Cavalier-Smith et von der Heyden, 2007.
Pseudoraphidiophrys discoidea (Dürrschmidt, 1985) Mikrjukov, 1997 [bas.: Acanthocystis discoidea Dürrschmidt, 1985; syn.: Pterocystis discoidea (Dürrschmidt, 1985) Siemensma, 1991] (рис. 2а).
Рис. 2.
Морфология чешуек солнечников сем. Raphidiophryidae: а – Pseudoraphidiophrys discoidea, б – P. formosa, в – P. veliformis, г – Raphidiophrys ovalis, д–ж – Choanocystis perpusilla, з–л – Pterocystis foliacea truncata. Использовали ТЭМ для а–ж, СЭМ – для з–л.

Обнаружено несколько наружных пластинчатых чешуек 2.75–3.70 мкм в диаметре округло-овальной формы. Разделены на две части – центральная часть имеет аксиальный гребень и радиально отходящие от него ребра, маргинальная часть бесфактурная. Внутренние чешуйки не исследованы.
Впервые описан в центре континентальной части Чили, в небольшом безымянном пруду в окрестностях г. Вальдивиа [Valdivia] [17].
Распространение: пресные воды Чили [17].
Pseudoraphidiophrys formosa (Dürrschmidt, 1985) Mikrjukov, 1997 [bas.: Acanthocystis formosa Dürrschmidt, 1985; syn.: Pterocystis formosa (Dürrschmidt, 1985) Siemensma, 1991] (рис. 2б).
Обнаружено несколько наружных пластинчатых чешуек 3.0–3.7 мкм в диаметре, стерна в виде утолщенной складки тянется от середины пластинки к краю, от нее к краям пластинки расходятся 30–35 радиальных тонких ребер. Края чешуйки неровные, слегка утолщены. Внутренние чешуйки не исследованы.
Впервые описан на Чилоэ, в сфагновом болоте в окрестностях г. Кастро [Castro] [17].
Распространение: пресные воды Чили [17], США [33, 35], Австралии [13].
Pseudoraphidiophrys veliformis (Dürrschmidt, 1985) Mikrjukov, 1997 [bas.: Acanthocystis veliformis Dürrschmidt, 1985; syn.: Pterocystis veliformis (Dürrschmidt, 1985) Siemensma, 1991] (рис. 2в).
Исследовано несколько наружных и внутренних пластинчатых чешуек. Наружные – крупные, парусовидные, 3–4 × 2–3 мкм. Стерна в виде продолговатого ребра, от которой к краям расходятся радиальные складки. От проксимального конца аксиального гребня выступает короткий стебелек в виде небольшого дорсального выпячивания пластинки. Внутренние чешуйки – небольшие, овальные, 1.0–1.5 × 0.5–1.0 мкм, в центре пластинки – аксиальный гребень.
Впервые описан в центре континентальной части Чили, в заболоченном водоеме около р. Крусес [Cruces] [16].
Распространение: пресноводные местообитания Чили [16], США [33].
Raphidiophrys ovalis (Dürrschmidt in Nicholls et Dürrschmidt, 1985) Siemensma et Roijackers, 1988 [bas.: R. orbicularis ssp. ovalis Dürrschmidt in Nicholls et Dürrschmidt, 1985; syn.: R. intermedia Penard, 1904 sensu Siemensma, 1981] (рис. 2г).
Исследовано несколько пластинчатых чешуек. Чешуйки овальной формы, 6.5–12.0 × 3.0–6.0 мкм. От края чешуйки к центру тянутся 50–60 длинных и коротких ребер. Длинные ребра доходят или почти доходят до центра, короткие достигают в длину 0.5–1.5 мкм.
Впервые описан на о. Чилоэ, в луже на пастбище в окрестностях г. Манао [Manao] [26]. Данный вид отличается от схожих с ним R. intermedia и R. elegans более узким маргинальным утолщением и меньшими размерами чешуек.
Распространение: пресные воды Чили [16, 26], Нидерландов, Швеции [30], США [35], Новой Зеландии, Японии [16].
Отряд Pterocystida Cavalier-Smith in Yabuki et al., 2012.
Семейство Choanocystidae Cavalier-Smith et von der Heyden, 2007; Choanocystis perpusilla (Petersen et Hansen, 1960) Siemensma, 1991 [bas.: Acanthocystis perpusilla Petersen et Hansen, 1960; syn.: C. cordiformis ssp. parvula Dürrschmidt, 1987] (рис. 2д–2ж).
Изучены чешуйки одной клетки и несколько отдельных чешуек. Радиальные чешуйки 2.5–6.0 мкм в длину, ствол слегка изогнут, дистальная часть несет две фурки. Базальная пластинка радиальной чешуйки сердцевидная, с прямыми краями, ~1.0–1.5 мкм в диаметре. Пластинчатые чешуйки овальной формы, бесфактурные, 2.0–2.5 × 1.5–2.0 мкм.
Морфология изученных чешуек в целом соответствует предыдущим описаниям. В работах [5, 20] приведены большие размеры чешуек.
Распространение: пресные воды Чили [16], eвропейской части России [5, 20], Эстонии [23]; соленые континентальные воды европейской части России [3]; морские воды Антарктики [14, 32].
Семейство Pterocystidae Cavalier-Smith et von der Heyden, 2007; Pterocystis foliacea subsp. truncata (Dürrschmidt, 1985) Siemensma, 1991 [bas.: Acanthocystis foliacea Dürrschmidt, 1985] (рис. 2з–2л).
Исследованы чешуйки двух клеток и несколько одиночных чешуек. Радиальные чешуйки 2.0–4.5 мкм в длину, листовидной формы. Ствол чешуйки в виде гребня, вдоль которого по всей длине располагаются широкие латеральные бесфактурные крылья, сужающиеся к дистальной части ствола. Чешуйка образует округлое выпячивание базальной части в дорсальную сторону, формируя небольшой “стебелек”. Пластинчатые чешуйки овальные, 2.5–3.0 × 1.8–2.0 мкм, имеют аксиальный гребень.
Впервые описан в центре континентальной части Чили, в болоте в окрестностях оз. Калафкен [Calafquen] [17]. Данный подвид отличается от Pterocystis foliacea ssp. elongata более расширенной формой латеральных крыльев радиальных чешуек, тянущихся вдоль всей длины ствола.
Распространение: пресные воды Чили [17], Украины [2], Индии [36], Австралии [13], Новой Зеландии, Шри-Ланки, Японии [16]; соленые континентальные воды eвропейской части России [27].
Pterocystis pyriformis (Dürrschmidt, 1987) Siemensma, 1991 [bas.: Acanthocystis pyriformis Dürrschmidt, 1987] (рис. 3а–3в).
Рис. 3.
Морфология чешуек солнечников сем. Pterocystidae, род Pterocystis: (а–в) – P. pyriformis, (г–ж) – P. quadrata. Использовали ТЭМ для а–в, СЭМ – для г–ж.
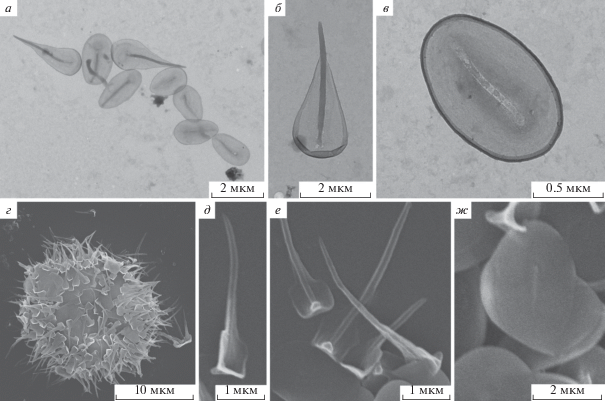
Обнаружено скопление радиальных и пластинчатых чешуек. Радиальные чешуйки грушевидной формы, 2.0–2.5 × 0.8–1.5 мкм. Ствол 1.5 мкм в длину, дистальная часть лишена латеральных крыльев и оканчивается тупой верхушкой. Латеральные крылья бесфактурные, с вогнутыми утолщенными краями, в основании ствола образуют выпячивание. Пластинчатые чешуйки овальной формы, 1.5 × 1.0 мкм, имеют аксиальный гребень.
Впервые описан в центре континентальной части Чили, в придорожной канаве в окрестностях оз. Кабурга [Caburga] [16].
Распространение: пресные воды Чили, Малайзии, Новой Зеландии [16].
Pterocystis quadrata* Cavalier-Smith et Von der Heyden, 2007 (рис. 3г–3ж).
Исследованы чешуйки двух клеток. Радиальные чешуйки 3–5 мкм в длину. Широкий ствол изогнут в дистальную сторону, постепенно сужается к заостренному дистальному концу. От основания вдоль ствола тянутся латеральные крылья 0.8–1.0 мкм в длину, параллельно друг другу, формируя “квадрат”. Крылья в дистальной части резко сужаются, образуя плечи. В основании крылья соединяются, образуя ковшевидное выпячивание базальной части чешуйки. Пластинчатые чешуйки 2.0–4.0 × 1.5–2.0 мкм, несут аксиальный гребень и слабо выраженные радиально отходящие ребра.
Радиальные чешуйки вида схожи по форме с чешуйками P. tropica (Dürrschmidt, 1987) Siemensma et Roijackers, 1988, отличительный признак – наличие у последнего небольшого числа ребер на латеральных крыльях и основании базальной части чешуйки, направленных перпендикулярно стволу чешуйки.
Распространение: пресные воды Великобритании [11].
Raineriophrys fortesca (Nicholls, 1983) Mikrjukov, 1999 [bas.: Acanthocystis fortesca Nicholls, 1983; syn.: A. pantopodeoides Nicholls, 1983; A. cuneiformis Dürrschmidt, 1985; Pterocystis fortesca (Nicholls, 1983) Siemensma et Roijackers, 1988] (рис. 4а–4г).
Рис. 4.
Морфология чешуек солнечников, род Raineriophrys: (а–г) – R. fortesca, (д–ж) – R. pteromorphos, (з–л) – R. erinaceoides. Использовали СЭМ для а–г, з–л, ТЭМ – для д–ж.

Обнаружены чешуйки одной клетки. Радиальные чешуйки 7.5–10.5 мкм в длину. Ствол полый, цилиндрический, 0.25–0.30 мкм в диаметре, оканчивается тупой верхушкой, несущей две–десять фурок. От основания ствола отходят латеральные крылья треугольной формы, соединяются в базальной части и образуют ковшеобразное выпячивание. Латеральные крылья тянутся вдоль ствола на 1.9–2.8 мкм, края слегка загибаются назад. Край базального крыла также слегка загибается назад, ширина основания 1.5–2.2 мкм. Пластинчатые чешуйки овальной или яйцевидной формы, (2.5–3.5) × (1.5–2.5) мкм, бесфактурные, с вогнутой округлой центральной частью и утолщенными краями.
Размеры исследованных чешуек соответствуют описанным ранее, однако на базальной пластинке отсутствуют радиальные чешуйки субпараллельных ребер [6].
Распространение: пресные воды европейской части России [22], Нидерландов, Швеции [29], США [35], Канады, как A. pantopodeoides и A. fortesca [25], Чили, как A. cuneiformis, Германии, Японии [17], Австралии, как A. pantopodeoides [12], Белое море [20].
Raineriophrys pteromorphos (Dürrschmidt, 1987) Mikrjukov, 2001 [bas.: Acanthocystis pteromorphos Dürrschmidt, 1987; syn.: Pterocystis pteromorphos (Dürrschmidt, 1987) Siemensma, 1991] (рис. 4д–4ж).
Обнаружены чешуйки одной клетки. Радиальные чешуйки 2–3 мкм в длину, ствол полый, слегка изогнут, сужается в тупую верхушку. Латеральные крылья тянутся вдоль всего ствола, постепенно сужаясь к дистальной части. От ствола отходят слабо выраженные радиальные ребра. В основании ствола латеральные крылья соединяются, образуя базальные выпячивания чешуйки. Пластинчатые чешуйки овальные, 1–2 мкм в длину, аксиальный гребень слабо заметен.
Впервые описан в центре континентальной части Чили, в болоте Крусес [15].
Распространение: пресные воды Чили [15].
Raineriophrys erinaceoides (Petersen et Hansen, 1960) Mikrjukov, 2001 [bas.: Acanthocystis erinaceoides Petersen et Hansen, 1960; syn.: Echinocystis erinaceoides (Petersen et Hansen, 1960) Mikrjukov, 1997; Pterocystis erinaceoides (Petersen et Hansen, 1960) Siemensma, 1991; Raineria erinaceoides (Petersen et Hansen, 1960) Mikrjukov, 1999] (рис. 4з–4л).
Обнаружены чешуйки одной клетки и несколько одиночных чешуек. Радиальные чешуйки 6–9 мкм в длину, слегка изогнуты в дорсальную сторону, ствол сужается в заостренную верхушку. Основание составляют латеральные крылья, соединенные в базальной части в виде ковша и простирающиеся вдоль ствола на 2/3 его длины. Пластинчатые чешуйки овально-яйцевидной формы, 4–5 × 2.0–2.5 мкм, с аксиальным гребнем.
Распространение: пресные воды Чили [16, 17], Eвропейской части России [20, 22, 38], Крыма [21], Украины [2], Эстонии [23], Монголии [18], Канады [25], Австралии [12], Шри-Ланки [16], соленые континентальные воды европейской части России [3, 27].
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Впервые подробно изучены видовой состав и внешняя морфология центрохелидных солнечников чилийской части Южной Патагонии и Огненной Земли, даны географическая и физико-химическая характеристика водных объектов. Обнаружено 14 видов солнечников класса Centroplasthelida из четырех семейств и шести родов, в том числе два новых для фауны страны вида: Acanthocystis nichollsi и Pterocystis quadrata. Большинство обнаруженных нами солнечников (12 видов) ранее выявлены в центральной части Чили, из них семь видов (Pseudoraphidiophrys discoidea, P. formosa, P. veliformis, Raphidiophrys ovalis, Pterocystis foliacea truncata, P. pyriformis, Raineriophrys pteromorphos) описаны там впервые. Максимальное количество видов принадлежит сем. Pterocystidae (шесть видов), на втором месте сем. Raphidiophryidae (четыре), далее идут сем. Acanthocystidae (три) и Choanocystidae (один). Наиболее часто встречается вид Acanthocystis penardi (в пяти пробах), реже – Choanocystis perpusilla, Pterocystis foliacea truncata и Raphidiophrys ovalis (каждый в трех пробах), только в одной пробе – Acanthocystis nichollsi, Pseudoraphidiophrys discoidea, P. veliformis, Raineriophrys fortesca и R. pteromorphos.
Максимальным видовым богатством (пять видов центрохелид) характеризуется временный водоем на лугу в окрестностях оз. Бланко (табл. 2), что может быть связано с поступлением терригенной органики с талыми водами и хорошим прогревом неглубокого водоема. Также высокое видовое богатство (четыре вида) отмечено в мочажине сфагнового болота в окрестностях г. Пунта-Аренас, что подтверждает выводы других авторов [4, 8, 28] о высоком видовом богатстве солнечников в сфагновых местообитаниях. Всего в сфагновом болоте обнаружено семь видов солнечников (50% общего числа видов) (Acanthocystis penardi, Choanocystis perpusilla, Pseudoraphidiophrys discoidea, P. formosa, P. veliformis, Raphidiophrys ovalis, Pterocystis pyriformis). Во временных водоемах зарегистрировано 11 видов (78.6%) (Acanthocystis pectinata, A. penardi, Choanocystis perpusilla, Pseudoraphidiophrys formosa, Raphidiophrys ovalis Pterocystis foliacea truncata, P. pyriformis, P. quadrata, Raineriophrys fortesca, R. Pteromorphos, R. erinaceoides), в постоянных – два вида (14.3%) (Acanthocystis penardi и Pterocystis foliacea truncatа). В постоянных водотоках найдено пять видов (35.7%) (Acanthocystis nichollsi, A. pectinata, A. penardi, Choanocystis perpusilla, Pterocystis foliacea truncatа). Таким образом, высоким видовым богатством отличались временные водоемы и сфагновые болота, минимальным – постоянные водоемы.
Более половины обнаруженных солнечников (восемь видов, или 57.1% общего числа) найдены только в пресных водах: Acanthocystis penardi, Pseudoraphidiophrys discoidea, P. formosa, P. veliformis, Raphidiophrys ovalis, Pterocystis pyriformis, P. quadrata и Raineriophrys pteromorphos. Остальные (шесть видов, или 42.9%) выявлены также в соленых континентальных и морских водах: Acanthocystis nichollsi, A. pectinata, Choanocystis perpusilla, Pterocystis foliacea truncata, Raineriophrys erinaceoides и R. fortesca [3, 14, 20, 27, 32]. Большинство найденных центрохелид имеют широкое распространение по всему миру, и только три вида крайне редко упоминаются в фаунистических списках, из них Pseudoraphidiophrys discoidea и Raineriophrys pteromorphos ранее встречались только в Чили [15, 17], Pterocystis quadrata – только в Великобритании [11].
Морфология чешуек обнаруженных клеток солнечников в целом соответствует предыдущим описаниям, однако имеются некоторые вариации в размерах и форме, которые выходят за рамки известной внутривидовой изменчивости данных солнечников. Так, чешуйки Acanthocystis nichollsi, A. pectinata, A. penardi и Choanocystis perpusilla оказались несколько меньше описываемых ранее [5, 15, 18, 20, 25]. У радиальных чешуек Raineriophrys fortesca отсутствовали заметные радиальные ребра на базальном крыле. У пластинчатых чешуек Acanthocystis nichollsi были слабо развиты или отсутствовали радиальные ребра. Таким образом, полученные данные дополнили наши представления о внутривидовой изменчивости морфологии чешуек.
Выводы. Фауна центрохелидных солнечников юга Чили в видовом отношении (14 видов) более чем втрое беднее таковой центральной части страны (53) и изучена недостаточно, что подтверждается находками двух новых для региона видов. Характерная особенность фауны – максимальное видовое богатство сем. Pterocystidae за счет родов Pterocystis и Raineriophrys. Фаунистические комплексы отдельных водных объектов невелики – в среднем два вида, максимальное число (5) отмечено в прогреваемом временном водоеме.
Авторы благодарны А.А. Прокину и Д.А. Филиппову за помощь в сборе материала.
Работа выполнена в рамках государственного задания № АААА-А18-118012690098-5 и при поддержке Российского фонда фундаментальных исследований (№ 14-04-00500, 17-04-00565 и 15-29-02518 офи_м).
Список литературы
Галай И.П., Жучкевич В.А., Рылюк Г.Я. Физическая география материков и океанов. Ч. 2: Северная Америка, Южная Америка, Африка, Австралия, Океания, Антарктида, Мировой океан. Минск: Изд-во Университетское, 1988. 366 с.
Гапонова Л.П. Центрохелидни солнцевики (Centrohelea Cavalier-Smith, 1993) Киивського та Чернигивського полисся: Канд. дис. … Ин-т зоологии им. И.И. Шмальгаузена НАН Украины. Киев, 2009. 150 с.
Герасимова Е.А., Плотников А.О. Разнообразие амебоидных протистов в соленых водоемах Оренбургской области // Вестн. Оренбург. гос. ун-та. 2015. № 10(185). С. 130–133.
Златогурский В.В. Солнечники. Протисты: руководство по зоологии. СПб; М.: Товарищество науч. изданий КМК, 2011. С. 205–269.
Леонов М.М., Мыльников А.П. Центрохелидные солнечники из Южной Карелии // Зоол. журн. 2012. Т. 91. № 5. С. 515–525.
Микрюков К.А. Интересные находки солнечников (Protista) в Черном море и Крыму: к вопросу о единстве морской и пресноводной фауны этих организмов // Зоол. журн. 1999. Т. 78. С. 517–527.
Микрюков К.А. Морские и солоноватоводные центрохелидные солнечники (Centroheliozoa, Sarcodina) Кандалакшского залива Белого моря // Зоол. журн. 1994. Т. 73. Вып. 6. С. 5–17.
Микрюков К.А. Центрохелидные солнечники (Centroheliozoa). М.: Товарищество науч. изданий КМК, 2002. 136 с.
Тихоненков Д.В., Мыльников А.П. Новый солнечник Choanocystis antarctica sp. n. (Centrohelida) из литоральной зоны острова Кинг-Джорж (южные Шетландские острова, Антарктика) // Зоол. журн. 2010. Т. 89. № 11. С. 1283–1287.
Burger O. Nuevos estudios sobre protozoos chilenos del aqua dulce // Anales de la Universidad de Chile. 1908. V. 122. P. 137–204.
Cavalier-Smith T., von der Heyden S. Molecular phylogeny, scale evolution and taxonomy of Centrohelid heliozoan // Mol. Phylogenet. Evol. 2007. V. 44. P. 1186–1203.
Croome R.L. Observations of the heliozoan genera Acanthocystis and Raphidocystis from Australia // Archiv für Protistenkunde. 1986. V. 131. P. 189–199.
Croome R.L. Observations of heliozoan genera Acanthocystis, Raphidiophrys, Clathrulina, and Pompholyxophrys from Australian freshwaters // Archiv für Protistenkunde. 1987. V. 133(3/4). P. 237–243.
Croome R.L., van den Hoff J., Burton H.R. Observations of the Heliozoean genera Pinasiophora and Acanthocystis (Heliozoea, Sarcodina, Protozoa) from Ellis Fjord, Antarctica // Polar Biol. 1987. V. 8. P. 23–28.
Dürrschmidt M. An electron microscopical study of freshwater Heliozoa (genus Acanthocystis, Centrohelidia) from Chile, New Zealand, Malaysia and Sri Lanka. II // Archiv für Protistenkunde. 1987. V. 133. P. 21–48.
Dürrschmidt M. An electron microscopical study of freshwater Heliozoa (genus Acanthocystis, Centrohelidia) from Chile, New Zealand, Malaysia and Sri Lanka. III // Archiv für Protistenkunde. 1987. V. 133. P. 49–79.
Dürrschmidt M. Electron microscopic observations on scales of species of the genus Acanthocystis (Centrohelidia, Heliozoa) from Chile. I // Archiv für Protistenkunde. 1985. V. 129. P. 55–87.
Kosolapova N.G., Mylnikov A.P. First record of Centrohelid heliozoans (Centrohelida) in Mongolia // Inland Water Biol. 2015. V. 8. № 3. P. 232–235. doi 10.1134/S1995082915030074
Leonov M.M. Heliozoan fauna of waterbodies and watercourses of the Central Russian Upland forest-steppe // Inland Water Biol. 2009. V. 2. № 1. P. 6–12. doi 10.1134/S1995082909010027
Leonov M.M. Heliozoans (Heliozoa, Sarcodina, Protista) of fresh and marine water of the European part of Russia: species composition, morphology, and description // Inland Water Biol. 2010. V. 3. № 4. P. 344–355. doi 10.1134/S1995082910040073
Mikrjukov K.A. Interesting findings of heliozoan (Protista) in Black Sea and Crimea: data on the similarity of sea and freshwater fauna of these organisms // Rus. J. Zool. 1999. V. 3. № 2. P. 240–249.
Mikrjukov K.A. Observations on Centroheliozoa (Sarcodina, Protozoa) of the Volga basin // Zoosystematica Rossica. 1993. V. 2. № 2. P. 201–209.
Mikrjukov K.A. On centrohelid and rotosphaerid Heliozoa from the environs of the Vôrtsjärv limnological station in Estonia // Proceedings of the Estonian Academy of Sciences, Biology and Ecology. 1993. V. 422. P. 154–160.
Moestrup Ø., Thomsen H.A. Preparations of shadow cast whole mounts // Handbook of Phycological Methods. Cambridge: Cambridge Univ. Press, 1980. P. 385–390.
Nicholls K.H. Little-known and new heliozoeans: the centrohelid genus Acanthocystis, including descriptions of nine new species // Can. J. Zool. 1983. V. 61. № 6. P. 1369–1386.
Nicholls K.H., Dürrschmidt M. Scale structure and taxonomy of some species of Raphidiophrys, Raphidiocystis and Pompholyxophrys (Heliozoa) including descriptions of six new taxa // Can. J. Zool. 1985. V. 63. P. 1944–1961.
Plotnikov A.O., Gerasimova E.A. Heliozoa (Centrohelea, Haptista, Hacrobia) of Saline and Brackish Water Bodies and Watercourses of Russia // Inland Water Biol. 2017. V. 10. № 2. P. 121–129. doi 10.1134/S1995082917020109
Siemensma F.J. Klasse Heliozoa Haeckel, 1866. Nackte Rhizopoda und Heliozoea. Stuttgart; N.Y.: Gustav Fischer Verlag, 1991. P. 171–297.
Siemensma F.J., Roijackers M.M. A study of new and little-known acanthocystid heliozoans, and a proposed devision of the genus Acanthocystis (Actinopoda, Heliozoea) // Archiv für Protistenkunde. 1988. V. 135. P. 197–212.
Siemensma F.J., Roijackers M.M. The genus Raphidiophrys (Actinopoda, Heliozoea): scale morphology and species distinctions // Archiv für Protistenkunde. 1988. V. 136. P. 237–248.
Thomasson K. Araucanian Lakes // Acta Phytogeographica Suecica. 1963. V. 47. P. 1–139.
Tong S., Vørs N., Patterson D.J. Heterotrophic flagellates, centrohelid heliozoan and filose amoebae from marine and freshwater sites in the Antarctic // Polar Biol. 1997. V. 18. P. 91–106.
Wujek D.E. Freshwater heliozoa from Florida // Florida Scientist. 2006. V. 69. P. 177–191.
Wujek D.E. Freshwater heliozoan (Protista, Heliozoa) from India // Proceeding of the Indian Academy of Science. 2003. V. 112. № 2. P. 169–174.
Wujek D.E. Identification, ecology, and distribution of scale bearing amoeba, ciliates, flagellates and heliozoan from the Carolinas // J. North Carolina Acad. Sci. 2005. V. 121. № 1. P. 1–16.
Wujek D.E., Saha L.C. Freshwater scaled Heterotrophic Flagellates and Heliozoa from India including a description of a new species of Choanocystis // J. Bombay Natur. History Soc. 2006. V. 103. № 1. P. 71–81.
Yabuki A., Chao E.E., Ishida K.I., Cavalier-Smith T. Microheliella maris (Microhelida ord. n.), an ultrastructurally highly distinctive new axopodial protist species and genus, and the unity of phylum Heliozoa // Protist. 2012. V. 163. № 3. P. 356–388.
Zlatogursky V.V. Puzzle-like cyst wall in centrohelid heliozoans Raphidiophrys heterophryoidea and Raineriophrys erinaceoides // Acta Protozool. 2013. V. 52. P. 229–236.
Дополнительные материалы отсутствуют.
Инструменты
Биология внутренних вод